Figure 1.
Secreted levels of 53 proteins from n = 8 normal rectal biopsies. Normal rectal biopsies were cultured for 24 h, and the resultant NCM was screened for the expression of 54 inflammatory mediators using a multiplex ELISA. (A) Secreted levels of vascular injury proteins CRP, ICAM-1, SAA and VCAM-1. (B) Secreted levels of angiogenic factors bFGF, Flt-1, P1GF, Tie-2, VEGF-A, VEGF-C and VEGF-D. (C) Inflammatory protein secretions IFN-γ, IL-10, IL-12p70, IL-13, IL-1β, IL-2, IL-4, IL-6, IL-8(HA) and TNF-α. (D) Secretion of Th17 proteins IL-21, IL-22, IL-23, IL-27, IL-31 and CCL20. (E) Chemokine secreted proteins CCL22, CCL3, CCL4, CCL17, CCL11, CCL26, CXCL10, CCL2 and CCL13. (F) Secretion of cytokine panel 1 proteins GM-CSF, IL-12/IL-23p40, IL-15, IL-16, IL-17A, IL-1α, IL-5, IL-7 and TNF-β. (G) Secretion of cytokine 2 panel proteins IL-17A/F, IL-17B, IL-17C, IL-17D, IL-1RA, IL-3, IL-9 and TSLP. All protein secretions are normalised to protein content of the biopsies.
Figure 1.
Secreted levels of 53 proteins from n = 8 normal rectal biopsies. Normal rectal biopsies were cultured for 24 h, and the resultant NCM was screened for the expression of 54 inflammatory mediators using a multiplex ELISA. (A) Secreted levels of vascular injury proteins CRP, ICAM-1, SAA and VCAM-1. (B) Secreted levels of angiogenic factors bFGF, Flt-1, P1GF, Tie-2, VEGF-A, VEGF-C and VEGF-D. (C) Inflammatory protein secretions IFN-γ, IL-10, IL-12p70, IL-13, IL-1β, IL-2, IL-4, IL-6, IL-8(HA) and TNF-α. (D) Secretion of Th17 proteins IL-21, IL-22, IL-23, IL-27, IL-31 and CCL20. (E) Chemokine secreted proteins CCL22, CCL3, CCL4, CCL17, CCL11, CCL26, CXCL10, CCL2 and CCL13. (F) Secretion of cytokine panel 1 proteins GM-CSF, IL-12/IL-23p40, IL-15, IL-16, IL-17A, IL-1α, IL-5, IL-7 and TNF-β. (G) Secretion of cytokine 2 panel proteins IL-17A/F, IL-17B, IL-17C, IL-17D, IL-1RA, IL-3, IL-9 and TSLP. All protein secretions are normalised to protein content of the biopsies.
Figure 2.
Secreted levels of 53 proteins from n = 12 rectal cancer biopsies. Rectal cancer biopsies were cultured for 24 h, and the resultant TCM was screened for the expression of 54 inflammatory mediators using a multiplex ELISA. (A) Secreted levels of vascular injury proteins CRP, ICAM-1, SAA and VCAM-1. (B) Secreted levels of angiogenic factors bFGF, Flt-1, P1GF, Tie-2, VEGF-A, VEGF-C and VEGF-D. (C) Inflammatory secreted proteins IFN-γ, IL-10, IL-12p70, IL-13, IL-1β, IL-2, IL-4, IL-6, IL-8(HA) and TNF-α. (D) Secretion of Th17 proteins IL-21, IL-22, IL-23, IL-27, IL-31 and CCL20. (E) Chemokine secreted proteins CCL22, CCL3, CCL4, CCL17, CCL11, CCL26, CXCL10, CCL2 and CCL13. (F) Secretion of cytokine panel 1 proteins GM-CSF, IL-12/IL-23p40, IL-15, IL-16, IL-17A, IL-1α, IL-5, IL-7 and TNF-β. (G) Secretion of cytokine 2 panel proteins IL-17A/F, IL-17B, IL-17C, IL-17D, IL-1RA, IL-3, IL-9 and TSLP. All protein secretions are normalised to protein content of the biopsies.
