

Research Trend and Detailed Insights into the Molecular Mechanisms of Food Bioactive Compounds against Cancer: A Comprehensive Review with Special Emphasis on Probiotics
 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
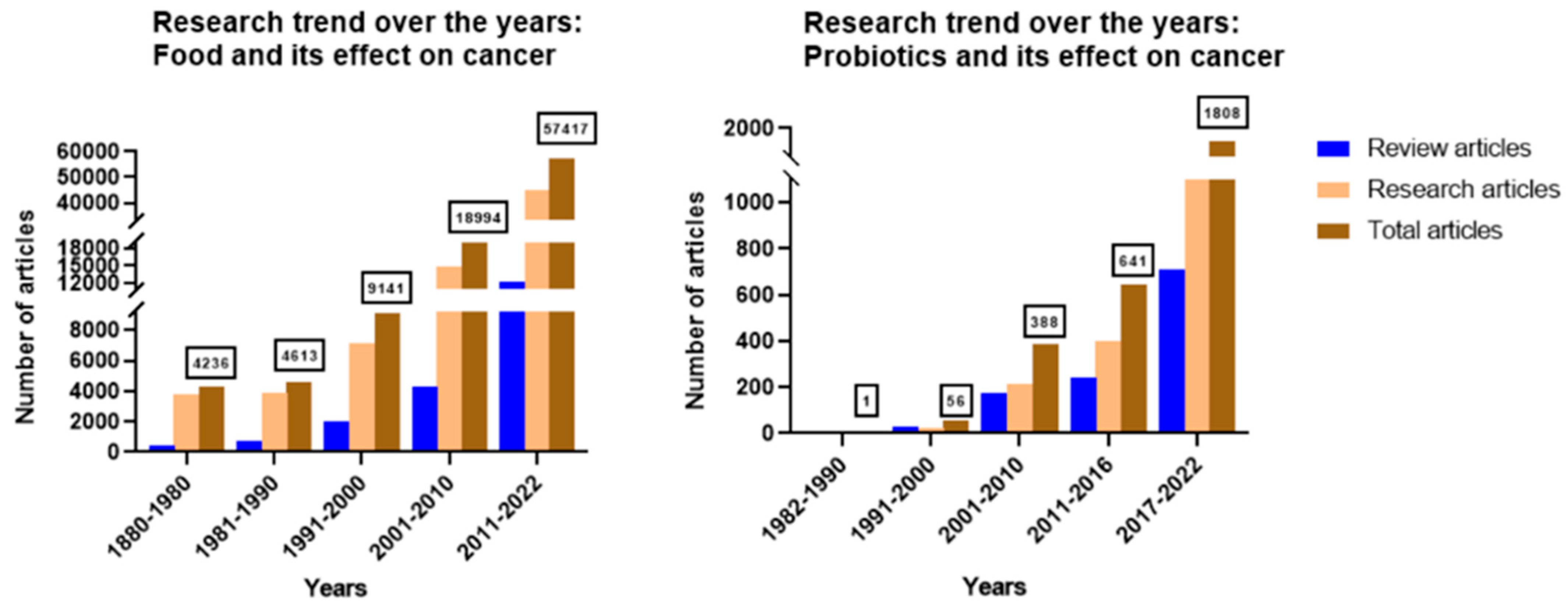
1. Introduction
2. Plant-Based Food Components
2.1. Peppers and Other Nightshade Vegetables
2.1.1. Black Pepper (Piper nigrum)
Piperine
Piperidine
2.1.2. Long Pepper (Piper longum)
2.1.3. Chili Pepper (Capsicum annuum)
2.1.4. Other Nightshade Vegetables
Eggplant (Solanum melongena)
Tomato (Solanum lycopersicum)
2.2. Spices
2.2.1. Cloves (Syzygium aromaticum)
2.2.2. Turmeric (Curcuma longa)
2.3. Cruciferous Vegetables
2.3.1. Isothiocyanates (ITC)
Benzyl Isothiocyanate (BITC)
Phenethyl Isothiocyanate (PEITC)
Sulforaphane (SFN)
2.3.2. Diindolylmethane (DIM)
2.4. Cucurbitaceous Foods
2.5. Root Vegetables
2.5.1. Ginger (Zingiber officinalis)
2.5.2. Garlic (Allium sativum) and Onion (Allium sepa)
2.5.3. Beetroot (Beta vulgaris Subsp. vulgaris)
2.6. Tropical Fruits
2.6.1. Guava (Psidium guajava)
2.6.2. Dragon Fruit (Selenicereus undatus)
2.7. Grass Family Members:
2.7.1. Lemongrass (Cymbopogon citratus)
2.7.2. Wheatgrass (Thinopyrum intermedium)
2.8. Caffeinated Plants (Tea (Camellia sinensis) and Coffee (Coffea arabica))
2.9. Other Plant-Based Foods
3. Probiotics in Cancer
4. Combination Strategies—The Therapeutic Opportunity
4.1. Combination of Two or More Food Bioactive Compounds
4.2. Combination of Food Bioactive Compounds with Other Physiologically Active Phytochemical/s
4.3. Combination of Food Bioactive Compound along with Chemotherapeutic Agents
5. Novel Strategies for Enhanced Delivery of Natural Food Bioactive Agents for Cancer Chemoprevention
6. Discussion
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Note
Appendix A
References
- What Is Cancer? Available online: https://www.cancer.org/treatment/understanding-your-diagnosis/what-is-cancer.html (accessed on 6 January 2022).
- Gaikwad, S.; Srivastava, S.K. Role of Phytochemicals in Perturbation of Redox Homeostasis in Cancer. Antioxidants 2021, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Rayburn, E.R.; Ezell, S.; Zhang, R. Anti-Inflammatory Agents for Cancer Therapy. Mol. Cell. Pharmacol. 2009, 1, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, I.; Srivastava, S.K. GABAA receptor agonist suppresses pediatric medulloblastoma progression by inhibiting PKA-Gli1 signaling axis. Mol. Ther. 2022, 30, 2584–2602. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gaikwad, S.; Kaushik, I.; Wright, S.; Markiewski, M.; Srivastava, S. Atovaquone Suppresses Triple-Negative Breast Tumor Growth by Reducing Immune-Suppressive Cells. Int. J. Mol. Sci. 2021, 22, 5150. [Google Scholar] [CrossRef]
- Ramachandran, S.; Srivastava, S.K. Repurposing Pimavanserin, an Anti-Parkinson Drug for Pancreatic Cancer Therapy. Mol. Ther.-Oncolytics 2020, 19, 19–32. [Google Scholar] [CrossRef]
- Yadav, A.; Alnakhli, A.; Vemana, H.P.; Bhutkar, S.; Muth, A.; Dukhande, V.V. Repurposing an Antiepileptic Drug for the Treatment of Glioblastoma. Pharm. Res. 2022, 39, 2871–2883. [Google Scholar] [CrossRef]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [Green Version]
- Bang, J.S.; Oh, D.H.; Choi, H.M.; Sur, B.J.; Lim, S.J.; Kim, J.Y.; Yang, H.I.; Yoo, M.C.; Hahm, D.H.; Kim, K.S. Anti-inflammatory and antiarthritic effects of piperine in human interleukin 1β-stimulated fibroblast-like synoviocytes and in rat arthritis models. Arthritis Res. Ther. 2009, 11, R49. [Google Scholar] [CrossRef] [Green Version]
- Kapadia, G.J.; Tokuda, H.; Konoshima, T.; Nishino, H. Chemoprevention of lung and skin cancer by Beta vulgaris (beet) root extract. Cancer Lett. 1996, 100, 211–214. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Kim, S.-H.; Srivastava, S.K. Piperine Causes G1 Phase Cell Cycle Arrest and Apoptosis in Melanoma Cells through Checkpoint Kinase-1 Activation. PLoS ONE 2014, 9, e94298. [Google Scholar] [CrossRef]
- Huovinen, M.; Loikkanen, J.; Myllynen, P.; Vähäkangas, K.H. Characterization of human breast cancer cell lines for the studies on p53 in chemical carcinogenesis. Toxicol. In Vitro 2011, 25, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Yang, R.; Lin, R.; Yang, S. Piperine functions as a tumor suppressor for human ovarian tumor growth via activation of JNK/p38 MAPK-mediated intrinsic apoptotic pathway. Biosci. Rep. 2018, 38, BSR20180503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Khoi, P.N.; Yoon, H.J.; Lian, S.; Joo, Y.E.; Chay, K.O.; Kim, K.K.; Jung, Y.D. Piperine inhibits IL-1β-induced IL-6 expression by suppressing p38 MAPK and STAT3 activation in gastric cancer cells. Mol. Cell. Biochem. 2014, 398, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, S.; Caldon, E.C.; Tilley, W.; Wang, S. Cyclin-Dependent Kinase 2 Inhibitors in Cancer Therapy: An Update. J. Med. Chem. 2019, 62, 4233–4251. [Google Scholar] [CrossRef]
- Jeong, S.; Jung, S.; Park, G.-S.; Shin, J.; Oh, J.-W. Piperine synergistically enhances the effect of temozolomide against temozolomide-resistant human glioma cell lines. Bioengineered 2020, 11, 791–800. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Liao, H.; Li, L.; Pan, L. Piperine induces apoptosis of lung cancer A549 cells via p53-dependent mitochondrial signaling pathway. Tumor Biol. 2014, 35, 3305–3310. [Google Scholar] [CrossRef]
- Siddiqui, S.; Ahamad, S.; Jafri, A.; Afzal, M. Arshad Piperine Triggers Apoptosis of Human Oral Squamous Carcinoma Through Cell Cycle Arrest and Mitochondrial Oxidative Stress. Nutr. Cancer 2017, 69, 791–799. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Chaudhary, A.K.; Yadav, N.; Bhat, T.A.; O’Malley, J.; Kumar, S.; Chandra, D. A potential role of X-linked inhibitor of apoptosis protein in mitochondrial membrane permeabilization and its implication in cancer therapy. Drug Discov. Today 2016, 21, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Gnanasekar, M.; Thirugnanam, S.; Zheng, G.; Chen, A.; Ramaswamy, K. Gene silencing of translationally controlled tumor protein (TCTP) by siRNA inhibits cell growth and induces apoptosis of human prostate cancer cells. Int. J. Oncol. 2009, 34, 1241–1246. [Google Scholar] [CrossRef]
- Guo, S.; Sun, F.; Guo, Z.; Li, W.; Alfano, A.; Chen, H.; Magyar, C.E.; Huang, J.; Chai, T.; Qiu, S.; et al. Tyrosine Kinase ETK/BMX Is Up-Regulated in Bladder Cancer and Predicts Poor Prognosis in Patients with Cystectomy. PLoS ONE 2011, 6, e17778. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Page, C.; Reynolds, R.; Lin, J. Constitutive Activation of Stat 3 Oncogene Product in Human Ovarian Carcinoma Cells. Gynecol. Oncol. 2000, 79, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Samykutty, A.; Shetty, A.V.; Dakshinamoorthy, G.; Bartik, M.M.; Johnson, G.L.; Webb, B.; Zheng, G.; Chen, A.; Kalyanasundaram, R.; Munirathinam, G. Piperine, a Bioactive Component of Pepper Spice Exerts Therapeutic Effects on Androgen Dependent and Androgen Independent Prostate Cancer Cells. PLoS ONE 2013, 8, e65889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaffe, P.B.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine impairs cell cycle progression and causes reactive oxygen species-dependent apoptosis in rectal cancer cells. Exp. Mol. Pathol. 2013, 94, 109–114. [Google Scholar] [CrossRef]
- Sherr, C.J.; Roberts, J.M. CDK inhibitors: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Loo, G. Redox-sensitive mechanisms of phytochemical-mediated inhibition of cancer cell proliferation (review). J. Nutr. Biochem. 2003, 14, 64–73. [Google Scholar] [CrossRef]
- Han, S.-Z.; Liu, H.-X.; Yang, L.-Q.; Cui, L.-D.; Xu, Y. Piperine (PP) enhanced mitomycin-C (MMC) therapy of human cervical cancer through suppressing Bcl-2 signaling pathway via inactivating STAT3/NF-κB. Biomed. Pharmacother. 2017, 96, 1403–1410. [Google Scholar] [CrossRef]
- Li, N.; Wen, S.; Chen, G.; Wang, S. Antiproliferative potential of piperine and curcumin in drug-resistant human leukemia cancer cells are mediated via autophagy and apoptosis induction, S-phase cell cycle arrest and inhibition of cell invasion and migration. J. BUON Off. J. Balk. Union Oncol. 2021, 26, 1181. [Google Scholar]
- Zhang, X.; Li, X.-R.; Zhang, J. Current status and future perspectives of PI3K and mTOR inhibitor as anticancer drugs in breast cancer. Curr. Cancer Drug Targets 2013, 13, 175–187. [Google Scholar] [CrossRef]
- Greenshields, A.L.; Doucette, C.D.; Sutton, K.M.; Madera, L.; Annan, H.; Yaffe, P.B.; Knickle, A.F.; Dong, Z.; Hoskin, D.W. Piperine inhibits the growth and motility of triple-negative breast cancer cells. Cancer Lett. 2015, 357, 129–140. [Google Scholar] [CrossRef]
- Twiddy, D.; Brown, D.G.; Adrain, C.; Jukes, R.; Martin, S.J.; Cohen, G.M.; MacFarlane, M.; Cain, K. Pro-apoptotic Proteins Released from the Mitochondria Regulate the Protein Composition and Caspase-processing Activity of the Native Apaf-1/Caspase-9 Apoptosome Complex. J. Biol. Chem. 2004, 279, 19665–19682. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.C.; Wilkinson, A.S.; Scott, F.L.; Csomos, R.A.; Salvesen, G.S.; Duckett, C.S. Neutralization of Smac/Diablo by Inhibitors of Apoptosis (IAPs). A caspase-independent mechanism for apoptotic inhibition. J. Biol. Chem. 2004, 279, 51082–51090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Li, K.; Zou, Z.; Li, L.; Zhu, L.; Wang, Q.; Gao, W.; Wang, Y.; Huang, W.; Liu, R.; et al. Piperidine nitroxide Tempol enhances cisplatin-induced apoptosis in ovarian cancer cells. Oncol. Lett. 2018, 16, 4847–4854. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-T.; Lin, B.-R.; Chang, C.-C.; Chu, C.-Y.; Su, H.-J.; Chen, S.-T.; Jeng, Y.-M.; Kuo, M.-L. IL-6 induces AGS gastric cancer cell invasionvia activation of the c-Src/RhoA/ROCK signaling pathway. Int. J. Cancer 2007, 120, 2600–2608. [Google Scholar] [CrossRef] [PubMed]
- Łukaszewicz-Zajac, M.; Mroczko, B.; Szmitkowski, M. The role of interleukin-6 and C-reactive protein in gastric cancer. Polski Merkur. Lek. Organ Polskiego Towar. Lek. 2010, 29, 382–386. [Google Scholar]
