
Pharmacologic Targeting of MMP2/9 Decreases Peritoneal Metastasis Formation of Colorectal Cancer in a Human Ex Vivo Peritoneum Culture Model

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
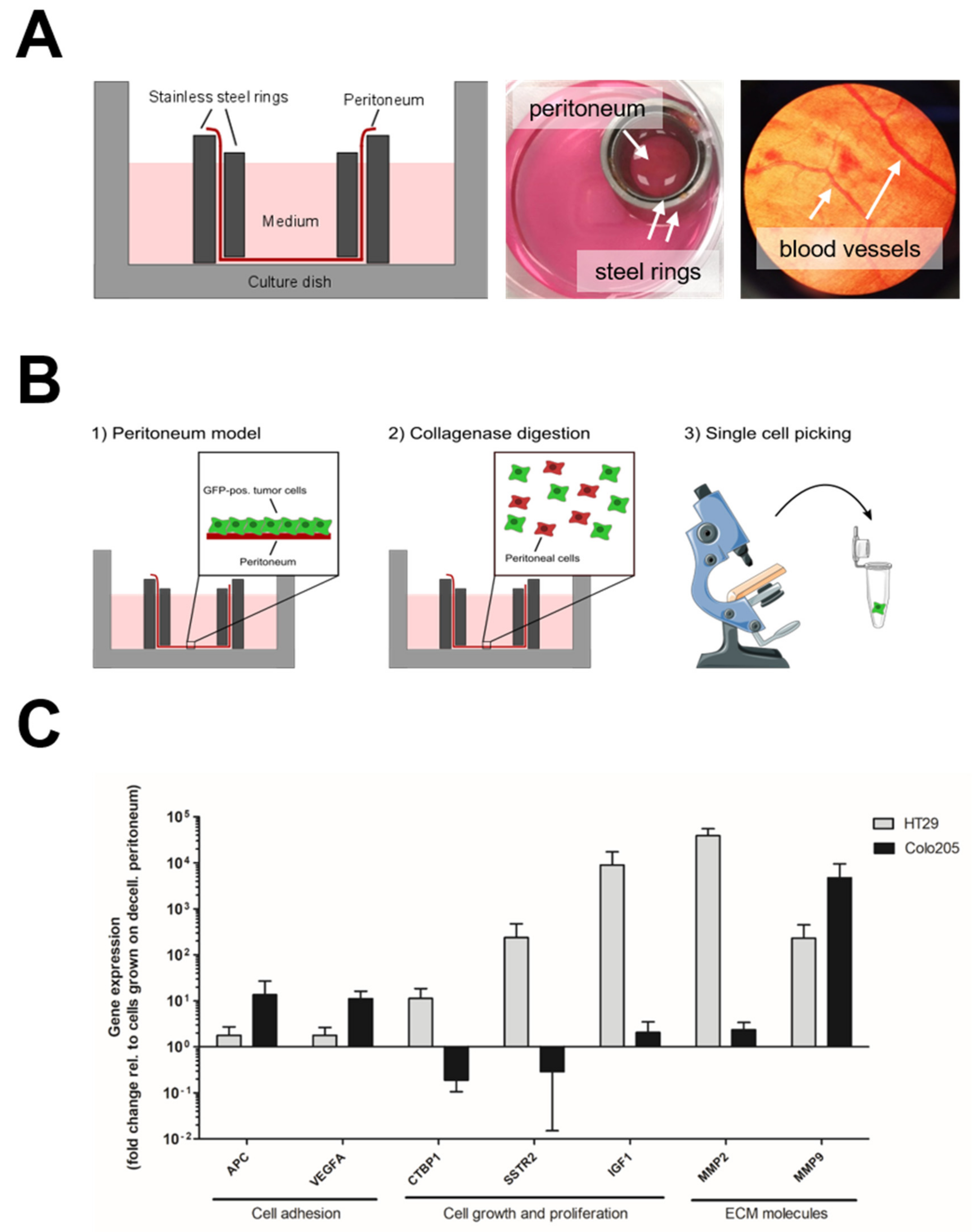
2.2. Peritoneal Culture
2.3. Decellularisation of Peritoneal Tissue
2.4. Co-Culture
2.5. MMP Inhibitor Treatment
2.6. Cell Counting
2.7. Single-Cell Isolation
2.8. Whole-Transcriptome Analysis (WTA) of Single Cells
2.9. Quantitative Real-Time PCR (RT2 and qRT-PCR Assays)
2.10. siRNA Treatment
2.11. Gel Electrophoresis and Immunoblotting
2.12. Protein Quantification
2.13. Immunohistochemistry
2.14. Statistical Analysis
3. Results
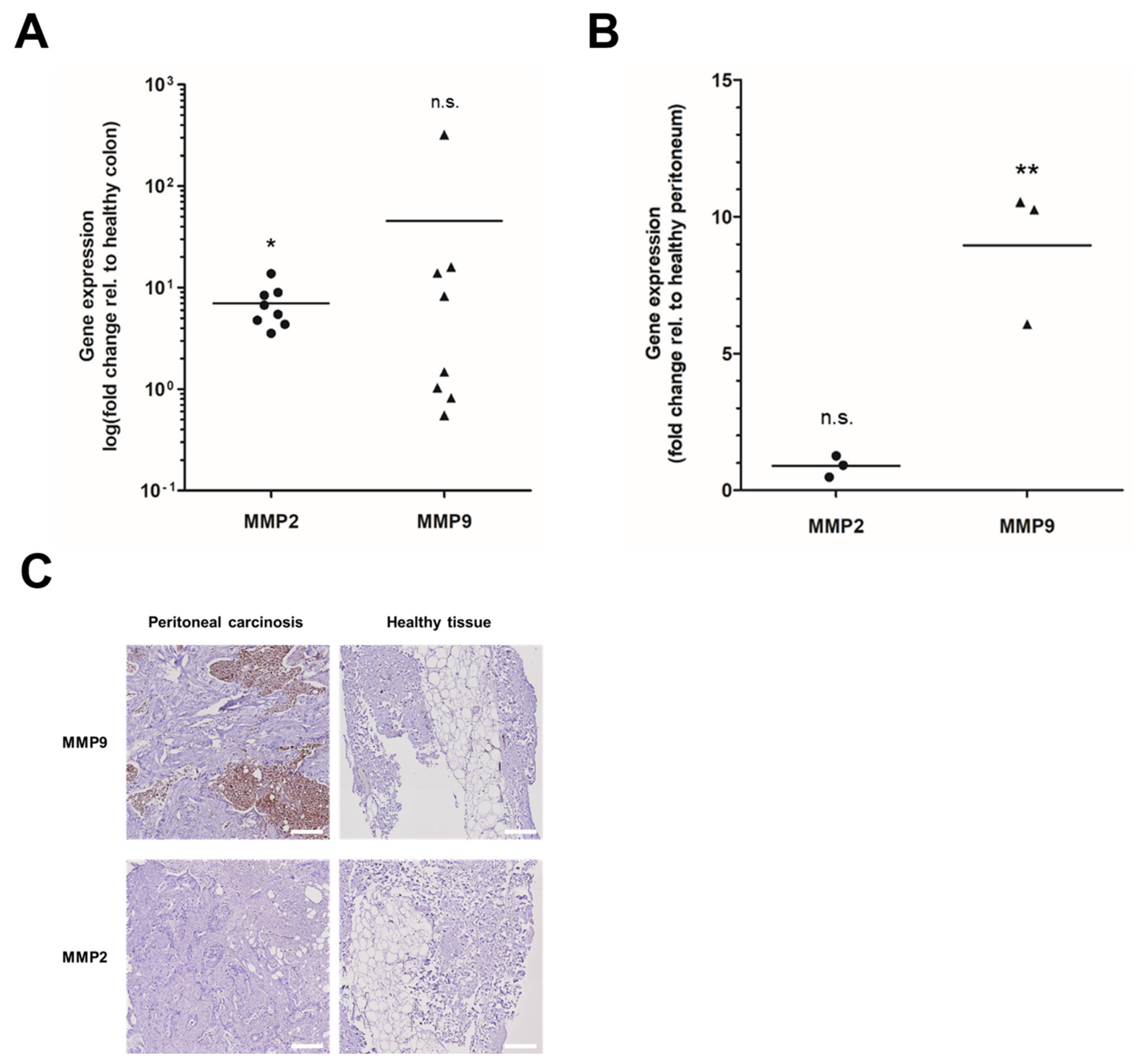
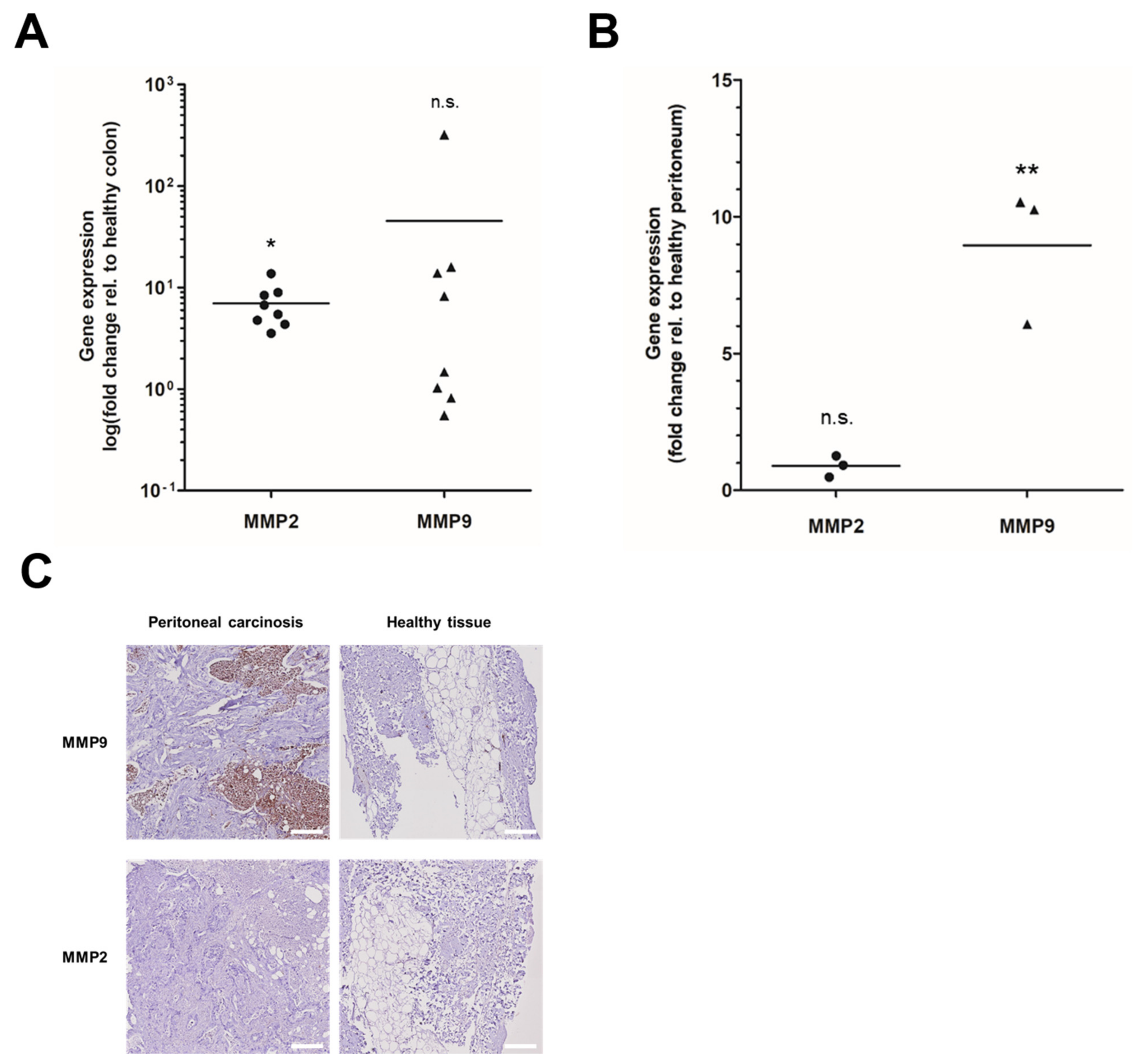
3.1. MMP Overexpression in Primary and Peritoneal Human CRC Samples
3.2. Upregulation of MMPs in Colonising Peritoneal Cancer Cells in a Human Ex Vivo Model
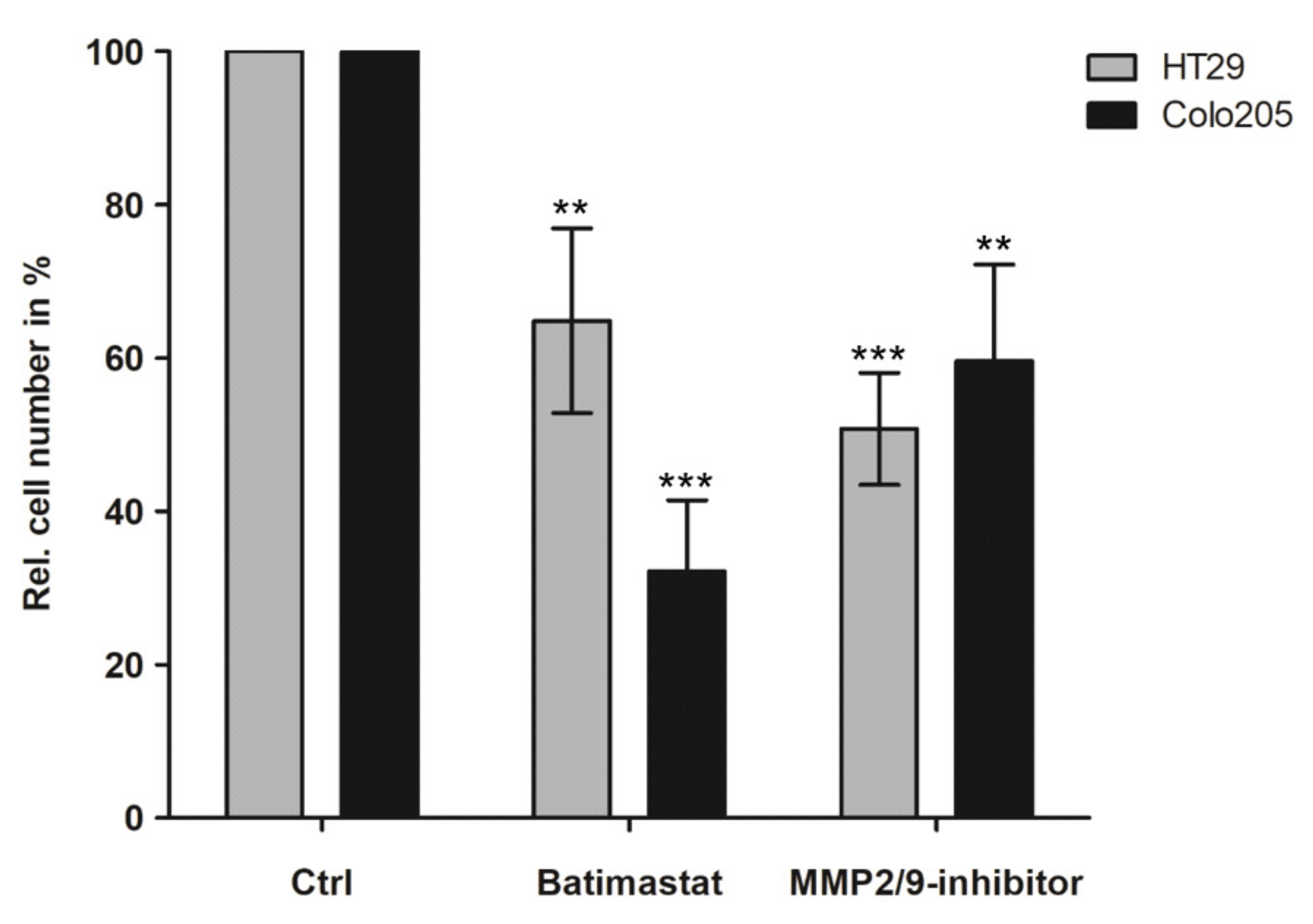
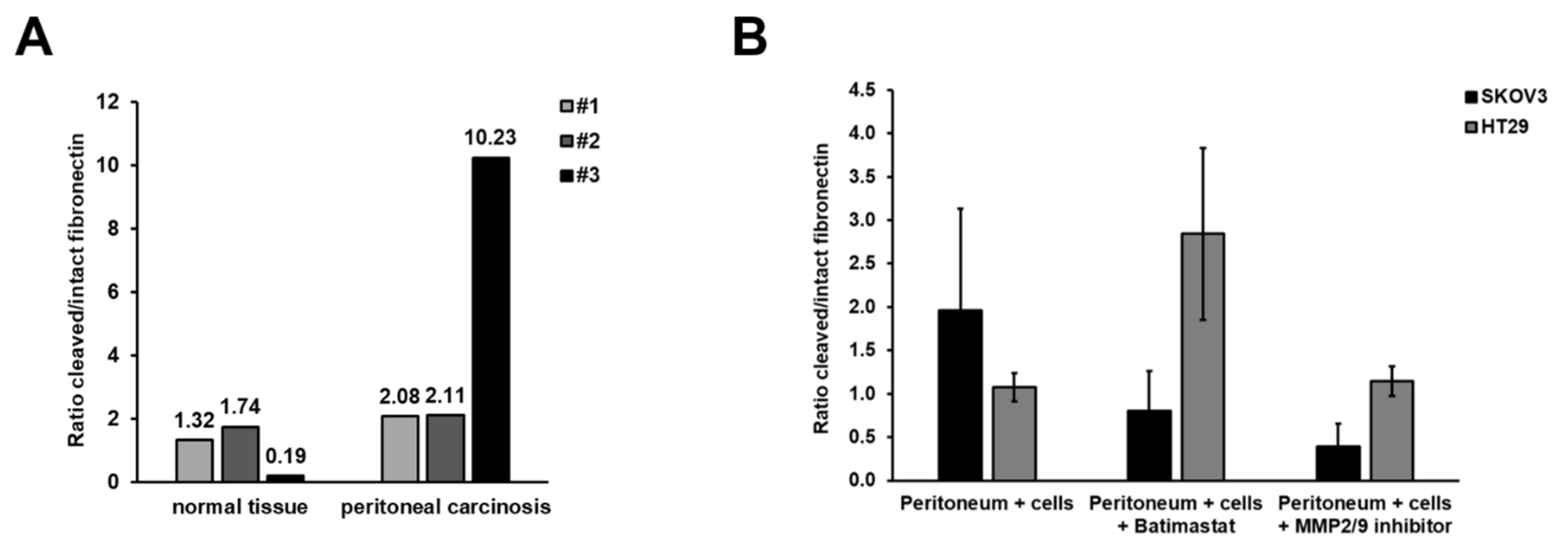