Figure 2.
Secreted levels of 53 proteins from n = 12 rectal cancer biopsies. Rectal cancer biopsies were cultured for 24 h, and the resultant TCM was screened for the expression of 54 inflammatory mediators using a multiplex ELISA. (A) Secreted levels of vascular injury proteins CRP, ICAM-1, SAA and VCAM-1. (B) Secreted levels of angiogenic factors bFGF, Flt-1, P1GF, Tie-2, VEGF-A, VEGF-C and VEGF-D. (C) Inflammatory secreted proteins IFN-γ, IL-10, IL-12p70, IL-13, IL-1β, IL-2, IL-4, IL-6, IL-8(HA) and TNF-α. (D) Secretion of Th17 proteins IL-21, IL-22, IL-23, IL-27, IL-31 and CCL20. (E) Chemokine secreted proteins CCL22, CCL3, CCL4, CCL17, CCL11, CCL26, CXCL10, CCL2 and CCL13. (F) Secretion of cytokine panel 1 proteins GM-CSF, IL-12/IL-23p40, IL-15, IL-16, IL-17A, IL-1α, IL-5, IL-7 and TNF-β. (G) Secretion of cytokine 2 panel proteins IL-17A/F, IL-17B, IL-17C, IL-17D, IL-1RA, IL-3, IL-9 and TSLP. All protein secretions are normalised to protein content of the biopsies.
Figure 3.
The protein secretome differs between rectal cancer tissue and normal rectal tissue. NCM and TCM from cultured normal rectal and rectal cancer biopsies were screened for the expression of 54 inflammatory secretions using a multiplex ELISA. Nineteen factors were secreted at significantly higher levels in the TCM compared to the NCM. There were significantly higher levels of (A) Flt-1, (B) P1GF, (C) IFN-γ, (D) IL-6, (E) IL-10, (F) CCL20, (G) CCL26, (H) CCL22, (I) CCL3, (J) CCL4, (K) CCL17, (L) GM-CSF, (M) IL-12/IL-23p40, (N) IL-17A, (O) IL-1α, (P) IL-17A/F, (Q) IL-1RA, (R) TSLP and (S) CXCL10 in the rectal cancer secretome. All data expressed as mean ± SEM. Statistical analysis by Mann Whitney U-test. n = 8 for normal, n = 12 for cancer, n = 7 normal for P1GF, n = 5 normal for CCL17, n = 11 cancer for Flt-1, P1GF, CCL17 and n = 10 cancer for IL-17A/F. **** p < 0.0001, *** p < 0.001, ** p < 0.01, * p < 0.05.
Figure 3.
The protein secretome differs between rectal cancer tissue and normal rectal tissue. NCM and TCM from cultured normal rectal and rectal cancer biopsies were screened for the expression of 54 inflammatory secretions using a multiplex ELISA. Nineteen factors were secreted at significantly higher levels in the TCM compared to the NCM. There were significantly higher levels of (A) Flt-1, (B) P1GF, (C) IFN-γ, (D) IL-6, (E) IL-10, (F) CCL20, (G) CCL26, (H) CCL22, (I) CCL3, (J) CCL4, (K) CCL17, (L) GM-CSF, (M) IL-12/IL-23p40, (N) IL-17A, (O) IL-1α, (P) IL-17A/F, (Q) IL-1RA, (R) TSLP and (S) CXCL10 in the rectal cancer secretome. All data expressed as mean ± SEM. Statistical analysis by Mann Whitney U-test. n = 8 for normal, n = 12 for cancer, n = 7 normal for P1GF, n = 5 normal for CCL17, n = 11 cancer for Flt-1, P1GF, CCL17 and n = 10 cancer for IL-17A/F. **** p < 0.0001, *** p < 0.001, ** p < 0.01, * p < 0.05.
Figure 4.