- Johnson, G.L.; Lapadat, R. Mitogen-Activated Protein Kinase Pathways Mediated by ERK, JNK, and p38 Protein Kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [Green Version]
- Cahill, C.M.; Rogers, J.T. Interleukin (IL) 1β Induction of IL-6 Is Mediated by a Novel Phosphatidylinositol 3-Kinase-dependent AKT/IκB Kinase α Pathway Targeting Activator Protein. J. Biol. Chem. 2008, 283, 25900–25912. [Google Scholar] [CrossRef] [Green Version]
- Karpinich, N.O.; Tafani, M.; Rothman, R.J.; Russo, M.A.; Farber, J.L. The Course of Etoposide-induced Apoptosis from Damage to DNA and p53 Activation to Mitochondrial Release of Cytochromec. J. Biol. Chem. 2002, 277, 16547–16552. [Google Scholar] [CrossRef] [Green Version]
- Bunz, F.; Dutriaux, A.; Lengauer, C.; Waldman, T.; Zhou, S.; Brown, J.P.; Sedivy, J.M.; Kinzler, K.W.; Vogelstein, B. Requirement for p53 and p21 to Sustain G2 Arrest After DNA Damage. Science 1998, 282, 1497–1501. [Google Scholar] [CrossRef]
- Banerjee, S.; Katiyar, P.; Kumar, V.; Saini, S.S.; Varshney, R.; Krishnan, V.; Sircar, D.; Roy, P. Black pepper and piperine induce anticancer effects on leukemia cell line. Toxicol. Res. 2021, 10, 169–182. [Google Scholar] [CrossRef]
- He, G.; Yu, G.-Y.; Temkin, V.; Ogata, H.; Kuntzen, C.; Sakurai, T.; Sieghart, W.; Peck-Radosavljevic, M.; Leffert, H.L.; Karin, M. Hepatocyte IKKβ/NF-κB Inhibits Tumor Promotion and Progression by Preventing Oxidative Stress-Driven STAT3 Activation. Cancer Cell 2010, 17, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, G.C.; Oliveira, L.F.S.; Predes, D.; Fokoue, H.H.; Kuster, R.M.; Oliveira, F.L.; Mendes, F.A.; Abreu, J.G. Piperine suppresses the Wnt/β-catenin pathway and has anti-cancer effects on colorectal cancer cells. Sci. Rep. 2020, 10, 11681. [Google Scholar] [CrossRef] [PubMed]
- Arun, A.; Ansari, M.; Popli, P.; Jaiswal, S.; Mishra, A.; Dwivedi, A.; Hajela, K.; Konwar, R. New piperidine derivative DTPEP acts as dual-acting anti-breast cancer agent by targeting ERα and downregulating PI3K/Akt-PKCα leading to caspase-dependent apoptosis. Cell Prolif. 2018, 51, e12501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.-J.; Liu, S.-M.; Yang, J.-J.; Li, J. Novel piperidine derivatives as colchicine binding site inhibitors induce apoptosis and inhibit epithelial-mesenchymal transition against prostate cancer PC3 cells. J. Enzym. Inhib. Med. Chem. 2020, 35, 1403–1413. [Google Scholar] [CrossRef]
- Yaffe, P.B.; Coombs, M.R.P.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1 arrest and apoptosis triggered by endoplasmic reticulum stress. Mol. Carcinog. 2015, 54, 1070–1085. [Google Scholar] [CrossRef]
- Wang, J.; Li, B.; Zhao, K.; Su, X. 2-Amino-4-(1-piperidine) pyridine exhibits inhibitory effect on colon cancer through suppression of FOXA2 expression. 3 Biotech 2019, 9, 384. [Google Scholar] [CrossRef]
- Golovine, K.; Makhov, P.; Naito, S.; Raiyani, H.; Tomaszewski, J.; Mehrazin, R.; Tulin, A.; Kutikov, A.; Uzzo, R.G.; Kolenko, V.M. Piperlongumine and its analogs down-regulate expression of c-Met in renal cell carcinoma. Cancer Biol. Ther. 2015, 16, 743–749. [Google Scholar] [CrossRef]
- Conde, J.; Pumroy, R.A.; Baker, C.; Rodrigues, T.; Guerreiro, A.; Sousa, B.B.; Marques, M.C.; de Almeida, B.P.; Lee, S.; Leites, E.P.; et al. Allosteric Antagonist Modulation of TRPV2 by Piperlongumine Impairs Glioblastoma Progression. ACS Cent. Sci. 2021, 7, 868–881. [Google Scholar] [CrossRef]
- Harshbarger, W.; Gondi, S.; Ficarro, S.B.; Hunter, J.; Udayakumar, D.; Gurbani, D.; Singer, W.D.; Liu, Y.; Li, L.; Marto, J.A.; et al. Structural and Biochemical Analyses Reveal the Mechanism of Glutathione S-Transferase Pi 1 Inhibition by the Anti-cancer Compound Piperlongumine. J. Biol. Chem. 2017, 292, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Hoch-Ligeti, C. Production of liver tumours by dietary means; effect of feeding chilies [Capsicum frutescens and annuum (Linn.)] to rats. Acta-Unio Int. Contra Cancrum 1951, 7, 606–611. [Google Scholar]
- Yang, Y.; Zhang, J.; Weiss, N.S.; Guo, L.; Zhang, L.; Jiang, Y.; Yang, Y. The consumption of chili peppers and the risk of colorectal cancer: A matched case-control study. World J. Surg. Oncol. 2019, 17, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Humphreys, I.; Sahu, R.P.; Shi, Y.; Srivastava, S.K. In vitro and in vivo induction of apoptosis by capsaicin in pancreatic cancer cells is mediated through ROS generation and mitochondrial death pathway. Apoptosis 2008, 13, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.C.; Boreddy, S.R.; Srivastava, S.K. Role of Mitochondrial Electron Transport Chain Complexes in Capsaicin Mediated Oxidative Stress Leading to Apoptosis in Pancreatic Cancer Cells. PLoS ONE 2011, 6, e20151. [Google Scholar] [CrossRef]
- Bai, H.; Li, H.; Zhang, W.; Matkowskyj, K.A.; Liao, J.; Srivastava, S.K.; Yang, G.-Y. Inhibition of chronic pancreatitis and pancreatic intraepithelial neoplasia (PanIN) by capsaicin in LSL-KrasG12D/Pdx1-Cre mice. Carcinogenesis 2011, 32, 1689–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pramanik, K.C.; Srivastava, S.K. Apoptosis Signal-Regulating Kinase 1–Thioredoxin Complex Dissociation by Capsaicin Causes Pancreatic Tumor Growth Suppression by Inducing Apoptosis. Antioxid. Redox Signal. 2012, 17, 1417–1432. [Google Scholar] [CrossRef] [Green Version]
- Reilly, C.A.; Henion, F.; Bugni, T.S.; Ethirajan, M.; Stockmann, C.; Pramanik, K.C.; Srivastava, S.K.; Yost, G.S. Reactive Intermediates Produced from the Metabolism of the Vanilloid Ring of Capsaicinoids by P450 Enzymes. Chem. Res. Toxicol. 2013, 26, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Boreddy, S.R.; Srivastava, S.K. Pancreatic cancer chemoprevention by phytochemicals. Cancer Lett. 2013, 334, 86–94. [Google Scholar] [CrossRef]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and clinical uses of capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef]
- Oyagbemi, A.; Saba, A.B.; I Azeez, O. Capsaicin: A novel chemopreventive molecule and its underlying molecular mechanisms of action. Indian J. Cancer 2010, 47, 53–58. [Google Scholar] [CrossRef]
- Islam, A.; Yang, Y.T.; Wu, W.H.; Chueh, P.J.; Lin, M.H. Capsaicin attenuates cell migration via SIRT1 targeting and inhibition to enhance cortactin and beta-catenin acetylation in bladder cancer cells. Am. J. Cancer Res. 2019, 9, 1172–1182. [Google Scholar]
- Afshari, F.S.H.; Hashemi, Z.S.; Timajchi, M.; Ensiyeh, O.; Ladan, G.; Asadi, M.; Elyasi, Z.; GanjiBakhsh, M. The Cytotoxic Effects of Eggplant Peel Extract on Human Gastric Adenocarcinoma Cells and Normal Cells. Mod. Med. Lab. J. 2017, 1, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-R.; Kozukue, N.; Han, J.-S.; Park, J.-H.; Chang, E.-Y.; Baek, E.-J.; Chang, J.-S.; Friedman, M. Glycoalkaloids and Metabolites Inhibit the Growth of Human Colon (HT29) and Liver (HepG2) Cancer Cells. J. Agric. Food Chem. 2004, 52, 2832–2839. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Sasaki, K.; Kito, H.; Haga, A.; Sato, T. Inhibitory Effect of Delphinidin from Solanum melongena on Human Fibrosarcoma HT-1080 Invasiveness in vitro. Planta Med. 1998, 64, 216–219. [Google Scholar] [CrossRef]
- Shen, K.-H.; Hung, J.-H.; Chang, C.-W.; Weng, Y.-T.; Wu, M.-J.; Chen, P.-S. Solasodine inhibits invasion of human lung cancer cell through downregulation of miR-21 and MMPs expression. Chem. Biol. Interact. 2017, 268, 129–135. [Google Scholar] [CrossRef]
- Zhao, B.; Tomoda, Y.; Mizukami, H.; Makino, T. 9-Oxo-(10E,12E)-octadecadienoic acid, a cytotoxic fatty acid ketodiene isolated from eggplant calyx, induces apoptosis in human ovarian cancer (HRA) cells. J. Nat. Med. 2015, 69, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Fekry, M.; Ezzat, S.M.; Salama, M.M.; AlShehri, O.Y.; Al-Abd, A.M. Bioactive glycoalkaloides isolated from Solanum melongena fruit peels with potential anticancer properties against hepatocellular carcinoma cells. Sci. Rep. 2019, 9, 1746. [Google Scholar] [CrossRef] [Green Version]
- Fujimaki, J.; Sayama, N.; Shiotani, S.; Suzuki, T.; Nonaka, M.; Uezono, Y.; Oyabu, M.; Kamei, Y.; Nukaya, H.; Wakabayashi, K.; et al. The Steroidal Alkaloid Tomatidine and Tomatidine-Rich Tomato Leaf Extract Suppress the Human Gastric Cancer-Derived 85As2 Cells In Vitro and In Vivo via Modulation of Interferon-Stimulated Genes. Nutrients 2022, 14, 1023. [Google Scholar] [CrossRef]
- Chen, X.; Yang, G.; Liu, M.; Quan, Z.; Wang, L.; Luo, C.; Wu, X.; Zheng, Y. Lycopene enhances the sensitivity of castration-resistant prostate cancer to enzalutamide through the AKT/EZH2/androgen receptor signaling pathway. Biochem. Biophys. Res. Commun. 2022, 613, 53–60. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Mukhtar, H. Cancer Chemoprevention Through Dietary Antioxidants: Progress and Promise. Antioxid. Redox Signal. 2008, 10, 475–510. [Google Scholar] [CrossRef]
- Gupta, P.; Bhatia, N.; Bansal, M.P.; Koul, A. Lycopene modulates cellular proliferation, glycolysis and hepatic ultrastructure during hepatocellular carcinoma. World J. Hepatol. 2016, 8, 1222–1233. [Google Scholar] [CrossRef]
- Palozza, P.; Simone, R.E.; Catalano, A.; Mele, M.C. Tomato Lycopene and Lung Cancer Prevention: From Experimental to Human Studies. Cancers 2011, 3, 2333–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, M.J.; Das Gupta, S.; Wahler, J.; Suh, N. Role of dietary bioactive natural products in estrogen receptor-positive breast cancer. Semin. Cancer Biol. 2016, 40–41, 170–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowles, J.; Smith, J.W.; Applegate, C.C.; Miller, R.J.; A Wallig, M.; Kaur, A.; Sarol, J.N.; Musaad, S.; Clinton, S.K.; O’Brien, W.D.; et al. Dietary Tomato or Lycopene Do Not Reduce Castration-Resistant Prostate Cancer Progression in a Murine Model. J. Nutr. 2020, 150, 1808–1817. [Google Scholar] [CrossRef]
