
Predictive and Prognostic Value of Non-Coding RNA in Breast Cancer

 ,
,  ,
,  , and
, and 
Abstract
Simple Summary
Abstract
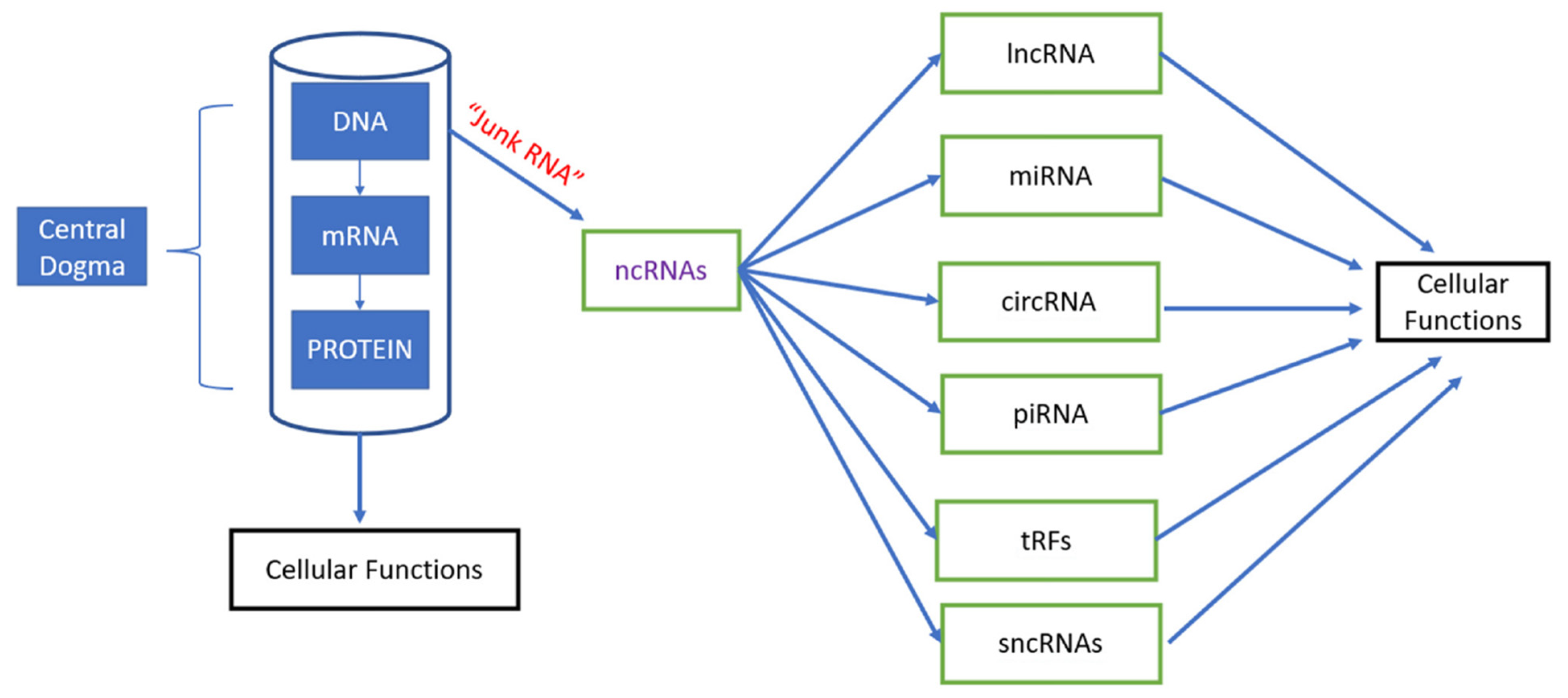
1. Introduction
2. Long Non-Coding RNA Molecular Mechanisms and Cancer Involvement
2.1. LncRNA Nuclear Accumulation, Splicing, and Gene Regulation
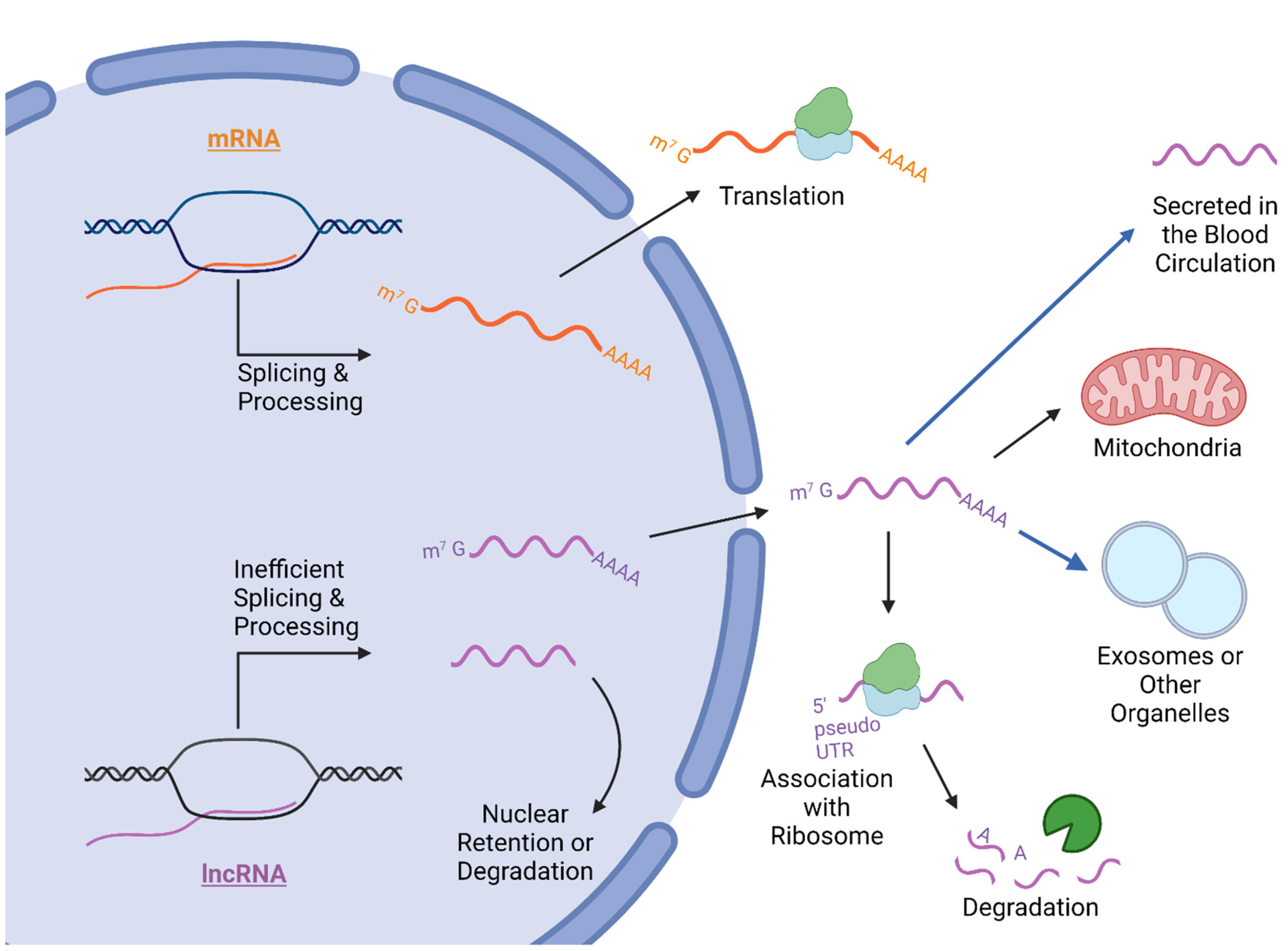
2.2. Export and Cellular Compartmentalization of LncRNA
2.3. LncRNA in Cancer Processes and Patient Serum
3. Clinical Value of Non-Coding RNAs in BC
4. Discussions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- On Protein Synthesis-PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/13580867/ (accessed on 20 January 2022).
