
Pentraxin 3 Inhibits the Angiogenic Potential of Multiple Myeloma Cells

 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Reagents
2.2. Western Blot Analysis
2.3. RT-qPCR
2.4. Gene Expression Profiling (GEP)
2.5. Cytofluorimetric Analyses
2.6. MM/HUVE Cells Co-Cultures
2.7. In Vitro Immunofluorescence Analysis
2.8. Chick Embryo Chorioallantoic Membrane (CAM) Assay
2.9. Zebrafish Embryo Model
2.10. Subcutaneous Human Xenografts
2.11. Systemic Human Xenograft
2.12. Histological Analyses
2.13. Two-Photon Microscopy
2.14. Statistical Analyses
3. Results
3.1. PTX3 Produced by MM Cells Hampers the Proliferation of Endothelial Cells
3.2. PTX3 Reduces the Angiogenic Potential of MM Cells
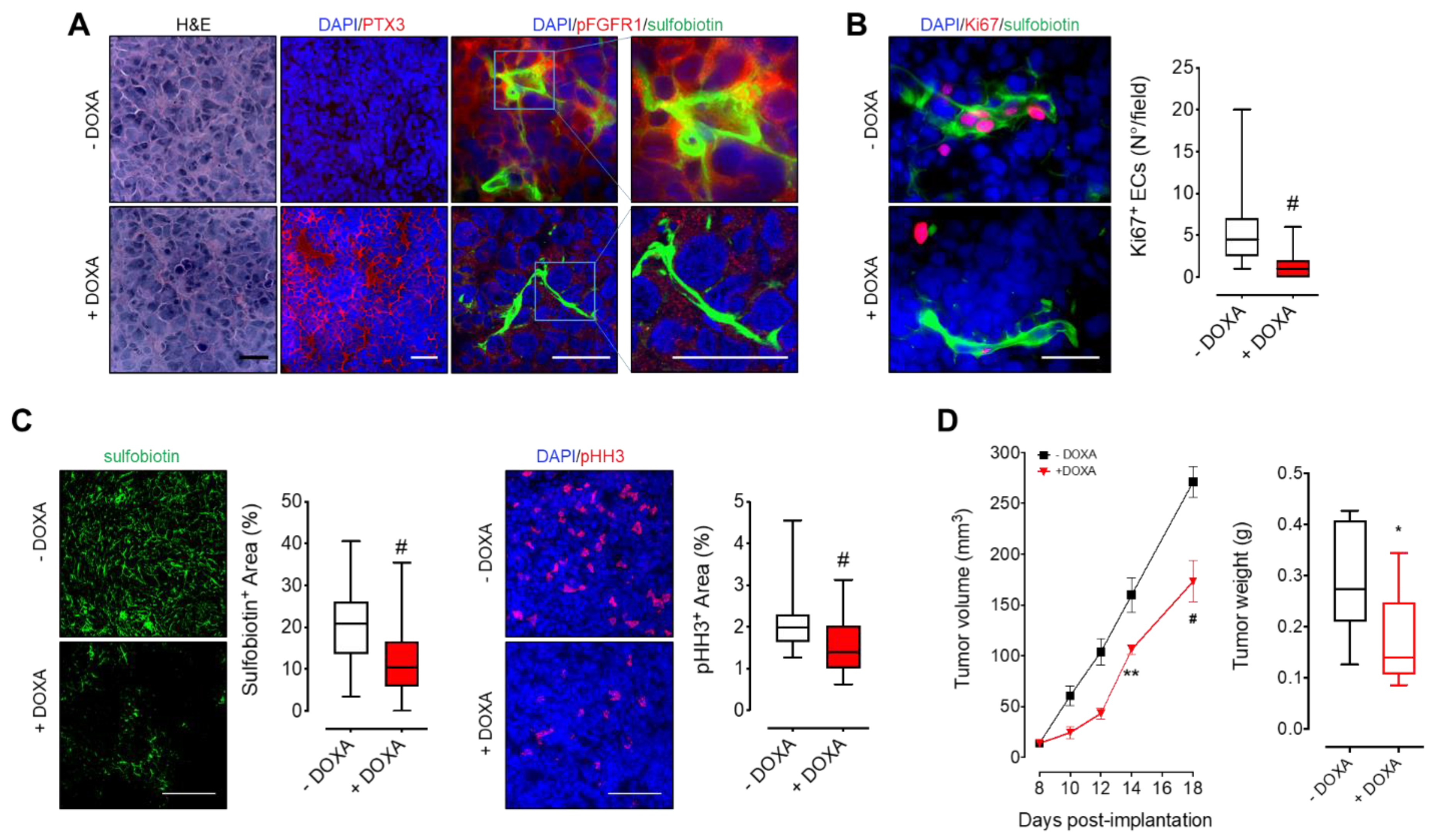
3.3. MM-Released PTX3 Inhibits Tumor Angiogenesis and Growth In Vivo
3.4. PTX3 Reduces BM Niche Vascularization and Colonization by MM Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreau, P.; Kumar, S.K.; Miguel, J.S.; Davies, F.; Zamagni, E.; Bahlis, N.; Ludwig, H.; Mikhael, J.; Terpos, E.; Schjesvold, F.; et al. Treatment of relapsed and refractory multiple myeloma: Recommendations from the International Myeloma Working Group. Lancet Oncol. 2021, 22, e105–e118. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A. New insights in anti-angiogenesis in multiple myeloma. Int. J. Mol. Sci. 2018, 19, 2031. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Vacca, A. Importance of the bone marrow microenvironment in inducing the angiogenic response in multiple myeloma. Oncogene 2006, 25, 4257–4266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Witzig, T.E.; Timm, M.; Haug, J.; Wellik, L.; Kimlinger, T.K.; Greipp, P.R.; Rajkumar, S.V. Bone marrow angiogenic ability and expression of angiogenic cytokines in myeloma: Evidence favoring loss of marrow angiogenesis inhibitory activity with disease progression. Blood 2004, 104, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, S.V.; Mesa, R.A.; Fonseca, R.; Schroeder, G.; Plevak, M.F.; Dispenzieri, A.; Lacy, M.Q.; Lust, J.A.; Witzig, T.E.; Gertz, M.A.; et al. Bone marrow angiogenesis in 400 patients with monoclonal gammopathy of undetermined significance, multiple myeloma, and primary amyloidosis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2002, 8, 2210–2216. [Google Scholar]