- E Moran, N.; Thomas-Ahner, J.M.; Wan, L.; E Zuniga, K.; Erdman, J.W.; Clinton, S.K. Tomatoes, Lycopene, and Prostate Cancer: What Have We Learned from Experimental Models? J. Nutr. 2022, 152, 1381–1403. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.S.; Febriyanti, R.M.; Sofyan, F.F.; E Luftimas, D.; Abdulah, R. Anticancer potential of Syzygium aromaticum L. in MCF-7 human breast cancer cell lines. Pharmacogn. Res. 2014, 6, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, V.; Shrivastava, R.; Hussain, S.; Ganguly, C.; Bharadwaj, M. Comparative anticancer potential of clove (Syzygium aromaticum)—An Indian spice—Against cancer cell lines of various anatomical origin. Asian Pac. J. Cancer Prev. 2011, 12, 1989–1993. [Google Scholar]
- Bhamarapravati, S.; Pendland, S.L.; Mahady, G.B. Extracts of spice and food plants from Thai traditional medicine inhibit the growth of the human carcinogen Helicobacter pylori. In Vivo 2003, 17, 541–544. [Google Scholar]
- Liu, H.; Schmitz, J.C.; Wei, J.; Cao, S.; Beumer, J.H.; Strychor, S.; Cheng, L.; Liu, M.; Wang, C.; Wu, N.; et al. Clove Extract Inhibits Tumor Growth and Promotes Cell Cycle Arrest and Apoptosis. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2014, 21, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Kubatka, P.; Uramova, S.; Kello, M.; Kajo, K.; Kruzliak, P.; Mojzis, J.; Vybohova, D.; Adamkov, M.; Jasek, K.; Lasabova, Z.; et al. Antineoplastic effects of clove buds (Syzygium aromaticum L.) in the model of breast carcinoma. J. Cell. Mol. Med. 2017, 21, 2837–2851. [Google Scholar] [CrossRef]
- Li, C.; Xu, H.; Chen, X.; Chen, J.; Li, X.; Qiao, G.; Tian, Y.; Yuan, R.; Su, S.; Liu, X.; et al. Aqueous extract of clove inhibits tumor growth by inducing autophagy through AMPK/ULK pathway. Phytother. Res. 2019, 33, 1794–1804. [Google Scholar] [CrossRef]
- Nirmala, M.J.; Durai, L.; Gopakumar, V.; Nagarajan, R. Anticancer and antibacterial effects of a clove bud essential oil-based nanoscale emulsion system. Int. J. Nanomed. 2019, 14, 6439–6450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, R.P.; Batra, S.; Srivastava, S.K. Activation of ATM/Chk1 by curcumin causes cell cycle arrest and apoptosis in human pancreatic cancer cells. Br. J. Cancer 2009, 100, 1425–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Hu, R.; Dai, A. Curcumin Increased the Sensitivity of Non-Small-Cell Lung Cancer to Cisplatin through the Endoplasmic Reticulum Stress Pathway. Evid.-Based Complement. Altern. Med. 2022, 2022, 6886366. [Google Scholar] [CrossRef]
- Liu, W.; Wang, J.; Zhang, C.; Bao, Z.; Wu, L. Curcumin nanoemulsions inhibit oral squamous cell carcinoma cell proliferation by PI3K/Akt/mTOR suppression and miR-199a upregulation: A preliminary study. Oral Dis. 2022. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Y.; Li, L.; Xie, S. Curcumin Inhibits Papillary Thyroid Cancer Cell Proliferation by Regulating lncRNA LINC. Anal. Cell. Pathol. 2022, 2022, 5946670. [Google Scholar] [CrossRef]
- Ryskalin, L.; Biagioni, F.; Busceti, C.L.; Lazzeri, G.; Frati, A.; Fornai, F. The Multi-Faceted Effect of Curcumin in Glioblastoma from Rescuing Cell Clearance to Autophagy-Independent Effects. Molecules 2020, 25, 4839. [Google Scholar] [CrossRef]
- Talib, W.H.; Al-Hadid, S.A.; Ali, M.B.W.; Al-Yasari, I.H.; Ali, M.R.A. Role of curcumin in regulating p53 in breast cancer: An overview of the mechanism of action. Breast Cancer Targets Ther. 2018, 10, 207–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, J.-R.; Liu, P.-L.; Chen, Y.-H.; Chou, S.-H.; Cheng, Y.-J.; Hwang, J.-J.; Chong, I.-W. Curcumin Inhibits Non-Small Cell Lung Cancer Cells Metastasis through the Adiponectin/NF-κb/MMPs Signaling Pathway. PLoS ONE 2015, 10, e0144462. [Google Scholar] [CrossRef] [Green Version]
- Lagisetty, P.; Vilekar, P.; Sahoo, K.; Anant, S.; Awasthi, V. CLEFMA—An anti-proliferative curcuminoid from structure–activity relationship studies on 3,5-bis(benzylidene)-4-piperidones. Bioorg. Med. Chem. 2010, 18, 6109–6120. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, K.; Dozmorov, M.G.; Anant, S.; Awasthi, V. The curcuminoid CLEFMA selectively induces cell death in H441 lung adenocarcinoma cells via oxidative stress. Investig. New Drugs 2010, 30, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.R.; Sahoo, K.; Awasthi, V. Preclinical evaluation of 4-[3,5-bis(2-chlorobenzylidene)-4-oxo-piperidine-1-yl]-4-oxo-2-butenoic acid, in a mouse model of lung cancer xenograft. J. Cereb. Blood Flow Metab. 2013, 170, 1436–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billen, L.P.; Shamas-Din, A.; Andrews, D.W. Bid: A Bax-like BH3 protein. Oncogene 2008, 27, S93–S104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traenckner, E.; Wilk, S.; Baeuerle, P. A proteasome inhibitor prevents activation of NF-kappa B and stabilizes a newly phosphorylated form of I kappa B-alpha that is still bound to NF-kappa B. EMBO J. 1994, 13, 5433–5441. [Google Scholar] [CrossRef] [PubMed]
- McDade, T.P.; Perugini, R.A.; Vittimberga, F.J., Jr.; Callery, M.P. Ubiquitin-proteasome inhibition enhances apoptosis of human pancreatic cancer cells. Surgery 1999, 126, 371–377. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.-P.; Li, S.; Li, H.-B. Spices for Prevention and Treatment of Cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, Q.-H.; Xu, K. Are isothiocyanates potential anti-cancer drugs? Acta Pharmacol. Sin. 2009, 30, 501–512. [Google Scholar] [CrossRef]
- Sehrawat, A.; Kim, S.-H.; Vogt, A.; Singh, S.V. Suppression of FOXQ1 in benzyl isothiocyanate-mediated inhibition of epithelial–mesenchymal transition in human breast cancer cells. Carcinogenesis 2012, 34, 864–873. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhu, Y.; Yan, H.; Liu, B.; Li, Y.; Zhou, Q.; Xu, K. Isothiocyanates induce oxidative stress and suppress the metastasis potential of human non-small cell lung cancer cells. BMC Cancer 2010, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Lim, D.Y.; Kwon, G.T.; Kim, J.H.; Huang, Z.; Song, H.; Oh, Y.S.; Kang, Y.-H.; Lee, K.W.; Dong, Z.; et al. Benzyl Isothiocyanate Inhibits Prostate Cancer Development in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest. Int. J. Mol. Sci. 2016, 17, 264. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Thornalley, P.J. Studies on the mechanism of the inhibition of human leukaemia cell growth by dietary isothiocyanates and their cysteine adducts in vitro. Biochem. Pharmacol. 2000, 60, 221–231. [Google Scholar] [CrossRef]
- Lai, K.-C.; Huang, A.-C.; Hsu, S.-C.; Kuo, C.-L.; Yang, J.-S.; Wu, S.-H.; Chung, J.-G. Benzyl Isothiocyanate (BITC) Inhibits Migration and Invasion of Human Colon Cancer HT29 Cells by Inhibiting Matrix Metalloproteinase-2/-9 and Urokinase Plasminogen (uPA) through PKC and MAPK Signaling Pathway. J. Agric. Food Chem. 2010, 58, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Li, W.; Dong, X.; Chen, Y.; Lu, Y.; Lin, B.; Guo, J.; Li, M. Benzyl-isothiocyanate Induces Apoptosis and Inhibits Migration and Invasion of Hepatocellular Carcinoma Cells in vitro. J. Cancer 2017, 8, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Sahu, R.P.; Srivastava, S.K. The Role of STAT-3 in the Induction of Apoptosis in Pancreatic Cancer Cells by Benzyl Isothiocyanate. JNCI J. Natl. Cancer Inst. 2009, 101, 176–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pasqua, A.J.; Hong, C.; Wu, M.Y.; McCracken, E.; Wang, X.; Mi, L.; Chung, F.-L. Sensitization of Non-small Cell Lung Cancer Cells to Cisplatin by Naturally Occurring Isothiocyanates. Chem. Res. Toxicol. 2010, 23, 1307–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Loganathan, S.; Humphreys, I.; Srivastava, S.K. Benzyl Isothiocyanate-Induced DNA Damage Causes G2/M Cell Cycle Arrest and Apoptosis in Human Pancreatic Cancer Cells. J. Nutr. 2006, 136, 2728–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boreddy, S.R.; Pramanik, K.C.; Srivastava, S.K. Pancreatic Tumor Suppression by Benzyl Isothiocyanate Is Associated with Inhibition of PI3K/AKT/FOXO Pathway. Clin. Cancer Res. 2011, 17, 1784–1795. [Google Scholar] [CrossRef]
- Boreddy, S.R.; Sahu, R.P.; Srivastava, S.K. Benzyl Isothiocyanate Suppresses Pancreatic Tumor Angiogenesis and Invasion by Inhibiting HIF-α/VEGF/Rho-GTPases: Pivotal Role of STAT-3. PLoS ONE 2011, 6, e25799. [Google Scholar] [CrossRef]
- Xie, B.; Nagalingam, A.; Kuppusamy, P.; Muniraj, N.; Langford, P.; Győrffy, B.; Saxena, N.K.; Sharma, D. Benzyl Isothiocyanate potentiates p53 signaling and antitumor effects against breast cancer through activation of p53-LKB1 and p73-LKB1 axes. Sci. Rep. 2017, 7, 40070. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Adkins, C.; Lockman, P.; Srivastava, S.K. Metastasis of Breast Tumor Cells to Brain Is Suppressed by Phenethyl Isothiocyanate in a Novel In Vivo Metastasis Model. PLoS ONE 2013, 8, e67278. [Google Scholar] [CrossRef]
- Gupta, P.; Srivastava, S.K. Antitumor activity of phenethyl isothiocyanate in HER2-positive breast cancer models. BMC Med. 2012, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Wright, S.E.; Srivastava, S.K. PEITC treatment suppresses myeloid derived tumor suppressor cells to inhibit breast tumor growth. OncoImmunology 2015, 4, e981449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyanapalli, S.S.; Li, W.; Fuentes, F.; Guo, Y.; Ramirez, C.N.; Gonzalez, X.-P.; Pung, D.; Kong, A.-N.T. Epigenetic reactivation of RASSF1A by phenethyl isothiocyanate (PEITC) and promotion of apoptosis in LNCaP cells. Pharmacol. Res. 2016, 114, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Li, S.; Sargsyan, D.; Yin, R.; Kuo, H.; Peter, R.; Wang, L.; Hudlikar, R.; Liu, X.; Kong, A. DNA methylome, transcriptome, and prostate cancer prevention by phenethyl isothiocyanate in TRAMP mice. Mol. Carcinog. 2021, 60, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, G.; Pelicano, H.; Liao, J.; Huang, J.; Feng, L.; Keating, M.J.; Huang, P. Targeting p53-deficient chronic lymphocytic leukemia cells in vitro and in vivo by ROS-mediated mechanism. Oncotarget 2016, 7, 71378–71389. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Kadam, S.; Brahme, A.; Agrawal, M.; Apte, K.; Narke, G.; Ghosh, B.; Madas, S.; Salvi, S. Immuno-Metabolic Alterations in Systemic Immune Cells of Tobacco-Smoke Associated Chronic Obstructive Pulmonary Disease (COPD) Subjects. Am. J. Respir. Crit. Care. 2019, 20, 171. [Google Scholar] [CrossRef]