- Crick, F. Central Dogma of Molecular Biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Non–coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquienelll, A.E.; Bettlnger, J.C.; Rougvle, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Bejerano, G.; Pheasant, M.; Makunin, I.; Stephen, S.; Kent, W.J.; Mattick, J.S.; Haussler, D. Ultraconserved elements in the human genome. Science 2004, 304, 1321–1325. [Google Scholar] [CrossRef]
- Johnsson, P.; Lipovich, L.; Grandér, D.; Morris, K.V. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 1063–1071. [Google Scholar] [CrossRef]
- Shan, N.; Li, N.; Dai, Q.; Hou, L.; Yan, X.; Amei, A.; Lu, L.; Wang, Z. Interplay of tRNA-Derived Fragments and T Cell Activation in Breast Cancer Patient Survival. Cancers 2020, 12, 2230. [Google Scholar] [CrossRef]
- Li, N.; Shan, N.; Lu, L.; Wang, Z. tRFtarget: A database for transfer RNA-derived fragment targets. Nucleic Acids Res. 2021, 49, D254–D260. [Google Scholar] [CrossRef]
- Mendell, J.T. MicroRNAs: Critical regulators of development, cellular physiology and malignancy. Cell Cycle 2005, 4, 1179–1184. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Aravin, A.A.; Sachidanandam, R.; Girard, A.; Fejes-Toth, K.; Hannon, G.J. Developmentally regulated piRNA clusters implicate MILI in transposon control. Science 2007, 316, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2017, 18, 5–18. [Google Scholar] [CrossRef]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- Kapranov, P.; St Laurent, G.; Raz, T.; Ozsolak, F.; Reynolds, C.P.; Sorensen, P.H.B.; Reaman, G.; Milos, P.; Arceci, R.J.; Thompson, J.F.; et al. The majority of total nuclear-encoded non-ribosomal RNA in a human cell is “dark matter” un-annotated RNA. BMC Biol. 2010, 8, 149. [Google Scholar] [CrossRef]
- Huang, J.; Wu, S.; Wang, P.; Wang, G. Non-coding RNA Regulated Cross-Talk Between Mitochondria and Other Cellular Compartments. Front. Cell Dev. Biol. 2021, 9, 2095. [Google Scholar] [CrossRef]
- Dunham, I.; Kundaje, A.; Aldred, S.F.; Collins, P.J.; Davis, C.A.; Doyle, F.; Epstein, C.B.; Frietze, S.; Harrow, J.; Kaul, R.; et al. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2020, 22, 96–118. [Google Scholar] [CrossRef]
- Guo, C.J.; Ma, X.K.; Xing, Y.H.; Zheng, C.C.; Xu, Y.F.; Shan, L.; Zhang, J.; Wang, S.; Wang, Y.; Carmichael, G.G.; et al. Distinct Processing of lncRNAs Contributes to Non-conserved Functions in Stem Cells. Cell 2020, 181, 621–636.e22. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775. [Google Scholar] [CrossRef] [PubMed]
- Schlackow, M.; Nojima, T.; Gomes, T.; Dhir, A.; Carmo-Fonseca, M.; Proudfoot, N.J. Distinctive Patterns of Transcription and RNA Processing for Human lincRNAs. Mol. Cell 2017, 65, 25. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Lu, J.Y.; Zhang, X.; Shao, W.; Xu, Y.; Li, P.; Hong, Y.; Cui, L.; Shan, G.; Tian, B.; et al. U1 snRNP regulates chromatin retention of noncoding RNAs. Nature 2020, 580, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Nojima, T.; Tellier, M.; Foxwell, J.; Ribeiro de Almeida, C.; Tan-Wong, S.M.; Dhir, S.; Dujardin, G.; Dhir, A.; Murphy, S.; Proudfoot, N.J. Deregulated Expression of Mammalian lncRNA through Loss of SPT6 Induces R-Loop Formation, Replication Stress, and Cellular Senescence. Mol. Cell 2018, 72, 970. [Google Scholar] [CrossRef] [PubMed]
- Vos, S.M.; Farnung, L.; Boehning, M.; Wigge, C.; Linden, A.; Urlaub, H.; Cramer, P. Structure of activated transcription complex Pol II–DSIF–PAF–SPT6. Nature 2018, 560, 607–612. [Google Scholar] [CrossRef]
- Khan, M.; Hou, S.; Azam, S.; Lei, H. Sequence-dependent recruitment of SRSF1 and SRSF7 to intronless lncRNA NKILA promotes nuclear export via the TREX/TAP pathway. Nucleic Acids Res. 2021, 49, 6420. [Google Scholar] [CrossRef]
- Zuckerman, B.; Ron, M.; Mikl, M.; Segal, E.; Ulitsky, I. Gene Architecture and Sequence Composition Underpin Selective Dependency of Nuclear Export of Long RNAs on NXF1 and the TREX Complex. Mol. Cell 2020, 79, 251–267. [Google Scholar] [CrossRef]
- Li, J.; Tong, H.; Li, D.; Jiang, Q.; Zhang, Y.; Tang, W.; Jin, D.; Chen, S.; Qin, X.; Zhang, S.; et al. The long non-coding RNA DKFZp434J0226 regulates the alternative splicing process through phosphorylation of SF3B6 in PDAC. Mol. Med. 2021, 27, 95. [Google Scholar] [CrossRef]
- Lee, J.T. Lessons from X-chromosome inactivation: Long ncRNA as guides and tethers to the epigenome. Genes Dev. 2009, 23, 1831. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Wutz, A. Gene silencing in X-chromosome inactivation: Advances in understanding facultative heterochromatin formation. Nat. Rev. Genet. 2011, 12, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Brockdorff, N.; Bowness, J.S.; Wei, G. Progress toward understanding chromosome silencing by Xist RNA. Genes Dev. 2020, 34, 733. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Das, S.; Das, B. Localization elements and zip codes in the intracellular transport and localization of messenger RNAs in Saccharomyces cerevisiae. Wiley Interdiscip. Rev. RNA 2020, 11, e1591. [Google Scholar] [CrossRef] [PubMed]
- Lubelsky, Y.; Ulitsky, I. Sequences enriched in Alu repeats drive nuclear localization of long RNAs in human cells. Nature 2018, 555, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Hou, S.; Zhu, B.; Wang, W.; Hao, T.; Bu, X.; Khan, M.; Lei, H. Nuclear retention element recruits U1 snRNP components to restrain spliced lncRNAs in the nucleus. RNA Biol. 2019, 16, 1001. [Google Scholar] [CrossRef]
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. exoRBase: A database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2018, 46, D106. [Google Scholar] [CrossRef]
- Statello, L.; Maugeri, M.; Garre, E.; Nawaz, M.; Wahlgren, J.; Papadimitriou, A.; Lundqvist, C.; Lindfors, L.; Collén, A.; Sunnerhagen, P.; et al. Identification of RNA-binding proteins in exosomes capable of interacting with different types of RNA: RBP-facilitated transport of RNAs into exosomes. PLoS ONE 2018, 13, e0195969. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Hoffman, A.R.; Cui, J.; Hu, J.F. The Nucleus/Mitochondria-Shuttling LncRNAs Function as New Epigenetic Regulators of Mitophagy in Cancer. Front. Cell Dev. Biol. 2021, 9, 2519. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Q.; Ji, M.; Guo, X.; Li, L.; Su, X. Exosomal lncRNA UCA1 modulates cervical cancer stem cell self-renewal and differentiation through microRNA-122-5p/SOX2 axis. J. Transl. Med. 2021, 19, 229. [Google Scholar] [CrossRef]
- Dai, W.; Jin, X.; Han, L.; Huang, H.; Ji, Z.; Xu, X.; Tang, M.; Jiang, B.; Chen, W. Exosomal lncRNA DOCK9-AS2 derived from cancer stem cell-like cells activated Wnt/β-catenin pathway to aggravate stemness, proliferation, migration, and invasion in papillary thyroid carcinoma. Cell Death Dis. 2020, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Hamada, M. Identifying sequence features that drive ribosomal association for lncRNA. BMC Genom. 2018, 19, 906. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Song, R.; Regev, A.; Struhl, K. Many lncRNAs, 5’UTRs, and pseudogenes are translated and some are likely to express functional proteins. eLife 2015, 4, e08890. [Google Scholar] [CrossRef] [PubMed]
- Bazin, J.; Baerenfaller, K.; Gosai, S.J.; Gregory, B.D.; Crespi, M.; Bailey-Serres, J. Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E10018–E10027. [Google Scholar] [CrossRef]
- Pircher, A.; Gebetsberger, J.; Polacek, N. Ribosome-associated ncRNAs: An emerging class of translation regulators. RNA Biol. 2014, 11, 1335. [Google Scholar] [CrossRef]
- Carlevaro-Fita, J.; Rahim, A.; Guigó, R.; Vardy, L.A.; Johnson, R. Cytoplasmic long noncoding RNAs are frequently bound to and degraded at ribosomes in human cells. RNA 2016, 22, 867–882. [Google Scholar] [CrossRef]
- Mercer, T.R.; Neph, S.; Dinger, M.E.; Crawford, J.; Smith, M.A.; Shearwood, A.M.J.; Haugen, E.; Bracken, C.P.; Rackham, O.; Stamatoyannopoulos, J.A.; et al. The human mitochondrial transcriptome. Cell 2011, 146, 645–658. [Google Scholar] [CrossRef]
- Rackham, O.; Shearwood, A.M.J.; Mercer, T.R.; Davies, S.M.K.; Mattick, J.S.; Filipovska, A. Long noncoding RNAs are generated from the mitochondrial genome and regulated by nuclear-encoded proteins. RNA 2011, 17, 2085. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; Abdelmohsen, K.; Yoon, J.H.; Panda, A.C.; Munk, R.; Kim, J.; Curtis, J.; Moad, C.A.; Wohler, C.M.; et al. HuR and GRSF1 modulate the nuclear export and mitochondrial localization of the lncRNA RMRP. Genes Dev. 2016, 30, 1224–1239. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, L.; Li, H.; Sun, T.; Wen, X.; Li, X.; Meng, Y.; Li, Y.; Liu, M.; Liu, S.; et al. Nuclear-Encoded lncRNA MALAT1 Epigenetically Controls Metabolic Reprogramming in HCC Cells through the Mitophagy Pathway. Mol. Ther. Nucleic Acids 2021, 23, 264. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution-trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2018, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Hirsch, A.G.; Bender, L.; Lingner, T.; Salinas, G.; Correspondence, H.K.; Krebber, H. Nuclear Pre-snRNA Export Is an Essential Quality Assurance Mechanism for Functional Spliceosomes Article Nuclear Pre-snRNA Export Is an Essential Quality Assurance Mechanism for Functional Spliceosomes. Cell Rep. 2019, 27, 3199–3214.e3. [Google Scholar] [CrossRef] [PubMed]
- Reichow, S.L.; Hamma, T.; Ferré-D’Amaré, A.R.; Varani, G. The structure and function of small nucleolar ribonucleoproteins. Nucleic Acids Res. 2007, 35, 1452–1464. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, P.; Wu, Q.; Fang, H.; Wang, Y.; Xiao, Y.; Cong, M.; Wang, T.; He, Y.; Ma, C.; et al. Long non-coding RNA NR2F1-AS1 induces breast cancer lung metastatic dormancy by regulating NR2F1 and ΔNp63. Nat. Commun. 2021, 12, 5232. [Google Scholar] [CrossRef]
- Calle, A.S.; Yamamoto, T.; Kawamura, Y.; Hironaka-Mitsuhashi, A.; Ono, M.; Tsuda, H.; Shimomura, A.; Tamura, K.; Takeshita, F.; Ochiya, T.; et al. Long non-coding NR2F1-AS1 is associated with tumor recurrence in estrogen receptor-positive breast cancers. Mol. Oncol. 2020, 14, 2271. [Google Scholar] [CrossRef]
- Kim, J.; Piao, H.L.; Kim, B.J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long non-coding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705. [Google Scholar] [CrossRef]
