
Agonistic CD40 Antibodies in Cancer Treatment


Abstract
:Simple Summary
Abstract
1. Introduction
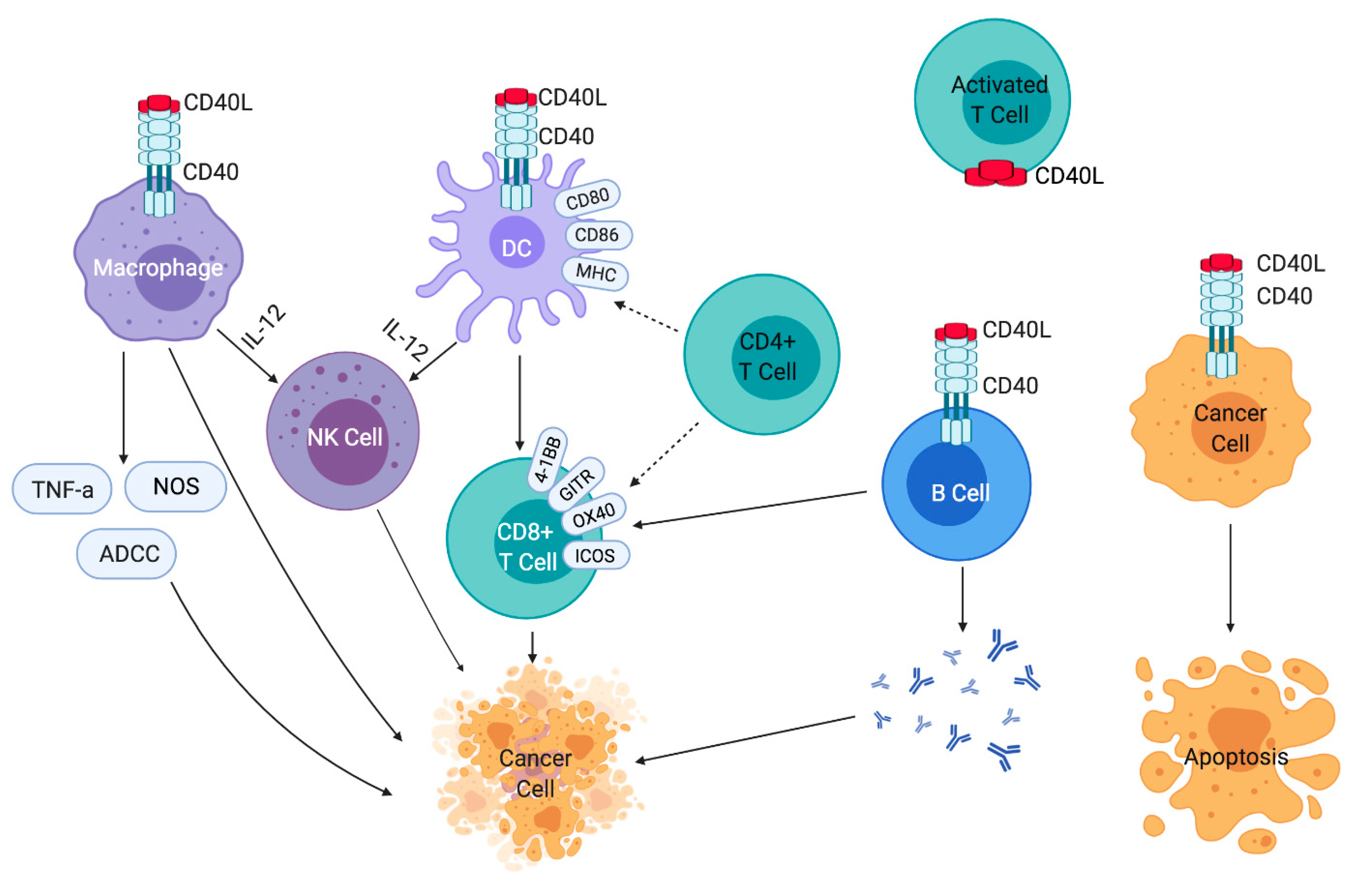
2. Expression of CD40 and CD40L and Downstream Signaling
3. Effects of CD40 Activation on Immune Cells
4. Expression of CD40 in Various Tumor Types
5. Pre-Clinical Studies Supporting Development of Agonistic CD40 Antibodies for Cancer
5.1. CD40 Agonists in Combination with Chemotherapy and the Sequencing of Treatments
5.2. Synergism with Other Macrophage Modulating Drugs
5.3. Combinations of CD40 Agonists and PD-1/PD-L1 Inhibitors, with or without Chemotherapy, and Effects on T Cell Infiltration
5.4. Combinations of CD40 Agonists and Other Immune Modulatory Approaches
5.5. Combinations of CD40 Agonists and Inhibitors of Angiogenesis
6. Development of Human Agonistic CD40 Antibodies for Clinical Use
6.1. Selicrelumab
6.2. CDX-1140
6.3. APX005M
6.4. ADC-1013
6.5. SEA-CD40
6.6. ChiLob7/4
7. Summary
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, P.; Hu-Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, adaptive, and acquired resistance to cancer immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, J.S.; Teng, M.W.L.; Smyth, M.J. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat. Rev. Clin. Oncol. 2019, 16, 151–167. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Paz-Ares, L.; Bernabe Caro, R.; Zurawski, B.; Kim, S.W.; Carcereny Costa, E.; Park, K.; Alexandru, A.; Lupinacci, L.; de la Mora Jimenez, E.; et al. Nivolumab plus ipilimumab in advanced non-small-cell lung cancer. N. Engl. J. Med. 2019, 381, 2020–2031. [Google Scholar] [CrossRef]
- Motzer, R.J.; Tannir, N.M.; McDermott, D.F.; Frontera, O.A.; Melichar, B.; Choueiri, T.K.; Plimack, E.R.; Barthelemy, P.; Porta, C.; George, S.; et al. Nivolumab plus ipilimumab versus sunitinib in advanced renal-cell carcinoma. N. Engl. J. Med. 2018, 378, 1277–1290. [Google Scholar] [CrossRef]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall survival with combined nivolumab and ipilimumab in advanced melanoma. N. Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Hodi, F.S.; Chesney, J.; Pavlick, A.C.; Robert, C.; Grossmann, K.F.; McDermott, D.F.; Linette, G.P.; Meyer, N.; Giguere, J.R.K.; Agarwala, S.S.; et al. Combined nivolumab and ipilimumab versus ipilimumab alone in patients with advanced melanoma: 2-year overall survival outcomes in a multicentre, randomised, controlled, phase 2 trial. Lancet Oncol. 2016, 17, 1558–1568. [Google Scholar] [CrossRef] [Green Version]
- Hammers, H.J.; Plimack, E.R.; Infante, J.R.; Rini, B.I.; McDermott, D.F.; Lewis, L.D.; Voss, M.H.; Sharma, P.; Pal, S.K.; Razak, A.R.A.; et al. Safety and efficacy of nivolumab in combination with ipilimumab in metastatic renal cell carcinoma: The checkmate 016 study. J. Clin. Oncol. 2017, 35, 3851–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cella, D.; Grunwald, V.; Escudier, B.; Hammers, H.J.; George, S.; Nathan, P.; Grimm, M.O.; Rini, B.I.; Doan, J.; Ivanescu, C.; et al. Patient-reported outcomes of patients with advanced renal cell carcinoma treated with nivolumab plus ipilimumab versus sunitinib (checkmate 214): A randomised, phase 3 trial. Lancet Oncol. 2019, 20, 297–310. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Rizvi, N.A.; Goldman, J.W.; Gettinger, S.N.; Borghaei, H.; Brahmer, J.R.; Ready, N.E.; Gerber, D.E.; Chow, L.Q.; Juergens, R.A.; et al. Nivolumab plus ipilimumab as first-line treatment for advanced non-small-cell lung cancer (checkmate 012): Results of an open-label, phase 1, multicohort study. Lancet Oncol. 2017, 18, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Wang, Y.; Chlewicki, L.K.; Zhang, Y.; Guo, J.; Liang, W.; Wang, J.; Wang, X.; Fu, Y.X. Facilitating T cell infiltration in tumor microenvironment overcomes resistance to pd-l1 blockade. Cancer Cell 2016, 30, 500. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Powis de Tenbossche, C.G.; Cane, S.; Colau, D.; van Baren, N.; Lurquin, C.; Schmitt-Verhulst, A.M.; Liljestrom, P.; Uyttenhove, C.; Van den Eynde, B.J. Resistance to cancer immunotherapy mediated by apoptosis of tumor-infiltrating lymphocytes. Nat. Commun. 2017, 8, 1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgueta, R.; Benson, M.J.; de Vries, V.C.; Wasiuk, A.; Guo, Y.; Noelle, R.J. Molecular mechanism and function of CD40/CD40L engagement in the immune system. Immunol. Rev. 2009, 229, 152–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewal, I.S.; Flavell, R.A. The role of CD40 ligand in costimulation and T-cell activation. Immunol. Rev. 1996, 153, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, C.; Rocha, B.; Tanchot, C. A role for CD40 expression on CD8+ T cells in the generation of CD8+ T cell memory. Science 2002, 297, 2060–2063. [Google Scholar] [CrossRef] [PubMed]
- Chand Dakal, T.; Dhabhai, B.; Agarwal, D.; Gupta, R.; Nagda, G.; Meena, A.R.; Dhakar, R.; Menon, A.; Mathur, R.; Mona; et al. Mechanistic basis of co-stimulatory CD40-CD40L ligation mediated regulation of immune responses in cancer and autoimmune disorders. Immunobiology 2020, 225, 151899. [Google Scholar] [CrossRef] [PubMed]