- Brahme, A.; Kadam, S.; Agrawal, M.; Apte, K.; Narke, G.; Madas, S.; Salvi, S.; Agarwal, A. Metabolic paralysis in systemic immune cells of Chronic Obstructive Pulmonary Disease (COPD) subjects. Eur. Respir. J. 2018, 52, PA934. [Google Scholar] [CrossRef]
- Agarwal, A.R.; Kadam, S.; Brahme, A.; Manas, A.; Apte, K.; Narke, G.; Madas, S.; Salvi, S.S. Systemic Immune-Metabolic Deficiency in Tobacco-and Biomass-Smoke Exposed Chronic Obstructive Pulmonary Disease (COPD) Subjects. Am. J. Respir. Crit. Care 2018, 197, A1190. [Google Scholar]
- Agarwal, A.R.; Kadam, S.; Brahme, A.; Agrawal, M.; Apte, K.; Narke, G.; Kekan, K.; Madas, S.; Salvi, S. Systemic Immuno-metabolic alterations in chronic obstructive pulmonary disease (COPD). Respir. Res. 2019, 20, 120. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Kadam, S.; Brahme, A.; Agrawal, M.; Apte, K.; Narke, G.; Madas, S.; Salvi, S. Systemic Immunometabolism in Asthmatics and Biomass or Tobacco Smoke Exposed Chronic Obstructive Pulmonary Disease (COPD) Patients. Am. J. Respir. Crit. Care 2019, 199, A3802. [Google Scholar] [CrossRef]
- Yuan, J.-M.; Stepanov, I.; Murphy, S.E.; Wang, R.; Allen, S.; Jensen, J.; Strayer, L.; Adams-Haduch, J.; Upadhyaya, P.; Le, C.; et al. Clinical Trial of 2-Phenethyl Isothiocyanate as an Inhibitor of Metabolic Activation of a Tobacco-Specific Lung Carcinogen in Cigarette Smokers. Cancer Prev. Res. 2016, 9, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Jiang, X.; Meng, L.; Dong, X.; Shen, Y.; Xin, Y. Anticancer Activity of Sulforaphane: The Epigenetic Mechanisms and the Nrf2 Signaling Pathway. Oxidative Med. Cell. Longev. 2018, 2018, 5438179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandala, P.K.; Srivastava, S.K. Activation of Checkpoint Kinase 2 by 3,3′-Diindolylmethane Is Required for Causing G2/M Cell Cycle Arrest in Human Ovarian Cancer Cells. Mol. Pharmacol. 2010, 78, 297–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandala, P.K.; Wright, S.E.; Srivastava, S.K. Blocking Epidermal Growth Factor Receptor Activation by 3,3′-Diindolylmethane Suppresses Ovarian Tumor Growth In Vitro and In Vivo. J. Pharmacol. Exp. Ther. 2012, 341, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Kandala, P.K.; Srivastava, S.K. Diindolylmethane suppresses ovarian cancer growth and potentiates the effect of cisplatin in tumor mouse model by targeting signal transducer and activator of transcription 3 (STAT3). BMC Med. 2012, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Kandala, P.K.; Srivastava, S.K. Regulation of macroautophagy in ovarian cancer cells in vitro and in vivo by controlling Glucose regulatory protein 78 and AMPK. Oncotarget 2012, 3, 435–449. [Google Scholar] [CrossRef] [Green Version]
- Fofaria, N.M.; Srivastava, S.K. STAT3 induces anoikis resistance, promotes cell invasion and metastatic potential in pancreatic cancer cells. Carcinogenesis 2015, 36, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandala, P.K.; Srivastava, S.K. Diindolylmethane-mediated Gli1 Protein Suppression Induces Anoikis in Ovarian Cancer Cells in Vitro and Blocks Tumor Formation Ability in Vivo. J. Biol. Chem. 2012, 287, 28745–28754. [Google Scholar] [CrossRef]
- Shilpa, G.; Lakshmi, S.; Jamsheena, V.; Lankalapalli, R.S.; Prakash, V.; Anbumani, S.; Priya, S. Studies on the mode of action of synthetic diindolylmethane derivatives against triple negative breast cancer cells. Basic Clin. Pharmacol. Toxicol. 2022, 131, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Munakarmi, S.; Shrestha, J.; Shin, H.-B.; Lee, G.-H.; Jeong, Y.-J. 3,3′-Diindolylmethane Suppresses the Growth of Hepatocellular Carcinoma by Regulating Its Invasion, Migration, and ER Stress-Mediated Mitochondrial Apoptosis. Cells 2021, 10, 1178. [Google Scholar] [CrossRef]
- Wang, T.T.Y.; Pham, Q.; Kim, Y.S. Elucidating the Role of CD84 and AHR in Modulation of LPS-Induced Cytokines Production by Cruciferous Vegetable-Derived Compounds Indole-3-Carbinol and 3,3′-Diindolylmethane. Int. J. Mol. Sci. 2018, 19, 339. [Google Scholar] [CrossRef] [Green Version]
- Popolo, A.; Pinto, A.; Daglia, M.; Nabavi, S.F.; Farooqi, A.A.; Rastrelli, L. Two likely targets for the anti-cancer effect of indole derivatives from cruciferous vegetables: PI3K/Akt/mTOR signalling pathway and the aryl hydrocarbon receptor. Semin. Cancer Biol. 2017, 46, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Srivastava, S.K. Inhibition of HER2-integrin signaling by Cucurbitacin B leads to in vitro and in vivo breast tumor growth suppression. Oncotarget 2014, 5, 1812–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Srivastava, S.K. HER2 mediated de novo production of TGFβ leads to SNAIL driven epithelial-to-mesenchymal transition and metastasis of breast cancer. Mol. Oncol. 2014, 8, 1532–1547. [Google Scholar] [CrossRef]
- Ma, W.; Xiang, Y.; Yang, R.; Zhang, T.; Xu, J.; Wu, Y.; Liu, X.; Xiang, K.; Zhao, H.; Liu, Y.; et al. Cucurbitacin B induces inhibitory effects via the CIP2A/PP2A/C-KIT signaling axis in t(8;21) acute myeloid leukemia. J. Pharmacol. Sci. 2019, 139, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Liu, Y.; Liu, W.; Ma, F.; Zhou, Y.; Chen, M.; Chang, J.; Wang, Y.; Yang, G.; He, G. Cucurbitacin B inhibits growth and induces apoptosis through the JAK2/STAT3 and MAPK pathways in SH-SY5Y human neuroblastoma cells. Mol. Med. Rep. 2014, 10, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Bian, Z.-G.; Zhang, Y.; Wang, J.-H.; Kan, L.; Wang, X.; Niu, H.-Y.; He, P. Cucurbitacin B inhibits proliferation and induces apoptosis via STAT3 pathway inhibition in A549 lung cancer cells. Mol. Med. Rep. 2014, 10, 2905–2911. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Sun, C.; Shan, X.; Yang, X.; Li-Ling, J.; Deng, Y. Inhibition of Pancreatic Cancer Cell Growth by Cucurbitacin B Through Modulation of Signal Transducer and Activator of Transcription 3 Signaling. Pancreas 2010, 39, 923–929. [Google Scholar] [CrossRef]
- Saglam, A.S.Y.; Alp, E.; Elmazoglu, Z.; Menevse, S. Treatment with cucurbitacin B alone and in combination with gefitinib induces cell cycle inhibition and apoptosis via EGFR and JAK/STAT pathway in human colorectal cancer cell lines. Hum. Exp. Toxicol. 2016, 35, 526–543. [Google Scholar] [CrossRef]
- Gundala, S.R.; Mukkavilli, R.; Yang, C.; Yadav, P.; Tandon, V.; Vangala, S.; Prakash, S.; Aneja, R. Enterohepatic recirculation of bioactive ginger phytochemicals is associated with enhanced tumor growth-inhibitory activity of ginger extract. Carcinogenesis 2014, 35, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.-K.; Tsai, Y.-H.; Korinek, M.; Hung, P.-H.; El-Shazly, M.; Cheng, Y.-B.; Wu, Y.-C.; Hsieh, T.-J.; Chang, F.-R. 6-Paradol and 6-Shogaol, the Pungent Compounds of Ginger, Promote Glucose Utilization in Adipocytes and Myotubes, and 6-Paradol Reduces Blood Glucose in High-Fat Diet-Fed Mice. Int. J. Mol. Sci. 2017, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Kotowski, U.; Kadletz, L.; Schneider, S.; Foki, E.; Schmid, R.; Seemann, R.; Thurnher, D.; Heiduschka, G. 6-shogaol induces apoptosis and enhances radiosensitivity in head and neck squamous cell carcinoma cell lines. Phytother. Res. 2018, 32, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Vasudevan, S.; Sengupta, S. 6-Shogaol Inhibits Breast Cancer Cells and Stem Cell-Like Spheroids by Modulation of Notch Signaling Pathway and Induction of Autophagic Cell Death. PLoS ONE 2015, 10, e0137614. [Google Scholar] [CrossRef] [PubMed]
- Mansingh, D.P.; Sunanda, O.J.; Sali, V.K.; Vasanthi, H.R. [6]-Gingerol-induced cell cycle arrest, reactive oxygen species generation, and disruption of mitochondrial membrane potential are associated with apoptosis in human gastric cancer (AGS) cells. J. Biochem. Mol. Toxicol. 2018, 32, e22206. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, G.; Kathiresan, S.; Kannappan, N. [6]-Shogaol, a dietary phenolic compound, induces oxidative stress mediated mitochondrial dependant apoptosis through activation of proapoptotic factors in Hep-2 cells. Biomed. Pharmacother. 2016, 82, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, S.; Govindhan, A. [6]-Shogaol, a Novel Chemopreventor in 7,12-Dimethylbenz[a]anthracene-induced Hamster Buccal Pouch Carcinogenesis. Phytotherapy Res. 2016, 30, 646–653. [Google Scholar] [CrossRef]
- Dai, Y.; Zhao, Y.; Nie, K. The Antiemetic Mechanisms of Gingerols against Chemotherapy-Induced Nausea and Vomiting. Evid.-Based Complement. Altern. Med. 2022, 2022, 1753430. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K. Ginger and Its Constituents: Role in Prevention and Treatment of Gastrointestinal Cancer. Gastroenterol. Res. Pract. 2015, 2015, 142979. [Google Scholar] [CrossRef]
- Babasheikhali, S.R.; Rahgozar, S.; Mohammadi, M. Ginger extract has anti-leukemia and anti-drug resistant effects on malignant cells. J. Cancer Res. Clin. Oncol. 2019, 145, 1987–1998. [Google Scholar] [CrossRef]
- Rhode, J.; Fogoros, S.; Zick, S.; Wahl, H.; A Griffith, K.; Huang, J.; Liu, J.R. Ginger inhibits cell growth and modulates angiogenic factors in ovarian cancer cells. BMC Complement. Altern. Med. 2007, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Seshadri, V.D.; Oyouni, A.A.A.; Bawazir, W.M.; Alsagaby, S.A.; Alsharif, K.F.; Albrakati, A.; Al-Amer, O.M. Zingiberene exerts chemopreventive activity against 7,12-dimethylbenz(a)anthracene-induced breast cancer in Sprague-Dawley rats. J. Biochem. Mol. Toxicol. 2022, 36, e23146. [Google Scholar] [CrossRef]