- Yin, H.; Hu, J.; Ye, Z.; Chen, S.; Chen, Y. Serum long non-coding RNA NNT-AS1 protected by exosome is a potential biomarker and functions as an oncogene via the miR-496/RAP2C axis in colorectal cancer. Mol. Med. Rep. 2021, 24, 585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, H.; Li, J.; Ma, X.; He, Z.; Liu, C.; Gao, C.; Li, H.; Wang, X.; Wu, J. 6-lncRNA Assessment Model for Monitoring and Prognosis of HER2-Positive Breast Cancer: Based on Transcriptome Data. Pathol. Oncol. Res. 2021, 27, 609083. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Miao, Z.; Wang, K.; Lv, Y.; Qiu, L.; Guo, L. Expression levels and clinical values of miR-92b-3p in breast cancer. World J. Surg. Oncol. 2021, 19, 239. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Lou, X.; Meng, N.; Li, Z.; Teng, Y.; Zou, Y.; Wang, F. Peripheral Blood-Based DNA Methylation of Long Non-Coding RNA H19 and Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promoters are Potential Non-Invasive Biomarkers for Gastric Cancer Detection. Cancer Control 2021, 28, 10732748211043667. [Google Scholar] [CrossRef] [PubMed]
- Said, M.N.; Muawia, S.; Helal, A.; Fawzy, A.; Allam, R.M.; Shafik, N.F. Regulation of CDK inhibitor p27 by microRNA 222 in breast cancer patients. Exp. Mol. Pathol. 2021, 123, 104718. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Cui, K.; Weng, L.; Yu, P.; Du, Y.; Zhang, T.; Liu, H.; Li, B.; Ma, W. A panel of 8-lncRNA predicts prognosis of breast cancer patients and migration of breast cancer cells. PLoS ONE 2021, 16, e0249174. [Google Scholar] [CrossRef]
- Wu, Q.; Li, Q.; Zhu, W.; Zhang, X.; Li, H. Identification of autophagy-related long non-coding RNA prognostic signature for breast cancer. J. Cell. Mol. Med. 2021, 25, 4088–4098. [Google Scholar] [CrossRef]
- Su, X.; Yu, Z.; Zhang, Y.; Chen, J.; Wei, L.; Sun, L. Construction and Analysis of the Dysregulated ceRNA Network and Identification of Risk Long Noncoding RNAs in Breast Cancer. Front. Genet. 2021, 12, 806. [Google Scholar] [CrossRef]
- Sheng, X.; Dai, H.; Du, Y.; Peng, J.; Sha, R.; Yang, F.; Zhou, L.; Lin, Y.; Xu, S.; Wu, Y.; et al. LncRNA CARMN overexpression promotes prognosis and chemosensitivity of triple negative breast cancer via acting as miR143-3p host gene and inhibiting DNA replication. J. Exp. Clin. Cancer Res. 2021, 40, 205. [Google Scholar] [CrossRef]
- Zhao, T.; Meng, W.; Chin, Y.; Gao, L.; Yang, X.; Sun, S.; Pan, X.; He, L. Identification of miR-25-3p as a tumor biomarker: Regulation of cellular functions via TOB1 in breast cancer. Mol. Med. Rep. 2021, 23, 406. [Google Scholar] [CrossRef]
- Kärkkäinen, E.; Heikkinen, S.; Tengström, M.; Kosma, V.M.; Mannermaa, A.; Hartikainen, J.M. The debatable presence of PIWI-interacting RNAs in invasive breast cancer. Cancer Med. 2021, 10, 3593–3603. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, X.; Zheng, Y.; Chen, W.; Zheng, Y.; Li, G.; Lou, W.; Wang, X. Construction of an mRNA-miRNA-lncRNA network prognostic for triple-negative breast cancer. Aging 2021, 13, 1153–1175. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Barwal, T.S.; Khandelwal, A.; Malhotra, A.; Rana, M.K.; Singh Rana, A.P.; Imyanitov, E.N.; Vasquez, K.M.; Jain, A. LncRNA ZFAS1 inhibits triple-negative breast cancer by targeting STAT3. Biochimie 2021, 182, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, R.; Tripathi, R.; Sridhar, T.S.; Korlimarla, A.; Choudhury, K.D.; Suryavanshi, M.; Mehta, A.; Doval, D.C. Prognostic role of microRNA 182 and microRNA 18a in locally advanced triple negative breast cancer. PLoS ONE 2020, 15, e0242190. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Li, C.; Kong, F.; Wu, Y.; Yuan, Q.; Hu, L. Prognostic and clinicopathological significance of MicroRNA-153 in human cancers: A meta-analysis. Medicine 2020, 99, e22833. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Wang, P.; Pan, B.; Nie, J.; Wang, S.; He, B. The diagnostic and prognostic values of microRNA-196a in cancer. Biosci. Rep. 2021, 41, BSR20203559. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zheng, S.; Peng, S.; Yu, Y.; Fang, J.; Tan, S.; Yao, F.; Guo, Z.; Shao, Y. Prognostic Value and Clinicopathological Features of MicroRNA-206 in Various Cancers: A Meta-Analysis. Biomed Res. Int. 2020, 2020, 2159704. [Google Scholar] [CrossRef]
- Gao, S.; Shi, P.; Tian, Z.; Yang, X.; Liu, N. Overexpression of miR-1225 promotes the progression of breast cancer, resulting in poor prognosis. Clin. Exp. Med. 2021, 21, 287–296. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Xiao, H.; Deng, X. Serum lncRNA TINCR Serve as a Novel Biomarker for Predicting the Prognosis in Triple-Negative Breast Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820965574. [Google Scholar] [CrossRef]
- Luo, Z.B.; Lai, G.E.; Jiang, T.; Cao, C.L.; Peng, T.; Liu, F.E. A Competing Endogenous RNA Network Reveals Novel lncRNA, miRNA and mRNA Biomarkers with Diagnostic and Prognostic Value for Early Breast Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820983293. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, L.; Yu, G.; Sun, Z.; Wang, T.; Tian, X.; Duan, X.; Zhang, C. Exosomal miR-1246 and miR-155 as predictive and prognostic biomarkers for trastuzumab-based therapy resistance in HER2-positive breast cancer. Cancer Chemother. Pharmacol. 2020, 86, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, X.; Xia, L.; Chen, Y.; Kuang, L.; Gu, X.; Li, T. A nine-lncRNA signature predicts distant relapse-free survival of HER2-negative breast cancer patients receiving taxane and anthracycline-based neoadjuvant chemotherapy. Biochem. Pharmacol. 2021, 189, 114285. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Yin, L.; Zhang, H.; Wei, B.; Chen, H.; Pu, T.; Yang, L.; Ye, F.; Zhang, Z.; Bu, H. High expression of microRNA20b is associated with malignant clinicopathological features and poor prognosis in breast phyllodes tumor. Int. J. Clin. Oncol. 2020, 25, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, J.; Ma, L.J.; Yang, H.B.; Jing, J.F.; Jia, M.M.; Zhang, X.J.; Guo, F.; Gao, J.N. Identification of serum exosomal miR-148a as a novel prognostic biomarker for breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7303–7309. [Google Scholar] [CrossRef]
- Han, S.; Li, P.; Wang, D.; Yan, H. Dysregulation of serum miR-1204 and its potential as a biomarker for the diagnosis and prognosis of breast cancer. Rev. Assoc. Med. Bras. 2020, 66, 732–736. [Google Scholar] [CrossRef]
- Zhang, S.; Qiu, D.; Xie, X.; Shen, Y. Clinicopathological and prognostic value of SNHG6 in cancers: A systematic review and a meta-analysis. BMC Cancer 2020, 20, 343. [Google Scholar] [CrossRef]
- Binabaj, M.M.; Bahrami, A.; Khazaei, M.; Avan, A.; Ferns, G.A.; Soleimanpour, S.; Ryzhikov, M.; Hassanian, S.M. The Prognostic Value of Small Noncoding microRNA-21 Expression in the Survival of Cancer Patients: A Meta-Analysis. Crit. Rev. Eukaryot. Gene Expr. 2020, 30, 207–221. [Google Scholar] [CrossRef]
- Zhang, X.; Zhuang, J.; Liu, L.; He, Z.; Liu, C.; Ma, X.; Li, J.; Ding, X.; Sun, C. Integrative transcriptome data mining for identification of core lncRNAs in breast cancer. PeerJ 2019, 7, e7821. [Google Scholar] [CrossRef]
- Feng, T.; Zhang, P.; Sun, Y.; Han, X.; Tong, J.; Hua, Z. Evaluation of the Role of hsa-mir-124 in Predicting Clinical Outcome in Breast Invasive Carcinoma Based on Bioinformatics Analysis. Biomed Res. Int. 2020, 2020, 1839205. [Google Scholar] [CrossRef]
- Li, W.J.; Xie, X.X.; Bai, J.; Wang, C.; Zhao, L.; Jiang, D.Q. Increased expression of miR-1179 inhibits breast cancer cell metastasis by modulating Notch signaling pathway and correlates with favorable prognosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8374–8382. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Z.; Fang, X.; Li, N.; Fang, J. Long noncoding RNA Breast cancer antiestrogen resistance 4 is associated with cancer progression and its significant prognostic value. J. Cell. Physiol. 2019, 234, 12956–12963. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Deng, K.; Liu, X.; Dai, M.; Chen, X.; Chen, J.; Chen, J.; Huang, Y.; Dai, S.; Chen, J. Molecular mechanism and role of microRNA-93 in human cancers: A study based on bioinformatics analysis, meta-analysis, and quantitative polymerase chain reaction validation. J. Cell. Biochem. 2019, 120, 6370–6383. [Google Scholar] [CrossRef]
- Zhang, X.J.; Li, X.; Sun, Y.; Xu, M. A three-microRNA signature predicts clinical outcome in breast cancer patients. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6396–6402. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.T.; Feng, C.X.; Zhao, J.C.; He, R.Q.; Ma, J.; Zhong, J.C. Upregulated miR-203a-3p and its potential molecular mechanism in breast cancer: A study based on bioinformatics analyses and a comprehensive meta-analysis. Mol. Med. Rep. 2018, 18, 4994–5008. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Yan, L.; Qi, Q.; Peng, X.; Edge, S.B.; Young, J.; Yao, S.; Liu, S.; Otsuji, E.; Takabe, K. Novel MicroRNA-Based Risk Score Identified by Integrated Analyses to Predict Metastasis and Poor Prognosis in Breast Cancer. Ann. Surg. Oncol. 2018, 25, 4037–4046. [Google Scholar] [CrossRef]
- Cui, X.; Yi, Q.; Jing, X.; Huang, Y.; Tian, J.; Long, C.; Xiang, Z.; Liu, J.; Zhang, C.; Tan, B.; et al. Mining Prognostic Significance of MEG3 in Human Breast Cancer Using Bioinformatics Analysis. Cell. Physiol. Biochem. 2018, 50, 41–51. [Google Scholar] [CrossRef]
- Fan, C.N.; Ma, L.; Liu, N. Systematic analysis of lncRNA-miRNA-mRNA competing endogenous RNA network identifies four-lncRNA signature as a prognostic biomarker for breast cancer. J. Transl. Med. 2018, 16, 264. [Google Scholar] [CrossRef]
- Fan, C.N.; Ma, L.; Liu, N. Comprehensive analysis of novel three-long noncoding RNA signatures as a diagnostic and prognostic biomarkers of human triple-negative breast cancer. J. Cell. Biochem. 2019, 120, 3185–3196. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, D.M.; Deng, S.H.; Han, R.; Liu, T.; Li, J.; Xu, Y. Integrated Analysis Reveals That Long Non-Coding RNA TUBA4B Can Be Used as a Prognostic Biomarker in Various Cancers. Cell. Physiol. Biochem. 2018, 49, 530–544. [Google Scholar] [CrossRef]
- Tang, J.; Ma, W.; Zeng, Q.; Tan, J.; Cao, K.; Luo, L. Identification of miRNA-Based Signature as a Novel Potential Prognostic Biomarker in Patients with Breast Cancer. Dis. Markers 2019, 2019, 3815952. [Google Scholar] [CrossRef]
- Xu, L.; Xu, K.; Xiang, L.; Yan, J. Circular RNA OMA1 regulates the progression of breast cancer via modulation of the miR-1276/SIRT4 axis. Mol. Med. Rep. 2021, 24, 728. [Google Scholar] [CrossRef]
- Dwedar, F.I.; Shams-Eldin, R.S.; Nayer Mohamed, S.; Mohammed, A.F.; Gomaa, S.H. Potential value of circulatory microRNA10b gene expression and its target E-cadherin as a prognostic and metastatic prediction marker for breast cancer. J. Clin. Lab. Anal. 2021, 35, e23887. [Google Scholar] [CrossRef] [PubMed]