- Michel, N.A.; Zirlik, A.; Wolf, D. CD40L and its receptors in atherothrombosis-an update. Front. Cardiovasc. Med. 2017, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.; Hohmann, J.D.; Wiedemann, A.; Bledzka, K.; Blankenbach, H.; Marchini, T.; Gutte, K.; Zeschky, K.; Bassler, N.; Hoppe, N.; et al. Binding of CD40L to mac-1’s i-domain involves the eqlkksktl motif and mediates leukocyte recruitment and atherosclerosis--but does not affect immunity and thrombosis in mice. Circ. Res. 2011, 109, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Andre, P.; Prasad, K.S.; Denis, C.V.; He, M.; Papalia, J.M.; Hynes, R.O.; Phillips, D.R.; Wagner, D.D. CD40L stabilizes arterial thrombi by a beta3 integrin--dependent mechanism. Nat. Med. 2002, 8, 247–252. [Google Scholar] [CrossRef]
- Banchereau, J.; Bazan, F.; Blanchard, D.; Briere, F.; Galizzi, J.P.; van Kooten, C.; Liu, Y.J.; Rousset, F.; Saeland, S. The CD40 antigen and its ligand. Annu. Rev. Immunol. 1994, 12, 881–922. [Google Scholar] [CrossRef] [PubMed]
- Ihle, J.N.; Kerr, I.M. Jaks and stats in signaling by the cytokine receptor superfamily. Trends Genet. 1995, 11, 69–74. [Google Scholar] [CrossRef]
- Saemann, M.D.; Diakos, C.; Kelemen, P.; Kriehuber, E.; Zeyda, M.; Bohmig, G.A.; Horl, W.H.; Baumruker, T.; Zlabinger, G.J. Prevention of CD40-triggered dendritic cell maturation and induction of T-cell hyporeactivity by targeting of janus kinase 3. Am. J. Transplant. 2003, 3, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Revy, P.; Hivroz, C.; Andreu, G.; Graber, P.; Martinache, C.; Fischer, A.; Durandy, A. Activation of the janus kinase 3-stat5a pathway after CD40 triggering of human monocytes but not of resting B cells. J. Immunol. 1999, 163, 787–793. [Google Scholar] [PubMed]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-kappaB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Hostager, B.S.; Bishop, G.A. CD40-mediated activation of the NF-kappaB2 pathway. Front. Immunol. 2013, 4, 376. [Google Scholar] [CrossRef] [Green Version]
- Rescigno, M.; Martino, M.; Sutherland, C.L.; Gold, M.R.; Ricciardi-Castagnoli, P. Dendritic cell survival and maturation are regulated by different signaling pathways. J. Exp. Med. 1998, 188, 2175–2180. [Google Scholar] [CrossRef]
- Dong, C.; Davis, R.J.; Flavell, R.A. Map kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, L.; Zhu, D.; Bae, D.; Nel, A.; Saxon, A. CD40-mediated p38 mitogen-activated protein kinase activation is required for immunoglobulin class switch recombination to IgE. J. Allergy Clin. Immunol. 2002, 110, 421–428. [Google Scholar] [CrossRef]
- Yu, Q.; Kovacs, C.; Yue, F.Y.; Ostrowski, M.A. The role of the p38 mitogen-activated protein kinase, extracellular signal-regulated kinase, and phosphoinositide-3-OH kinase signal transduction pathways in CD40 ligand-induced dendritic cell activation and expansion of virus-specific CD8+ T cell memory responses. J. Immunol. 2004, 172, 6047–6056. [Google Scholar]
- Mathur, R.K.; Awasthi, A.; Wadhone, P.; Ramanamurthy, B.; Saha, B. Reciprocal CD40 signals through p38MAPK and erk-1/2 induce counteracting immune responses. Nat. Med. 2004, 10, 540–544. [Google Scholar] [CrossRef]
- Foey, A.D.; Feldmann, M.; Brennan, F.M. CD40 ligation induces macrophage IL-10 and TNF-alpha production: Differential use of the PI3K and p42/44 mapk-pathways. Cytokine 2001, 16, 131–142. [Google Scholar] [CrossRef]
- Grewal, I.S.; Flavell, R.A. CD40 and CD154 in cell-mediated immunity. Annu. Rev. Immunol. 1998, 16, 111–135. [Google Scholar] [CrossRef]
- O’Sullivan, B.; Thomas, R. Recent advances on the role of CD40 and dendritic cells in immunity and tolerance. Curr. Opin. Hematol. 2003, 10, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifaz, L.; Bonnyay, D.; Mahnke, K.; Rivera, M.; Nussenzweig, M.C.; Steinman, R.M. Efficient targeting of protein antigen to the dendritic cell receptor DEC-205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8+ T cell tolerance. J. Exp. Med. 2002, 196, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Vonderheide, R.H. The immune revolution: A case for priming, not checkpoint. Cancer Cell 2018, 33, 563–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.G.; Rakhmilevich, A.L.; Burdelya, L.; Neal, Z.; Imboden, M.; Sondel, P.M.; Yu, H. Anti-CD40 antibody induces antitumor and antimetastatic effects: The role of NK cells. J. Immunol. 2001, 166, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Rakhmilevich, A.L.; Alderson, K.L.; Sondel, P.M. T-cell-independent antitumor effects of CD40 ligation. Int. Rev. Immunol. 2012, 31, 267–278. [Google Scholar] [CrossRef]
- Schoenberger, S.P.; Toes, R.E.; van der Voort, E.I.; Offringa, R.; Melief, C.J. T-cell help for cytotoxic t lymphocytes is mediated by CD40-CD40L interactions. Nature 1998, 393, 480–483. [Google Scholar] [CrossRef]