3.3. Inhibition of Peritoneal Colonisation of Cancer Cells by MMP2/9 Blockage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dahdaleh, F.S.; Turaga, K.K. Evolving Treatment Strategies and Outcomes in Advanced Gastric Cancer with Peritoneal Metastasis. Surg. Oncol. Clin. N. Am. 2018, 27, 519–537. [Google Scholar] [CrossRef] [PubMed]
- Verwaal, V.J.; van Ruth, S.; de Bree, E.; van Sloothen, G.W.; van Tinteren, H.; Boot, H.; Zoetmulder, F.A.N. Randomized Trial of Cytoreduction and Hyperthermic Intraperitoneal Chemotherapy versus Systemic Chemotherapy and Palliative Surgery in Patients with Peritoneal Carcinomatosis of Colorectal Cancer. J. Clin. Oncol. 2003, 21, 3737–3743. [Google Scholar] [CrossRef] [PubMed]
- Klaver, Y.L.B.; Hendriks, T.; Lomme, R.M.L.M.; Rutten, H.J.T.; Bleichrodt, R.P.; de Hingh, I.H.J.T. Hyperthermia and Intraperitoneal Chemotherapy for the Treatment of Peritoneal Carcinomatosis: An Experimental Study. Ann. Surg. 2011, 254, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.M.; Morris, M.C.; Sohal, D.; Sussman, J.J.; Wilson, G.C.; Ahmad, S.A.; Patel, S.H. Cytoreductive Surgery and Hyperthermic Intraperitoneal Chemotherapy for Colorectal Peritoneal Metastases: A Summary of Key Clinical Trials. J. Clin. Med. 2022, 11, 3406. [Google Scholar] [CrossRef]
- Quénet, F.; Elias, D.; Roca, L.; Goéré, D.; Ghouti, L.; Pocard, M.; Facy, O.; Arvieux, C.; Lorimier, G.; Pezet, D.; et al. Cytoreductive Surgery plus Hyperthermic Intraperitoneal Chemotherapy versus Cytoreductive Surgery Alone for Colorectal Peritoneal Metastases (PRODIGE 7): A Multicentre, Randomised, Open-Label, Phase 3 Trial. Lancet Oncol. 2021, 22, 256–266. [Google Scholar] [CrossRef]
- Klaver, C.E.L.; Wisselink, D.D.; Punt, C.J.A.; Snaebjornsson, P.; Crezee, J.; Aalbers, A.G.J.; Brandt, A.; Bremers, A.J.A.; Burger, J.W.A.; Fabry, H.F.J.; et al. Adjuvant Hyperthermic Intraperitoneal Chemotherapy in Patients with Locally Advanced Colon Cancer (COLOPEC): A Multicentre, Open-Label, Randomised Trial. Lancet Gastroenterol. Hepatol. 2019, 4, 761–770. [Google Scholar] [CrossRef]
- Goéré, D.; Glehen, O.; Quenet, F.; Guilloit, J.-M.; Bereder, J.-M.; Lorimier, G.; Thibaudeau, E.; Ghouti, L.; Pinto, A.; Tuech, J.-J.; et al. Second-Look Surgery plus Hyperthermic Intraperitoneal Chemotherapy versus Surveillance in Patients at High Risk of Developing Colorectal Peritoneal Metastases (PROPHYLOCHIP–PRODIGE 15): A Randomised, Phase 3 Study. Lancet Oncol. 2020, 21, 1147–1154. [Google Scholar] [CrossRef]
- Mikuła-Pietrasik, J.; Uruski, P.; Tykarski, A.; Książek, K. The Peritoneal “Soil” for a Cancerous “Seed”: A Comprehensive Review of the Pathogenesis of Intraperitoneal Cancer Metastases. Cell. Mol. Life Sci. 2018, 75, 509–525. [Google Scholar] [CrossRef]
- Sawada, K.; Mitra, A.K.; Radjabi, A.R.; Bhaskar, V.; Kistner, E.O.; Tretiakova, M.; Jagadeeswaran, S.; Montag, A.; Becker, A.; Kenny, H.A.; et al. Loss of E-Cadherin Promotes Ovarian Cancer Metastasis via Alpha 5-Integrin, Which Is a Therapeutic Target. Cancer Res. 2008, 68, 2329–2339. [Google Scholar] [CrossRef] [Green Version]
- Said, A.H.; Raufman, J.-P.; Xie, G. The Role of Matrix Metalloproteinases in Colorectal Cancer. Cancers 2014, 6, 366–375. [Google Scholar] [CrossRef]
- Langers, A.M.J.; Verspaget, H.W.; Hawinkels, L.J.A.C.; Kubben, F.J.G.M.; van Duijn, W.; van der Reijden, J.J.; Hardwick, J.C.H.; Hommes, D.W.; Sier, C.F.M. MMP-2 and MMP-9 in Normal Mucosa Are Independently Associated with Outcome of Colorectal Cancer Patients. Br. J. Cancer 2012, 106, 1495–1498. [Google Scholar] [CrossRef] [Green Version]
- Falk, P.; Ruiz-Jasbon, F.; Strigård, K.; Gunnarsson, U.; Ivarsson, M.-L. An Ex Vivo Model Using Human Peritoneum to Explore Mesh-Tissue Integration. Biol. Open 2017, 6, 1391–1395. [Google Scholar] [CrossRef] [Green Version]
- Klein, C.A.; Zohlnhöfer, D.; Petat-Dutter, K.; Wendler, N. Gene Expression Analysis of a Single or Few Cells. Curr. Protoc. Mol. Biol. 2003. [Google Scholar] [CrossRef]
- Werner-Klein, M.; Grujovic, A.; Irlbeck, C.; Obradović, M.; Hoffmann, M.; Koerkel-Qu, H.; Lu, X.; Treitschke, S.; Köstler, C.; Botteron, C.; et al. Interleukin-6 Trans-Signaling Is a Candidate Mechanism to Drive Progression of Human DCCs during Clinical Latency. Nat. Commun. 2020, 11, 4977. [Google Scholar] [CrossRef]
- Mönch, D.; Koch, J.; Maaß, A.; Janssen, N.; Mürdter, T.; Renner, P.; Fallier-Becker, P.; Solaß, W.; Schwab, M.; Dahlke, M.-H.; et al. A Human Ex Vivo Coculture Model to Investigate Peritoneal Metastasis and Innovative Treatment Options. Pleura Peritoneum 2021, 6. [Google Scholar] [CrossRef]
- Kenny, H.A.; Kaur, S.; Coussens, L.M.; Lengyel, E. The Initial Steps of Ovarian Cancer Cell Metastasis Are Mediated by MMP-2 Cleavage of Vitronectin and Fibronectin. J. Clin. Investig. 2008, 118, 1367–1379. [Google Scholar] [CrossRef]
- Botos, I.; Scapozza, L.; Zhang, D.; Liotta, L.A.; Meyer, E.F. Batimastat, a Potent Matrix Mealloproteinase Inhibitor, Exhibits an Unexpected Mode of Binding. Proc. Natl. Acad. Sci. USA 1996, 93, 2749–2754. [Google Scholar] [CrossRef] [Green Version]
- Pezeshkian, Z.; Nobili, S.; Peyravian, N.; Shojaee, B.; Nazari, H.; Soleimani, H.; Asadzadeh-Aghdaei, H.; Ashrafian Bonab, M.; Nazemalhosseini-Mojarad, E.; Mini, E. Insights into the Role of Matrix Metalloproteinases in Precancerous Conditions and in Colorectal Cancer. Cancers 2021, 13, 6226. [Google Scholar] [CrossRef]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Hua, H.; Li, M.; Luo, T.; Yin, Y.; Jiang, Y. Matrix Metalloproteinases in Tumorigenesis: An Evolving Paradigm. Cell. Mol. Life Sci. 2011, 68, 3853–3868. [Google Scholar] [CrossRef]
- Gabelloni, P.; Da Pozzo, E.; Bendinelli, S.; Costa, B.; Nuti, E.; Casalini, F.; Orlandini, E.; Da Settimo, F.; Rossello, A.; Martini, C. Inhibition of Metalloproteinases Derived from Tumours: New Insights in the Treatment of Human Glioblastoma. Neuroscience 2010, 168, 514–522. [Google Scholar] [CrossRef]
- Wang, K.; Zheng, J.; Yu, J.; Wu, Y.; Guo, J.; Xu, Z.; Sun, X. Knockdown of MMP-1 Inhibits the Progression of Colorectal Cancer by Suppressing the PI3K/Akt/C-myc Signaling Pathway and EMT. Oncol. Rep. 2020, 43, 1103–1112. [Google Scholar] [CrossRef]
- Wojtowicz-Praga, S.; Low, J.; Marshall, J.; Ness, E.; Dickson, R.; Barter, J.; Sale, M.; McCann, P.; Moore, J.; Cole, A.; et al. Phase I Trial of a Novel Matrix Metalloproteinase Inhibitor Batimastat (BB-94) in Patients with Advanced Cancer. Investig. New Drugs 1996, 14, 193–202. [Google Scholar] [CrossRef]
- Beattie, G.J.; Smyth, J.F. Phase I Study of Intraperitoneal Metalloproteinase Inhibitor BB94 in Patients with Malignant Ascites. Clin. Cancer Res. 1998, 4, 1899–1902. [Google Scholar]
- Parsons, S.L.; Watson, S.A.; Steele, R.J. Phase I/II Trial of Batimastat, a Matrix Metalloproteinase Inhibitor, in Patients with Malignant Ascites. Eur. J. Surg. Oncol. 1997, 23, 526–531. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, J.; Mönch, D.; Maaß, A.; Mangold, A.; Gužvić, M.; Mürdter, T.; Leibold, T.; Dahlke, M.-H.; Renner, P. Pharmacologic Targeting of MMP2/9 Decreases Peritoneal Metastasis Formation of Colorectal Cancer in a Human Ex Vivo Peritoneum Culture Model. Cancers 2022, 14, 3760. https://doi.org/10.3390/cancers14153760
Koch J, Mönch D, Maaß A, Mangold A, Gužvić M, Mürdter T, Leibold T, Dahlke M-H, Renner P. Pharmacologic Targeting of MMP2/9 Decreases Peritoneal Metastasis Formation of Colorectal Cancer in a Human Ex Vivo Peritoneum Culture Model. Cancers. 2022; 14(15):3760. https://doi.org/10.3390/cancers14153760
Chicago/Turabian StyleKoch, Jana, Dina Mönch, Annika Maaß, Alina Mangold, Miodrag Gužvić, Thomas Mürdter, Tobias Leibold, Marc-H. Dahlke, and Philipp Renner. 2022. "Pharmacologic Targeting of MMP2/9 Decreases Peritoneal Metastasis Formation of Colorectal Cancer in a Human Ex Vivo Peritoneum Culture Model" Cancers 14, no. 15: 3760. https://doi.org/10.3390/cancers14153760
APA StyleKoch, J., Mönch, D., Maaß, A., Mangold, A., Gužvić, M., Mürdter, T., Leibold, T., Dahlke, M.-H., & Renner, P. (2022). Pharmacologic Targeting of MMP2/9 Decreases Peritoneal Metastasis Formation of Colorectal Cancer in a Human Ex Vivo Peritoneum Culture Model. Cancers, 14(15), 3760. https://doi.org/10.3390/cancers14153760