Radiation alters the secretome of normal rectal tissue. NCM from mock-irradiated and irradiated normal rectal biopsies were screened for the secretion of 54 inflammatory proteins using a multiplex ELISA. Following a single fraction of 1.8 Gy radiation, there were significantly higher levels of (A) IL-15 and (B) CCL22 in the normal rectal secretome. All data expressed as mean ± SEM. Statistical analysis by Wilcoxon signed-rank test. n = 8 for CCL22 and n = 7 for IL-15, * p < 0.05.
Figure 4.
Radiation alters the secretome of normal rectal tissue. NCM from mock-irradiated and irradiated normal rectal biopsies were screened for the secretion of 54 inflammatory proteins using a multiplex ELISA. Following a single fraction of 1.8 Gy radiation, there were significantly higher levels of (A) IL-15 and (B) CCL22 in the normal rectal secretome. All data expressed as mean ± SEM. Statistical analysis by Wilcoxon signed-rank test. n = 8 for CCL22 and n = 7 for IL-15, * p < 0.05.
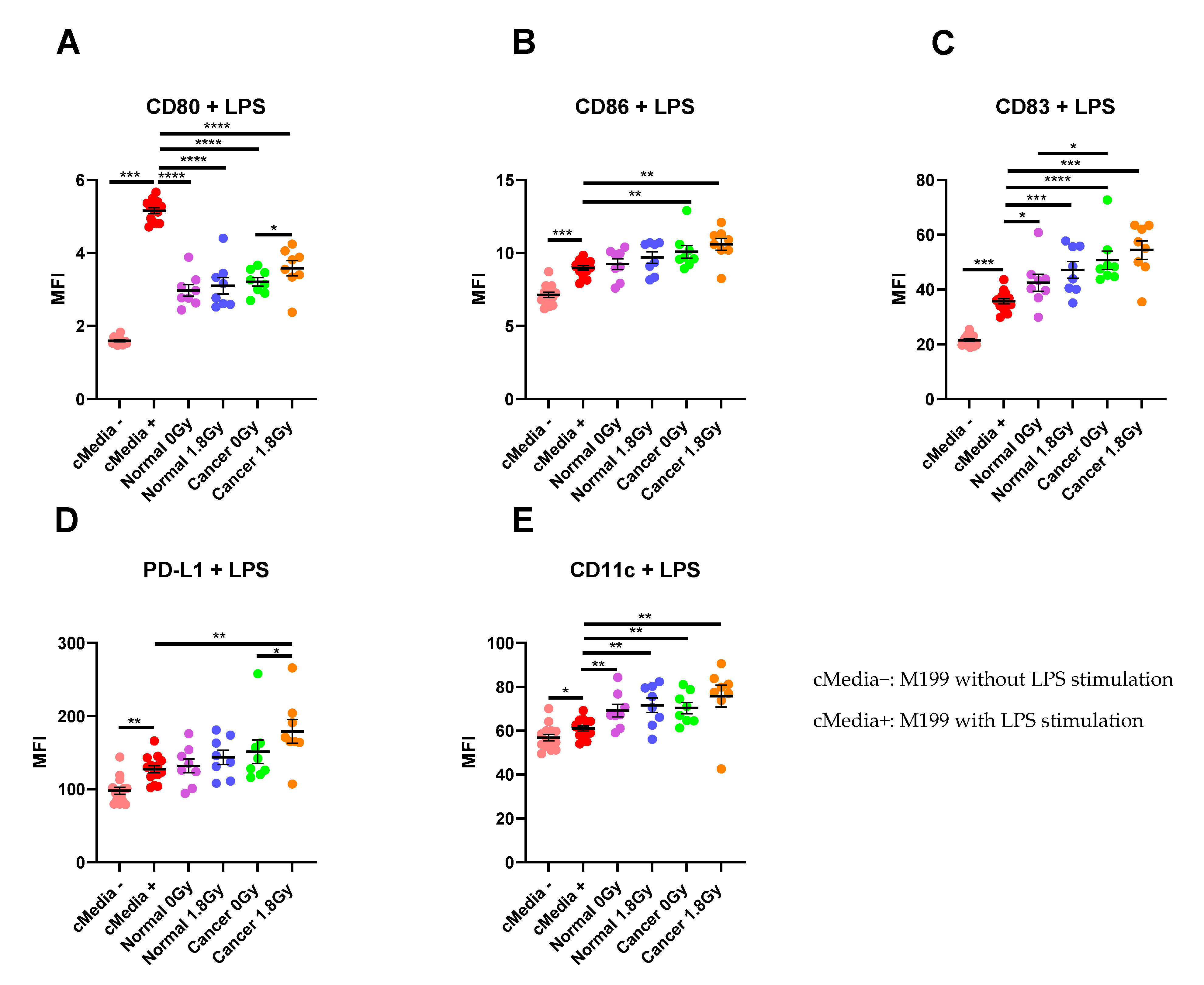
Figure 5.