- Sotomayor, E.M.; Borrello, I.; Tubb, E.; Rattis, F.M.; Bien, H.; Lu, Z.; Fein, S.; Schoenberger, S.; Levitsky, H.I. Conversion of tumor-specific CD4+ T-cell tolerance to T-cell priming through in vivo ligation of CD40. Nat. Med. 1999, 5, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Jackaman, C.; Cornwall, S.; Graham, P.T.; Nelson, D.J. CD40-activated B cells contribute to mesothelioma tumor regression. Immunol. Cell Biol. 2011, 89, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, A.G.; Davies, C.; Knox, P.G.; Gallagher, N.J.; Afford, S.C.; Adams, D.H.; Young, L.S. CD40 induces apoptosis in carcinoma cells through activation of cytotoxic ligands of the tumor necrosis factor superfamily. Mol. Cell. Biol. 2000, 20, 5503–5515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, S.; Engelmann, H. A novel function of CD40: Induction of cell death in transformed cells. J. Exp. Med. 1996, 183, 159–167. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Morishima, N.; Mizoguchi, I.; Shimizu, M.; Nagai, H.; Oniki, S.; Oka, M.; Nishigori, C.; Mizuguchi, J. Antiproliferative activity of IL-27 on melanoma. J. Immunol. 2008, 180, 6527–6535. [Google Scholar] [CrossRef] [Green Version]
- Oniki, S.; Nagai, H.; Horikawa, T.; Furukawa, J.; Belladonna, M.L.; Yoshimoto, T.; Hara, I.; Nishigori, C. Interleukin-23 and interleukin-27 exert quite different antitumor and vaccine effects on poorly immunogenic melanoma. Cancer Res. 2006, 66, 6395–6404. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Zhang, Y.; Li, B.; Zhang, J.; Dong, Z.; Hu, X.; Wan, Y. Trim21 mediates ubiquitination of snail and modulates epithelial to mesenchymal transition in breast cancer cells. Int. J. Biol. Macromol. 2019, 124, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, S.; Stiles, J.K. The emerging role of CXCL10 in cancer (review). Oncol. Lett. 2011, 2, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Luster, A.D.; Ravetch, J.V. Biochemical characterization of a gamma interferon-inducible cytokine (ip-10). J. Exp. Med. 1987, 166, 1084–1097. [Google Scholar] [CrossRef]
- Dyer, K.D.; Percopo, C.M.; Fischer, E.R.; Gabryszewski, S.J.; Rosenberg, H.F. Pneumoviruses infect eosinophils and elicit MyD88-dependent release of chemoattractant cytokines and interleukin-6. Blood 2009, 114, 2649–2656. [Google Scholar] [CrossRef]
- Lund, R.; Ahlfors, H.; Kainonen, E.; Lahesmaa, A.M.; Dixon, C.; Lahesmaa, R. Identification of genes involved in the initiation of human Th1 or Th2 cell commitment. Eur. J. Immunol. 2005, 35, 3307–3319. [Google Scholar] [CrossRef] [PubMed]
- Turman, M.A.; Yabe, T.; McSherry, C.; Bach, F.H.; Houchins, J.P. Characterization of a novel gene (NKG7) on human chromosome 19 that is expressed in natural killer cells and T cells. Hum. Immunol. 1993, 36, 34–40. [Google Scholar] [CrossRef]
- Fairfax, B.P.; Taylor, C.A.; Watson, R.A.; Nassiri, I.; Danielli, S.; Fang, H.; Mahe, E.A.; Cooper, R.; Woodcock, V.; Traill, Z.; et al. Peripheral CD8(+) T cell characteristics associated with durable responses to immune checkpoint blockade in patients with metastatic melanoma. Nat. Med. 2020, 26, 193–199. [Google Scholar] [CrossRef]
- Nowak, A.K.; Robinson, B.W.; Lake, R.A. Synergy between chemotherapy and immunotherapy in the treatment of established murine solid tumors. Cancer Res. 2003, 63, 4490–4496. [Google Scholar]
- Byrne, K.T.; Vonderheide, R.H. CD40 stimulation obviates innate sensors and drives T cell immunity in cancer. Cell Rep. 2016, 15, 2719–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanderley, C.W.; Colon, D.F.; Luiz, J.P.M.; Oliveira, F.F.; Viacava, P.R.; Leite, C.A.; Pereira, J.A.; Silva, C.M.; Silva, C.R.; Silva, R.L.; et al. Paclitaxel reduces tumor growth by reprogramming tumor-associated macrophages to an M1 profile in a TLR4-dependent manner. Cancer Res. 2018, 78, 5891–5900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannesdottir, L.; Tymoszuk, P.; Parajuli, N.; Wasmer, M.H.; Philipp, S.; Daschil, N.; Datta, S.; Koller, J.B.; Tripp, C.H.; Stoitzner, P.; et al. Lapatinib and doxorubicin enhance the Stat1-dependent antitumor immune response. Eur. J. Immunol. 2013, 43, 2718–2729. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Buque, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic cell death in cancer and infectious disease. Nat. Rev. Immunol. 2017, 17, 97–111. [Google Scholar] [CrossRef]
- Tomihara, K.; Fuse, H.; Heshiki, W.; Takei, R.; Zhang, B.; Arai, N.; Nakamori, K.; Noguchi, M. Gemcitabine chemotherapy induces phenotypic alterations of tumor cells that facilitate antitumor T cell responses in a mouse model of oral cancer. Oral. Oncol. 2014, 50, 457–467. [Google Scholar] [CrossRef]
- Dammeijer, F.; De Gooijer, C.J.; van Gulijk, M.; Lukkes, M.; Klaase, L.; Lievense, L.A.; Waasdorp, C.; Jebbink, M.; Bootsma, G.P.; Stigt, J.A.; et al. Immune monitoring in mesothelioma patients identifies novel immune-modulatory functions of gemcitabine associating with clinical response. EBioMedicine 2020, 103160. [Google Scholar]
