
Strong Humoral but Not Cellular Immune Responses against SARS-CoV-2 in Individuals with Oncohematological Disease Who Were Treated with Rituximab before Receiving a Vaccine Booster
 , , , ,
, , , ,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethical Statement
2.3. Samples Processing and Materials
2.4. Phenotyping of B Lymphocytes
2.5. SARS-CoV-2 Serology
2.6. Pseudovirus Neutralization Assays
2.7. Antibody-Dependent Cellular Cytotoxicity Assay
2.8. Pseudotyped SARS-CoV-2 Infection for Direct Cellular Cytotoxicity Assay
2.9. Statistical Analysis
3. Results
3.1. Patients’ Characteristics
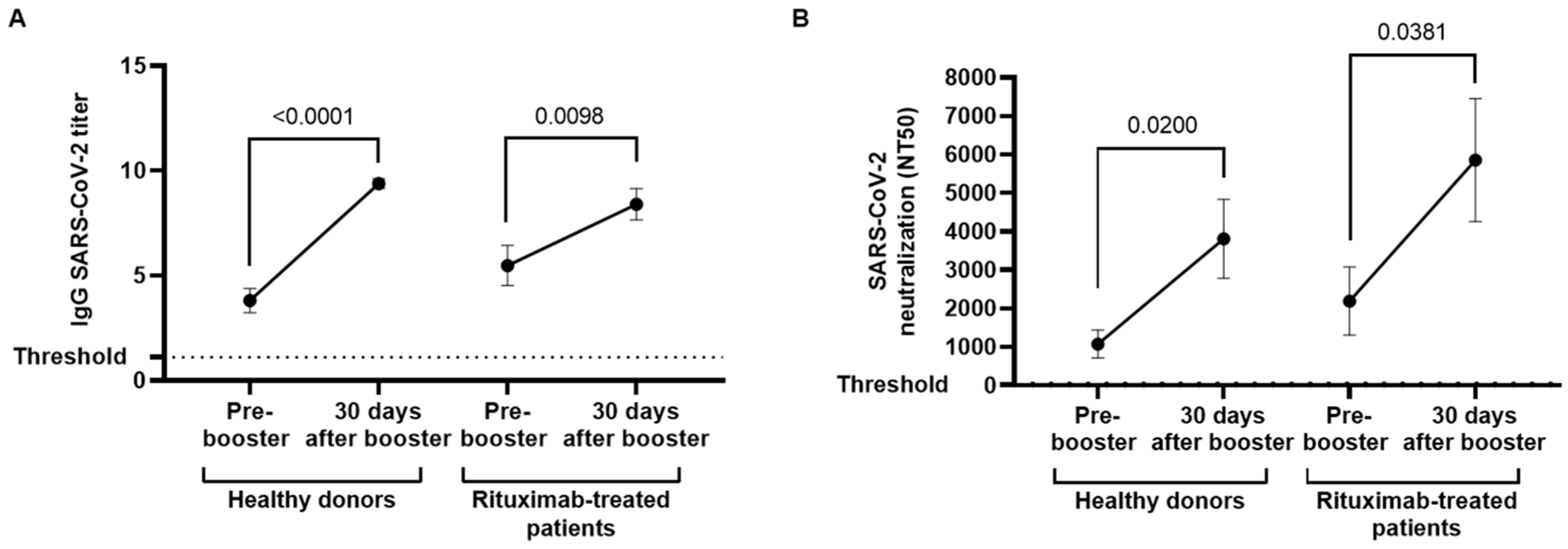
3.2. Serological Response against COVID-19 Vaccination
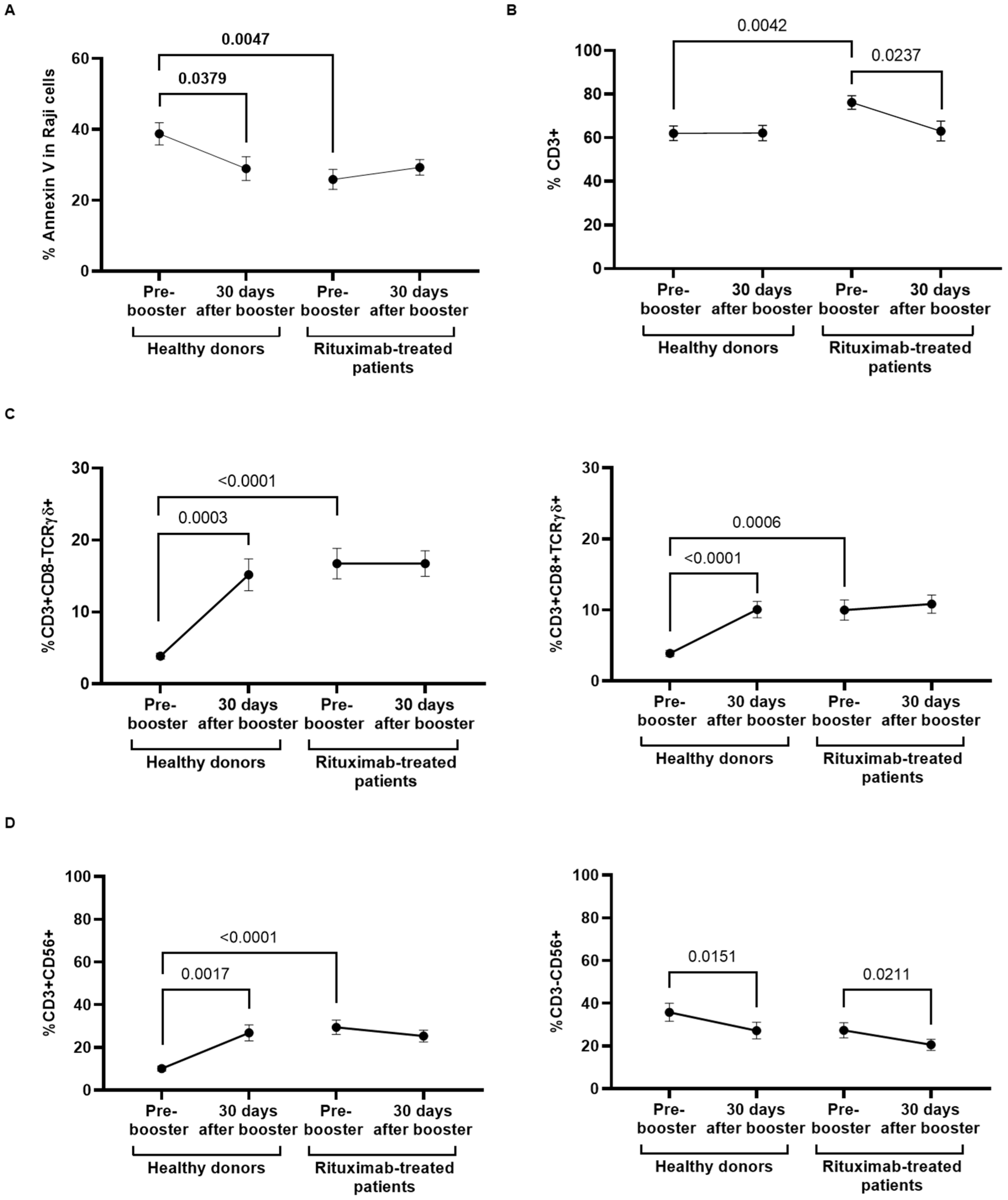
3.3. Analysis of B-Cell Subpopulations
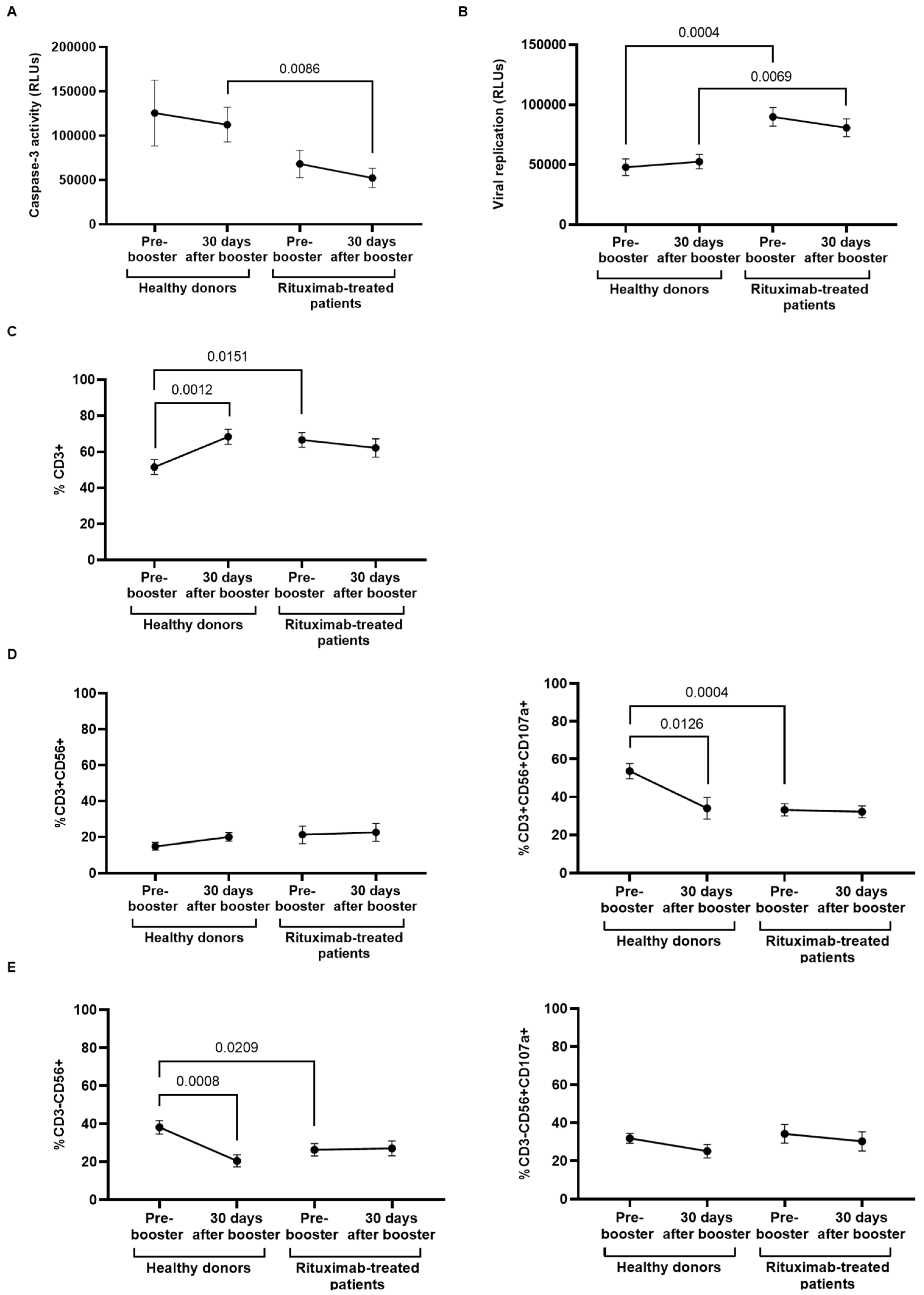
3.4. Antibody-Dependent Cellular Cytotoxic Response of PBMCs from Rituximab-Treated Individuals
3.5. Specific Direct Cellular Cytotoxicity of PBMCs and Viral Neutralization
3.6. Breakthrough Infections
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eisenberg, R.A.; Jawad, A.F.; Boyer, J.; Maurer, K.; McDonald, K.; Prak, E.T.; Sullivan, K.E. Rituximab-treated patients have a poor response to influenza vaccination. J. Clin. Immunol. 2013, 33, 388–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Kolk, L.E.; Baars, J.W.; Prins, M.H.; van Oers, M.H. Rituximab treatment results in impaired secondary humoral immune responsiveness. Blood 2002, 100, 2257–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijenthira, A.; Gong, I.; Betschel, S.D.; Cheung, M.; Hicks, L.K. Vaccine response following anti-CD20 therapy: A systematic review and meta-analysis of 905 patients. Blood Adv. 2021, 5, 2624–2643. [Google Scholar] [CrossRef]