The effect of NCM and TCM on LPS-induced dendritic cell (DC) maturation. CD11c+ DCs were treated with NCM and TCM from mock-irradiated and irradiated biopsies, and the effect on DC maturation markers was assessed by flow cytometry. (A) NCM and TCM from both mock-irradiated and irradiated biopsies significantly inhibit LPS-induced expression of CD80. TCM from irradiated rectal cancer tissue has a less inhibitory effect than TCM from mock-irradiated rectal cancer tissue. (B) TCM from both irradiated and mock-irradiated rectal cancer tissue caused significant enhancement of LPS-induced expression of CD86. (C) NCM and TCM from both irradiated and mock-irradiated normal rectal and rectal cancer biopsies caused a significant enhancement of LPS-induced expression of CD83. TCM from mock-irradiated rectal cancer biopsies caused significant elevation of CD83 expression levels compared to NCM from mock-irradiated biopsies. (D) PD-L1 expression was significantly enhanced by TCM from irradiated rectal cancer tissue. (E) LPS-induced expression of CD11c was significantly elevated by NCM and TCM from irradiated and mock-irradiated biopsies. All data expressed as mean ± SEM. Statistical analysis by Wilcoxon signed-rank test when comparing the same tissue type, e.g., Cancer 0 Gy vs. Cancer 1.8 Gy and Mann Whitney U-test when comparing different tissue types and comparing to media control. n = 14 for cMedia, n = 8 for normal and cancer, **** p < 0.0001, *** p < 0.001, ** p < 0.01, * p < 0.05.
Figure 5.
The effect of NCM and TCM on LPS-induced dendritic cell (DC) maturation. CD11c+ DCs were treated with NCM and TCM from mock-irradiated and irradiated biopsies, and the effect on DC maturation markers was assessed by flow cytometry. (A) NCM and TCM from both mock-irradiated and irradiated biopsies significantly inhibit LPS-induced expression of CD80. TCM from irradiated rectal cancer tissue has a less inhibitory effect than TCM from mock-irradiated rectal cancer tissue. (B) TCM from both irradiated and mock-irradiated rectal cancer tissue caused significant enhancement of LPS-induced expression of CD86. (C) NCM and TCM from both irradiated and mock-irradiated normal rectal and rectal cancer biopsies caused a significant enhancement of LPS-induced expression of CD83. TCM from mock-irradiated rectal cancer biopsies caused significant elevation of CD83 expression levels compared to NCM from mock-irradiated biopsies. (D) PD-L1 expression was significantly enhanced by TCM from irradiated rectal cancer tissue. (E) LPS-induced expression of CD11c was significantly elevated by NCM and TCM from irradiated and mock-irradiated biopsies. All data expressed as mean ± SEM. Statistical analysis by Wilcoxon signed-rank test when comparing the same tissue type, e.g., Cancer 0 Gy vs. Cancer 1.8 Gy and Mann Whitney U-test when comparing different tissue types and comparing to media control. n = 14 for cMedia, n = 8 for normal and cancer, **** p < 0.0001, *** p < 0.001, ** p < 0.01, * p < 0.05.

Table 1.
Correlation between DC maturation markers and secreted factors in the microenvironment of irradiated normal rectal tissue, i.e., Normal 1.8 Gy.
Table 1.