- Cullis, J.; Siolas, D.; Avanzi, A.; Barui, S.; Maitra, A.; Bar-Sagi, D. Macropinocytosis of Nab-paclitaxel drives macrophage activation in pancreatic cancer. Cancer Immunol. Res. 2017, 5, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diggs, L.P.; Ruf, B.; Ma, C.; Heinrich, B.; Cui, L.; Zhang, Q.; McVey, J.C.; Wabitsch, S.; Heinrich, S.; Rosato, U.; et al. CD40-mediated immune cell activation enhances response to anti-PD1 in murine intrahepatic cholangiocarcinoma. J. Hepatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Byrne, K.T.; Leisenring, N.H.; Bajor, D.L.; Vonderheide, R.H. CSF-1R-dependent lethal hepatotoxicity when agonistic CD40 antibody is given before but not after chemotherapy. J. Immunol. 2016, 197, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komohara, Y.; Fujiwara, Y.; Ohnishi, K.; Takeya, M. Tumor-associated macrophages: Potential therapeutic targets for anti-cancer therapy. Adv. Drug Deliv. Rev. 2016, 99, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.J.; Munoz-Rojas, A.R.; Meeth, K.M.; Kellman, L.N.; Amezquita, R.A.; Thakral, D.; Du, V.Y.; Wang, J.X.; Damsky, W.; Kuhlmann, A.L.; et al. Myeloid-targeted immunotherapies act in synergy to induce inflammation and antitumor immunity. J. Exp. Med. 2018, 215, 877–893. [Google Scholar] [CrossRef] [Green Version]
- Hoves, S.; Ooi, C.H.; Wolter, C.; Sade, H.; Bissinger, S.; Schmittnaegel, M.; Ast, O.; Giusti, A.M.; Wartha, K.; Runza, V.; et al. Rapid activation of tumor-associated macrophages boosts preexisting tumor immunity. J. Exp. Med. 2018, 215, 859–876. [Google Scholar] [CrossRef]
- Latchman, Y.E.; Liang, S.C.; Wu, Y.; Chernova, T.; Sobel, R.A.; Klemm, M.; Kuchroo, V.K.; Freeman, G.J.; Sharpe, A.H. PD-L1-deficient mice show that PD-L1 on T cells, antigen-presenting cells, and host tissues negatively regulates T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 10691–10696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taube, J.M.; Anders, R.A.; Young, G.D.; Xu, H.; Sharma, R.; McMiller, T.L.; Chen, S.; Klein, A.P.; Pardoll, D.M.; Topalian, S.L.; et al. Colocalization of inflammatory response with B7-h1 expression in human melanocytic lesions supports an adaptive resistance mechanism of immune escape. Sci. Transl. Med. 2012, 4, 127ra137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thumar, J.R.; Kluger, H.M. Ipilimumab: A promising immunotherapy for melanoma. Oncology 2010, 24, 1280–1288. [Google Scholar]
- Kluger, H.M.; Zito, C.R.; Turcu, G.; Baine, M.K.; Zhang, H.; Adeniran, A.; Sznol, M.; Rimm, D.L.; Kluger, Y.; Chen, L.; et al. PD-L1 studies across tumor types, its differential expression and predictive value in patients treated with immune checkpoint inhibitors. Clin. Cancer Res. 2017, 23, 4270–4279. [Google Scholar] [CrossRef] [Green Version]
- Zippelius, A.; Schreiner, J.; Herzig, P.; Muller, P. Induced PD-L1 expression mediates acquired resistance to agonistic anti-CD40 treatment. Cancer Immunol. Res. 2015, 3, 236–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luheshi, N.M.; Coates-Ulrichsen, J.; Harper, J.; Mullins, S.; Sulikowski, M.G.; Martin, P.; Brown, L.; Lewis, A.; Davies, G.; Morrow, M.; et al. Transformation of the tumour microenvironment by a CD40 agonist antibody correlates with improved responses to PD-L1 blockade in a mouse orthotopic pancreatic tumour model. Oncotarget 2016, 7, 18508–18520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngiow, S.F.; Young, A.; Blake, S.J.; Hill, G.R.; Yagita, H.; Teng, M.W.; Korman, A.J.; Smyth, M.J. Agonistic CD40 mab-driven IL12 reverses resistance to anti-pd1 in a T-cell-rich tumor. Cancer Res. 2016, 76, 6266–6277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rech, A.J.; Dada, H.; Kotzin, J.J.; Henao-Mejia, J.; Minn, A.J.; Twyman-Saint Victor, C.; Vonderheide, R.H. Radiotherapy and CD40 activation separately augment immunity to checkpoint blockade in cancer. Cancer Res. 2018, 78, 4282–4291. [Google Scholar] [CrossRef] [Green Version]
- Winograd, R.; Byrne, K.T.; Evans, R.A.; Odorizzi, P.M.; Meyer, A.R.; Bajor, D.L.; Clendenin, C.; Stanger, B.Z.; Furth, E.E.; Wherry, E.J.; et al. Induction of T-cell immunity overcomes complete resistance to PD-1 and CTLA-4 blockade and improves survival in pancreatic carcinoma. Cancer Immunol. Res. 2015, 3, 399–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahonen, C.L.; Doxsee, C.L.; McGurran, S.M.; Riter, T.R.; Wade, W.F.; Barth, R.J.; Vasilakos, J.P.; Noelle, R.J.; Kedl, R.M. Combined tlr and CD40 triggering induces potent CD8+ T cell expansion with variable dependence on type I IFN. J. Exp. Med. 2004, 199, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.W.; Cowled, C.J.; Farzaneh, F.; Noble, A. Combined triggering of dendritic cell receptors results in synergistic activation and potent cytotoxic immunity. J. Immunol. 2008, 181, 3422–3431. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Chen, H.L.; Girgis, K.R.; Cunningham, H.T.; Meny, G.M.; Nadaf, S.; Kavanaugh, D.; Carbone, D.P. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat. Med. 1996, 2, 1096–1103. [Google Scholar] [CrossRef]
- Gabrilovich, D.; Ishida, T.; Oyama, T.; Ran, S.; Kravtsov, V.; Nadaf, S.; Carbone, D.P. Vascular endothelial growth factor inhibits the development of dendritic cells and dramatically affects the differentiation of multiple hematopoietic lineages in vivo. Blood 1998, 92, 4150–4166. [Google Scholar] [CrossRef]