- Jakob, C.; Sterz, J.; Zavrski, I.; Heider, U.; Kleeberg, L.; Fleissner, C.; Kaiser, M.; Sezer, O. Angiogenesis in multiple myeloma. Eur. J. Cancer 2006, 42, 1581–1590. [Google Scholar] [CrossRef]
- Vacca, A.; Ribatti, D. Bone marrow angiogenesis in multiple myeloma. Leukemia 2006, 20, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Ronca, R.; Benkheil, M.; Mitola, S.; Struyf, S.; Liekens, S. Tumor angiogenesis revisited: Regulators and clinical implications. Med. Res. Rev. 2017, 37, 1231–1274. [Google Scholar] [CrossRef] [Green Version]
- Presta, M.; Chiodelli, P.; Giacomini, A.; Rusnati, M.; Ronca, R. Fibroblast growth factors (FGFs) in cancer: FGF traps as a new therapeutic approach. Pharmacol. Ther. 2017, 179, 171–187. [Google Scholar] [CrossRef]
- Sacco, A.; Federico, C.; Giacomini, A.; Caprio, C.; Maccarinelli, F.; Todoerti, K.; Favasuli, V.; Anastasia, A.; Motta, M.; Russo, D.; et al. Halting the FGF/FGFR axis leads to anti-tumor activity in Waldenstrom′s macroglobulinemia by silencing MYD88. Blood 2020. [Google Scholar] [CrossRef]
- Ronca, R.; Ghedini, G.C.; Maccarinelli, F.; Sacco, A.; Locatelli, S.L.; Foglio, E.; Taranto, S.; Grillo, E.; Matarazzo, S.; Castelli, R.; et al. FGF trapping inhibits multiple myeloma growth through c-Myc degradation-induced mitochondrial oxidative stress. Cancer Res. 2020, 80, 2340–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomini, A.; Ghedini, G.C.; Presta, M.; Ronca, R. Long pentraxin 3: A novel multifaceted player in cancer. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 53–63. [Google Scholar] [CrossRef]
- Annese, T.; Ronca, R.; Tamma, R.; Giacomini, A.; Ruggieri, S.; Grillo, E.; Presta, M.; Ribatti, D. PTX3 modulates neovascularization and immune inflammatory infiltrate in a murine model of fibrosarcoma. Int. J. Mol. Sci. 2019, 20, 4599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzola, S.; Ronca, R.; Loda, A.; Nawaz, M.I.; Tobia, C.; Paganini, G.; Maccarinelli, F.; Giacomini, A.; Semeraro, F.; Mor, M.; et al. The autocrine FGF/FGFR System in both skin and uveal melanoma: FGF trapping as a possible therapeutic approach. Cancers 2019, 11, 1305. [Google Scholar] [CrossRef] [Green Version]