- Zheng, A.; Zhang, L.; Song, X.; Jin, F. Clinical significance of SPRY4-IT1 in efficacy and survival prediction in breast cancer patients undergoing neoadjuvant chemotherapy. Histol. Histopathol. 2020, 35, 361–370. [Google Scholar] [CrossRef]
- Lasham, A.; Fitzgerald, S.J.; Knowlton, N.; Robb, T.; Tsai, P.; Black, M.A.; Williams, L.; Mehta, S.Y.; Harris, G.; Shelling, A.N.; et al. A Predictor of Early Disease Recurrence in Patients with Breast Cancer Using a Cell-free RNA and Protein Liquid Biopsy. Clin. Breast Cancer 2020, 20, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Nama, S.; Muhuri, M.; Di Pascale, F.; Quah, S.; Aswad, L.; Fullwood, M.; Sampath, P. MicroRNA-138 is a Prognostic Biomarker for Triple-Negative Breast Cancer and Promotes Tumorigenesis via TUSC2 repression. Sci. Rep. 2019, 9, 12718. [Google Scholar] [CrossRef] [PubMed]
- Zou, B.; Wang, D.; Xu, K.; Liu, J.L.; Yuan, D.Y.; Meng, Z.; Zhang, B. Prognostic value of long non-coding RNA plasmacytoma variant translocation1 in human solid tumors: A meta-analysis. Medicine 2019, 98, e16087. [Google Scholar] [CrossRef]
- Jia, X.; Wei, L.; Zhang, Z. NEAT1 Overexpression Indicates a Poor Prognosis and Induces Chemotherapy Resistance via the miR-491-5p/SOX3 Signaling Pathway in Ovarian Cancer. Front. Genet. 2021, 12, 616220. [Google Scholar] [CrossRef]
- Lukianova, N.Y.; Borikun, T.V.; Chekhun, V.F. Tumor microenvironment-derived miRNAs as prognostic markers of breast cancer. Exp. Oncol. 2019, 41, 242–247. [Google Scholar] [CrossRef]
- Tsiakou, A.; Zagouri, F.; Zografos, E.; Samelis, G.; Gazouli, M.; Kalapanida, D.; Giannos, A.; Marinopoulos, S.; Dimitrakakis, K.; Lazaris, C.A.; et al. Prognostic significance of miR-34 rs4938723 T>C polymorphism in triple negative breast cancer patients. Clin. Biochem. 2019, 68, 9–14. [Google Scholar] [CrossRef]
- Pan, C.; Luo, J.; Zhang, J. Computational Identification of RNA-Seq Based miRNA-Mediated Prognostic Modules in Cancer. IEEE J. Biomed. Health Inform. 2020, 24, 626–633. [Google Scholar] [CrossRef]
- Fan, C.; Liu, N. Identification of dysregulated microRNAs associated with diagnosis and prognosis in triple-negative breast cancer: An In Silico study. Oncol. Rep. 2019, 41, 3313–3324. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liao, C.; Zhong, Q.; Dong, H.; Zhang, T.; Jin, R. CeNETs analysis reveals the prognostic value of a signature integration from five lncRNAs in breast cancer. J. Cell. Biochem. 2019, 120, 13509–13519. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Soliman, S.E.; Habib, M.S.E.; Gohar, S.F.; Abo-Zeid, G.S. Potential value of circulatory microRNA122 gene expression as a prognostic and metastatic prediction marker for breast cancer. Mol. Biol. Rep. 2019, 46, 2809–2818. [Google Scholar] [CrossRef] [PubMed]
- Arabkari, V.; Clancy, E.; Dwyer, R.M.; Kerin, M.J.; Kalinina, O.; Holian, E.; Newell, J.; Smith, T.J. Relative and Absolute Expression Analysis of MicroRNAs Associated with Luminal A Breast Cancer-A Comparison. Pathol. Oncol. Res. 2020, 26, 833–844. [Google Scholar] [CrossRef]
- Meng, Y.; Liu, Y.L.; Li, K.; Fu, T. Prognostic value of long non-coding RNA breast cancer anti-estrogen resistance 4 in human cancers: A meta-analysis. Medicine 2019, 98, e15793. [Google Scholar] [CrossRef]
- Salavaty, A.; Motlagh, F.M.; Barabadi, M.; Cheshomi, H.; Esmatabadi, M.J.D.; Shahmoradi, M.; Soleimanpour-Lichaei, H.R. Potential role of RAB6C-AS1 long noncoding RNA in different cancers. J. Cell. Physiol. 2018, 234, 891–903. [Google Scholar] [CrossRef]
- Papadopoulos, E.I.; Papachristopoulou, G.; Ardavanis, A.; Scorilas, A. A comprehensive clinicopathological evaluation of the differential expression of microRNA-331 in breast tumors and its diagnostic significance. Clin. Biochem. 2018, 60, 24–32. [Google Scholar] [CrossRef]
- Kim, C.; Go, E.J.; Kim, A. Recurrence prediction using microRNA expression in hormone receptor positive breast cancer during tamoxifen treatment. Biomarkers 2018, 23, 804–811. [Google Scholar] [CrossRef]
- Jiang, M.; Xiao, Y.; Liu, D.; Luo, N.; Gao, Q.; Guan, Y. Overexpression of long noncoding RNA LINC01296 indicates an unfavorable prognosis and promotes tumorigenesis in breast cancer. Gene 2018, 675, 217–224. [Google Scholar] [CrossRef]
- Liu, W.; Xu, Y.; Guan, H.; Meng, H. Clinical potential of miR-940 as a diagnostic and prognostic biomarker in breast cancer patients. Cancer Biomark. 2018, 22, 487–493. [Google Scholar] [CrossRef]
- Lu, R.; Zhang, J.; Zhang, W.; Huang, Y.; Wang, N.; Zhang, Q.; Qu, S. Circulating HOTAIR expression predicts the clinical response to neoadjuvant chemotherapy in patients with breast cancer. Cancer Biomark. 2018, 22, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, S.H.; Kong, P.; Zhang, L.Y.; Zhang, L.L.; Zhang, N.Q.; Gu, H. Increased expression of miR-330-3p: A novel independent indicator of poor prognosis in human breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1726–1730. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Pongor, L.; Bottai, G.; Li, X.; Budczies, J.; Szabó, A.; Hatzis, C.; Pusztai, L.; Santarpia, L. An integrative bioinformatics approach reveals coding and non-coding gene variants associated with gene expression profiles and outcome in breast cancer molecular subtypes. Br. J. Cancer 2018, 118, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.W.; Zhu, W.J.; Li, Y.; Liu, L.; Yin, G.; Gao, P. A 4-microRNA signature predicts lymph node metastasis and prognosis in breast cancer. Hum. Pathol. 2018, 76, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Abak, A.; Amini, S.; Estiar, M.; Montazeri, V.; Sakhinia, E.; Abhari, A. Analysis of miRNA-221 Expression Level in Tumors and Marginal Biopsies from Patients with Breast Cancer (Cross-Sectional Observational Study). Clin. Lab. 2018, 64, 169–175. [Google Scholar] [CrossRef]
- Wang, K.; Li, J.; Xiong, Y.F.; Zeng, Z.; Zhang, X.; Li, H.Y. A Potential Prognostic Long Noncoding RNA Signature to Predict Recurrence among ER-positive Breast Cancer Patients Treated with Tamoxifen. Sci. Rep. 2018, 8, 3179. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Liu, D.J.; Yuan, R.B.; Zhang, D.H.; Li, S.R.; Zhang, S.H.; Zhang, L.Y. Low expression of miR-597 is correlated with tumor stage and poor outcome in breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 456–460. [Google Scholar] [CrossRef]
- Papachristopoulou, G.; Papadopoulos, E.I.; Nonni, A.; Rassidakis, G.Z.; Scorilas, A. Expression Analysis of miR-29b in Malignant and Benign Breast Tumors: A Promising Prognostic Biomarker for Invasive Ductal Carcinoma With a Possible Histotype-Related Expression Status. Clin. Breast Cancer 2018, 18, 305–312.e3. [Google Scholar] [CrossRef]
- Zheng, J.Z.; Huang, Y.N.; Yao, L.; Liu, Y.R.; Liu, S.; Hu, X.; Liu, Z.B.; Shao, Z.M. Elevated miR-301a expression indicates a poor prognosis for breast cancer patients. Sci. Rep. 2018, 8, 2225. [Google Scholar] [CrossRef]
- Zidan, H.E.; Karam, R.A.; El-Seifi, O.S.; Abd Elrahman, T.M. Circulating long non-coding RNA MALAT1 expression as molecular biomarker in Egyptian patients with breast cancer. Cancer Genet. 2018, 220, 32–37. [Google Scholar] [CrossRef]
- Liu, L.; Chi, Y.; Chen, J.; Xue, J.; Deng, L.; Huang, N.; Shao, J.; Wu, J. Long non-coding RNA00544 serves as a potential novel predictive and prognostic marker for HR+ HER2− subtype breast cancer. Sci. Rep. 2017, 7, 12382. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.H.; Kim, H.J.; Gwak, J.M.; Chung, Y.R.; Park, S.Y. Prognostic value of microRNA-9 and microRNA-155 expression in triple-negative breast cancer. Hum. Pathol. 2017, 68, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, C.; Tang, H.; Zhang, C.; Tang, J.; Li, X.; Chen, B.; Xie, X. miR-629-3p may serve as a novel biomarker and potential therapeutic target for lung metastases of triple-negative breast cancer. Breast Cancer Res. 2017, 19, 72. [Google Scholar] [CrossRef]
- Li, C.Y.; Xiong, D.D.; Huang, C.Q.; He, R.Q.; Liang, H.W.; Pan, D.H.; Wang, H.L.; Wang, Y.W.; Zhu, H.W.; Chen, G. Clinical Value of miR-101-3p and Biological Analysis of Its Prospective Targets in Breast Cancer: A Study Based on the Cancer Genome Atlas (TCGA) and Bioinformatics. Med. Sci. Monit. 2017, 23, 1857. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Slater, C.M.; Zhou, Y.; Devarajan, K.; Ruth, K.J.; Li, Y.; Cai, K.Q.; Daly, M.; Chen, X. LincIN, a novel NF90-binding long non-coding RNA, is overexpressed in advanced breast tumors and involved in metastasis. Breast Cancer Res. 2017, 19, 62. [Google Scholar] [CrossRef]
- Nie, Z.L.; Wang, Y.S.; Mei, Y.P.; Lin, X.; Zhang, G.X.; Sun, H.L.; Wang, Y.L.; Xia, Y.X.; Wang, S.K. Prognostic significance of long noncoding RNA Z38 as a candidate biomarker in breast cancer. J. Clin. Lab. Anal. 2018, 32, e22193. [Google Scholar] [CrossRef]
- Raychaudhuri, M.; Bronger, H.; Buchner, T.; Kiechle, M.; Weichert, W.; Avril, S. MicroRNAs miR-7 and miR-340 predict response to neoadjuvant chemotherapy in breast cancer. Breast Cancer Res. Treat. 2017, 162, 511–521. [Google Scholar] [CrossRef]
- Wu, A.; Chen, Y.; Liu, Y.; Lai, Y.; Liu, D. miR-199b-5p inhibits triple negative breast cancer cell proliferation, migration and invasion by targeting DDR1. Oncol. Lett. 2018, 16, 4889. [Google Scholar] [CrossRef]
- Huang, N.S.; Chi, Y.Y.; Xue, J.Y.; Liu, M.Y.; Huang, S.; Mo, M.; Zhou, S.L.; Wu, J. Long non-coding RNA metastasis associated in lung adenocarcinoma transcript 1 (MALAT1) interacts with estrogen receptor and predicted poor survival in breast cancer. Oncotarget 2016, 7, 37957. [Google Scholar] [CrossRef]
- Zelli, V.; Compagnoni, C.; Capelli, R.; Cannita, K.; Sidoni, T.; Ficorella, C.; Capalbo, C.; Zazzeroni, F.; Tessitore, A.; Alesse, E. Circulating MicroRNAs as Prognostic and Therapeutic Biomarkers in Breast Cancer Molecular Subtypes. J. Pers. Med. 2020, 10, 98. [Google Scholar] [CrossRef]
- Meseure, D.; Vacher, S.; Lallemand, F.; Alsibai, K.D.; Hatem, R.; Chemlali, W.; Nicolas, A.; De Koning, L.; Pasmant, E.; Callens, C.; et al. Prognostic value of a newly identified MALAT1 alternatively spliced transcript in breast cancer. Br. J. Cancer 2016, 114, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Zehentmayr, F.; Hauser-Kronberger, C.; Zellinger, B.; Hlubek, F.; Schuster, C.; Bodenhofer, U.; Fastner, G.; Deutschmann, H.; Steininger, P.; Reitsamer, R.; et al. Hsa-miR-375 is a predictor of local control in early stage breast cancer. Clin. Epigenetics 2016, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Study on the Relationship between miR-520g and the Development of Breast Cancer. Available online: https://www.europeanreview.org/article/10343 (accessed on 14 February 2022).