- Sehrawat, A.; Arlotti, J.A.; Murakami, A.; Singh, S.V. Zerumbone causes Bax- and Bak-mediated apoptosis in human breast cancer cells and inhibits orthotopic xenograft growth in vivo. Breast Cancer Res. Treat. 2012, 136, 429–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Dulcey, L.; Tomasin, R.; Naves, M.A.; Da Silva, J.A.; Cominetti, M.R. Autophagy-dependent apoptosis is triggered by a semi-synthetic [6]-gingerol analogue in triple negative breast cancer cells. Oncotarget 2018, 9, 30787–30804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Seo, E.Y.; E Kang, N.; Kim, W.K. [6]-Gingerol inhibits metastasis of MDA-MB-231 human breast cancer cells. J. Nutr. Biochem. 2008, 19, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.C.B.; Fuzer, A.M.; Becceneri, A.B.; da Silva, J.A.; Tomasin, R.; Denoyer, D.; Kim, S.-H.; McIntyre, K.A.; Pearson, H.B.; Yeo, B.; et al. [10]-gingerol induces apoptosis and inhibits metastatic dissemination of triple negative breast cancer in vivo. Oncotarget 2017, 8, 72260–72271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, A.; Blando, J.; Silver, E.; Beltran, L.; Sessler, J.; DiGiovanni, J. 6-Shogaol from Dried Ginger Inhibits Growth of Prostate Cancer Cells Both In Vitro and In Vivo through Inhibition of STAT3 and NF-κB Signaling. Cancer Prev. Res. 2014, 7, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Karna, P.; Chagani, S.; Gundala, S.R.; Rida, P.C.G.; Asif, G.; Sharma, V.; Gupta, M.V.; Aneja, R. Benefits of whole ginger extract in prostate cancer. Br. J. Nutr. 2011, 107, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Wani, N.A.; Zhang, B.; Teng, K.Y.; Barajas, J.M.; Motiwal, T.; Hu, P.; Yu, L.B.; Bruschweiler, R.; Ghoshal, K.; Jacob, S.T. Reprogramming of Glucose Metabolism by Zerumbone Suppresses Hepatocarcinogenesis. Mol. Cancer Res. 2022, 20, 256–268. [Google Scholar] [CrossRef]
- Hessien, M.; El-Gendy, S.; Donia, T.; Sikkena, M.A. Growth inhibition of human non-small lung cancer cells h460 by green tea and ginger polyphenols. Anti-Cancer Agents Med. Chem. 2012, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, K.; Ando, T.; Maeda, O.; Ohmiya, N.; Niwa, Y.; Kadomatsu, K.; Goto, H. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem. Biophys. Res. Commun. 2007, 362, 218–223. [Google Scholar] [CrossRef]
- Oh, T.-I.; Jung, H.-J.; Lee, Y.-M.; Lee, S.; Kim, G.-H.; Kan, S.-Y.; Kang, H.; Oh, T.; Ko, H.M.; Kwak, K.-C.; et al. Zerumbone, a Tropical Ginger Sesquiterpene of Zingiber officinale Roscoe, Attenuates α-MSH-Induced Melanogenesis in B16F10 Cells. Int. J. Mol. Sci. 2018, 19, 3149. [Google Scholar] [CrossRef] [Green Version]
- Zaid, A.; Haw, X.R.; Alkatib, H.H.; Sasidharan, S.; Marriott, P.J.; Wong, Y.F. Phytochemical Constituents and Antiproliferative Activities of Essential Oils from Four Varieties of Malaysian Zingiber officinale Roscoe against Human Cervical Cancer Cell Line. Plants 2022, 11, 1280. [Google Scholar] [CrossRef]
- Saud, S.M.; Li, W.; Gray, Z.; Matter, M.S.; Colburn, N.H.; Young, M.R.; Kim, Y.S. Diallyl Disulfide (DADS), a Constituent of Garlic, Inactivates NF-κB and Prevents Colitis-Induced Colorectal Cancer by Inhibiting GSK-3β. Cancer Prev. Res. 2016, 9, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Lea, M.; Randolph, V.M.; Patel, M. Increased acetylation of histones induced by diallyl disulfide and structurally related molecules. Int. J. Oncol. 1999, 15, 347–399. [Google Scholar] [CrossRef] [PubMed]
- Herman-Antosiewicz, A.; A Powolny, A.; Singh, S.V. Molecular targets of cancer chemoprevention by garlic-derived organosulfides. Acta Pharmacol. Sin. 2007, 28, 1355–1364. [Google Scholar] [CrossRef]
- Hu, X.; Benson, P.J.; Srivastava, S.K.; Mack, L.M.; Xia, H.; Gupta, V.; Zaren, H.A.; Singh, S.V. GlutathioneS-Transferases of Female A/J Mouse Liver and Forestomach and Their Differential Induction by Anti-carcinogenic Organosulfides from Garlic. Arch. Biochem. Biophys. 1996, 336, 199–214. [Google Scholar] [CrossRef]
- Hu, X.; Singh, S.V. GlutathioneS-Transferases of Female A/J Mouse Lung and Their Induction by Anticarcinogenic Organosulfides from Garlic. Arch. Biochem. Biophys. 1997, 340, 279–286. [Google Scholar] [CrossRef]
- Hu, X.; Benson, P.J.; Srivastava, S.K.; Xia, H.; Bleicher, R.J.; Zaren, H.A.; Awasthi, S.; Awasthi, Y.C.; Singh, S.V. Induction of glutathione S-transferase pi as a bioassay for the evaluation of potency of inhibitors of benzo(a)pyrene-induced cancer in a murine model. Int. J. Cancer 1997, 73, 897–902. [Google Scholar] [CrossRef]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and Onions: Their Cancer Prevention Properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, K.J.; Mohaideen, N.S.M.H.; Srinivasan, H. Phytocompounds of Onion Target Heat Shock Proteins (HSP70s) to Control Breast Cancer Malignancy. Appl. Biochem. Biotechnol. 2022, 194, 4836–4851. [Google Scholar] [CrossRef]
- Zielińska-Przyjemska, M.; Olejnik, A.; Dobrowolska-Zachwieja, A.; Łuczak, M.; Baer-Dubowska, W. DNA damage and apoptosis in blood neutrophils of inflammatory bowel disease patients and in Caco-2 cells in vitro exposed to betanin. Postep. Hig. Med. Dosw. 2016, 70, 265–271. [Google Scholar] [CrossRef]
- Sreekanth, D.; Arunasree, M.; Roy, K.R.; Reddy, T.C.; Reddy, G.V.; Reddanna, P. Betanin a betacyanin pigment purified from fruits of Opuntia ficus-indica induces apoptosis in human chronic myeloid leukemia Cell line-K562. Phytomedicine 2007, 14, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; An, D.; Nguyen, C.T.T.; Patil, B.S.; Kim, J.; Yoo, K.S. Betalain and Betaine Composition of Greenhouse- or Field-Produced Beetroot (Beta vulgaris L.) and Inhibition of HepG2 Cell Proliferation. J. Agric. Food Chem. 2014, 62, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-J.; Yu, Q.; Yan, H.; Khan, A.; Feng, M.-Y.; Li, P.-P.; Hao, X.-J.; An, L.-K.; Liu, H.-Y. Meroterpenoids with Antitumor Activities from Guava (Psidium guajava). J. Agric. Food Chem. 2017, 65, 4993–4999. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Longato, G.; Ruiz, A.L.T.G.; Tinti, S.; Possenti, A.; Vendramini-Costa, D.B.; Sartoratto, A.; Figueira, G.; Silva, F.; Eberlin, M.; et al. In Vitro, In Vivo and In Silico Analysis of the Anticancer and Estrogen-like Activity of Guava Leaf Extracts. Curr. Med. Chem. 2014, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.-H.; Wang, Z.-H.; Meng, D.-L.; Li, X. Cytotoxic and antioxidant constituents from the leaves of Psidium guajava. Bioorg. Med. Chem. Lett. 2015, 25, 2193–2198. [Google Scholar] [CrossRef]
- Shao, M.; Wang, Y.; Huang, X.-J.; Fan, C.-L.; Zhang, Q.-W.; Zhang, X.-Q.; Ye, W.-C. Four new triterpenoids from the leaves of Psidium guajava. J. Asian Nat. Prod. Res. 2012, 14, 348–354. [Google Scholar] [CrossRef]
- Jiang, L.; Lu, J.; Qin, Y.; Jiang, W.; Wang, Y. Antitumor effect of guava leaves on lung cancer: A network pharmacology study. Arab. J. Chem. 2020, 13, 7773–7797. [Google Scholar] [CrossRef]
- Lin, H.-C.; Lin, J.-Y. GSF3, a polysaccharide from guava (Psidium guajava L.) seeds, inhibits MCF-7 breast cancer cell growth via increasing Bax/Bcl-2 ratio or Fas mRNA expression levels. Int. J. Biol. Macromol. 2020, 161, 1261–1271. [Google Scholar] [CrossRef]
- Ryu, N.H.; Park, K.-R.; Kim, S.-M.; Yun, H.-M.; Nam, D.; Lee, S.-G.; Jang, H.-J.; Ahn, K.S.; Shim, B.S.; Choi, S.-H.; et al. A Hexane Fraction of Guava Leaves (Psidium guajava L.) Induces Anticancer Activity by Suppressing AKT/Mammalian Target of Rapamycin/Ribosomal p70 S6 Kinase in Human Prostate Cancer Cells. J. Med. Food 2012, 15, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-C.; Chiang, C.-C.; Lin, C.-H.; Chen, C.-S.; Wei, C.-W.; Lin, S.-Y.; Yiang, G.-T.; Yu, Y. Anti-cancer therapeutic benefit of red guava extracts as a potential therapy in combination with doxorubicin or targeted therapy for triple-negative breast cancer cells. Int. J. Med. Sci. 2020, 17, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Paśko, P.; Galanty, A.; Zagrodzki, P.; Luksirikul, P.; Barasch, D.; Nemirovski, A.; Gorinstein, S. Dragon Fruits as a Reservoir of Natural Polyphenolics with Chemopreventive Properties. Molecules 2021, 26, 2158. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.K.; Alexander-Lindo, R.L.; Nair, M.G. Relative Inhibition of Lipid Peroxidation, Cyclooxygenase Enzymes, and Human Tumor Cell Proliferation by Natural Food Colors. J. Agric. Food Chem. 2005, 53, 9268–9273. [Google Scholar] [CrossRef]
- Trang, D.T.; Van Hoang, T.K.; Nguyen, T.T.M.; Van Cuong, P.; Dang, N.H.; Dang, H.D.; Quang, T.N.; Dat, N.T. Essential Oils of Lemongrass (Cymbopogon citratus Stapf) Induces Apoptosis and Cell Cycle Arrest in A549 Lung Cancer Cells. BioMed Res. Int. 2020, 2020, 5924856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudai, N.; Weinstein, Y.; Krup, M.; Rabinski, T.; Ofir, R. Citral is a New Inducer of Caspase-3 in Tumor Cell Lines. Planta Med. 2005, 71, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Maruoka, T.; Kitanaka, A.; Kubota, Y.; Yamaoka, G.; Kameda, T.; Imataki, O.; Dobashi, H.; Bandoh, S.; Kadowaki, N.; Tanaka, T. Lemongrass essential oil and citral inhibit Src/Stat3 activity and suppress the proliferation/survival of small-cell lung cancer cells, alone or in combination with chemotherapeutic agents. Int. J. Oncol. 2018, 52, 1738–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balusamy, S.R.; Perumalsamy, H.; Veerappan, K.; Huq, A.; Rajeshkumar, S.; Lakshmi, T.; Kim, Y.J. Citral Induced Apoptosis through Modulation of Key Genes Involved in Fatty Acid Biosynthesis in Human Prostate Cancer Cells: In Silico and In Vitro Study. BioMed Res. Int. 2020, 2020, 6040727. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Tian, L.; Gan, L.; Liu, Z.; Sun, C. Mark4 promotes adipogenesis and triggers apoptosis in 3T3-L1 adipocytes by activating JNK1 and inhibiting p38MAPK pathways. Biol. Cell 2014, 106, 294–307. [Google Scholar] [CrossRef]
- Naz, F.; Khan, F.I.; Mohammad, T.; Khan, P.; Manzoor, S.; Hasan, G.M.; Lobb, K.A.; Luqman, S.; Islam, A.; Ahmad, F.; et al. Investigation of molecular mechanism of recognition between citral and MARK4: A newer therapeutic approach to attenuate cancer cell progression. Int. J. Biol. Macromol. 2018, 107, 2580–2589. [Google Scholar] [CrossRef]
- Sheikh, B.Y.; Sarker, M.R.; Kamarudin, M.N.A.; Mohan, G. Antiproliferative and apoptosis inducing effects of citral via p53 and ROS-induced mitochondrial-mediated apoptosis in human colorectal HCT116 and HT29 cell lines. Biomed. Pharmacother. 2017, 96, 834–846. [Google Scholar] [CrossRef]
- Kuzu, B.; Cüce, G.; Ayan, I.; Gültekin, B.; Canbaz, H.T.; Dursun, H.G.; Şahin, Z.; Keskin, I.; Kalkan, S.S. Evaluation of Apoptosis Pathway of Geraniol on Ishikawa Cells. Nutr. Cancer 2020, 73, 2532–2537. [Google Scholar] [CrossRef]