- Oyama, T.; Ran, S.; Ishida, T.; Nadaf, S.; Kerr, L.; Carbone, D.P.; Gabrilovich, D.I. Vascular endothelial growth factor affects dendritic cell maturation through the inhibition of nuclear factor-kappa B activation in hemopoietic progenitor cells. J. Immunol. 1998, 160, 1224–1232. [Google Scholar]
- Sfiligoi, C.; de Luca, A.; Cascone, I.; Sorbello, V.; Fuso, L.; Ponzone, R.; Biglia, N.; Audero, E.; Arisio, R.; Bussolino, F.; et al. Angiopoietin-2 expression in breast cancer correlates with lymph node invasion and short survival. Int. J. Cancer 2003, 103, 466–474. [Google Scholar] [CrossRef]
- Coffelt, S.B.; Chen, Y.Y.; Muthana, M.; Welford, A.F.; Tal, A.O.; Scholz, A.; Plate, K.H.; Reiss, Y.; Murdoch, C.; De Palma, M.; et al. Angiopoietin 2 stimulates TIE2-expressing monocytes to suppress T cell activation and to promote regulatory T cell expansion. J. Immunol. 2011, 186, 4183–4190. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, S.; Raundhal, M.; Patidar, A.; Saha, B. Anti-VEGF antibody enhances the antitumor effect of CD40. Int. J. Cancer 2014, 135, 1983–1988. [Google Scholar] [CrossRef]
- Kashyap, A.S.; Schmittnaegel, M.; Rigamonti, N.; Pais-Ferreira, D.; Mueller, P.; Buchi, M.; Ooi, C.H.; Kreuzaler, M.; Hirschmann, P.; Guichard, A.; et al. Optimized antiangiogenic reprogramming of the tumor microenvironment potentiates CD40 immunotherapy. Proc. Natl. Acad. Sci. USA 2020, 117, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, A.L.; Chan, H.T.; Roghanian, A.; French, R.R.; Mockridge, C.I.; Tutt, A.L.; Dixon, S.V.; Ajona, D.; Verbeek, J.S.; Al-Shamkhani, A.; et al. Interaction with fcgammariib is critical for the agonistic activity of anti-CD40 monoclonal antibody. J. Immunol. 2011, 187, 1754–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Ravetch, J.V. Inhibitory Fc gamma receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies. Science 2011, 333, 1030–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, A.L.; Chan, H.T.; French, R.R.; Willoughby, J.; Mockridge, C.I.; Roghanian, A.; Penfold, C.A.; Booth, S.G.; Dodhy, A.; Polak, M.E.; et al. Conformation of the human immunoglobulin G2 hinge imparts superagonistic properties to immunostimulatory anticancer antibodies. Cancer Cell. 2015, 27, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Barr, T.A.; Heath, A.W. Functional activity of CD40 antibodies correlates to the position of binding relative to CD154. Immunology. 2001, 102, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Bjorck, P.; Braesch-Andersen, S.; Paulie, S. Antibodies to distinct epitopes on the CD40 molecule co-operate in stimulation and can be used for the detection of soluble CD40. Immunology 1994, 83, 430–437. [Google Scholar]
- Ellmark, P.; Ottosson, C.; Borrebaeck, C.A.; Malmborg Hager, A.C.; Furebring, C. Modulation of the CD40-CD40 ligand interaction using human anti-CD40 single-chain antibody fragments obtained from the n-CoDeR phage display library. Immunology 2002, 106, 456–463. [Google Scholar] [CrossRef]
- Yamniuk, A.P.; Suri, A.; Krystek, S.R.; Tamura, J.; Ramamurthy, V.; Kuhn, R.; Carroll, K.; Fleener, C.; Ryseck, R.; Cheng, L.; et al. Functional antagonism of human CD40 achieved by targeting a unique species-specific epitope. J. Mol. Biol. 2016, 428, 2860–2879. [Google Scholar] [CrossRef] [Green Version]
- Richman, L.P.; Vonderheide, R.H. Role of crosslinking for agonistic CD40 monoclonal antibodies as immune therapy of cancer. Cancer Immunol. Res. 2014, 2, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahan, R.; Barnhart, B.C.; Li, F.; Yamniuk, A.P.; Korman, A.J.; Ravetch, J.V. Therapeutic activity of agonistic, human anti-CD40 monoclonal antibodies requires selective Fc gamma R engagement. Cancer Cell. 2016, 29, 820–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Chan, H.T.C.; Orr, C.M.; Dadas, O.; Booth, S.G.; Dahal, L.N.; Penfold, C.A.; O’Brien, L.; Mockridge, C.I.; French, R.R.; et al. Complex interplay between epitope specificity and isotype dictates the biological activity of anti-human CD40 antibodies. Cancer Cell 2018, 33, 664–675 e664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonderheide, R.H.; Flaherty, K.T.; Khalil, M.; Stumacher, M.S.; Bajor, D.L.; Hutnick, N.A.; Sullivan, P.; Mahany, J.J.; Gallagher, M.; Kramer, A.; et al. Clinical activity and immune modulation in cancer patients treated with CP-870,893, a novel CD40 agonist monoclonal antibody. J. Clin. Oncol. 2007, 25, 876–883. [Google Scholar] [CrossRef]
- Bajor, D.L.; Xu, X.; Torigian, D.A.; Mick, R.; Garcia, L.R.; Richman, L.P.; Desmarais, C.; Nathanson, K.L.; Schuchter, L.M.; Kalos, M.; et al. Immune activation and a 9-year ongoing complete remission following CD40 antibody therapy and metastasectomy in a patient with metastatic melanoma. Cancer Immunol. Res. 2014, 2, 1051–1058. [Google Scholar] [CrossRef] [Green Version]