- Madhavan, D.; Peng, C.; Wallwiener, M.; Zucknick, M.; Nees, J.; Schott, S.; Rudolph, A.; Riethdorf, S.; Trumpp, A.; Pantel, K.; et al. Circulating miRNAs with prognostic value in metastatic breast cancer and for early detection of metastasis. Carcinogenesis 2016, 37, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Liu, T.; Li, Y.; Li, S. Overexpression of long non-coding RNA CCAT1 is a novel biomarker of poor prognosis in patients with breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 9440. [Google Scholar]
- Dong, L.L.; Chen, L.M.; Wang, W.M.; Zhang, L.M. Decreased expression of microRNA-124 is an independent unfavorable prognostic factor for patients with breast cancer. Diagn. Pathol. 2015, 10, 45. [Google Scholar] [CrossRef][Green Version]
- Sørensen, K.P.; Thomassen, M.; Tan, Q.; Bak, M.; Cold, S.; Burton, M.; Larsen, M.J.; Kruse, T.A. Long non-coding RNA expression profiles predict metastasis in lymph node-negative breast cancer independently of traditional prognostic markers. Breast Cancer Res. 2015, 17, 55. [Google Scholar] [CrossRef]
- Gökmen-Polar, Y.; Vladislav, I.T.; Neelamraju, Y.; Janga, S.C.; Badve, S. Prognostic impact of HOTAIR expression is restricted to ER-negative breast cancers. Sci. Rep. 2015, 5, 8765. [Google Scholar] [CrossRef]
- Wang, G.; Wang, L.; Sun, S.; Wu, J.; Wang, Q. Quantitative Measurement of Serum MicroRNA-21 Expression in Relation to Breast Cancer Metastasis in Chinese Females. Ann. Lab. Med. 2015, 35, 226. [Google Scholar] [CrossRef]
- Zheng, R.; Pan, L.; Gao, J.; Ye, X.; Chen, L.; Zhang, X.; Tang, W.; Zheng, W. Prognostic value of miR-106b expression in breast cancer patients. J. Surg. Res. 2015, 195, 158–165. [Google Scholar] [CrossRef]
- Agarwal, S.; Hanna, J.; Sherman, M.E.; Figueroa, J.; Rimm, D.L. Quantitative assessment of miR34a as an independent prognostic marker in breast cancer. Br. J. Cancer 2014, 112, 61–68. [Google Scholar] [CrossRef][Green Version]
- Müller, V.; Gade, S.; Steinbach, B.; Loibl, S.; Von Minckwitz, G.; Untch, M.; Schwedler, K.; Lübbe, K.; Schem, C.; Fasching, P.A.; et al. Changes in serum levels of miR-21, miR-210, and miR-373 in HER2-positive breast cancer patients undergoing neoadjuvant therapy: A translational research project within the Geparquinto trial. Breast Cancer Res. Treat. 2014, 147, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Sun, Q.; Liang, Z.; Cui, X.; Ren, X.; Chen, H.; Zhang, X.; Zhou, Y. A Prognostic Model of Triple-Negative Breast Cancer Based on miR-27b-3p and Node Status. PLoS ONE 2014, 9, e100664. [Google Scholar] [CrossRef] [PubMed]
- Parrella, P.; Barbano, R.; Pasculli, B.; Fontana, A.; Copetti, M.; Valori, V.M.; Poeta, M.L.; Perrone, G.; Righi, D.; Castelvetere, M.; et al. Evaluation of microRNA-10b prognostic significance in a prospective cohort of breast cancer patients. Mol. Cancer 2014, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, C.; Rani, S.; Breslin, S.; Gogarty, M.; Ghobrial, I.M.; Crown, J.; O’Driscoll, L. MiR-630 targets IGF1R to regulate response to HER-targeting drugs and overall cancer cell progression in HER2 over-expressing breast cancer. Mol. Cancer 2014, 13, 71. [Google Scholar] [CrossRef]
- Pérez-Rivas, L.G.; Jerez, J.M.; Carmona, R.; Luque, V.; Vicioso, L.; Claros, M.G.; Viguera, E.; Pajares, B.; Sańchez, A.; Ribelles, N.; et al. A microRNA signature associated with early recurrence in breast cancer. PLoS ONE 2014, 9, e91884. [Google Scholar] [CrossRef]
- Gasparini, P.; Cascione, L.; Fassan, M.; Lovat, F.; Guler, G.; Balci, S.; Irkkan, C.; Morrison, C.; Croce, C.M.; Shapiro, C.L.; et al. microRNA expression profiling identifies a four microRNA signature as a novel diagnostic and prognostic biomarker in triple negative breast cancers. Oncotarget 2014, 5, 1174. [Google Scholar] [CrossRef]
- Bueno, R.C.; Canevari, R.A.; Villacis, R.A.R.; Domingues, M.A.C.; Caldeira, J.R.F.; Rocha, R.M.; Drigo, S.A.; Rogatto, S.R. ATM down-regulation is associated with poor prognosis in sporadic breast carcinomas. Ann. Oncol. 2014, 25, 69–75. [Google Scholar] [CrossRef]
- Sørensen, K.P.; Thomassen, M.; Tan, Q.; Bak, M.; Cold, S.; Burton, M.; Larsen, M.J.; Kruse, T.A. Long non-coding RNA HOTAIR is an independent prognostic marker of metastasis in estrogen receptor-positive primary breast cancer. Breast Cancer Res. Treat. 2013, 142, 529–536. [Google Scholar] [CrossRef]
- Redis, R.S.; Sieuwerts, A.M.; Look, M.P.; Tudoran, O.; Ivan, C.; Spizzo, R.; Zhang, X.; de Weerd, V.; Shimizu, M.; Ling, H.; et al. CCAT2, a novel long non-coding RNA in breast cancer: Expression study and clinical correlations. Oncotarget 2013, 4, 1748–1762. [Google Scholar] [CrossRef]
- Yan, M.; Xu, H.; Waddell, N.; Shield-Artin, K.; Haviv, I.; McKay, M.J.; Fox, S.B. Enhanced RAD21 cohesin expression confers poor prognosis in BRCA2 and BRCAX, but not BRCA1 familial breast cancers. Breast Cancer Res. 2012, 14, R69. [Google Scholar] [CrossRef]
- Le Quesne, J.; Jones, J.; Warren, J.; Dawson, S.J.; Raza Ali, H.; Bardwell, H.; Blows, F.; Pharoah, P.; Caldas, C. Biological and prognostic associations of miR-205 and let-7b in breast cancer revealed by in situ hybridization analysis of micro-RNA expression in arrays of archival tumour tissue. J. Pathol. 2012, 227, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Rothé, F.; Ignatiadis, M.; Chaboteaux, C.; Haibe-Kains, B.; Kheddoumi, N.; Majjaj, S.; Badran, B.; Fayyad-Kazan, H.; Desmedt, C.; Harris, A.L.; et al. Global microRNA expression profiling identifies MiR-210 associated with tumor proliferation, invasion and poor clinical outcome in breast cancer. PLoS ONE 2011, 6, e20980. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.A.M.; Slewa, A.; Gudlaugsson, E.; Jonsdottir, K.; Skaland, I.; Søiland, H.; Baak, J.P.A. Biologic profiling of lymph node negative breast cancers by means of microRNA expression. Mod. Pathol. 2010, 23, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Grelier, G.; Voirin, N.; Ay, A.S.; Cox, D.G.; Chabaud, S.; Treilleux, I.; Léon-Goddard, S.; Rimokh, R.; Mikaelian, I.; Venoux, C.; et al. Prognostic value of Dicer expression in human breast cancers and association with the mesenchymal phenotype. Br. J. Cancer 2009, 101, 673. [Google Scholar] [CrossRef]
- Liu, Y.R.; Jiang, Y.Z.; Xu, X.E.; Hu, X.; Yu, K.D.; Shao, Z.M. Comprehensive Transcriptome Profiling Reveals Multigene Signatures in Triple-Negative Breast Cancer. Clin. Cancer Res. 2016, 22, 1653–1662. [Google Scholar] [CrossRef]
- Xu, S.; Wang, P.; You, Z.; Meng, H.; Mu, G.; Bai, X.; Zhang, G.; Zhang, J.; Pang, D. The long non-coding RNA EPB41L4A-AS2 inhibits tumor proliferation and is associated with favorable prognoses in breast cancer and other solid tumors. Oncotarget 2016, 7, 20704. [Google Scholar] [CrossRef]
- Lánczky, A.; Nagy, Á.; Bottai, G.; Munkácsy, G.; Szabó, A.; Santarpia, L.; Győrffy, B. miRpower: A web-tool to validate survival-associated miRNAs utilizing expression data from 2178 breast cancer patients. Breast Cancer Res. Treat. 2016, 160, 439–446. [Google Scholar] [CrossRef]
- Hu, Y.; Qiu, Y.; Yagüe, E.; Ji, W.; Liu, J.; Zhang, J. miRNA-205 targets VEGFA and FGF2 and regulates resistance to chemotherapeutics in breast cancer. Cell Death Dis. 2016, 7, e2291. [Google Scholar] [CrossRef]
- Gong, C.; Tan, W.; Chen, K.; You, N.; Zhu, S.; Liang, G.; Xie, X.; Li, Q.; Zeng, Y.; Ouyang, N.; et al. Prognostic Value of a BCSC-associated MicroRNA Signature in Hormone Receptor-Positive HER2-Negative Breast Cancer. eBioMedicine 2016, 11, 199–209. [Google Scholar] [CrossRef]
- Zhou, M.; Zhong, L.; Xu, W.; Sun, Y.; Zhang, Z.; Zhao, H.; Yang, L.; Sun, J. Discovery of potential prognostic long non-coding RNA biomarkers for predicting the risk of tumor recurrence of breast cancer patients. Sci. Rep. 2016, 6, 31038. [Google Scholar] [CrossRef]
- Xu, L.; Fengji, L.; Changning, L.; Liangcai, Z.; Yinghui, L.; Yu, L.; Shanguang, C.; Jianghui, X. Comparison of the Prognostic Utility of the Diverse Molecular Data among lncRNA, DNA Methylation, microRNA, and mRNA across Five Human Cancers. PLoS ONE 2015, 10, e0142433. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, X.; Huang, Z.; Xu, L.; Zhu, W.; Liu, P. An ER-associated miRNA signature predicts prognosis in ER-positive breast cancer. J. Exp. Clin. Cancer Res. 2014, 33, 94. [Google Scholar] [CrossRef] [PubMed]
- Dedes, K.J.; Natrajan, R.; Lambros, M.B.; Geyer, F.C.; Lopez-Garcia, M.A.; Savage, K.; Jones, R.L.; Reis-Filho, J.S. Down-regulation of the miRNA master regulators Drosha and Dicer is associated with specific subgroups of breast cancer. Eur. J. Cancer 2011, 47, 138–150. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference Number | Non-Coding RNA/s Investigated | N° of Patients vs. Healthy Controls | Technique (TCGA, qRT-PCR or ddPCR) | Main Observation |
|---|---|---|---|---|
| [56] | 6 lncRNAs (AC046168.1, AC010595.1, AC069277.1, AP000904.1 MIR762MG and LINC00528) | These RNA data were derived from 113 HER2-positive breast cancer tissues and 105 tumor-adjacent normal breast tissues. | TCGA | The 6-lncRNA model had a good predictive power for OS and 3-year survival in HER2+ BCs |
| [57] | miR-92b | 112 female BC patients and 108 healthy women. | qRT-PCR | Higher expression level of miR-92b-3p. AUC of 0.88 correlated with degree of differentiation, tumor size and TNM staging, lymph metastasis. miR-92b significantly positively correlated with the expression of carbohydrate antigen 125 (CA125) |
| [58] | RP11-1024P17.1, RP11-890B15.3, MFI2-AS1 and RP11-180N14.1 | Patients from TCGA and GEO databases. | TCGA and GEO databases | Useful to stratify patients into high and low risk groups and as prognostic biomarkers.RP11-890B15.3, RP11-180N14.1 and RP11-1024P17.1 could regulate more mRNAs by targeting various miRNAs. The MF12-AS1 regulated three mRNAs by sponging miR-3150a-3p. |
| [59] | lncRNA MIAT | 1057 BC and 103 normal specimens | First: TCGA Then: qRT-PCR | Its expression in serum positively correlated with TNM stage and lymph node metastasis |