- Matarazzo, S.; Melocchi, L.; Rezzola, S.; Grillo, E.; Maccarinelli, F.; Giacomini, A.; Turati, M.; Taranto, S.; Zammataro, L.; Cerasuolo, M.; et al. Long pentraxin-3 follows and modulates bladder cancer progression. Cancers 2019, 11, 1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, P.F.; Matarazzo, S.; Maccarinelli, F.; Foglio, E.; Giacomini, A.; Nunes, J.P.S.; Presta, M.; Dias, A.A.M.; Ronca, R. Long pentraxin 3-mediated fibroblast growth factor trapping impairs fibrosarcoma growth. Front. Oncol. 2018, 8, 472. [Google Scholar] [CrossRef] [Green Version]
- Presta, M.; Foglio, E.; Schuind, A.C.; Ronca, R. Long pentraxin-3 modulates the angiogenic activity of fibroblast growth factor-2. Front. Immunol. 2018, 9, 2327. [Google Scholar] [CrossRef] [Green Version]
- Ronca, R.; Giacomini, A.; Di Salle, E.; Coltrini, D.; Pagano, K.; Ragona, L.; Matarazzo, S.; Rezzola, S.; Maiolo, D.; Torrella, R.; et al. Long-pentraxin 3 derivative as a small-molecule fgf trap for cancer therapy. Cancer Cell 2015, 28, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Giacomini, A.; Taranto, S.; Rezzola, S.; Matarazzo, S.; Grillo, E.; Bugatti, M.; Scotuzzi, A.; Guerra, J.; Di Trani, M.; Presta, M.; et al. Inhibition of the FGF/FGFR system induces apoptosis in lung cancer cells via c-Myc downregulation and oxidative stress. Int. J. Mol. Sci. 2020, 21, 9376. [Google Scholar] [CrossRef]
- Castelli, R.; Giacomini, A.; Anselmi, M.; Bozza, N.; Vacondio, F.; Rivara, S.; Matarazzo, S.; Presta, M.; Mor, M.; Ronca, R. Synthesis, structural elucidation, and biological evaluation of NSC12, an orally available fibroblast growth factor (FGF) ligand trap for the treatment of FGF-dependent lung tumors. J. Med. Chem. 2016, 59, 4651–4663. [Google Scholar] [CrossRef] [Green Version]
- Basile, A.; Moschetta, M.; Ditonno, P.; Ria, R.; Marech, I.; De Luisi, A.; Berardi, S.; Frassanito, M.A.; Angelucci, E.; Derudas, D.; et al. Pentraxin 3 (PTX3) inhibits plasma cell/stromal cell cross-talk in the bone marrow of multiple myeloma patients. J. Pathol. 2013, 229, 87–98. [Google Scholar] [CrossRef]
- Ribatti, D.; Gualandris, A.; Bastaki, M.; Vacca, A.; Iurlaro, M.; Roncali, L.; Presta, M. New model for the study of angiogenesis and antiangiogenesis in the chick embryo chorioallantoic membrane: The gelatin sponge/chorioallantoic membrane assay. J. Vasc. Res. 1997, 34, 455–463. [Google Scholar] [CrossRef]