- Shakya, G.; Balasubramanian, S.; Hoda, M.; Rajagopalan, R. Inhibition of metastasis and angiogenesis in Hep-2 cells by wheatgrass extract—An in vitro and in silico approach. Toxicol. Mech. Methods 2018, 28, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.H.; Choi, M.-H.; Shin, H.-J.; Lee, J.-E. Wheatgrass Extract Ameliorates Hypoxia-induced Mucin Gene Expression in A549 cells. Pharmacogn. Mag. 2017, 13, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Avisar, A.; Cohen, M.; Brenner, B.; Bronshtein, T.; Machluf, M.; Bar-Sela, G.; Aharon, A. Extracellular Vesicles Reflect the Efficacy of Wheatgrass Juice Supplement in Colon Cancer Patients During Adjuvant Chemotherapy. Front. Oncol. 2020, 10, 1659. [Google Scholar] [CrossRef]
- Avisar, A.; Cohen, M.; Katz, R.; Kutiel, T.S.; Aharon, A.; Bar-Sela, G. Wheatgrass Juice Administration and Immune Measures during Adjuvant Chemotherapy in Colon Cancer Patients: Preliminary Results. Pharmaceuticals 2020, 13, 129. [Google Scholar] [CrossRef] [PubMed]
- Galeone, C.; Tavani, A.; Pelucchi, C.; Turati, F.; Winn, D.M.; Levi, F.; Yu, G.-P.; Morgenstern, H.; Kelsey, K.; Maso, L.D.; et al. Coffee and Tea Intake and Risk of Head and Neck Cancer: Pooled Analysis in the International Head and Neck Cancer Epidemiology Consortium. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1723–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.C.; Luo, D.; Pang, G.D.; Xiao, J.; Yang, X.R.; Zhang, Y.; Jia, H.Y. A dose–response meta-analysis of coffee consumption and thyroid cancer occurrence. Int. J. Food Sci. Nutr. 2019, 71, 176–185. [Google Scholar] [CrossRef]
- Larsson, S.C.; Wolk, A. Coffee Consumption and Risk of Liver Cancer: A Meta-Analysis. Gastroenterology 2007, 132, 1740–1745. [Google Scholar] [CrossRef]
- Montenegro, J.; Freitas-Silva, O.; Teodoro, A.J. Molecular Mechanisms of Coffee on Prostate Cancer Prevention. BioMed Res. Int. 2022, 2022, 3254420. [Google Scholar] [CrossRef]
- Friberg, E.; Orsini, N.; Mantzoros, C.S.; Wolk, A. Coffee drinking and risk of endometrial cancer-A population-based cohort study. Int. J. Cancer 2009, 125, 2413–2417. [Google Scholar] [CrossRef]
- Pietrocola, F.; Malik, S.A.; Mariño, G.; Vacchelli, E.; Senovilla, L.; Chaba, K.; Niso-Santano, M.; Maiuri, M.C.; Madeo, F.; Kroemer, G. Coffee induces autophagy in vivo. Cell Cycle 2014, 13, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-Y.; Lee, S.-H.; Lee, J.-H.; Lee, J.-H.; Jang, B.K.; Kim, M.K. Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 10509. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.S.; Asif, M.; Basheer, M.K.A.; Kang, C.W.; Al-Suede, F.S.; Ein, O.C.; Tang, J.; Majid, A.S.A.; Majid, A.M.S.A. Treatment of novel IL17A inhibitor in glioblastoma implementing 3rd generation co-culture cell line and patient-derived tumor model. Eur. J. Pharmacol. 2017, 803, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Bonafé, G.A.; Boschiero, M.N.; Sodré, A.R.; Ziegler, J.V.; Rocha, T.; Ortega, M.M. Natural Plant Compounds: Does Caffeine, Dipotassium Glycyrrhizinate, Curcumin, and Euphol Play Roles as Antitumoral Compounds in Glioblastoma Cell Lines? Front. Neurol. 2022, 12, 784330. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Aronson, W.J.; Huang, M.; Zhang, Y.; Lee, R.-P.; Heber, D.; Henning, S.M. Green Tea Polyphenols and Metabolites in Prostatectomy Tissue: Implications for Cancer Prevention. Cancer Prev. Res. 2010, 3, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.S.; Wang, X.; Lu, G.; Picinich, S.C. Cancer prevention by tea: Animal studies, molecular mechanisms and human relevance. Nat. Rev. Cancer 2009, 9, 429–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.-Y.; Lee, J.-K.; Jeon, Y.-K.; Kim, C.-W. Exosome derived from epigallocatechin gallate treated breast cancer cells suppresses tumor growth by inhibiting tumor-associated macrophage infiltration and M2 polarization. BMC Cancer 2013, 13, 421. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, T.; Nakajima, T.; Moriguchi, M.; Jo, M.; Sekoguchi, S.; Ishii, M.; Takashima, H.; Katagishi, T.; Kimura, H.; Minami, M.; et al. A green tea polyphenol, epigalocatechin-3-gallate, induces apoptosis of human hepatocellular carcinoma, possibly through inhibition of Bcl-2 family proteins. J. Hepatol. 2006, 44, 1074–1082. [Google Scholar] [CrossRef]
- Huang, C.-H.; Tsai, S.-J.; Wang, Y.-J.; Pan, M.-H.; Kao, J.-Y.; Way, T.-D. EGCG inhibits protein synthesis, lipogenesis, and cell cycle progression through activation of AMPK in p53 positive and negative human hepatoma cells. Mol. Nutr. Food Res. 2009, 53, 1156–1165. [Google Scholar] [CrossRef]
- Duhon, D.; Bigelow, R.L.H.; Coleman, D.T.; Steffan, J.J.; Yu, C.; Langston, W.; Kevil, C.G.; Cardelli, J.A. The polyphenol epigallocatechin-3-gallate affects lipid rafts to block activation of the c-Met receptor in prostate cancer cells. Mol. Carcinog. 2010, 49, 739–749. [Google Scholar] [CrossRef]
- Gupta, S.; Hussain, T.; Mukhtar, H. Molecular pathway for (−)-epigallocatechin-3-gallate-induced cell cycle arrest and apoptosis of human prostate carcinoma cells. Arch. Biochem. Biophys. 2002, 410, 177–185. [Google Scholar] [CrossRef]
- Mak, K.-K.; Wu, A.T.H.; Lee, W.-H.; Chang, T.-C.; Chiou, J.-F.; Wang, L.-S.; Wu, C.-H.; Huang, C.-Y.F.; Shieh, Y.-S.; Chao, T.-Y.; et al. Pterostilbene, a bioactive component of blueberries, suppresses the generation of breast cancer stem cells within tumor microenvironment and metastasis via modulating NF-κB/microRNA 448 circuit. Mol. Nutr. Food Res. 2013, 57, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Pestana, D.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C. Blueberry anthocyanins and pyruvic acid adducts: Anticancer properties in breast cancer cell lines. Phytother. Res. 2010, 24, 1862–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernández-Ledesma, B. In vitro chemopreventive properties of peptides released from quinoa (Chenopodium quinoa Willd.) protein under simulated gastrointestinal digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Murad, L.D.; Soares, N.d.C.P.; Brand, C.; Monteiro, M.C.; Teodoro, A.J. Effects of Caffeic and 5-Caffeoylquinic Acids on Cell Viability and Cellular Uptake in Human Colon Adenocarcinoma Cells. Nutr. Cancer 2015, 67, 532–542. [Google Scholar] [CrossRef]
- Ding, H.; Chin, Y.-W.; Kinghorn, A.D.; D’Ambrosio, S.M. Chemopreventive characteristics of avocado fruit. Semin. Cancer Biol. 2007, 17, 386–394. [Google Scholar] [CrossRef]
- Larrosa, M.; González-Sarrías, A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Urolithins, Ellagic Acid-Derived Metabolites Produced by Human Colonic Microflora, Exhibit Estrogenic and Antiestrogenic Activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Xu, S. Intakes of citrus fruit and risk of esophageal cancer: A meta-analysis. Medicine 2018, 97, e0018. [Google Scholar] [CrossRef]
- Song, J.-K.; Bae, J.-M. Citrus Fruit Intake and Breast Cancer Risk: A Quantitative Systematic Review. J. Breast Cancer 2013, 16, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Orlando, A.; Refolo, M.G.; Messa, C.; Amati, L.; Lavermicocca, P.; Guerra, V.; Russo, F. Antiproliferative and Proapoptotic Effects of Viable or Heat-Killed Lactobacillus paracasei IMPC2.1 and Lactobacillus rhamnosus GG in HGC-27 Gastric and DLD-1 Colon Cell Lines. Nutr. Cancer 2012, 64, 1103–1111. [Google Scholar] [CrossRef]
- Leblanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Kahouli, I.; Tomaro-Duchesneau, C.; Prakash, S. Probiotics in colorectal cancer (CRC) with emphasis on mechanisms of action and current perspectives. J. Med. Microbiol. 2013, 62, 1107–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soel, S.M.; Choi, O.S.; Bang, M.H.; Park, J.H.Y.; Kim, W.K. Influence of conjugated linoleic acid isomers on the metastasis of colon cancer cells in vitro and in vivo. J. Nutr. Biochem. 2007, 18, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Weng, W.; Peng, J.; Hong, L.; Yang, L.; Toiyama, Y.; Gao, R.; Liu, M.; Yin, M.; Pan, C.; et al. Fusobacterium nucleatum Increases Proliferation of Colorectal Cancer Cells and Tumor Development in Mice by Activating Toll-Like Receptor 4 Signaling to Nuclear Factor−κB, and Up-regulating Expression of MicroRNA-21. Gastroenterology 2017, 152, 851–866.e24. [Google Scholar] [CrossRef] [Green Version]
- Chandel, D.; Sharma, M.; Chawla, V.; Sachdeva, N.; Shukla, G. Isolation, characterization and identification of antigenotoxic and anticancerous indigenous probiotics and their prophylactic potential in experimental colon carcinogenesis. Sci. Rep. 2019, 9, 14769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Sankarapandian, V.; Maran, B.A.V.; Rajendran, R.L.; Jogalekar, M.P.; Gurunagarajan, S.; Krishnamoorthy, R.; Gangadaran, P.; Ahn, B.-C. An Update on the Effectiveness of Probiotics in the Prevention and Treatment of Cancer. Life 2022, 12, 59. [Google Scholar] [CrossRef]
- Pithva, S.P.; Ambalam, P.S.; Ramoliya, J.M.; Dave, J.M.; Vyas, B.R.M. Antigenotoxic and Antimutagenic Activities of Probiotic Lactobacillus rhamnosus Vc against N-Methyl-N′-Nitro-N-Nitrosoguanidine. Nutr. Cancer 2015, 67, 1142–1150. [Google Scholar] [CrossRef]
- Asoudeh-Fard, A.; Barzegari, A.; Dehnad, A.; Bastani, S.; Golchin, A.; Omidi, Y. Lactobacillus plantarum induces apoptosis in oral cancer KB cells through upregulation of PTEN and downregulation of MAPK signalling pathways. BioImpacts 2017, 7, 193–198. [Google Scholar] [CrossRef]
- Kadirareddy, R.H.; Vemuri, S.G.; Palempalli, U.M. Probiotic Conjugated Linoleic Acid Mediated Apoptosis in Breast Cancer Cells by Downregulation of NFkappaB. Asian Pac. J. Cancer Prev. 2016, 17, 3395–3403. [Google Scholar]
- Khosrovan, Z.; Haghighat, S.; Mahdavi, M. The Probiotic Bacteria Induce Apoptosis in Breast and Colon Cancer Cells: An Immunostimulatory Effect. ImmunoRegulation 2020, 3, 37–50. [Google Scholar] [CrossRef]