| [60] | miR-222 | 110 patients | qRT-PCR | Capacity to inhibit tumor suppressor CDK inhibitor p27 in BC. miR-222 was expressed at significantly higher levels in BC. Serum p27 and miR-222 could help differentiate between BC and controls |
| [61] | 8-IncRNA | 808 BC patients | First: TCGA Then: qRT-PCR | Potential prognostic biomarker for BC |
| [62] | LINC01871, MAPT-AS1, AL122010.1, AC090912.1, AC061992.1 | 1108 BC patients | TCGA | Their model offered an independent prognostic value, with a risk score significantly related to the TNM stage, PR, ER and HER2 status in BC patients |
| [63] | LINC01977, AP000851.1, MAFG-D, SIAH2-AS1 | 1222 BC patients | TCGA | Synergistically exerted functions related to cell cycle and DNA separation, DNA replication, and an independent prognostic marker for BC |
| [64] | IncRNA CARMN | 250 BC patients | qRT-PCR | CAMN could predict both better prognosis and higher response rate of cisplatin-based neoadjuvant chemotherapy in BC patients and inhibit DNA replication. |
| [65] | miR-25-3p | 25 BC patients | qRT-PCR | Patients with low expression levels of serum miR-25-3p had a higher survival compared to those with higher miR-25-3p expression. miR-25-3p could be a good biomarker for BC |
| [66] | PIWI-interacting RNAs (piRNAs): DQ570994, DQ571955, and DQ596932 | 227 fresh-frozen BC samples. | RNA sequencing | piRNSAs were upregulated in grade III tumors and DQ696932 was upregulated in estrogen receptor negative tumors only. DQ571955 showed shorter relapse-free survival and poorer BC-specific survival. DQ571955 can be a predictive biomarker for radiotherapy response in ER+ BCs. DQ570994 can be a predictive marker for tamoxifen and chemotherapy response |
| [67] | NEAT1 and MAL2 | 63 patients SCLC | GEO2R tool | Able to differentiate good prognosis vs. bad regression free survivals |
| [68] | ZFAS1 | 40 TNBC patients compared to 40 healthy individuals | qRT-PCR | ZFAS1 promoted proliferation of human breast cancer cell line MDA-MB-231 TNBC cells through the inhibition of the cyclin-dependent kinase (CDK) inhibitors p21 and p27. ZFAS1 could be a diagnostic and prognostic marker for TNBC that could be also used for therapy. |
| [69] | miR-182 and miR-18 | 50 TNBC patients | qRT-PCR | A significant correlation was observed with clinical nodal status, T-category, clinical response, pathological response with miR-18 and miR-182 |
| [70] | miR153 miR-196a | Meta-analysis collecting data from 933 patients from 11 articles. | Meta-analysis | Low miR-153 expression significantly correlated with poor OS miR-153 could be a very effective biomarker for tumor prognosis especially in BC and digestive tumors. |
| [71] | miR-196a | 17 articles were included | Meta-analysis | Tumor tissue or blood-derived miR-196a could be used a prognostic and diagnostic biomarker for cancers such as BC |
| [72] | miR-206 | 2095 patients | qRT-PCR | Potential prognostic biomarker. The pooled HR showed that low miR-206 expression was significantly associated with unfavorable OS Moreover, the expression of miR-206 predicted significantly negative association with tumor stage distant metastasis, lymph node status and invasion depth |
| [73] | miR-1225 | 120 BC patients | qRT-PCR | Significantly upregulated in BC and associated with TNM stage of BC. Overexpression of miR-1225 could be used as a biomarkers since it correlated with a poor prognosis of patients and promoted the progression of BC by targeting JAK1 |
| [74] | TINCR | 72 TNBC patients, 105 non-TNBC patients, 60 benign BC, and 86 heathy subjects | qRT-PCR | The lncRNA TINCR level was significantly increased in BC patients, particularly in TNBC. The clinicopathological features and clinical outcomes of TNBC were worse in patients with high circulating lncRNA TINCR |
| [75] | ADAMTS9-AS1, CDKN2B-AS1, IL-6, MMP11, has-miR-145-5p ahashsa-miR-182-5p | 787 early BC patients and 78 normal BC individuals. | qRT-PCR | AUC were 0.947, 0.862, 0.842, 0.993, 0.960, and 0.944, and the specificity and sensitivity 83.4% and 95.6%, 72.2% and 90.3%, 80.1%, and 74.3%, 96.2% and 96.5%, 90.1%, and 92.3%, and 88.7%, and 90.4%, respectively |
| [76] | TPT-AS1 | 316 BC patients | First: TCGA and GEOSecond: qRT-PCR | The low expression of TPT1-AS1 was correlated with lymph node metastasis, TNM stage, HER-2- status, shorter OS. TPT1-AS1 was an independent prognostic factor for BC patients |
| [77] | miR-1246 and miR-155 | 107 early-stage and 68 metastatic BC patients treated with trastuzumab-based chemotherapy | Meta-analysis | Both miRNAs were not associated with OS. The study showed that miR-146 and miR-155 could distinguish trastuzumab-resistant from sensitive patients |
| [78] | 9-IncRNA | 10,213 BC patients | Meta-analysis | The 9-lncRNA signature was a robust and effective model for the prediction of DRFS of patients with HER2- BC |
| [79] | miR-20b | 123 BC patients | qRT-PCR | Expression of miR-20b increased with increase in tumor grade and correlated with stromal overgrowth, high stromal atypia and cellularity, infiltrative tumor margin, tumor grade, high mitotic activity, local recurrence and metastasis, and shorter DFS miR-20b could be used as a prognostic biomarker in BC |
| [80] | miR-148a | 125 patients with BC and 50 patients with benign breast tumors | qRT-PCR | miR-148a is significantly reduced in BC patients and down-regulation of serum exosomal miR-148a was closely associated with unfavorable clinical outcome of BC. |
| [81] | miR-1204 | 144 BC patients and 38 healthy volunteers | qRT-PCR | The study suggested that miR-1204 could be used a prognostic and diagnostic biomarker for BC7 |
| [82] | SNHG6 | 914 patients from 13 studies | Meta-analysis | Expression could predict unfavorable OS. elevated level of SNHG6 was positively associated with tumor invasion depth, DM LNM and advanced TNM stage |
| [83] | miR-21 | 10,213 cancer patients | Meta-analysis | miR-21 correlated with shorter OS in breast cancer patients and could be a clinically useful biomarker for cancer progression. |
| [84] | Seven core lncRNAs. LINC00478, AL035610.1, AC005550.4, MIR143HG, MIR497HG, PGM5-AS1, RP11-175K6.1 | 837 BC | TCGA | Good single-factor diagnostic value for BC. Moreover, AC005550.2 had a prognostic value for BC. AC005550.4 and MIR497HG were able to distinguish better BC patients in early stage from patients at advanced stages. Overall, the 7-lncRNA could have a prognostic value in BC |
| [85] | Has-mir-124 | 742 BC patients | TCGA | OS of patients with high expression levels of has-mir-124-1 and has-mir-124-2 was better than that of patients with low expression levels of has-mir-124-1 and has-mir-124-2. Overall, has-mir-124 was associated with worse clinical outcomes and could be used as a biomarker for BRCA |
| [86] | miR-1179 | 164 BC patients | qRT-PCR | miR-1179 was frequently down-regulated in BC tissues and cell lines. low miR-1179 levels correlated with advanced clinical stage, shorter OS and lymph node metastasis Overall, miR-1179 could serve as a new prognostic biomarker or actionable target for new therapies |
| [87] | lncRNA in BC antiestrogen resistance 4 (BCAR4) | 1293 patients from 9 studies | Meta-analysis | BCAR4 expression was significantly correlated with poor, and high expression levels of BCAR4 correlated with worse clinical stage, distant metastases, lymph node metastasis. |
| [88] | miR-93 | Meta-analysis | qRT-PCR | The AUC for overall sensitivity and specificity were 0.76, 0.82, and 0.85), suggesting that miR-93 is a good prognostic biomarker. |
| [89] | 3-miRNA signature (miR-133a-2, miR-204, and miR-301b). | 1103 BC patients vs. 104 healthy samples | TCGA | The 3-miRNA signature could be a potential biomarker for BRCA |
| [90] | miR-2031-3p | 1077 BC tissues compared to 104 adjacent BC samples | TCGA | miR-2031-3p did not have prognostic value in BC. Overall, the study suggests that miR-2031-3p could enhance tumorigenesis in BC, but has not prognostic utility. |
| [91] | Three miRNAs risk score (miR-19a, miR-93, and miR-106a) | 1051 BC patients | TCGA | The miRNA-based risk score could predict worse survival and bone recurrence in BC patients |
| [92] | lncRNA maternally expressed gene 3 (MEG3) | The MEG3 was down-regulated in BC than in normal tissues. ER and progesterone receptor (PR) status positively correlated with MEG3 expression. MEG3 positively correlated with heparin sulfate proteoglycan 2 (HSPG2) expression and could be a good predictor of prognosis in BC with HSPG2. | ||
| [93] | 2150 DEmRNAs, 1061 DElncRNAs, and 82 DEmiRNAs | 1103 BC vs. 104 adjacent normal breast tissues | TCGA | The 4-lncRNA signature could independently predict OS in BC patients. The AUC of the 4-lncRNA signature associated with 3-year of survival was 0.696. Overall, the 4-lncRNA is a good prognostic tool for BC. |
| [94] | AC091043.1, AP000924.1, and FOXCUT.AC010343.3, AL354793.1, and FGF10-AS1 | 1097 BC samples | TCGA | AC091043.1, AP000924.1, and FOXCUT, may have a strong diagnostic value for the prediction of TNBC in both training and validation sets (AUC > 0.85).The signature could be efficient in predicting diagnosis and prognosis of TNBC |
| [95] | TUBA4B | 94 BC patients vs. 86 normal tissues | Meta-analysis | TUBA4B is significantly correlated with OS DFS and recurrence-free survival. Overall, the study suggested that low levels of TUBA4B are significantly associated with short OS, DFS, and RFS in cancers and that TUBA4B could therefore be a BC biomarker |
| [96] | hsa-let-7b, hsa-mir-9-3, hsa-mir-22, hsa-mir-30a, hsa-mir-31, hsa-mir-101-2, hsa-mir- 135a-2, hsa-mir-320b-1, hsa-mir-493, hsa-mir-556, hsa-mir-652, hsa-mir-874, hsa-mir-3130-1, hsa-mir-3678, hsa-mir-4662a, hsa-mir-4772 and hsa-mir-6733. On the other hand sa-mir-130a, hsa-mir-204, hsa-mir-217, hsa-mir-223, hsa-mir-24-2, hsa-mir-29b- 1, hsa-mir-363, hsa-mir-5001, hsa-mir-514a-1, hsa-mir-624, hsa-mir-639, hsa-mir-659, and hsa-mir-6892 | 1098 BC | TCGA | The ROC analysis validated the accuracy and stability of these two signatures as an independent prognostic indicators for BC patients. |