- Corsini, M.; Moroni, E.; Ravelli, C.; Grillo, E.; Presta, M.; Mitola, S. In situ DNA/protein interaction assay to visualize transcriptional factor activation. Methods Protoc. 2020, 3, 80. [Google Scholar] [CrossRef] [PubMed]
- Ronca, R.; Benzoni, P.; Leali, D.; Urbinati, C.; Belleri, M.; Corsini, M.; Alessi, P.; Coltrini, D.; Calza, S.; Presta, M.; et al. Antiangiogenic activity of a neutralizing human single-chain antibody fragment against fibroblast growth factor receptor 1. Mol. Cancer Ther. 2010, 9, 3244–3253. [Google Scholar] [CrossRef] [Green Version]
- Lavazza, C.; Carlo-Stella, C.; Giacomini, A.; Cleris, L.; Righi, M.; Sia, D.; Di Nicola, M.; Magni, M.; Longoni, P.; Milanesi, M.; et al. Human CD34+ cells engineered to express membrane-bound tumor necrosis factor-related apoptosis-inducing ligand target both tumor cells and tumor vasculature. Blood 2010, 115, 2231–2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belleri, M.; Coltrini, D.; Righi, M.; Ravelli, C.; Taranto, S.; Chiodelli, P.; Mitola, S.; Presta, M.; Giacomini, A. β-galactosylceramidase deficiency causes bone marrow vascular defects in an animal model of Krabbe disease. Int. J. Mol. Sci. 2019, 21, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronca, R.; Taranto, S.; Corsini, M.; Tobia, C.; Ravelli, C.; Rezzola, S.; Belleri, M.; De Cillis, F.; Cattaneo, A.; Presta, M.; et al. Pentraxin 3 Inhibits the Angiogenic Potential of Multiple Myeloma Cells. Cancers 2021, 13, 2255. https://doi.org/10.3390/cancers13092255
Ronca R, Taranto S, Corsini M, Tobia C, Ravelli C, Rezzola S, Belleri M, De Cillis F, Cattaneo A, Presta M, et al. Pentraxin 3 Inhibits the Angiogenic Potential of Multiple Myeloma Cells. Cancers. 2021; 13(9):2255. https://doi.org/10.3390/cancers13092255
Chicago/Turabian StyleRonca, Roberto, Sara Taranto, Michela Corsini, Chiara Tobia, Cosetta Ravelli, Sara Rezzola, Mirella Belleri, Floriana De Cillis, Annamaria Cattaneo, Marco Presta, and et al. 2021. "Pentraxin 3 Inhibits the Angiogenic Potential of Multiple Myeloma Cells" Cancers 13, no. 9: 2255. https://doi.org/10.3390/cancers13092255
APA StyleRonca, R., Taranto, S., Corsini, M., Tobia, C., Ravelli, C., Rezzola, S., Belleri, M., De Cillis, F., Cattaneo, A., Presta, M., & Giacomini, A. (2021). Pentraxin 3 Inhibits the Angiogenic Potential of Multiple Myeloma Cells. Cancers, 13(9), 2255. https://doi.org/10.3390/cancers13092255