- Taherian-Esfahani, Z.; Abedin-Do, A.; Nouri, Z.; Mirfakhraie, R.; Ghafouri-Fard, S.; Motevaseli, E. Lactobacilli Differentially Modulate mTOR and Wnt/β-Catenin Pathways in Different Cancer Cell Lines. Iran. J. Cancer Prev. 2016, 9, e5369. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Belzacq, A.-S.; Haouzi, D.; Rouault, A.; Métivier, D.; Kroemer, G.; Brenner, C. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Chumchalová, J.; Šmarda, J. Human tumor cells are selectively inhibited by colicins. Folia Microbiol. 2003, 48, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Shukla, G. Administration of Metabiotics Extracted from Probiotic Lactobacillus rhamnosus MD 14 Inhibit Experimental Colorectal Carcinogenesis by Targeting Wnt/β-Catenin Pathway. Front. Oncol. 2020, 10, 746. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-Free Supernatants from Probiotic Lactobacillus casei and Lactobacillus rhamnosus GG Decrease Colon Cancer Cell Invasion In Vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Escamilla, J.; Lane, M.A.; Maitin, V. Probiotic Lactobacilli Decrease Invasion of Metastatic Human Colon Cancer Cells In Vitro. FASEB J. 2010, 24, 928. [Google Scholar]
- Nomura, M.; Nagatomo, R.; Doi, K.; Shimizu, J.; Baba, K.; Saito, T.; Matsumoto, S.; Inoue, K.; Muto, M. Association of Short-Chain Fatty Acids in the Gut Microbiome with Clinical Response to Treatment with Nivolumab or Pembrolizumab in Patients with Solid Cancer Tumors. JAMA Netw. Open 2020, 3, e202895. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Sheng, J.; Wang, M.; Luo, H.; Zhu, J.; Zhang, B.; Liu, Z.; Yang, X. Combination Therapy of TGF-β Blockade and Commensal-derived Probiotics Provides Enhanced Antitumor Immune Response and Tumor Suppression. Theranostics 2019, 9, 4115–4129. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.M.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef]
- Seki, H.; Shiohara, M.; Matsumura, T.; Miyagawa, N.; Tanaka, M.; Komiyama, A.; Kurata, S. Prevention of antibiotic-associated diarrhea in children by Clostridium butyricum MIYAIRI. Pediatr. Int. 2003, 45, 86–90. [Google Scholar] [CrossRef]
- Derosa, L.; Zitvogel, L. A probiotic supplement boosts response to cancer immunotherapy. Nat. Med. 2022, 28, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vetizou, M.; Cogdill, A.P.; Khan, A.W.; Zhang, X.; White, M.G.; Peterson, C.B.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef]
- Bolat, Z.B.; Islek, Z.; Demir, B.N.; Yilmaz, E.N.; Sahin, F.; Ucisik, M.H. Curcumin- and Piperine-Loaded Emulsomes as Combinational Treatment Approach Enhance the Anticancer Activity of Curcumin on HCT116 Colorectal Cancer Model. Front. Bioeng. Biotechnol. 2020, 8, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, T.O.; Keum, Y.-S.; Lin, W.; Kim, J.-H.; Hu, R.; Shen, G.; Xu, C.; Gopalakrishnan, A.; Reddy, B.; Zheng, X.; et al. Combined Inhibitory Effects of Curcumin and Phenethyl Isothiocyanate on the Growth of Human PC-3 Prostate Xenografts in Immunodeficient Mice. Cancer Res. 2006, 66, 613–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaikwad, S.R.; Srivastava, S.K. Antioxidant Activity of Phytochemicals in Cancer. In Handbook of Oxidative Stress in Cancer: Therapeutic Aspects; Chakraborti, S., Ed.; Springer: Singapore, 2021; pp. 1–17. [Google Scholar]
- Shi, Y.; Sahu, R.P.; Srivastava, S.K. Triphala inhibits both in vitro and in vivo xenograft growth of pancreatic tumor cells by inducing apoptosis. BMC Cancer 2008, 8, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpa, E.S.; Emanuelli, M.; Frati, A.; Pozzi, V.; Antonini, E.; Diamantini, G.; Di Ruscio, G.; Sartini, D.; Armeni, T.; Palma, F.; et al. Betacyanins enhance vitexin-2-O-xyloside mediated inhibition of proliferation of T24 bladder cancer cells. Food Funct. 2016, 7, 4772–4780. [Google Scholar] [CrossRef]
- Karthika, C.; Sureshkumar, R.; Sajini, D.V.; Ashraf, G.M.; Rahman, H. 5-fluorouracil and curcumin with pectin coating as a treatment regimen for titanium dioxide with dimethylhydrazine-induced colon cancer model. Environ. Sci. Pollut. Res. 2022, 29, 63202–63215. [Google Scholar] [CrossRef]
- Han, H.-K. The effects of black pepper on the intestinal absorption and hepatic metabolism of drugs. Expert Opin. Drug Metab. Toxicol. 2011, 7, 721–729. [Google Scholar] [CrossRef]
- Sari, N.F.; Lestari, B.; Saputri, D.; Ahsani, A.F.; Santoso, R.A.; Sasmito, E.; Meiyanto, E. Reveal Cyto-toxicity and Antigenotoxicity of Piper nigrum L. Ethanolic Extract and its Combination with Doxorubicin on CHO-K1 Cells. Indones. J. Cancer Chemoprevention 2017, 8, 110–119. [Google Scholar]
- Tsao, C.-W.; Li, J.-S.; Lin, Y.-W.; Wu, S.-T.; Cha, T.-L.; Liu, C.-Y. Regulation of carcinogenesis and mediation through Wnt/β-catenin signaling by 3,3′-diindolylmethane in an enzalutamide-resistant prostate cancer cell line. Sci. Rep. 2021, 11, 1239. [Google Scholar] [CrossRef]
- Draz, H.; Goldberg, A.A.; Guns, E.S.T.; Fazli, L.; Safe, S.; Sanderson, J.T. Autophagy inhibition improves the chemotherapeutic efficacy of cruciferous vegetable-derived diindolymethane in a murine prostate cancer xenograft model. Investig. New Drugs 2018, 36, 718–725. [Google Scholar] [CrossRef]
- Cang, S.; Ma, Y.; Chiao, J.-W.; Liu, D. Phenethyl isothiocyanate and paclitaxel synergistically enhanced apoptosis and alpha-tubulin hyperacetylation in breast cancer cells. Exp. Hematol. Oncol. 2014, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Dey, S.; Bhattacharya, R.K.; Roy, M. Isothiocyanates sensitize the effect of chemotherapeutic drugs via modulation of protein kinase C and telomerase in cervical cancer cells. Mol. Cell. Biochem. 2009, 330, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Saraswat, A.; Wei, Z.; Agrawal, M.; Dukhande, V.; Reznik, S.; Patel, K. Development of Dual ARV-825 and Nintedanib-Loaded PEGylated Nano-Liposomes for Synergistic Efficacy in Vemurafnib-Resistant Melanoma. Pharmaceutics 2021, 13, 1005. [Google Scholar] [CrossRef] [PubMed]
- Vartak, R.; Menon, S.; Patki, M.; Billack, B.; Patel, K. Ebselen nanoemulgel for the treatment of topical fungal infection. Eur. J. Pharm. Sci. 2020, 148, 105323. [Google Scholar] [CrossRef]
- Bagde, A.; Patel, K.; Kutlehria, S.; Chowdhury, N.; Singh, M. Formulation of topical ibuprofen solid lipid nanoparticle (SLN) gel using hot melt extrusion technique (HME) and determining its anti-inflammatory strength. Drug Deliv. Transl. Res. 2019, 9, 816–827. [Google Scholar] [CrossRef]
- Rajan, R.; Sabnani, M.K.; Mavinkurve, V.; Shmeeda, H.; Mansouri, H.; Bonkoungou, S.; Le, A.D.; Wood, L.M.; Gabizon, A.A.; La-Beck, N.M. Liposome-induced immunosuppression and tumor growth is mediated by macrophages and mitigated by liposome-encapsulated alendronate. J. Control. Release 2017, 271, 139–148. [Google Scholar] [CrossRef]
- Abadi, A.V.M.; Karimi, E.; Oskoueian, E.; Mohammad, G.R.K.S.; Shafaei, N. Chemical investigation and screening of anti-cancer potential of Syzygium aromaticum L. bud (clove) essential oil nanoemulsion. 3 Biotech 2022, 12, 49. [Google Scholar] [CrossRef]
- Fofaria, N.M.; Qhattal, H.S.S.; Liu, X.; Srivastava, S.K. Nanoemulsion formulations for anti-cancer agent piplartine—Characterization, toxicological, pharmacokinetics and efficacy studies. Int. J. Pharm. 2015, 498, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Qhattal, H.S.S.; Wang, S.; Salihima, T.; Srivastava, S.K.; Liu, X. Nanoemulsions of Cancer Chemopreventive Agent Benzyl Isothiocyanate Display Enhanced Solubility, Dissolution, and Permeability. J. Agric. Food Chem. 2011, 59, 12396–12404. [Google Scholar] [CrossRef]
- Ranjan, A.; Fofaria, N.M.; Kim, S.-H.; Srivastava, S.K. Modulation of signal transduction pathways by natural compounds in cancer. Chin. J. Nat. Med. 2015, 13, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.; Agrawal, M.Y.; Kaushik, I.; Ramachandran, S.; Srivastava, S.K. Immune checkpoint proteins: Signaling mechanisms and molecular interactions in cancer immunotherapy. Semin. Cancer Biol. 2022, 86, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Arulampalam, V. Uncoupling the p38 MAPK kinase in IBD: A double edged sword? Gut 2002, 50, 446–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Lin, A. Role of JNK activation in apoptosis: A double-edged sword. Cell Res. 2005, 15, 36–42. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Food | Bioactive Component | Cancer/Organ Model | Cell-Line | Mechanism of Action | References |
|---|---|---|---|---|---|---|
| 1 | Black pepper | Piperine | Breast cancer | ↓pAKT, STAT3, p65, IκBα | [31] | |
| Ovarian cancer | A2780 | ↓JNK, p38 MAPK, STAT3 | [13,14] | |||
| Gastric cancer | TMK-1 | ↓IL-6 | [14] | |||
| Glioblastoma | ↓CDK2-cyclin-E, CDK-4/6-cyclin D Jnk/p38 G1/S cell cycle arrest | [16] | ||||
| Lung cancer | A549 | ↑p53 G2/M cell cycle arrest and induction of intrinsic apoptosis | [17] | |||
| Leukemia | HL60 and K-562 | Induction of both intrinsic and extrinsic apoptotic pathway | [29,41] | |||
| Colorectal cancer | HCT116, SW480 | ↓Wnt/β-catenin signaling | [43] | |||
| 2 | Black pepper | Piperidine | Breast cancer | MDA-MB-231 and MCF-7 cells | G0/G1 cell cycle arrest | [44] |
| Piperidine (17a) | Prostate cancer | PC3 | ↑Bax ↓Bcl-2 and XIAP | [24,45] | ||
| Piperidine (2-amino-4-(1-piperidine) | Colon cancer | HT29 and DLD-1 | G0/G1 cell cycle arrest ↓FOXA2, vimentin | [46,47] | ||
| Piperidine (CLEFMA) | Lung cancer | H441 cells | Induction of redox homeostasis Apoptosis induction S-phase cell cycle arrest ↓COX2, cyclin D, NFκB, CD31 | [90,92,94] | ||
| 3 | Long pepper | Piperlongumine | Renal cancer | PNX0010 | ↓c-met via ROS-dependent mechanism ↓STAT3, NFκB, PI3K/Akt, and ERK/MAPK | [48] |
| Piperlongumine | Glioblastoma | U87, U251 | ↑HTRPV2 | [49] | ||
| Piperlongumine | Cervical, pancreatic, and colorectal | HeLa, PANC1, and SW620 | ↓GSTP1 | [50] | ||
| 4 | Chili pepper | Capsaicin | Pancreatic cancer | AsPC-1 and BxPC-3 | Induction of ROS and apoptosis ↓Bcl-2, survivin, β-catenin, JNK | [53] |
| Capsaicin | Pancreatic cancer | AsPC-1 and BxPC-3 | ↑Inhibition of mitochondrial complex-1 and complex-3 followed by ROS generation | [54] | ||
| Capsaicin | Bladder cancer | ↓SIRT-1 deacetylase | [61] | |||