- Ruter, J.; Antonia, S.J.; Burris, H.A.; Huhn, R.D.; Vonderheide, R.H. Immune modulation with weekly dosing of an agonist CD40 antibody in a phase I study of patients with advanced solid tumors. Cancer Biol. Ther. 2010, 10, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Beatty, G.L.; Torigian, D.A.; Chiorean, E.G.; Saboury, B.; Brothers, A.; Alavi, A.; Troxel, A.B.; Sun, W.; Teitelbaum, U.R.; Vonderheide, R.H.; et al. A phase I study of an agonist CD40 monoclonal antibody (CP-870,893) in combination with gemcitabine in patients with advanced pancreatic ductal adenocarcinoma. Clin. Cancer Res. 2013, 19, 6286–6295. [Google Scholar] [CrossRef] [Green Version]
- Vonderheide, R.H.; Burg, J.M.; Mick, R.; Trosko, J.A.; Li, D.; Shaik, M.N.; Tolcher, A.W.; Hamid, O. Phase I study of the CD40 agonist antibody CP-870,893 combined with carboplatin and paclitaxel in patients with advanced solid tumors. Oncoimmunology 2013, 2, e23033. [Google Scholar] [CrossRef] [Green Version]
- Beatty, G.L.; Chiorean, E.G.; Fishman, M.P.; Saboury, B.; Teitelbaum, U.R.; Sun, W.; Huhn, R.D.; Song, W.; Li, D.; Sharp, L.L.; et al. CD40 agonists alter tumor stroma and show efficacy against pancreatic carcinoma in mice and humans. Science 2011, 331, 1612–1616. [Google Scholar] [CrossRef] [Green Version]
- Nowak, A.K.; Cook, A.M.; McDonnell, A.M.; Millward, M.J.; Creaney, J.; Francis, R.J.; Hasani, A.; Segal, A.; Musk, A.W.; Turlach, B.A.; et al. A phase 1b clinical trial of the CD40-activating antibody CP-870,893 in combination with cisplatin and pemetrexed in malignant pleural mesothelioma. Ann. Oncol. 2015, 26, 2483–2490. [Google Scholar] [CrossRef] [PubMed]
- Bajor, D.L.; Mick, R.; Riese, M.J.; Huang, A.C.; Sullivan, B.; Richman, L.P.; Torigian, D.A.; George, S.M.; Stelekati, E.; Chen, F.; et al. Long-term outcomes of a phase I study of agonist CD40 antibody and CLTA-4 blockade in patients with metastatic melanoma. Oncoimmunology 2018, 7, e1468956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machiels, J.P.; Gomez-Roca, C.; Michot, J.M.; Zamarin, D.; Mitchell, T.; Catala, G.; Eberst, L.; Jacob, W.; Jegg, A.M.; Cannarile, M.A.; et al. Phase Ib study of anti-CSF-1R antibody emactuzumab in combination with CD40 agonist selicrelumab in advanced solid tumor patients. J. Immunother. Cancer 2020, 8, e001153. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, M.H.; O’Reilly, E.M.; Varadhachary, G.; Wolff, R.A.; Wainberg, Z.A.; Ko, A.H.; Fisher, G.; Rahma, O.; Lyman, J.P.; Cabanski, C.R.; et al. CD40 agonistic monoclonal antibody APX005M (sotigalimab) and chemotherapy, with or without nivolumab, for the treatment of metastatic pancreatic adenocarcinoma: An open-label, multicentre, phase 1b study. Lancet Oncol. 2021, 22, 118–131. [Google Scholar] [CrossRef]
- Irenaeus, S.M.M.; Nielsen, D.; Ellmark, P.; Yachnin, J.; Deronic, A.; Nilsson, A.; Norlen, P.; Veitonmaki, N.; Wennersten, C.S.; Ullenhag, G.J. First-in-human study with intratumoral administration of a CD40 agonistic antibody, ADC-1013, in advanced solid malignancies. Int. J. Cancer 2019, 145, 1189–1199. [Google Scholar] [CrossRef]
- Johnson, P.; Challis, R.; Chowdhury, F.; Gao, Y.; Harvey, M.; Geldart, T.; Kerr, P.; Chan, C.; Smith, A.; Steven, N.; et al. Clinical and biological effects of an agonist anti-CD40 antibody: A cancer research UK phase I study. Clin. Cancer Res. 2015, 21, 1321–1328. [Google Scholar] [CrossRef] [Green Version]
- Vitale, L.A.; Thomas, L.J.; He, L.Z.; O’Neill, T.; Widger, J.; Crocker, A.; Sundarapandiyan, K.; Storey, J.R.; Forsberg, E.M.; Weidlick, J.; et al. Development of CDX-1140, an agonist CD40 antibody for cancer immunotherapy. Cancer Immunol. Immunother. 2019, 68, 233–245. [Google Scholar] [CrossRef]
- Sanborn, R.E.; Gabrail, N.Y.; Bhardwaj, N.; Gordon, M.S.; O’Hara, M.; Khalil, D.; Hawthorne, T.; Gedrich, R.; Vitale, L.; Rogalski, M.; et al. First-in-human phase I study of the CD40 agonist MAB CDX-1140 and in combination with CDX-301 (rhFLT3l) in patients with advanced cancers: Interim results. In Proceedings of the American Association for Cancer Research Annual Meeting, Atlanta, GA, USA, 29 March–3 April 2019. [Google Scholar]
- Kluger, H.; Weiss, S.A.; Olszanski, A.J.; Schuchter, L.; Linette, G.P.; Garland, L.; Iannotti, N.O.; Johnson, M.; Avsar, E.; Srivastava, M.K.; et al. Phase Ib/II of CD40 Agonistic Antibody APX005M in Combination with Nivolumab (Nivo) in Subjects with Metastatic Melanoma (M) or Non-Small Cell Lung Cancer (NSCLC). In Proceedings of the American Association for Cancer Research Annual Meeting, Atlanta, GA, USA, 29 March–3 April 2019. [Google Scholar]
- Mangsbo, S.M.; Broos, S.; Fletcher, E.; Veitonmaki, N.; Furebring, C.; Dahlen, E.; Norlen, P.; Lindstedt, M.; Totterman, T.H.; Ellmark, P. The human agonistic CD40 antibody ADC-1013 eradicates bladder tumors and generates T-cell-dependent tumor immunity. Clin. Cancer Res. 2015, 21, 1115–1126. [Google Scholar] [CrossRef] [Green Version]