| [97] | circRNA OMA1 | 64 pairs of BC tissues and adjacent normal tissues | qRT-PCR | Associated with tumor size and lymph node metastasis. circOMA1 promoted viability, migration, and invasion of BC cells. circOMA1 promoted tumor progression by upregulating sirtuin 4 (SIR4) and miR-1276. circOMA1 together with mir-1276/SirT4 could be prognostic markers for BC |
| [98] | miR-10b | 61 BC patients and 48 healthy volunteers | qRT-PCR | The expression levels of serum miR-10b was progressively up-regulated in advanced stages of BC with higher levels in metastatic BC. Overall, the study evinced that simultaneous detections of miRNA10b and E-cadherin could be a robust serum biomarker to determine diagnosis and prognosticate BC metastasis. |
| [99] | SPRY4IT1 | 93 BC patients | qRT-PCR | SPRY4-IT1 was significantly expressed at higher levels in cancer tissues than normal tissues and its higher expression correlated also with higher rates of lymph node metastasis and recurrence and poor clinical response SPRY4-IT1 expression was a prognostic biomarker of poor clinical response in BC. |
| [100] | miR-923 and CA 15-3 | 253 BC patients | ddPCR | The miR-923 and CA 15-3 were significantly associated with prognosis. Combination of miR-923 and CA 15-3 detected from serum are a good preoperative, non-invasive prognostic marker of BC. |
| [101] | miR-138 | 30 BC and 10 healthy individuals | ddPCR | The level of miR-138 increased significantly in TNBC and correlated with poor prognosis. miR-138 was a good diagnostic and prognostic biomarker of TNBC. |
| [102] | ncRNA plasmacytoma variant translocation 1 (PVT1) | 3,974 patients (39 articles). | Meta-analysis | PVT1 correlated with low OS TNM stage, tumor size, lymph node metastasis, and distant metastasis. |
| [103] | HOTAIR | 15 BC patients treated surgically and 15 healthy individuals enrolled as controls; 25 BC patients received neoadjuvant chemotherapy before surgery and another 25 BC patients received tamoxifen hormone treatment after surgery | qRT-PCR | Potential diagnostic and prognostic biomarker since its high expression correlated with poor response to neoadjuvant chemotherapy and tamoxifen response |
| [104] | miR-155, miR-320a, and miR-205 | 28 studies | qRT-PCR | miR-155, miR-320a, and miR-205 could provide just some information on the major clinical-pathological features of BC, but not being used as a biomarker |
| [105] | miR-34 | 114 TNBC and blood samples from 124 healthy donors | IHC | TC and CC alleles associated with unfavorable prognosis in TNBC and that they could be used as a prognostic biomarker |
| [106] | miRNA-mediated prognostic modules | ProModule | New computational approach | |
| [107] | hsa-miR-10a, hsa-miR-18a, hsa-miR-135b and hsa-miR-577 | 1098 TNBC | TCGA | The 4-miRNA signature was an independent prognostic factor of clinical variables in TNBC patients andit could be used as a prognostic biomarker for TNBC patients |
| [108] | ACTA2-AS1, RP11-384P7.7, RP11- 327J17.9, RP11-124N14.3, and RP11-645C24.5 | 511 breast cancer tissues vs. 59 normal tissues | TCGA | Potential biomarkers to predict potential diagnosis and prognosis of BC |
| [109] | circmiR-122 | 90 BC and 60 healthy controls | qRT-PCR | miR-122 could predict metastasis at a cutoff value of 10.9 with a sensitivity of 95.83% and a specificity of 65.15%. miR-122 expression could be a biomarker for OS and PFS |
| [110] | miR-16, miR-145, miR-155, miR- 451a, miR-21 and miR-486 | 38 luminal A BC patients vs. 20 healthy controls | qRT-PCR | The absolute value of the three miRNAs had a prognostic value in luminal A BC |
| [111] | IncRNA BCAR4 | 890 BC | Meta-analysis | High lncRNA BCAR4 expression correlated with poor OS. The higher levels of lncRNA BCAR4 significantly correlated with increased tumor stage, lymph node, and distant metastases |
| [112] | miR-331 | 130 malignant and 66 benign breast cancer surgically resected from primary tumors | q-RT-PCR | miR-331 significantly correlated with malignant breast tumors compared to their benign counterparts. miR-331 could be considered a good prognostic marker for BC. |
| [113] | miR-134, miR-125b-5P, miRNA-30a, miR-10a-5p and miR-222 | 176 BC patients | qRT-PCR | These 5 miRNAs could be used to predict distant recurrence during tamoxifen treatment. |
| [114] | LINC01296 | 55 BC patients | RT-qPCR | LINC01296 could be a negative prognostic biomarker that could be used to predict disease progression as well as an actionable target. |
| [115] | miR-940 | 128 BC patients | qRT-PCR | miR-940 could be an independent prognostic factor and a reliable biomarker for diagnosis and prognosis of BC. |
| [116] | HOTAIR | 112 BC patients | qRT-PCR | The high expression levels of HOTAIR correlated with response to neoadjuvant chemotherapy as well as to a worse BC prognosis. |
| [91] | miR-330-3p | 233 BC patients | qRT-PCR | miR-330-3p upregulation associated with prognosis in BC patients, suggesting that it could be a prognostic biomarker and an actionable treatment. |
| [92] | ncRNA variants associated with BC profiles | 930 BC patients | TCGA | The authors observed that the overall mutation rate in coding and non-coding regions were significantly higher in ER− /HER2+ tumors |
| [93] | miR-191-5p, miR-214-3p, miR-451a, and miR-489 | 449 BC patients | qRT-PCR | Higher expression levels of hsa-miR-221-3p was observed in BC tissues than in adjacent healthy tissuebut there was no significant correlation between hsa-miR-221-3p and clinicopathological characteristics |
| [94] | hsa-miR221-3p | 40 | qRT-PCR | Higher expression levels of hsa-miR-221-3p was observed in BC tissues than in adjacent non-cancerous breast biopsies |
| [95] | PINK1.AS, RP11.259N19.1, KLF3.AS1, LINC00339, LINC00472, RP11.351I21.11, KB.1460A1.5, PKD1P6.NPIPP1, PDCD4.AS1, KLF3.AS1 PP14571, RP11.69E11.4 | 298 from GEO and 160 from TCGA | TCGA | Reliable prognostic and predictive biomarkers for disease relapse in BC patients receiving tamoxifen. |
| [96] | miR-597 | 190 paired fresh BC and non-cancerous BC | qRT-PCR | A close correlation was found between low miR-597 expression with positive lymph node metastasis advanced TNM stage poorer tumor differentiation, unfavorable OS and higher expression levels of miR-597. miR-597 can be an independent prognostic indicator of BC |
| [97] | miR-29b | 121 BC and 56 benign breast tissue specimens | qRT-PCR | Increased levels of miR-29b had a significantly longer disease-free survival and a lower risk to relapse |
| [98] | miR-301 | 380 BC samples | TCGA | Higher expression of miR-301a in BC cases is correlated with reduction of 5-year DFS and OS compared to BC with low levels of miR-301a expression. |
| [99] | MALAT1 | 80 BC cases compared to 80 controls. | qRT-PCR | A positive correlation was observed between MALAT1 expression with lymph node status, ER status, tumor stage, and histological grade indicating its possible prognostic value. |
| [100] | lncRNA00544 | 373 primary BC samples of 49 BC tissues and pair-matched metastatic axillary nodes | qRT-PCR | Elevated expression of lncRNA00544 was correlated with poor disease-free survival. lncRNA00544 can represent a novel predictive and prognostic biomarker in luminal BC patients. |
| [117] | miR-9 and miR-155 | 190 TNBC | qRT-PCR | Increased miR-9 levels showed significant association with poor PFS and distant metastasis–free survival (DMFS) in TNBC, whereas high level of miR-155 expression was associated with better DMFS miR-9 and miR-155 can be prognostic biomarkers in TNBCs. |
| [118] | miR-629-3p | 669 patients without de novo stage IV TNBC | qRT-PCR and IHC | miR-629-3p was correlated with poor OS and DFS in the validation set, but it failed to show significance after multivariate analysis. |
| [119] | miR-101 | 781 patients with BC | TCGA | Low levels of miR-101-2 expression might represent a diagnostic) marker, whereas the miR-101-1 was a prognostic marker. There was a close correlation between ER, PR, and HER2, while miR-101-2 was correlated with the tumor (T), lymph node (N), and metastasis (M) stages of BC. |
| [120] | LincIN | 781 BC patients | TCGA | Overexpression of LincIN was associated with BC aggressiveness and shorter OS. Ablation of LincIN showed inhibition of tumor cell migration and invasion in vitro and diminished lung metastasis in a mouse tail vein injection model. |
| [121] | SPRY4-IT1 | 110 BC | qRT-PCR | Increased expression of Z38 was found in BC compared to controls, advanced TNM stage, presence of lymph node metastasis and unfavorable OS. |
| [122] | 10 miRNAs, including (miR-7,-21,-29a,-29b,-34a,-125b,-155,-200c,-340,-451) | 64 BC patients | PCR | The patients with miR-7low or miR-340 high profile might not have complete response. |
| [123] | miR-199b-5p | 19 BC samples | qRT-PCR | Low expression of MiR-199b-5p showed close association with advanced TNM stage, positive lymph node metastasis and poor OS miR-199b-5p might be a possible marker for BC. |
| [124] | MALAT1 | 33 pairs of primary non-metastatic BC and their matched adjacent normal tissues 204 BC tissues | TCGA | Up-regulation of MALAT1 was associated with poor RFS in tamoxifen-treated ER-positive BC patients, which might present as a candidate biomarker to predict endocrine treatment sensitivity. |
| [125] | 12 circulating miRNAs in serum of inflammatory and non-inflammatory BC | 1014 BC patients | TCGA | Overexpression of miR-335 in premenopausal non-inflammatory BC patients, whereas miR-24 was significantly upregulated in non-inflammatory BC with postmenopausal status. |
| [126] | MALAT1 | 446 unilateral invasive primary BC | RT-PCR | The authors reported a complex expression pattern of various MALAT1 transcript variants in BC cases and the prognostic and predictive role of MALAT1 should be considered conservatively. |
| [127] | Hsa-miR-375 | 115 patients (30 relapses versus 85 controls) | qRT-PCR | Positive association between the levels of has-miR-375 with local relapshashsa-miR-375 can distinguish between relapse and control groups |