| 5 | Clove | Whole extract | Colon cancer | HT29 | G0/G1 phase arrest ↓E2F1, thymidylate synthase ↑p21, WAF1/Cip1, γ-H2AX | [79] |
| Whole extract | Breast cancer | MCF-7 | ↑ALDH1, caspase-3, and Bax ↓CD44, CD24, and ALDH1 | [80] | ||
| Aqueous extract | Pancreatic and colon cancer | ASPC-1 and human colon HT-29 | ↑AMPK and ULK | [81] | ||
| Nanoemulsion | Thyroid cancer | Hth-7, B-CPAP, BHT-101, and KTC-1 cell line | Apoptosis induction | [82] | ||
| 6 | Turmeric | Curcumin | Pancreatic cancer | BxPC-3 | G2/M phase arrest ↑pH2A.X, CHK1, p-ATM, ↓Cyclin B1 DNA damage effect is regulated via ER stress pathway | [83,84] |
| Curcumin | Oral squamous carcinoma | HSC-3 cells | Akt/PI3K/mTOR ↑miR-199a | [86] | ||
| Curcumin | Glioblastoma | ↓MMP, NF-κB, STAT3, Akt | [87] | |||
| 7 | Cruciferous vegetables | Benzyl isothiocyanate | Pancreatic cancer | Capan-2 | ↑DNA damage G2/M phase arrest and apoptosis induction | [106] |
| Benzyl isothiocyanate | Pancreatic cancer | BxPC3, MiaPaCa2, and Panc-1 | ↓STAT-3, PI3K/AKT/FOXO | [104,107] | ||
| Benzyl isothiocyanate | Colon cancer | HT29 | ↓Urokinase-type plasminogen activator (uPA) protein kinase C (PKC), MAPK signaling pathway, and MMP-2/9 pathway | [102] | ||
| Phenethyl isothiocyanate (PEITC) | Breast cancer | MDA-MB-231-BR (BR-brain seeking) | ↓HER2 | [110] | ||
| PEITC | Breast cancer | MDA-MB-231 (in vivo xenografts in artificial immune environment) | ↓MDSC cells | [112] | ||
| PEITC | Prostate cancer | LNCap cells | Reactivation of tumor suppressor gene RASSF1A and apoptosis induction Cell cycle arrest and impact on global CpG epigenome | [113,114] | ||
| PEITC | Chronic lymphocytic leukemia | Primary leukemia cells deficient in p53. | ROS generation via glutathione depletion | [115] | ||
| Diindolylmethane (DIM) | Ovarian cancer | SKOV-3, TOV-21G, and OVCAR-3 | ↑p-H2A.X, Chk2 ↓p-EGFR, MEK, ERK, p-STAT3, VEGF, and HIF-1α Induction of macroautophagy with AMPK activation | [123,124,126] | ||
| DIM | Ovarian cancer | A2780 and OVCAR-429 cells | ↓Gli-1 | [128] | ||
| DIM | Breast cancer | MDA-MB-231 | ↓MMP-2, MMP-9, and Akt/PI3K/mTOR signaling axis, EGFR | [129] | ||
| DIM | Hepatocellular carcinoma | Hep3B and HuhCell | Suppression of EMT via ER stress induction | [130] | ||
| Sulphoraphane | Prostate, colon, breast, lung, and oral | ↑Nrf2 activation ↓HDAC Apoptosis induction, cell cycle arrest and inhibition of angiogenesis. | [122] | |||
| 8 | Cucurbitaceous food | Cucurbitacin B (CuB) | Breast cancer | ↓HER2 and ITGA and ITGA4 ↑ITGB1 and ITGB3 | [133] | |
| CuB | Colorectal cancer, lung cancer, pancreatic cancer, breast cancer, neuroblastoma, and acute myeloid leukemia | Inhibition of STAT3 signaling | [133,135] | |||
| CuB | Colorectal cancer | HT-29 and HCT-116 cell lines | Inhibition of EGFR and JAK/STAT pathway | [139] | ||
| CuB | Neuroblastoma | SHSY5Y cells | Inhibition of MAPK pathway | [136] | ||
| CuB | Myeloid leukemia | Kasumi-1, acute promyelocytic leukemia (HL60), acute myelomonocytic leukemia (U937), chronic myelogenous leukemia (K562), Burkitt’s lymphoma (Raji) and T cell acute lymphoblastic leukemia (Molt-4) | CIP2A/PP2A/C-KIT signaling axis | [135] | ||
| 9 | Ginger | 6-Shogaol | Breast cancer | MCF-7 and MDA-MB-231 | Autophagic cell death and modulation of Notch signaling | [143] |
| Gingerol, zerumbone, zingiberene | Breast cancer | MCF-7 and MDA-MB-231 Human brain seeking (MDA-MB-231BrM) | Metastatic inhibition and cell cycle arrest | [152,154,155] | ||
| Zerumbone and 6-shogaol | Prostate cancer | DU-145 | Cell cycle arrest in G1 phase ↓STAT-3 and NF-κB | [156] | ||
| Zerumbone | Hepatocellular carcinoma | HepG2, SNU-182, Hep3B, SNU-449, Sk-Hep-1, and Huh-7 cells | G2/M arrest ↓PI3K/Akt/mTOR and STAT3 signaling axis | [158] | ||
| 10 | Garlic | Diallyl disulfide (DADS) | Colon cancer | SW480 cells | ↓NF-κB | [163] |
| Diallyl sulfide (DAS), DADS, and diallyl trisulfide (DATS) | Lung, liver, and forestomach | ↑GST enzymes (α (mGSTA1-2, mGSTA3-3, mGSTA4-4), µ (mGSTM1-1), and π (mGSTP1-1)) | [166,167,168] | |||
| 11 | Onion | Organosulfur compounds | Breast cancer | ↑Heat shock proteins (HSP70) and binding immunoglobulin protein (BiP). | [170] | |
| 12 | Beetroot | Betanin | Chronic myeloid leukemia | K562 cells | Induction of apoptosis by cytochrome c release from mitochondria | [172] |
| Betalain and betanine | Hepatocellular cancer | HepG2 cells | Free radical scavenging activity | [173] | ||
| 13 | Guava | Guava seed polysaccharide fraction 3 (GSF3) | Breast cancer | MCF-7 cells | Apoptosis induction by increasing Bax/Bcl-2 ratio | [179] |
| Guava leaf hexane fraction (GHF), ß-eudesmol, α-copaene, α-patchoulene, ß-caryophyllene oxide (CPO), octadecane, α-terpineol | Prostate cancer | PC-3 and LNCaP cells | ↓Akt/mTOR/S6K and MAPK signaling | [180] | ||
| Total extracts and smaller molecular weight (<30 kDa) extracts from guava fruit | Breast cancer | MDA-MB-231 and MDA-MB-468 cells | Apoptotic and necrotic cell death induction | [181] | ||
| 14 | Dragon fruit | Betalain | Breast cancer, colon cancer, stomach, glioblastoma, and lung cancer | MCF-7, HCT-116, AGS, SF268, and NCI-H460 cells | Inhibition of lipid peroxidation and COX enzymes | [183] |
| 15 | Lemongrass | Lemongrass oil terpenes | Lung cancer | A549, NCI-H1975, NCI-H1650, and NCI-H1299 | ↑Caspase-3 G2/M cell cycle arrest | [184,185] |
| Citral | Small-cell lung cancer (SCLC) | LU135 SCLC cell line | Inhibition of Src/STAT3 activity | [186] | ||
| Citral | Prostate cancer | PC-3 and PC-3M (metastatic) | Suppression of lipogenesis through inhibition of p-AMPK and downregulation of sterol regulatory element-binding protein (SREBP1), 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGR), fatty acid synthase (FASN), and acetyl coA carboxylase (ACC) | [187] | ||
| Citral | Colon cancer | HT29 and HCT-116 | ↑p53 and apoptosis induction via ROS generation | [190] | ||
| Geraniol | Endometrial adenocarcinoma | Ishikawa cells | ↑Bax, caspase3, caspase-8, cytochrome C, and Fas genes | [191] | ||
| 16 | Wheatgrass | Methanol extract of wheatgrass (MEWG) | Epithelial carcinoma | HEp2 | ↓VEGF, MMP-9, and COX-2 along with inhibition of Akt/PI3K signaling. | [192] |
| Lung adenocarcinoma | A549 cells | ↓HIF-1α, MUC5A, MUC5B, and MUC8 | [193] | |||
| 17 | Coffee | Kahweol | Hepatocellular Carcinoma | Hep3B, SNU182, and SNU423 | Inhibition of Src/mTOR/STAT3 signaling axis | [202] |
| PT93 FLVM and FLVZ | Glioblastoma | U87 MG and DBTRG MG | PT93 inhibits MMP-2 and MMP-9 expression FLVM and FLVZ target IL17A, HIF-1α, and VEGF | [203] | ||
| 18 | Green tea | Epigallocatechin-3-gallate (EGCG) | Hepatocellular carcinoma | HLE, HepG2, HuH-7, and PLC/PRF/5 | Inactivation of AMPK and NF-κB | [208,209] |
| EGCG | Prostate cancer | DU145 cells | ↓Phosphorylation of c-Met, Akt, and Erk | [210] | ||
| 19 | Blueberries | Pterostilbene | Breast cancer | MCF-7 and MDA-MB-231 | ↓NFkB/microRNA 448 circuit | [212] |
| Anthocyanin | Breast cancer | MCF-7 and MDA-MB-231 | - | [213] | ||
| 20 | Quinoa | Caffeic acid | Colon cancer | HT29 | ↑Gastrointestinal digestion, apoptosis, and G0/G1 cell cycle arrest | [214,215] |
| 21 | Avocado | Chloroform extract of avocado | Hepatocellular, oral, prostate, and breast cancer | ↑Cleaved caspase-3 and cleaved PARP leading to apoptosis | [216] | |
| 22 | Pomegranate juice | Ellagic acid, urolithin A and urolithin B | Breast cancer | MCF-7 | ↓17 Beta estradiol | [217] |
| 23 | Citrus fruits | - | Esophageal and breast cancers | - | - | [218,219] |
| 24 | Tomatoes | Tomatidine | Gastric cancer | - | Regulation of ISG genes | [68] |
| Lycopene | Prostate cancer | - | Modulation of AKT/EZH2/androgen receptor signaling pathway | [69] | ||
| Whole powder | Prostate cancer | - | - | [70] | ||
| Lycopene | Hepatocellular carcinoma | - | ↓ cyclin D1, HIF-1α, and PCNA | [71] |
| No. | Probiotic | Type of Cancer | Cell-Line | Mechanism | References |
|---|---|---|---|---|---|
| 1. | Lactobacillus paracasei Lactobacillus rhamnosus | Colon and gastric cancer | DLD-1 (colon) and HGC-27 (gastric) | Induction of apoptosis | [220] |
| 2. | Lactobacillus plantarum (LPCLA) | Breast cancer | MDA-MB-231 | ↓NF-κB pathway | [230] |
| 3. | Lactobacillus crispatus and L. rhamnosus | Various cancer types | HeLa, MDA-MB-231, and HT-29 | ↓Wnt/β-catenin pathway | [232] |
| 4. | Propionibacterium | Colon cancer | HT-29 and Caco-2 | Induction of apoptosis by secreting propionate and acetate | [233] |
| 5. | Escherichia coli | Breast cancer | MCF-7, MDA-MB-231 | G1 phase cell cycle arrest and pore formation through production of a chemical called colicin | [234] |
| 6. | L. rhamnosus | Colon cancer | Animal study | ↑Tumor suppressor p53 | [235] |
| 7. | Lacticaseibacillus casei and L. rhamnosus | Colon cancer | HCT-116 | Inhibit metastatic spread by downregulating MMP-9 | [236,237] |
| 8. | Escherichia coli strain Nissle 1917 (EcN) | Liver and breast cancer | 4T1 and H22 cell lines | ↑Infiltration of T cells and dendritic cells | [239] |
| 9. | Bacteroides fragilis | Sarcoma, melanoma, and colon carcinoma | MCA205 (sarcoma), MC38 (colon), and Ret melanoma model | ↑Response to CTLA -4 blockade | [240] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrawal, M.Y.; Gaikwad, S.; Srivastava, S.; Srivastava, S.K. Research Trend and Detailed Insights into the Molecular Mechanisms of Food Bioactive Compounds against Cancer: A Comprehensive Review with Special Emphasis on Probiotics. Cancers 2022, 14, 5482. https://doi.org/10.3390/cancers14225482
Agrawal MY, Gaikwad S, Srivastava S, Srivastava SK. Research Trend and Detailed Insights into the Molecular Mechanisms of Food Bioactive Compounds against Cancer: A Comprehensive Review with Special Emphasis on Probiotics. Cancers. 2022; 14(22):5482. https://doi.org/10.3390/cancers14225482
Chicago/Turabian StyleAgrawal, Manas Yogendra, Shreyas Gaikwad, Sangeeta Srivastava, and Sanjay K. Srivastava. 2022. "Research Trend and Detailed Insights into the Molecular Mechanisms of Food Bioactive Compounds against Cancer: A Comprehensive Review with Special Emphasis on Probiotics" Cancers 14, no. 22: 5482. https://doi.org/10.3390/cancers14225482