- Fransen, M.F.; Sluijter, M.; Morreau, H.; Arens, R.; Melief, C.J. Local activation of CD8 T cells and systemic tumor eradication without toxicity via slow release and local delivery of agonistic CD40 antibody. Clin. Cancer Res. 2011, 17, 2270–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandin, L.C.; Orlova, A.; Gustafsson, E.; Ellmark, P.; Tolmachev, V.; Totterman, T.H.; Mangsbo, S.M. Locally delivered CD40 agonist antibody accumulates in secondary lymphoid organs and eradicates experimental disseminated bladder cancer. Cancer Immunol. Res. 2014, 2, 80–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, E.; Moreno, V.; Perets, R.; Yablonski-Peretz, T.; Fourneau, N.; Girgis, S.; Guo, Y.; Hellemans, P.; Hokey, D.; Pendas Franco, N.; et al. A phase I study to assess safety, pharmacokinetics (PK), and pharmacodynamics (PD) of JNJ-64457107, a CD40 agonistic monoclonal antibody, in patients (pts) with advanced solid tumors. J. Clin. Oncol. 2019, 37, 2527. [Google Scholar] [CrossRef]
- Lewis, T.S.; McCormick, R.S.; Emmerton, K.; Lau, J.T.; Yu, S.F.; McEarchern, J.A.; Grewal, I.S.; Law, C.L. Distinct apoptotic signaling characteristics of the anti-CD40 monoclonal antibody dacetuzumab and rituximab produce enhanced antitumor activity in non-hodgkin lymphoma. Clin. Cancer Res. 2011, 17, 4672–4681. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.; Berenson, J.R.; Niesvizky, R.; Munshi, N.; Matous, J.; Sobecks, R.; Harrop, K.; Drachman, J.G.; Whiting, N. A phase I multidose study of dacetuzumab (SGN-40; humanized anti-CD40 monoclonal antibody) in patients with multiple myeloma. Haematologica 2010, 95, 845–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grilley-Olson, J.E.; Curti, B.D.; Smith, D.C.; Goel, S.; Gajewski, T.; Markovic, S.; Rixe, O.; Bajor, D.L.; Gutierrez, M.; Kuzel, T.; et al. SEA-CD40, a non-fucosylated CD40 agonist: Interim results from a phase 1 study in advanced solid tumors. J. Clin. Oncol. 2018, 36, 3093. [Google Scholar] [CrossRef]

{kind=link}
| Pancreatic Adeno Carcinoma | Spearman Rho | Adjusted p-Value 1 | Melanoma | Spearman Rho | Adjusted p-Value 2 | Clear Cell Renal Cell Carcinoma | Spearman Rho | Adjusted p-Value 3 |
|---|---|---|---|---|---|---|---|---|
| ARHGDIB | 0.57 | 2.85 × 10−13 | COTL1 | 0.37 | 4.25 × 10−15 | NSFL1C | 0.35 | 1.76 × 10−14 |
| TNFAIP8L2 | 0.56 | 4.79 × 10−13 | LILRB5 | 0.37 | 4.25 × 10−15 | WAS | 0.35 | 1.76 × 10−14 |
| WAS | 0.56 | 4.79 × 10−13 | BTN3A1 | 0.37 | 25.0 × 10−15 | ZNF691 | 0.35 | 1.76 × 10−14 |
| NCF1 | 0.56 | 4.79 × 10−13 | C11orf93 | 0.37 | 4.25 × 10−15 | TRADD | 0.35 | 1.76 × 10−14 |
| DOK3 | 0.56 | 6.83 × 10−13 | SPAG4 | 0.37 | 4.25 × 10−15 | EMP3 | 0.35 | 1.76 × 10−14 |
| GPSM3 | 0.55 | 1.24 × 10−12 | STX4 | 0.37 | 4.25 × 10−15 | IL18BP | 0.35 | 1.76 × 10−14 |
| TREML1 | 0.55 | 1.24 × 10−12 | IDO2 | 0.37 | 4.25 × 10−15 | SQRDL | 0.34 | 1.76 × 10−14 |
| HLA-DPB1 | 0.55 | 1.27 × 10−12 | EID3 | 0.37 | 4.25 × 10−15 | CXCR4 | 0.34 | 1.76 × 10−14 |
| FGD3 | 0.55 | 1.27 × 10−12 | MOBKL1B | 0.37 | 4.25 × 10−15 | LOC84856 | 0.34 | 1.76 × 10−14 |
| SIPA1 | 0.55 | 1.27 × 10−12 | SYNE1 | 0.37 | 4.25 × 10−15 | RPL11 | 0.34 | 3.17 × 10−14 |
| ARHGAP9 | 0.55 | 1.35 × 10−12 | JAK1 | 0.37 | 8.39 × 10−15 | SH3BP1 | 0.34 | 3.17 × 10−14 |
| LSP1 | 0.55 | 1.35 × 10−12 | IL4 | 0.37 | 8.39 × 10−15 | TRNAU1AP | 0.34 | 3.17 × 10−14 |
| SP110 | 0.55 | 1.46 × 10−12 | MAN1A1 | 0.37 | 8.39 × 10−15 | RSPH9 | 0.34 | 3.17 × 10−14 |
| BTN2A2 | 0.55 | 1.57 × 10−12 | CASS4 | 0.37 | 8.39 × 10−15 | IRF1 | 0.34 | 3.17 × 10−14 |
| CD53 | 0.55 | 1.76 × 10−12 | GPR55 | 0.37 | 8.39 × 10−15 | CD48 | 0.34 | 3.17 × 10−14 |
| GMFG | 0.55 | 1.76 × 10−12 | HLA-DPB2 | 0.37 | 8.39 × 10−15 | PRR14 | 0.34 | 3.17 × 10−14 |
| CD72 | 0.55 | 1.93 × 10−12 | C19orf36 | 0.37 | 8.39 × 10−15 | TRIM55 | 0.34 | 3.17 × 10−14 |
| SASH3 | 0.55 | 1.93 × 10−12 | GRAP2 | 0.37 | 8.39 × 10−15 | CCDC102A | 0.34 | 4.65 × 10−14 |
| MCOLN2 | 0.54 | 2.86 × 10−12 | XBP1 | 0.37 | 8.39 × 10−15 | EML3 | 0.34 | 4.65 × 10−14 |
| CORO1A | 0.54 | 2.86 × 10−12 | BIN1 | 0.37 | 8.39 × 10−15 | C5orf56 | 0.34 | 4.65 × 10−14 |
| SIRPG | 0.54 | 2.95 × 10−12 | RAB8B | 0.37 | 8.39 × 10−15 | C21orf63 | 0.34 | 5.85 × 10−14 |
| HLA-DMB | 0.54 | 2.95 × 10−12 | ATF7IP2 | 0.37 | 8.39 × 10−15 | PRKD2 | 0.34 | 5.85 × 10−14 |
| NECAP2 | 0.54 | 3.27 × 10−12 | VAMP5 | 0.37 | 8.39 × 10−15 | CYB5R3 | 0.34 | 5.85 × 10−14 |
| BTK | 0.54 | 3.27 × 10−12 | GNAI2 | 0.37 | 8.39 × 10−15 | ZNRD1 | 0.34 | 5.85 × 10−14 |
| JAK3 | 0.54 | 3.30 × 10−12 | RAB8A | 0.37 | 1.25 × 10−14 | RGS12 | 0.34 | 8.56 × 10−14 |
| PARVG | 0.54 | 3.55 × 10−12 | KLHL33 | 0.37 | 1.25 × 10−14 | PSMB8 | 0.34 | 1.11 × 10−13 |
| KMO | 0.54 | 3.69 × 10−12 | C1orf38 | 0.37 | 1.25 × 10−14 | RNF113A | 0.33 | 1.33 × 10−13 |
| ARHGAP25 | 0.54 | 4.35 × 10−12 | MMP25 | 0.37 | 1.25 × 10−14 | HYI | 0.33 | 1.33 × 10−13 |
| RAC2 | 0.54 | 4.35 × 10−12 | CDRT4 | 0.37 | 1.66 × 10−14 | RIBC1 | 0.33 | 1.33 × 10−13 |
| PLEK | 0.54 | 4.86 × 10−12 | CYB5R4 | 0.37 | 1.66 × 10−14 | BATF2 | 0.33 | 1.56 × 10−13 |
| NAPSB | 0.54 | 5.74 × 10−12 | GBGT1 | 0.37 | 1.66 × 10−14 | AP4M1 | 0.33 | 1.56 × 10−13 |
| ITGAL | 0.54 | 6.00 × 10−12 | CAPZB | 0.37 | 1.66 × 10−14 | NDUFA4L2 | 0.33 | 1.80 × 10−13 |
| FGD2 | 0.53 | 6.26 × 10−12 | VAMP8 | 0.37 | 1.66 × 10−14 | HSPB11 | 0.33 | 1.80 × 10−13 |