Correlation between DC maturation markers and secreted factors in the microenvironment of irradiated normal rectal tissue, i.e., Normal 1.8 Gy.
| DC Marker | Secreted Factor | r Value | p Value | n |
|---|
| CD80 | IL-10 | −0.8571 | 0.01 | 8 |
Table 2.
Correlation between DC maturation and phenotypic markers and secreted factors in the microenvironment of rectal cancer tissue.
Table 2.
Correlation between DC maturation and phenotypic markers and secreted factors in the microenvironment of rectal cancer tissue.
| CANCER 0 GY | CANCER 1.8 GY |
|---|
| DC Marker | Secreted Factor | r Value | p Value | n | DC Marker | Secreted Factor | r Value | p Value | n |
|---|
| CD80 | Flt-1 | 0.7857 | 0.04 | 7 | CD80 | ICAM-1 | 0.8214 | 0.03 | 7 |
| CD80 | IL-27 | −0.8214 | 0.03 | 7 | CD11c | IL-1RA | 0.7619 | 0.03 | 8 |
| CD83 | SAA | 0.7857 | 0.04 | 7 | | | | | |
Table 3.
Correlation between DC maturation and phenotypic markers and secreted factors from DCs treated with NCM.
Table 3.
Correlation between DC maturation and phenotypic markers and secreted factors from DCs treated with NCM.
| NORMAL 1.8 GY |
|---|
| DC Marker | Secreted Factor | r Value | p Value | n |
|---|
| CD83 | ICAM-1 | −0.7857 | 0.02 | 8 |
| CD11c | ICAM-1 | −0.7857 | 0.02 | 8 |
Table 4.
Correlation between DC maturation and phenotypic markers and secreted factors from DCs treated with TCM.
Table 4.
Correlation between DC maturation and phenotypic markers and secreted factors from DCs treated with TCM.
| CANCER 0 GY | CANCER 1.8 GY |
|---|
| DC Marker | Secreted Factor | r Value | p Value | n | DC Marker | Secreted Factor | r Value | p Value | n |
|---|
| PD-L1 | TNF-α | 0.8333 | 0.01 | 8 | CD86 | VEGF-C | −0.9276 | 0.01 | 6 |
| | | | | | CD11c | VEGF-C | −0.8857 | 0.03 | 6 |
Table 5.
Correlation between body composition parameters and factors secreted from mock-irradiated rectal cancer tissue, i.e., Cancer 0 Gy.
Table 5.
Correlation between body composition parameters and factors secreted from mock-irradiated rectal cancer tissue, i.e., Cancer 0 Gy.
| Body Composition Parameter | Secreted Factor | r Value | p Value | n |
|---|
| Skeletal muscle | Flt-1 | −0.6273 | 0.04 | 11 |
| Skeletal muscle | IL-12/IL-23p40 | 0.6573 | 0.02 | 12 |
| Skeletal muscle | IL-1α | 0.5874 | 0.04 | 12 |
| Skeletal muscle | VEGF-A | 0.6224 | 0.03 | 12 |
| Visceral fat area | CCL20 | 0.6783 | 0.01 | 12 |
Table 6.
Correlation between body composition parameters and factors secreted from irradiated rectal cancer tissue, i.e., Cancer 1.8 Gy.
Table 6.
Correlation between body composition parameters and factors secreted from irradiated rectal cancer tissue, i.e., Cancer 1.8 Gy.
| Body Composition Parameter | Secreted Factor | r Value | p Value | n |
|---|
| Skeletal muscle | Flt-1 | −0.7182 | 0.01 | 11 |
| Skeletal muscle | VEGF-D | −1 | 0.01 | 5 |
| Intermuscular fat | CCL20 | 0.7133 | 0.01 | 12 |
| Intermuscular fat | VEGF-A | 0.6084 | 0.03 | 12 |
| Intermuscular fat | IL-1RA | 0.6084 | 0.03 | 12 |
| Visceral fat area | CCL20 | 0.6643 | 0.02 | 12 |
| Visceral fat area | IL-1RA | 0.6503 | 0.02 | 12 |


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}