| [128] | miR-520g | 86 cases with BC | qRT-PCR | Higher levels of miR-520g were found in BC patients with lymph node metastatic and low differentiation degree grade mammary gland invasion and low expression of p53. miR-520g might be a potential prognostic factor in BC. |
| [129] | miR-200a, miR-200b, miR-200c, miR-210, miR-215 and miR-486-5p | Primary BC with metastasis (M1, n = 67) at diagnosis/blood collection, and patients without metastasis at diagnosis (M0, n = 265) plasma miRNAs of 40 MBC patients 237 MBC patients | TaqMan low density arrays | A significant correlation was found between miR-200a, miR-200b, miR-200c, miR-210, miR-215, and miR-486-5p with metastasis development before clinical manifestation of BC. |
| [130] | CCAT1 | 92 pairs of BC cancer tissues and adjacent normal tissues | qRT-PCR | Significant correlation between CCAT1 with poor differentiation grade, advanced TNM stage, presence of lymph node metastases, and shorter OS and PFS. CCAT1 could be a possible prognostic marker for BC progression. |
| [131] | miR-124 | 133 BC patients | qRT-PCR | This group showed that miR-124 can be an indicator of tumor progression and poor prognosis in BC cases. |
| [132] | lncRNA microarray data from 164 primary breast tumors from adjuvant naïve patients | 82 patients cases with detectable distant metastasis were compared to 82 patients with no metastases. | Microarray analysis | That lncRNA profiles could distinguish metastatic patients from non-metastatic patients with sensitivities above 90% and specificities of 64–65%. |
| [133] | HOTAIR | 133 BC cases | TCGA | HOTAIR might be an indicator of lymphatic metastases rather than hematogenous metastases in ER—BC. |
| [134] | miR-21 | 549 cases (326 with breast cancer, 223 without breast cancer) | (RQ)-PCR | Increased expression ofmiR-21 was reported in tissues and serum of BC patients versus healthy control groups in the Chinese population and it can be an indicator of recurrence and disease-free survival |
| [135] | miR-106b | Plasma samples of 173 patients with primary BC and a set of 50 women with fibroadenoma | qRT-PCR and in situ hybridization. | Increased levels of miR-106b were correlated with higher Ki67 expression, lymph node metastasis, shorter PFS, and OS. miR-106b might be a high risk of recurrence of BC. |
| [136] | miR34a | Three independent primary BC cohorts (Cohort 1 with 461, Cohort 2 with 279 and Cohort 3 with 795 patients) | quantitative in situ hybridisation assay (qISH) | Loss of miR34a can distinguish patients with poor PSF among node-negative patients, but not in the node-positive population. Loss of miR34a might be an indicator of a subgroup of BC patients with unfavorable disease-specific survival |
| [137] | miR-21, miR-210, and miR-373 | Serum of 127 HER2-postive BC patients before and after neoadjuvant therapy and 19 healthy controls | TaqMan MicroRNA assays | Close association between neoadjuvant therapy and the serum levels of miR-21, miR-210, and miR-373 in BC cases with a prognostic value for miR-21. |
| [138] | miR-27b-3p, miR-107, and miR-103a-3p | 99 TNBC patients including a training set of 58 patients with invasive ductal TNBC further validated in a separate set of 41 TNBC patients | qRT-PCR | Expressions of miR-27b-3p, miR-107, and miR-103a-3p were significantly up-regulated in the metastatic group versus the disease-free. Lymph node status and miR-27b-3p were independent predictors of poor prognosis. |
| [139] | miR-10b | 101 paired tumor and normal specimens | qRT-PCR | Adding miR-10b RERs to the prognostic factors used in clinical routine could improve the prediction abilities for overall mortality as well as progression in BC patients. |
| [140] | miR-630 | all breast tissue (n = 56) and HER2+ breast tissue (n = 6) | qRT-PCR | Induction of miR-630 into cells with innate- or acquired- resistance to HER-drugs significantly restored the efficacy of lapatinib, neratinib and afatinib |
| [141] | miR-149, miR-10a, miR-20b, miR-30a-3p, and miR-342-5p, | 71 primary BC | Microarray and RT-qPCR | These five 5-miRNA signatures determined a high-risk group of patients with shorter relapse-free survival as well as non-relapsing versus early-relapsing patients. possible prognostic value to identify patients with metastasis development after primary breast surgery. |
| [142] | miR-155, miR-493, miR-30e and miR-27a | 160 TNBC | First: miRNA expression profiling with GEO and TCGA Second: TaqMan® qRT-PCR assay | TNBC subclassification based on the 5 IHC markers and on the miR-155, miR-493, miR-30e, miR-27a expression levels are a powerful diagnostic approach. |
| [143] | miR-26a, miR-26b, miR-203, miR-421, miR-664, miR-576-5p and miR-18a | 52 BC and 3 normal breast samples | qRT-PCR | Increased expression of miR-421 was detected in 36.5% of cases which exhibit lower ATM transcript levels. It is clear that ATM protein expression might represent an independent prognostic marker in sporadic BC. |
| [144] | HOTAIR | 164 primary BC without adjuvant therapy | microarray | HOTAIR expression may provide an independent biomarker for the prediction of the risk of metastasis in ER + BC patients. |
| [145] | CCAT2 | 997 primary BC | qRT-PCR and ISH | CCAT2 up-regulates cell migration and down-regulates chemosensitivity to 5’FU in a rs6983267-independent manner. |
| [146] | RAD21 | 28 BRCA1, 27 BRCA2, and 39 BRCAX | IHC | RAD21 is a potential predictive and prognostic biomarker in familial breast cancers. |
| [147] | miR-205 | 40 FFPE archival BC | in situ hybridization analysisof micro-RNA expression in arrays | Expression of miR-205 is associated with tumors of ductal morphology and thus this molecule can be considered as a prognostic marker within these tumours. |
| [148] | miR-210 | 56 systemically untreated BC patients | miRNA microarray hybridization qRT-PCR | The effects of MiR-210 were analyzed on the BC cells, including MCF7 and MDA-MB-231. MiR-210 expression showed that this molecule was involved in cell proliferation, migration and invasion. |
| [149] | miR-106b | 103 lymph node negative BC | miRCURY LNA Array | Also demonstrated the presence of several microRNAs, including miR532-5p, miR-500, miR362-5p, and miR502-3p, located at Xp11.23 in cancers with a triple-negative signature, and the increased expression of several miR-17 cluster members in ER− tumors. |
| [150] | Dicer expression | 104 BC patients | qRT-PCR | A close correlation was reported between Dicer protein expression and hormone receptor status and subtypes in BC (Dicer expression might be an indicator of distant metastases in BC cases. |
| [151] | 8 mRNAs and 2 lncRNAs | In the training cohort, a total of 198 frozen tissues from 165 consecutive TNBC patients (including 33 pairs of tumor and adjacent normal tissues) 266 frozen TNBC samples and 33 adjacent normal breast tissue | First: GEO Second: validation with qRT-PCR | Tumor-specific mRNAs and lncRNAs were identified and correlated with patients’ recurrence-free survival (RFS). |
| [152] | EPB41L4A-AS2 | 250 BC tissues and 50 adjacent normal tissues | qRT-PCR | Induction of EPB41L4A-AS2 expression inhibited breast tumor cell proliferation. It can be concluded that evaluation of this long non-coding RNA might provide a possible prognostic biomarker in the clinical management of BC. |
| [153] | miR-29c and miR-101 | 2178 BC | GEO, EGA, TCGA | The authors demonstrated that miR-29c and miR-101 might have prognostic value in BC patients. |
| [154] | miR-205 | 30 BC patients | qRT-PCR | miR-205 may be valuable for prediction of the TAC regimen as well as a possible therapeutic target in BC treatment. |
| [155] | miR-7, miR-22, miR-21, miR-30c, miR-181a, miR-181c, miR-125b, miR-200a, miR-135b, and miR-200c | 818 BC patients | qRT-PCR | The 10-miRNAs were a good prognostic biomarker to predict distant relapse free survival (DRFS) in BC. |
| [156] | 12 differentially expressed lncRNAs | 473 BC patients | GEO | This lncRNAs was closely associated with tumor recurrence of BC from discovery cohort, which was capable to classify patints into high-risk and low-risk with recurrence-free survival that was significantly |
| [157] | Holistic IDFO approach of prioritization of cancers | 5 human cancers, including breast cancer, in 3198 samples | TCGA | lncRNAs closely associated with tumor recurrence of BC from discovery cohort, which was capable to classify patients into high-risk and low-risk with recurrence-free survival that was significantly different |
| [158] | 14 miRNAs | Training (n = 596) and Testing set (n = 319) | TCGA | Patients could be characterized as high and low score according to the risk scores calculated for each miRNA. The signature could be used as prognostic marker in ER+ BC. |
| [159] | Drosha and Dicer | 245 patients receiving adjuvant anthracycline-based chemotherapy | qRT-PCR | Concurrent down-regulation of Drosha and dicer in 15% of cases and a significant association with both high grade and ki-67 index. No significant association between the down-regulation of Drosha and/or Dicer and outcomes. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobhani, N.; Chahwan, R.; Roudi, R.; Morris, R.; Volinia, S.; Chai, D.; D’Angelo, A.; Generali, D. Predictive and Prognostic Value of Non-Coding RNA in Breast Cancer. Cancers 2022, 14, 2952. https://doi.org/10.3390/cancers14122952
Sobhani N, Chahwan R, Roudi R, Morris R, Volinia S, Chai D, D’Angelo A, Generali D. Predictive and Prognostic Value of Non-Coding RNA in Breast Cancer. Cancers. 2022; 14(12):2952. https://doi.org/10.3390/cancers14122952
Chicago/Turabian StyleSobhani, Navid, Richard Chahwan, Raheleh Roudi, Rachel Morris, Stefano Volinia, Dafei Chai, Alberto D’Angelo, and Daniele Generali. 2022. "Predictive and Prognostic Value of Non-Coding RNA in Breast Cancer" Cancers 14, no. 12: 2952. https://doi.org/10.3390/cancers14122952
APA StyleSobhani, N., Chahwan, R., Roudi, R., Morris, R., Volinia, S., Chai, D., D’Angelo, A., & Generali, D. (2022). Predictive and Prognostic Value of Non-Coding RNA in Breast Cancer. Cancers, 14(12), 2952. https://doi.org/10.3390/cancers14122952