| MYO1G | 0.53 | 6.83 × 10−12 | GNLY | 0.36 | 1.66 × 10−14 | RHOG | 0.33 | 2.16 × 10−13 |
| HMHA1 | 0.53 | 6.83 × 10−12 | UGT2B15 | 0.36 | 2.08 × 10−14 | DEF6 | 0.33 | 2.24 × 10−13 |
| GNGT2 | 0.53 | 6.83 × 10−12 | GPR84 | 0.36 | 2.48 × 10−14 | TMEM44 | 0.33 | 2.24 × 10−13 |
| PDCD1 | 0.53 | 6.83 × 10−12 | TC2N | 0.36 | 2.48 × 10−14 | PPIE | 0.33 | 2.24 × 10−13 |
| CD52 | 0.53 | 7.21 × 10−12 | CSF1 | 0.36 | 2.48 × 10−14 | TMSB10 | 0.33 | 2.35 × 10−13 |
| CTLA4 | 0.53 | 7.27 × 10−12 | EMR4P | 0.36 | 2.48 × 10−14 | CLIC1 | 0.33 | 2.64 × 10−13 |
| CTSW | 0.53 | 7.27 × 10−12 | TMEM155 | 0.36 | 2.48 × 10−14 | INSL3 | 0.33 | 2.64 × 10−13 |
| ARHGAP30 | 0.53 | 7.27 × 10−12 | TMEM37 | 0.36 | 2.48 × 10−14 | TICAM2 | 0.33 | 2.73 × 10−13 |
| ANKRD58 | 0.53 | 7.27 × 10−12 | SNX29 | 0.36 | 2.89 × 10−14 | TBX21 | 0.33 | 2.73 × 10−13 |
| STK17A | 0.53 | 7.72 × 10−12 | C12orf77 | 0.36 | 3.29 × 10−14 | NFKB2 | 0.33 | 2.83 × 10−13 |
| VAV1 | 0.53 | 7.72 × 10−12 | NCRNA00204B | 0.36 | 3.29 × 10−14 | ALKBH2 | 0.33 | 3.24 × 10−13 |
| CD2 | 0.53 | 8.50 × 10−12 | KCNN3 | 0.36 | 3.29 × 10−14 | KIAA1949 | 0.33 | 3.45 × 10−13 |
| RASAL3 | 0.53 | 8.73 × 10−12 | CLEC4M | 0.36 | 3.70 × 10−14 | IRF9 | 0.33 | 4.05 × 10−13 |
| Study/ Reference | Phase | Status | CD40 Agonist | Cancer Type | Response |
|---|---|---|---|---|---|
| [95] | 1 | Completed | Selicrelumab | Solid tumors | 14% PR, 24% SD |
| [97] | 1 | Completed | Selicrelumab | Solid tumors | 26% SD |
| [98,100] | 1 | Completed | Selicrelumab + gemcitabine | PDAC | 19% PR, 52% SD |
| [99] | 1 | Completed | Selicrelumab + paclitaxel and carboplatin | Solid tumors | 20% PR,40% SD |
| [101] | 1b | Completed | Selicrelumab + cisplatin pemetrexed | Mesothelioma | 40% PR, 53% SD |
| [102] | 1 | Completed | Selicrelumab + tremelimumab | Melanoma | 27% ORR, 9%CR, 18% PR |
| [103] | 1b | Completed | Selicrelumab + emactuzumab | Solid tumors | 40.5% SD |
| NCT03892525 | 1 | Suspended | Selicrelumab + atezolizumab | B Cell Lymphoma | NR |
| NCT02304393 | 1 | Completed | Selicrelumab + atezolizumab | Solid tumors | NR |
| NCT02665416 | 1 | Completed | Selicrelumab + vanucizumab or bevacizumab | Metastatic solid tumors | NR |
| NCT03193190 | 1b/2 | Recruiting | Selicrelumab + chemotherapy + atezolizumab | PDAC | NR |
| NCT03424005 | 1b/2 | Recruiting | Selicrelumab + atezolizumab + bevacizumab | Breast Cancer | NR |
| NCT03555149 | 1b/2 | Recruiting | Selicrelumab + bevacizumab + atezolizumab | CRC | NR |
| NCT04364230 | 1/2 | Recruiting | peptide vaccine with CDX-1140 and TLR3 agonists | Melanoma | NR |
| NCT03329950 | 1 | Recruiting | CDX-1140, CDX-301,pembrolizumab, chemotherapy | Advanced Malignancies | NR |
| NCT04536077 | 2 | Not yet recruiting | CDX-1140, CDX-301 | Pancreatic Cancer | NR |
| NCT04616248 | 1 | Not yet recruiting | CDX-1140, CDX-301, radiotherapy, and Poly-ICLC | Breast Cancer | NR |
| NCT04491084 | 1/2 | Not yet recruiting | CDX-1140, CDX-301 and SBRT | NSCLC | NR |
| NCT04520711 | 1/1b | Not yet recruiting | TCR-transduced T cells, CDX-1140, pembrolizumab | Epithelial Neoplasms | NR |
| NCT02482168 | 1 | Completed | APX005M | Solid tumors | NR |
| [104] | 2 | Completed | APX005M, gemcitabine, nab-paclitaxel, nivolumab | Pancreatic adeno ca. | 58% PR, 30% SD |
| NCT03123783 | 2 | Ongoing | APX005M + nivolumab | Melanoma, lung cancer | NR |
| NCT03502330 | 1 | Ongoing | APX005M + cabiralizumab w/w.o nivolumab | RCC, melanoma, NSCLC | NR |
| NCT03719430 | 2 | Recruiting | APX005M + doxorubicin | Advanced soft tissue ca. | NR |
| NCT03165994 | 2 | Recruiting | APX005M + radiation, paclitaxel, carboplatin | Esophageal cancers | NR |
| NCT02706353 | 2 | Recruiting | APX005M + pembrolizumab | Metastatic melanoma | 20% PR, 20% SD |
| NCT03389802 | 1 | Recruiting | APX005M | Pediatric CNS tumors | NR |
| NCT04337931 | 2 | Recruiting | APX005M | Melanoma | NR |
| NCT04130854 | 2 | Recruiting | Neoadjuvant therapy with and without APX005M | Rectal adeno ca | NR |
| NCT04495257 | 1 | Recruiting | APX005M, nivolumab and ipilimumab | Melanoma and RCC | NR |
| [105] | 1 | Completed | ADC-1013 (i.t vs. i.v) | Solid tumors | i.t well tolerated |
| NCT02379741 | 1 | Completed | ADC-1013 | Solid Tumors | NR |
| NCT02829099 | 1 | Ongoing | ADC-1013 | Solid tumors | 1% PR, 11–14% SD |
| NCT02376699 | 1 | Recruiting | SEA-CD40, pembrolizumab, nab-paclitaxel | Advanced malignancies | NR |
| [106] | 1 | Completed | ChiLob 7/4 | Solid tumors | SD |
| NCT01561911 | 1 | Completed | ChiLob 7/4 | Advanced malignancies | NR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djureinovic, D.; Wang, M.; Kluger, H.M. Agonistic CD40 Antibodies in Cancer Treatment. Cancers 2021, 13, 1302. https://doi.org/10.3390/cancers13061302
Djureinovic D, Wang M, Kluger HM. Agonistic CD40 Antibodies in Cancer Treatment. Cancers. 2021; 13(6):1302. https://doi.org/10.3390/cancers13061302
Chicago/Turabian StyleDjureinovic, Dijana, Meina Wang, and Harriet M. Kluger. 2021. "Agonistic CD40 Antibodies in Cancer Treatment" Cancers 13, no. 6: 1302. https://doi.org/10.3390/cancers13061302