- Cooksley, C.D.; Avritscher, E.B.; Bekele, B.N.; Rolston, K.V.; Geraci, J.M.; Elting, L.S. Epidemiology and outcomes of serious influenza-related infections in the cancer population. Cancer 2005, 104, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.; Marrie, T.J.; Garg, S.; Kellner, J.D.; Tyrrell, G.J.; Group, S. Increased risk of invasive pneumococcal disease in haematological and solid-organ malignancies. Epidemiol. Infect. 2010, 138, 1804–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanif, N.; Anwer, F. Rituximab. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Kasi, P.M.; Tawbi, H.A.; Oddis, C.V.; Kulkarni, H.S. Clinical review: Serious adverse events associated with the use of rituximab—A critical care perspective. Crit. Care 2012, 16, 231. [Google Scholar] [CrossRef] [Green Version]
- Cerny, T.; Borisch, B.; Introna, M.; Johnson, P.; Rose, A.L. Mechanism of action of rituximab. Anticancer Drugs 2002, 13 (Suppl. S2), S3–S10. [Google Scholar] [CrossRef]
- Buske, C.; Hoster, E.; Dreyling, M.; Eimermacher, H.; Wandt, H.; Metzner, B.; Fuchs, R.; Bittenbring, J.; Woermann, B.; Hohloch, K.; et al. The addition of rituximab to front-line therapy with CHOP (R-CHOP) results in a higher response rate and longer time to treatment failure in patients with lymphoplasmacytic lymphoma: Results of a randomized trial of the German Low-Grade Lymphoma Study Group (GLSG). Leukemia 2009, 23, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.A.; Curtis, J.R.; Winthrop, K.L. Impact of disease-modifying antirheumatic drugs on vaccine immunogenicity in patients with inflammatory rheumatic and musculoskeletal diseases. Ann. Rheum. Dis. 2021, 80, 1255–1265. [Google Scholar] [CrossRef]
- Atzrodt, C.L.; Maknojia, I.; McCarthy, R.D.P.; Oldfield, T.M.; Po, J.; Ta, K.T.L.; Stepp, H.E.; Clements, T.P. A Guide to COVID-19: A global pandemic caused by the novel coronavirus SARS-CoV-2. FEBS J. 2020, 287, 3633–3650. [Google Scholar] [CrossRef]
- Negahdaripour, M.; Shafiekhani, M.; Moezzi, S.M.I.; Amiri, S.; Rasekh, S.; Bagheri, A.; Mosaddeghi, P.; Vazin, A. Administration of COVID-19 vaccines in immunocompromised patients. Int. Immunopharmacol. 2021, 99, 108021. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food & Drug Administration. Spikevax and Moderna COVID-19 Vaccine. Available online: https://www.fda.gov/emergency-preparedness-and-response/coronavirus-disease-2019-covid-19/spikevax-and-moderna-covid-19-vaccine (accessed on 18 October 2022).
- U.S. Food & Drug Administration. Comirnaty and Pfizer-BioNTech COVID-19 Vaccine. Available online: https://www.fda.gov/emergency-preparedness-and-response/coronavirus-disease-2019-covid-19/pfizer-biontech-covid-19-vaccine (accessed on 18 October 2022).
- Fessenden, J. Inside the New mRNA Vaccines for COVID-19. Available online: https://www.umassmed.edu/news/news-archives/2020/12/inside-the-new-mrna-vaccines-for-covid-19/ (accessed on 18 October 2022).
- Patel, R.; Kaki, M.; Potluri, V.S.; Kahar, P.; Khanna, D. A comprehensive review of SARS-CoV-2 vaccines: Pfizer, Moderna & Johnson & Johnson. Hum. Vaccin. Immunother. 2022, 18, 2002083. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of Responses after SARS-CoV-2 mRNA-1273 Vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Feikin, D.R.; Higdon, M.M.; Abu-Raddad, L.J.; Andrews, N.; Araos, R.; Goldberg, Y.; Groome, M.J.; Huppert, A.; O’Brien, K.L.; Smith, P.G.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: Results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar] [CrossRef]
- Self, W.H.; Tenforde, M.W.; Rhoads, J.P.; Gaglani, M.; Ginde, A.A.; Douin, D.J.; Olson, S.M.; Talbot, H.K.; Casey, J.D.; Mohr, N.M.; et al. Comparative Effectiveness of Moderna, Pfizer-BioNTech, and Janssen (Johnson & Johnson) Vaccines in Preventing COVID-19 Hospitalizations Among Adults Without Immunocompromising Conditions—United States, March-August 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1337–1343. [Google Scholar] [CrossRef]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 COVID-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef]
- Mofaz, M.; Yechezkel, M.; Guan, G.; Brandeau, M.L.; Patalon, T.; Gazit, S.; Yamin, D.; Shmueli, E. Self-Reported and Physiologic Reactions to Third BNT162b2 mRNA COVID-19 (Booster) Vaccine Dose. Emerg. Infect. Dis. 2022, 28, 1375–1383. [Google Scholar] [CrossRef]
- Comirnaty and Spikevax: EMA Recommendations on Extra Doses and Boosters. Available online: https://www.ema.europa.eu/en/news/comirnaty-spikevax-ema-recommendations-extra-doses-boosters (accessed on 17 September 2022).
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Schietzel, S.; Anderegg, M.; Limacher, A.; Born, A.; Horn, M.P.; Maurer, B.; Hirzel, C.; Sidler, D.; Moor, M.B. Humoral and cellular immune responses on SARS-CoV-2 vaccines in patients with anti-CD20 therapies: A systematic review and meta-analysis of 1342 patients. RMD Open 2022, 8, e002036. [Google Scholar] [CrossRef] [PubMed]
- Mrak, D.; Simader, E.; Sieghart, D.; Mandl, P.; Radner, H.; Perkmann, T.; Haslacher, H.; Mayer, M.; Koblischke, M.; Hofer, P.; et al. Immunogenicity and safety of a fourth COVID-19 vaccination in rituximab-treated patients: An open-label extension study. Ann. Rheum Dis. 2022; published online first. [Google Scholar] [CrossRef]
- Jyssum, I.; Kared, H.; Tran, T.T.; Tveter, A.T.; Provan, S.A.; Sexton, J.; Jorgensen, K.K.; Jahnsen, J.; Kro, G.B.; Warren, D.J.; et al. Humoral and cellular immune responses to two and three doses of SARS-CoV-2 vaccines in rituximab-treated patients with rheumatoid arthritis: A prospective, cohort study. Lancet Rheumatol. 2022, 4, e177–e187. [Google Scholar] [CrossRef]
- Laing, A.G.; Lorenc, A.; Del Molino Del Barrio, I.; Das, A.; Fish, M.; Monin, L.; Munoz-Ruiz, M.; McKenzie, D.R.; Hayday, T.S.; Francos-Quijorna, I.; et al. A dynamic COVID-19 immune signature includes associations with poor prognosis. Nat. Med. 2020, 26, 1623–1635. [Google Scholar] [CrossRef]
- Ansariniya, H.; Seifati, S.M.; Zaker, E.; Zare, F. Comparison of Immune Response between SARS, MERS, and COVID-19 Infection, Perspective on Vaccine Design and Development. Biomed. Res. Int. 2021, 2021, 8870425. [Google Scholar] [CrossRef] [PubMed]
- Bange, E.M.; Han, N.A.; Wileyto, P.; Kim, J.Y.; Gouma, S.; Robinson, J.; Greenplate, A.R.; Hwee, M.A.; Porterfield, F.; Owoyemi, O.; et al. CD8(+) T cells contribute to survival in patients with COVID-19 and hematologic cancer. Nat. Med. 2021, 27, 1280–1289. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Moir, S.; Buckner, C.M.; Ho, J.; Wang, W.; Chen, J.; Waldner, A.J.; Posada, J.G.; Kardava, L.; O’Shea, M.A.; Kottilil, S.; et al. B cells in early and chronic HIV infection: Evidence for preservation of immune function associated with early initiation of antiretroviral therapy. Blood 2010, 116, 5571–5579. [Google Scholar] [CrossRef]
- Garcia-Perez, J.; Sanchez-Palomino, S.; Perez-Olmeda, M.; Fernandez, B.; Alcami, J. A new strategy based on recombinant viruses as a tool for assessing drug susceptibility of human immunodeficiency virus type 1. J. Med. Virol. 2007, 79, 127–137. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borobia, A.M.; Carcas, A.J.; Perez-Olmeda, M.; Castano, L.; Bertran, M.J.; Garcia-Perez, J.; Campins, M.; Portoles, A.; Gonzalez-Perez, M.; Garcia Morales, M.T.; et al. Immunogenicity and reactogenicity of BNT162b2 booster in ChAdOx1-S-primed participants (CombiVacS): A multicentre, open-label, randomised, controlled, phase 2 trial. Lancet 2021, 398, 121–130. [Google Scholar] [CrossRef]
- Vigon, L.; Garcia-Perez, J.; Rodriguez-Mora, S.; Torres, M.; Mateos, E.; Castillo de la Osa, M.; Cervero, M.; Malo De Molina, R.; Navarro, C.; Murciano-Anton, M.A.; et al. Impaired Antibody-Dependent Cellular Cytotoxicity in a Spanish Cohort of Patients With COVID-19 Admitted to the ICU. Front. Immunol. 2021, 12, 742631. [Google Scholar] [CrossRef] [PubMed]
- