1. Introduction
Over 2.3 million women are diagnosed with breast cancer every year, and ~685,000 will die as a result of this condition (
www.who.int) (accessed on 23 May 2022). Whilst improvements in targeted treatments for primary breast cancers have proven to be beneficial, with >80% of women making a full recovery, for patients whose tumours spread to distant organs, there are still no curative treatments. Around 70–80% of patients with late-stage breast cancer develop secondary tumours in their skeletons, and life expectancy for these patients drops to 2–3 years following diagnosis of bone involvement [
1]. There is, therefore, a clear need for improved therapies for patients whose tumours spread to bones. There is increasing evidence that the pro-inflammatory cytokine IL1β plays a critical role in breast cancer spread to the bone and metastatic outgrowth in this site: In patient samples, expression of IL1β by breast cancer cells is associated with an increased likelihood of future relapse in the bone [
2,
3]. Overexpression of IL1β in tumour cells facilitates epithelial to mesenchymal transition (EMT), migration and invasion towards bone cells in vitro, as well as inducing bone homing properties and promoting the dissemination of tumour cells in the bone in in vivo models [
2,
3,
4,
5,
6,
7,
8]. Once cancer cells are disseminated in the bone, interactions between tumour cells and the bone metastatic niche (osteoblasts, haematopoietic stem cells and blood vessels) lead to increased secretion of IL1β in the local environment. Environmental IL1β promotes the expansion of the niche, and IL1β driven wnt activation leads to metastatic outgrowth of tumour cells disseminated in this site [
2,
7,
9,
10]. Importantly, inhibition of IL-1 signalling with the anti-IL1β antibody Canakinumab, or the IL1R antagonist, Anakinra, inhibits bone metastasis in pre-clinical models of breast cancer. Similarly, Canakinumab has been shown to have potent anti-cancer effects in a clinical trial [
11], suggesting that targeting IL-1 may be an attractive therapeutic strategy for preventing/treating breast cancer-induced bone disease 1, [
12,
13].
In addition to its direct anti-metastatic properties, IL1β plays a central role in regulating immunity [
14], and it has recently become apparent that IL1β within the tumour microenvironment is important for recruiting anti-tumour immune cells, especially in soft tissue tumours [
7,
10]. Global removal of IL1β in IL1β knockout mice leads to increased growth of primary mammary tumours, and administration of Anakinra/Canakinumab also increases the growth of primary breast cancers in mice [
2,
13]. Anakinra also increases the growth of disseminated tumour cells in the lungs [
15,
16,
17]. Interestingly, IL1β appears to regulate anti-tumour immunity differently in the bone and soft tissue sites, promoting the recruitment of anti-tumour macrophages, neutrophils and Th1 cells in soft tissues and reducing myeloid cells in the bone [
13,
18]. Moreover, the administration of Canakinumab had been shown to significantly reduce the development of lung cancers in patients with atherosclerosis but had the adverse effect of increasing incidences of serious infections [
11,
19].
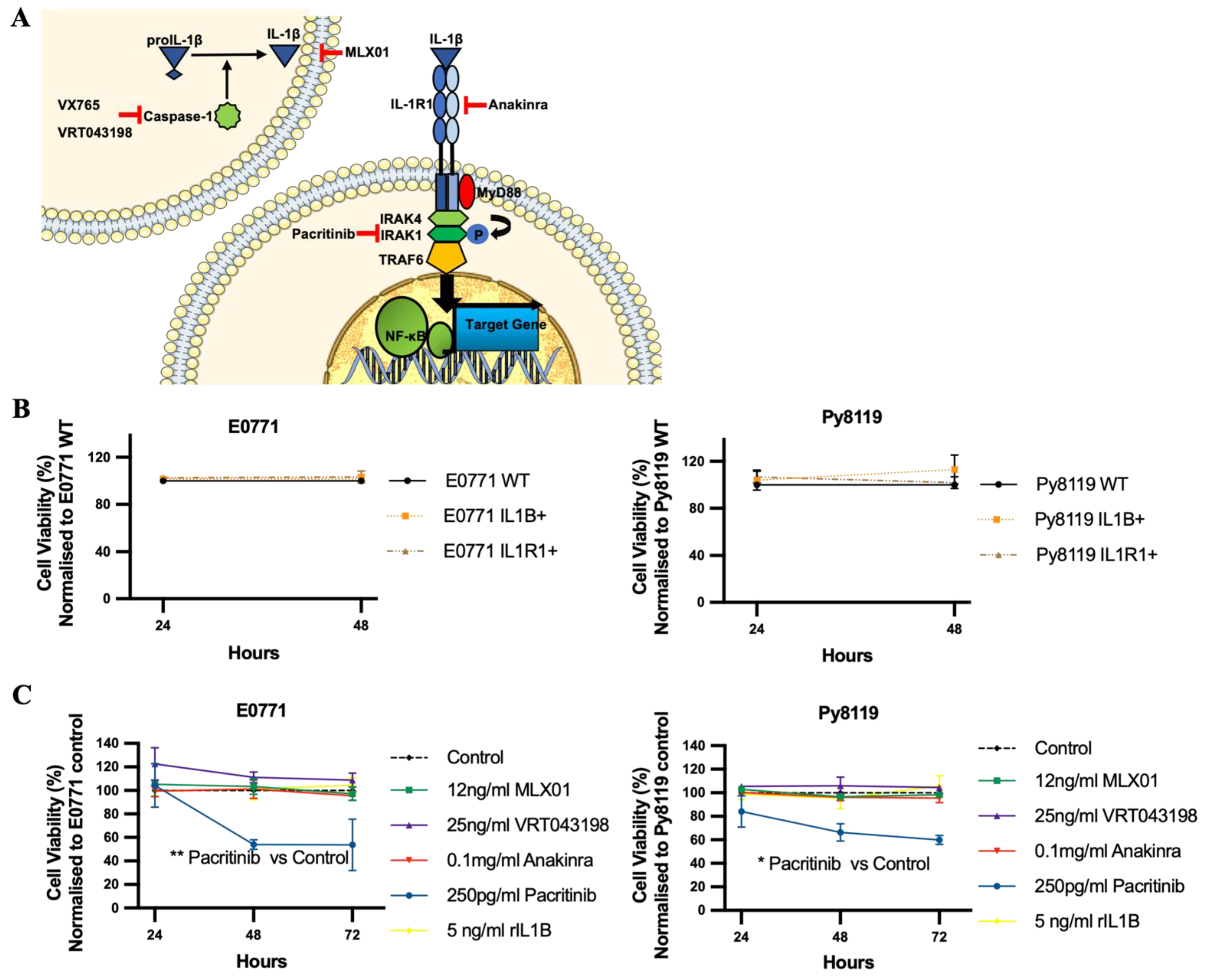
Given the promising outcomes of inhibiting IL-1 in suppressing breast cancer bone metastasis, we have evaluated the effects of small molecule inhibitors that target different parts of the IL1β signalling pathway (Caspase-1, IL1β and IRAK1, Interleukin 1 Receptor Associated Kinase 1) on breast cancer bone metastasis. Using in vitro and in vivo models, we have attempted to identify these inhibitors that could prevent IL1β-driven bone metastasis without stimulating tumour growth outside the bone or adversely affecting systemic immunity.
2. Materials and Methods
2.1. Cell Lines
Mouse mammary tumour E0771 were originally provided by Professor Sandra McAllister’s laboratory (Department of Medicine, Harvard Medical School, Boston, MA, USA) and Py8119 cells purchased from ATCC. These cells were manipulated to make E0771 Luc2 V5 GFP, E0771 Luc2 V5 IL1B GFP, E0771 Luc2 V5 IL1R1 cell lines and Py8119 Luc2 V5 GFP, Py8119 Luc2 V5 IL1B GFP, and Py8119 Luc2 V5 IL1R1 GFP as previously published and expression levels of IL1B and IL1R1 were confirmed by real-time PCR and ELISA (
Figure S1A–D) [
13]. E0771 and Py8119 Cell lines were cultured in RPMI 1640 Medium (61870-010, Gibco, Waltham, MA, USA) with 10%
v/
v fetal bovine serum (FBS) (F7524, Sigma, St. Louis, MI, USA). Pre-osteoblast OB1 cells were a kind gift from Marianna Kruithof de Julio (University of Bern, Bern, Switzerland), and this cell was cultured by DMEM medium (61966-021, Gibco) with 10%
v/
v FBS. HUVECs were purchased from PromoCell (Heidelberg, Germany) and cultured in the media supplied (Promocell, Heidelberg, Germany, C-12211 and C-39220).
2.2. Drugs
Anakinra (r-metHu1L-ra) was obtained from Amgen, Cambridge, UK. MLX01 was obtained from Michael Leach University of Greenwich, and the evidence of MLX01 inhibition of IL1β production in human cell lines is shown in
Figure S2A, and in mouse tissue is shown in
Figure S2B. VX765 (S2228) was obtained from Selleckchem (Boston, MA, USA). VRT043198 (244133-31-1) and Pacritinib (202571) were purchased from Medkoo (Morrisville, NC, USA). Recombinant Mouse IL1β (575106) was purchased from BioLegend (San Diego, CA, USA).
2.3. In Vitro Studies
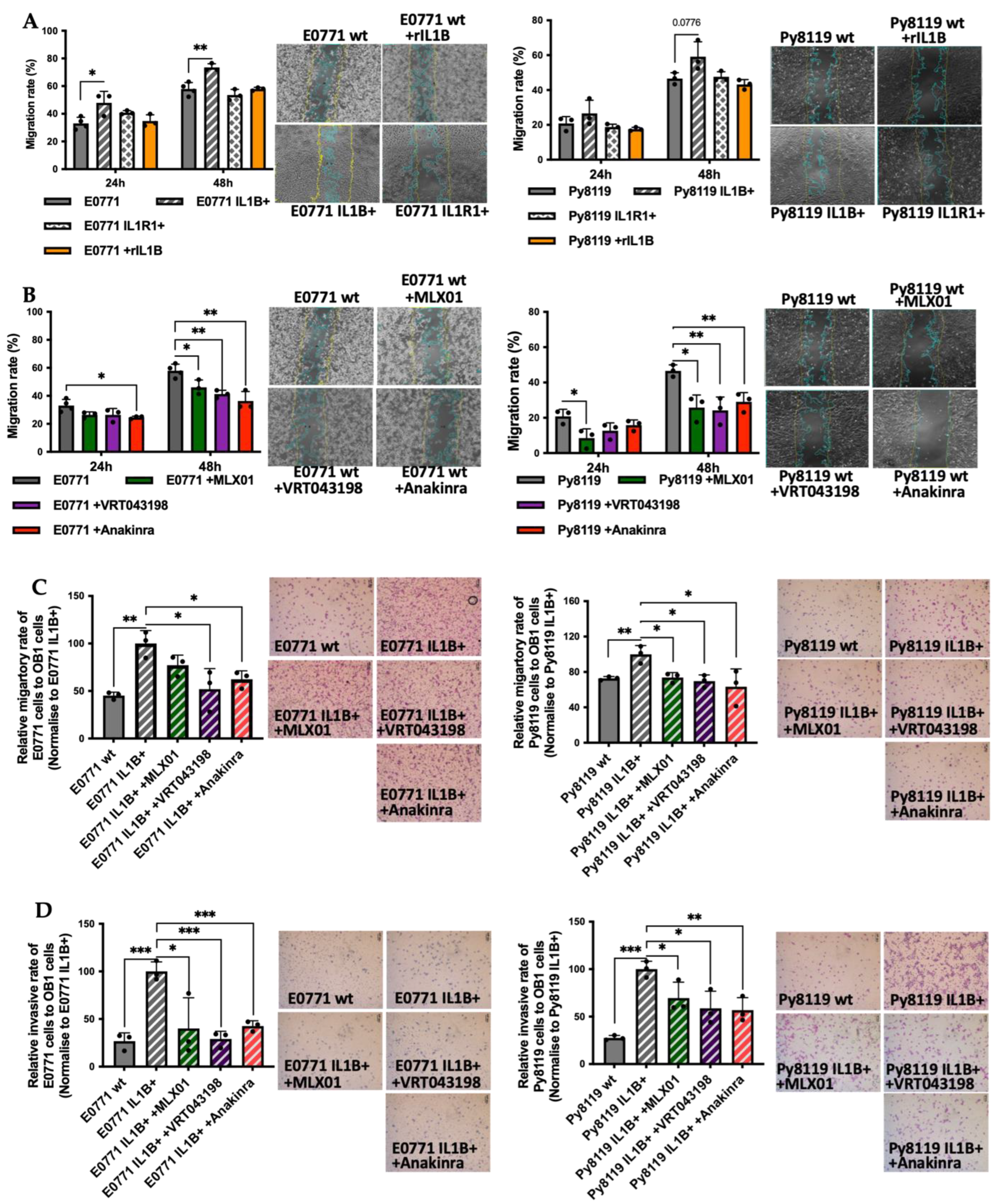
MTT: Cell viability was assessed by MTT according to the manufacturer’s instruction (M5655-1G, Sigma-Aldrich, St. Louis, MI, USA), and the plate was finally read at 570 nm by FlexStation3 Benchtop Multi-Mode Microplate Reader (Molecular Devices, San Jose, CA, USA). Scratch assay: Cell migration was assessed by scratch assay: 100% confluent monolayers of E0771 or Py8119 cells were treated with 10 µg/mL Mitomycin C (M-0503, Sigma-Aldrich) for 3 hours (Mitomycin C serves to inhibit cell proliferation from eliminating the impact of cell division on data analysis from scratch assay and transwell assays.) in a 12-well plate before a scratch was made by a 200µL-tip. Six different fields in each scratch were photographed by EVOS™ FL Auto 2 Imaging System (Fisher Scientific, Waltham, MA, USA) at 0 h, 24 h, and 48 h post the scratch and finally, T-scratch software (ETH Zurich, Zürich, Switzerland) [
20] was used to analyse % of open area in the image. Transwell assay: Two types of transwell inserts were respectively used to assess the migratory and invasive ability of E0771 and Py8119 cells towards pre-osteoblast, OB1 cells, Costar transwell permeable Supports 6.5 mm insert with 8um Polycarbonate Membrane (31019046) for migration and Corning BioCoat Matrigel Invasion insert with 8μm Polycarbonate Membrane (354480) for invasion. Firstly, pre-osteoblast OB1 cells or HUVECs were seeded at 30,000 cells/well in a 24-well-plate for 24 h. The next day, a 70% confluent monolayer of E0771/Py8119 cells was treated with 10 µg/mL Mitomycin C (Sigma-Aldrich, M-0503) for 3 hours before cells were collected and resuspended in 0.1%
w/
v BSA (A7030, Sigma) RPMI 1640 (FBS free) at 1 × 10
5 cells/well. A total of 200 μL of this cell suspension was subsequently added into the transwell insert. This insert was suspended above with OB1 cells following addition of 600 μL, fresh, complete DMEM or with HUVECs in 600 μL endothelial cell media supplied with these cells. After 24 h post cell seeding, un-invaded cells from the top of the membrane of the insert were wiped with a cotton bud before the membrane was fixed in 100% ethanol for 5 min and stained with haematoxylin solution (5 min) and 1%
w/
v eosin (1 min). The membrane was removed from the insert and mounted onto a glass slide using the aqueous mounting medium (S3025, DAKO, Glostrup, Denmark). Finally, the stained membrane was photographed by Leica microscope with Olympus DP73 colour camera (Tokyo, Japan), and the number of cells was counted.
2.4. In Vivo Studies
In vivo procedures were conducted in accordance with local guidelines and with UK Home Office approval under Project License (PPL) 70/8964 or P99922A2E, University of Sheffield, UK. Mice were maintained on a 12 h light:12 h dark cycle with food and water provided ad libitum.
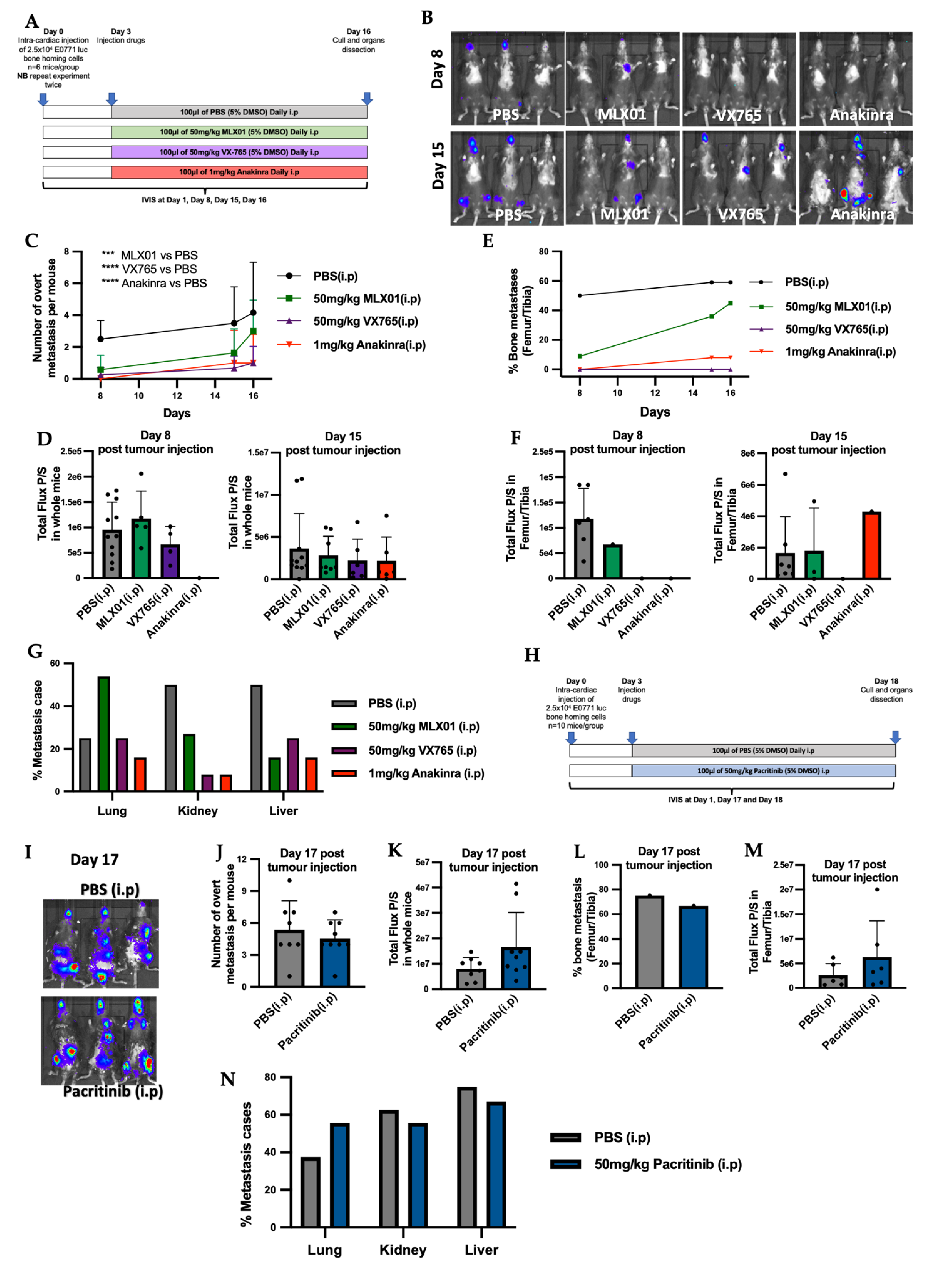
Model of metastatic outgrowth following tumour cell disseminated into metastatic organs (intra-cardiac injection): 2.5 × 104 E0771 luc2 GFP cells were administered via intra-cardiac injection into 5–6-week C57Bl/6J female mice. Five days post tumour cell injection, mice were imaged by IVIS and randomised into groups of equal tumour size. At this point mice received daily intraperitoneal injection of control (5% v/v DMSO/PBS), MLX01 (50 mg/kg), VX765 (50 mg/kg), Anakinra (1 mg/kg), or Pacritinib (50mg/kg) or orally gavage of control (5% v/v DMSO/PBS) or MLX01 (100 mg/kg). Model of spontaneous tumour cells metastasis (intra-ductal injection): 1.25 × 105 E0771 luc2 GFP cells were engrafted via injection into the 4th mammary ducts of 5–6-week C57Bl/6J female mice following subcutaneous injection of 0.002 mg Vetergesic. Seven days post tumour cell injection, when tumour volume reached around 0.027 cm3, mice were randomised into groups of equal tumour volume, and at this point, mice were given daily intraperitoneal injections of control (5% v/v DMSO/PBS), MLX01 (50 mg/kg), VX765 (50 mg/kg), Anakinra (1 mg/kg), respectively. Tumours were measured by digital vernier scale every three days, and mice were culled once their primary tumour reached 1 cm3.
2.5. In Vivo Imaging
Tumours in soft tissue or hind limbs were measured using bioluminescence-based in vivo imaging (IVIS Lumina II in Vivo Imaging System (PerkinElmer, Waltham, MA, USA) and Living Image® 4.5.4 software) 2-, 7-, 15- and 17-days post tumour cell injection, 5 minutes following subcutaneous injection of 6 mg/kg D-Luciferin (Invitrogen, Waltham, MA, USA). Maximum BLI thresholds were set as 5e5 photons, and minimum BLI thresholds were modified across experimental models to allow the visualisation of tumour-derived bioluminescent signals.
2.6. Sample Collection
After dissection and IVIS imaging, half of primary tumours, lungs and both tibias were fixed in 4% w/v PFA/PBS for 48 hours and stored in 70% v/v ethanol/distilled water for uCT analysis. The other half of the tumours and lungs and both femurs were snap-frozen by immersing in liquid nitrogen and then stored at −80 °C. Following blood collection, serum was collected by centrifugation and stored at −80 °C. Spleens were stored at −80 °C in 10% v/v DMSO/ FBS for future flow cytometric analysis.
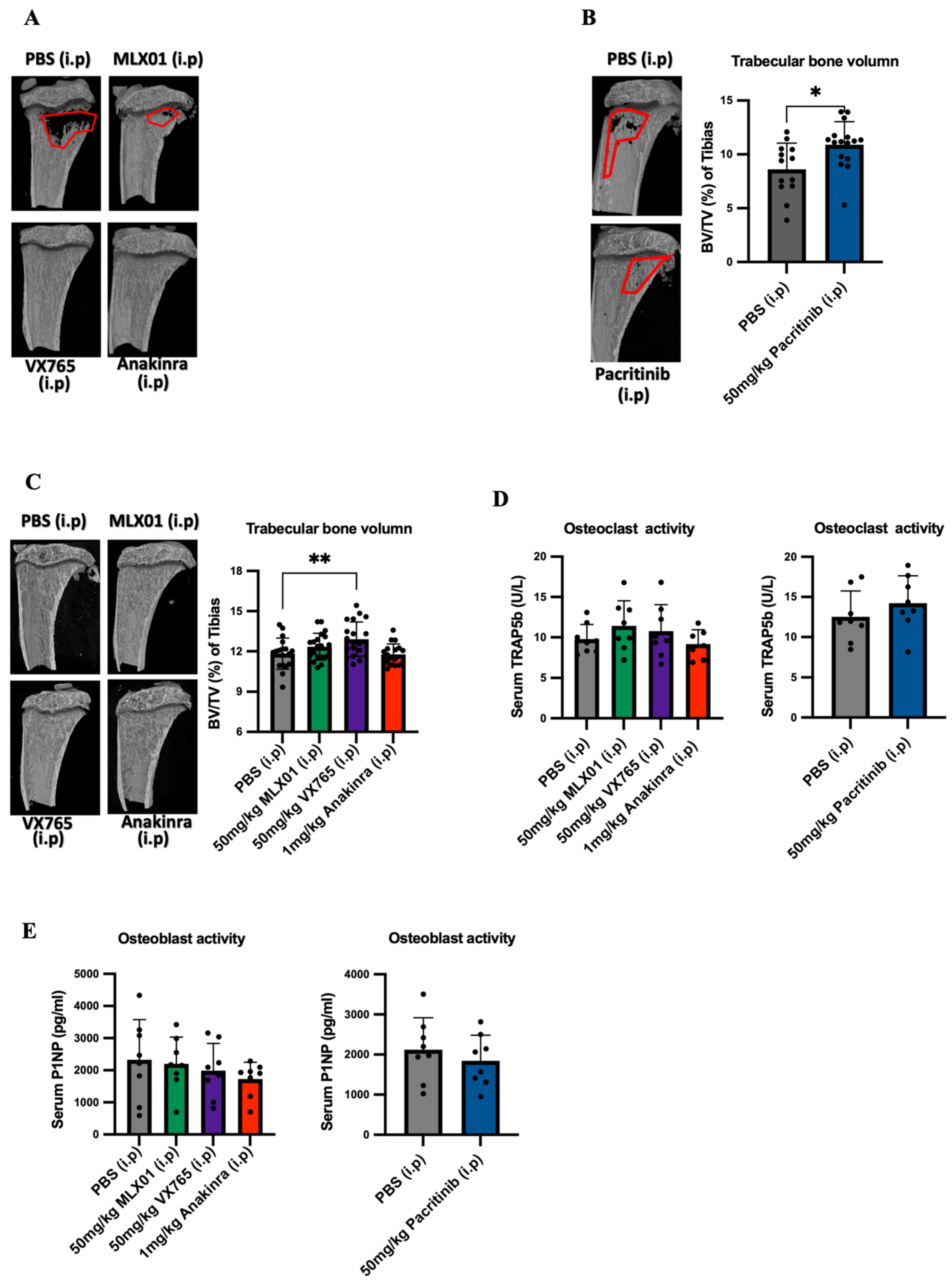
2.7. Microcomputed Tomography Imaging
Microcomputed tomography imaging analyses were carried out via a Skyscan 1172 X-ray–computed μCT scanner (Bruker, Billerica, MA, USA) equipped with the X-ray tube (voltage, 49 kV; current, 200 mA) and the 0.5 mm aluminium filter. Pixel size was set to 4.3 µm, and scanning initiated from the top of the proximal tibia as described previously [
21].
2.8. ELISA
Mouse serum TRAP5b was assessed by MouseTRAP™ (TRAcP 5b) ELISA (SB-TR103), and mouse serum P1NP was assessed by Elabscience P1NP (Procollagen 1 N-Terminal Propeptide) ELISA kit (E-EL-M0233) in accordance with manufacturer instructions. Mouse IL1β was assessed by ELISA MAX™ Deluxe Set Mouse IL1β (432604, BioLegend)
2.9. Flow Cytometry
Frozen spleens were quickly defrosted in a 37 °C water bath and then washed in PBS three times to remove DMSO. Deforested spleens were chopped and digested using Liberase digestion solution (Roche, 0540102001) and then filtered using a 100 µm cell strainer to single cells in 1%
v/
v FBS/PBS (FACS buffer). Samples were aliquoted and incubated with fluorochrome-conjugated antibodies and the viability dye (
Table S1) for 45 min on ice. Next, the cell pellet was resuspended with 0.5ml 1%
v/
v PFA/PBS. Finally, the cell suspension was transferred to a filtered FACS tube on the ice and analysis was performed by BD LSR II flow cytometer and FlowJo software v10.8.0 (Becton, Dickinson and Company, Franklin Lakes, NJ, USA). The gating strategy is shown in
Figures S10 and S11.
2.10. Statistical Analysis
Data analysis was performed using GraphPad Prism 9 (GraphPad Software Inc., San Diego, CA, USA). Data were analysed using unpaired t-tests for single timepoints and two-way ANOVA for time course experiments. All data from in vitro experiments are from a minimum of three repeats from three biological experiments, and their graphs represent mean + SD. All in vivo experiments show all individual points, and their graphs represent mean + SD. Statistical significance was defined as * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
4. Discussion
Our study shows that the small-molecule inhibitor that targets Caspase-1 has promising anti-tumour effects, inhibiting breast cancer bone metastasis: preventing metastatic outgrowth in the bone and reducing spontaneous metastasis to this site to the same extent or better than the IL1R antagonist, Anakinra (
Figure 3E and
Figure 4E), whilst having less impact on immunity (
Figure 6). Moreover, although the IL1β inhibitor MLX01 only slightly reduces breast cancer metastasis to the bone, it significantly delays the onset of bone metastasis and reduces tumour growth at the primary site (
Figure 3E,
Figure 4E and
Figure S6E). In contrast, inhibiting IRAK1 by Pacritinib had small cytotoxic effects in vitro but did not reduce migration or invasion and also failed to inhibit the development of bone metastases in vivo (
Figure 3L).
The expression of the IL-1-related genes, such as
Il1B, CASP1 and
IRAK1, have been identified to be significantly up-regulated in breast cancer compared with normal mammary tissue [
23,
24]. Meanwhile, high expression of IL1β, IRAK1 and Caspase-1 is closely correlated with reduced overall survival and distant recurrence of breast cancer [
3,
23]. Especially, a significant correlation between the expression of IL1β and bone metastasis has been reported: 37% of patients with IL1β-positive primary tumours developed bone metastasis compared with 5% of patients with IL1β-negative primary tumours [
3]. Therefore, targeting these molecules may provide an effective strategy for treating breast cancer bone metastasis. Our previous studies have shown that antibody-mediated IL1β inhibition or genetic ablation of IL1β in mice results in significantly reduced bone metastasis but increased growth of primary breast tumours in xenograft and allograft models, respectively [
2,
13]. An opposite result was demonstrated by Kiss et al., however, who showed delayed primary tumour growth in IL1β−/− (C57BL/6J background) mice after orthotopic injection with E0771 cells [
25]. Similarly, we have previously found that inhibiting IL1R1 with Anakinra reduces subcutaneous tumour growth in BALB/c nude mice, but the same dose did not reduce primary tumour growth in NOD SCID mice [
2,
12]. Other groups have found that knocking out Nlrp3 and Casp1 in mice significantly reduces the growth of primary breast tumours and lung metastases compared with wild-type mice [
26]. In addition, knockdown of IRAK1 in MDA-MB-231 tumour cells has been shown to reduce primary tumour growth in mouse mammary fat pads and subsequent lung metastasis in NOD/SCID mice [
23]. These studies suggest that although IL1β seems to have contrasting effects on the primary tumour, possibly dependent on the immune status of the mouse model used and the stage at which tumour cells are in when denied exposure to IL-1, low activity of IL-1/NFKB is consistently associated with significantly reduced metastasis, especially to the bone.
There are increasing numbers of studies highlighting that small molecule inhibitors of IL-1 may be promising options for the treatment of breast cancer bone metastasis. A small molecule inhibitor of IKK (a downstream target of IL-1) reduced metastases from disseminated MDA-MB-231 cells in the bone by 58% [
27]. Bishop et al. also demonstrated that an IKKe/TRAF6 (downstream target of IL-1) inhibitor could reduce osteoclastic bone destruction from 4T1 tumours growing in the bone; however, this treatment did not significantly reduce the percentage of mice that developed bone metastases [
28,
29]. Taking our new data together with these studies and previous studies showing near elimination of bone metastasis with Canakinumab and Anakinra, inhibiting IL1β or preventing IL1β binding to its receptor IL1R appears to almost prevent metastatic outgrowth of breast cancer in the bone [
2,
9,
12,
13]. However, inhibiting downstream targets of IL-1 (IRAK1, TRAF6, IKK) appears to be less effective, only slightly reducing the percentage of mice that develop the metastatic bone disease but reducing osteoclastic bone destruction. Therefore, we speculate that inhibiting downstream targets of IL-1 (IRAK1, TRAF6, IKK) has effects on reducing osteoclastic bone resorption, while IL1β inhibition not only reduces osteoclastic bone resorption but also blocks breast cancer metastasis to bone. In agreement with this, we have previously shown that inhibiting IL1β prevents metastatic outgrowth in the bone via several mechanisms. Firstly, inhibiting IL1β prevents wnt activation of cancer stem cells in the bone, holding tumour cells in a dormant state in this organ [
9] which might explain why IL1β inhibition is more effective at inhibiting metastatic outgrowth than reducing the growth of overt metastases. Secondly, reducing IL1β in the bone inhibits expansion of the bone metastatic niche (osteoblasts, haematopoietic stem cells and endovascular cells [
2]), which results in tumour cells being held in this site, preventing proliferation into overt metastases. Our μCT data from the current study show increased bone volume in mice following administration of MLX01, VX765, Anakinra and Pacritinib, indicating that these drugs all inhibit bone resorption and osteolysis (
Figure 5A,B and
Figure S8A). In our current study, we found that 13–15 consecutive daily treatments of these drugs had no significant effects on osteoblast or osteoclast activity (
Figure 5D,E). A likely reason for this is that osteoclast/osteoblast activities following intervention with drugs such as Anakinra or other factors that alter bone turnover, including ovariectomy, are commonly seen within a few hours/days, and these then normalise over time [
13,
30]. Therefore, 13–15 days after commencement of treatment may be too late to see the primary effects on these bone-resorbing and forming cells; however, the increased bone volume suggests that these IL-1 targeted drugs cause a net reduction in osteolysis, as previously reported for Anakinra [
13].
In line with IL1β inhibition holding tumours in a dormant state, we found that inhibition of IL1 was effective in reducing bone metastasis or delaying the development of bone metastasis; however, once overt metastases had developed, IL-1 inhibitors were no longer able to reduce tumour growth (
Figure 3F and
Figure S6F). IL1β is a pro-inflammatory cytokine, and inhibiting this molecule has been shown to prevent the infiltration of immune cells into primary tumours, facilitating their growth [
13]. In our models, none of the inhibitors used increased primary tumour growth; in contrast, MLX01 reduced primary tumour growth. However, since VX765 and Anakinra both stimulated the growth of metastatic tumour cells in the lung, we hypothesise that increased tumour growth at this site may be due to a reduced immune response. Our reasoning for this is that we have previously demonstrated that IL1β has opposing effects on tumour growth in the bone compared with soft tissue [
13] and that these differences in pro/anti-tumour effects are driven by differential, site-specific, immunological responses [
13]. It has been postulated that IL1β promotes the recruitment of anti-tumour neutrophils to the lung metastatic niche, inhibiting metastasis to this organ. Once tumour cells are disseminated in soft tissues, inhibiting IL1β signalling with Anakinra has been shown to reduce myeloid cell accumulation in E0771 soft tissue tumours [
16]. IL1β has also been shown to promote infiltration of monocytes and macrophage differentiation in soft tissue tumours [
13,
31]. In addition, in both melanoma and E0771 breast cancer models, IL1β has been shown to increase the anti-tumour potential of T helper (Th1) cells, and genetic knockout of IL1β in mice reduces cytotoxic CD8+ T cells in soft tissue tumours [
13,
18]. In contrast, IL1β appears to play a tumour-supporting function in the bone, and it has previously been demonstrated that IL1β controls myeloid cell number, reduces recruitment of anti-tumour neutrophils to bone and drives mobilisation of immune cells out of the bone marrow [
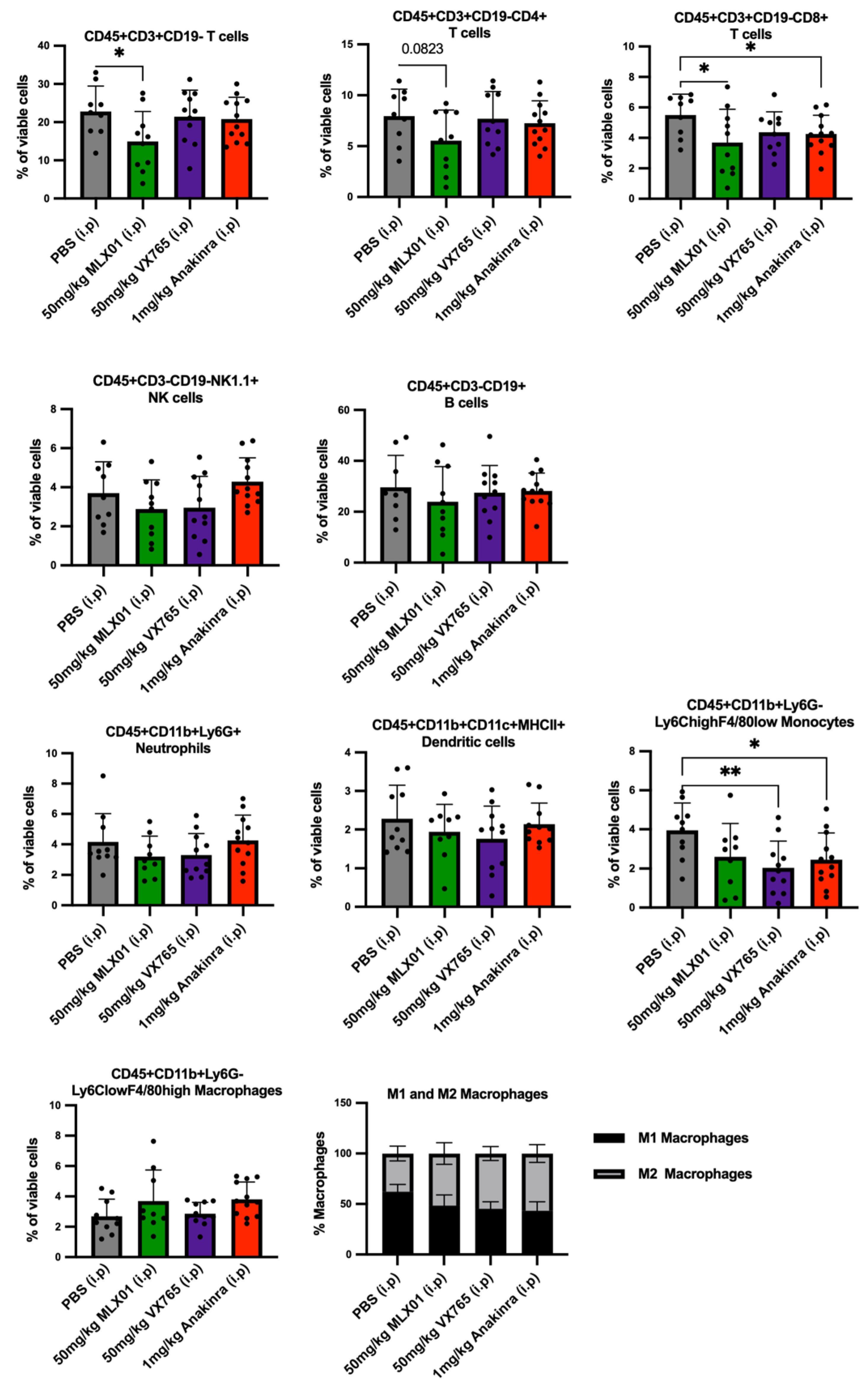
13]. Whilst there is increasing evidence suggesting differential roles of IL1β in immune cell regulation of tumour growth in the bone and soft tissue microenvironments, suitable samples for further analysis of this hypothesis using the small molecule inhibitors tested in the current study were not taken.
Previous studies have demonstrated that inhibiting IL1β significantly reduced breast cancer bone metastasis, but the administration of biological agents such as Canakinumab has severe adverse effects on the immune system of cancer patients [
11]. Therefore, the primary aim of our study was to identify a suitable small molecule inhibitor of the IL-1 signalling pathway that would reduce bone metastasis without seriously impacting immunity. Profiling of immune cells isolated from the spleen demonstrated that similar to previously tested inhibitors of IL-1, MLX01 reduced systemic immune cells, especially cytotoxic CD8+T cells. Whilst this may be the reason we saw a trend towards an increased percentage of mice with lung metastases, additional immune profiling of the lung tissue would be required to confirm this idea. However, despite reduced systemic immunity, MLX01 treatment delayed the growth of primary tumours; this may be because IL1β has been shown to drive both anti-tumour and pro-tumour effects via stimulating infiltration of innate anti-tumour immune cells and creating an immuno-suppressive microenvironment by increasing the infiltration of myeloid-derived suppressor cells (MDSCs), [
10,
32,
33]. Additional experiments would be required to confirm the apparent anti-tumour effects of this drug; however, its limited anti-tumour capacity and heightened adverse effects on the immune system suggest that this may not be the optimal treatment to take forward. In accordance with the published literature, administration of Anakinra suppressed both cytotoxic T cells and monocytes, potentially explaining why when tumours did grow following treatment with this drug, they grew significantly larger than tumours treated with PBS, MLX01 or VX765 [
13]. Importantly for our study, the small molecule inhibitor that was most effective at inhibiting bone metastasis, VX765, had the least effects on systemic immunity, slightly reducing monocytes, suggesting that this drug may be the best option for moving forwards into clinical trials for the prevention of future relapse in the bone.
To date, data from this current study and our previous studies [
2,
13] suggest that inhibiting IL-1 signalling is extremely effaceable against bone metastasis, inhibiting metastatic outgrowth and slowing the development of tumours in the bone; however, the same treatments appear to have the potential to increase metastatic outgrowth of tumour cells disseminated in lungs. It must be noted, however, that lung metastasis from mammary cancer is significantly more prevalent in syngeneic mouse models compared with human patients; therefore, increased lung metastasis observed in mice may not translate into humans [
34,
35,
36]. Despite relatively low lung metastasis from breast cancer in humans, care must be taken not to stimulate this. In breast cancer patients, tumour cell dissemination into metastatic organs is believed to be an early event, often occurring before the diagnosis of the primary disease [
37]. Therefore, whilst preventing the outgrowth of tumour cells disseminated in the bone is desirable, it is important that this is not performed at the expense of increasing metastatic outgrowth in the lung. Emerging studies suggest that this problem can be overcome by combining anti-IL-1 treatments with immune stimulatory chemotherapies and/or immune checkpoint inhibitors [
13,
31]. The reason for this is that IL1β is believed to exert its anti-tumour effects in the bone by inhibiting the expansion of the metastatic niche, bone resorption and the vicious cycle of bone metastasis, whereas IL1β promotes lung metastasis via reducing anti-tumour immune response [
2,
3,
7,
10,
13]. Therefore, giving an anti-IL-1 treatment along with an immune stimulatory drug enables the IL-1 targeted therapy to exert anti-tumour effects in the bone without adversely affecting tumour cells disseminated at other sites. There is emerging evidence that this idea may be true; previous studies have demonstrated that combining Anakinra and the immune stimulatory chemotherapy, doxorubicin, reduces primary tumour growth and almost eliminates bone metastasis by exerting opposing effects on immune regulation [
13], and a separate study demonstrated that combining Anakinra with PD-1 inhibitors resulted in significant, synergistic, anti-cancer effects in a mouse model of soft tissue tumour growth [
31]. Our new data suggest that targeting IL-1 signalling upstream at Caspase-1 with VX765 is even more potent at reducing bone metastasis than Anakinra; we, therefore, suggest further experiments focused on combining VX765 with immune stimulatory therapies before considering future clinical trials.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





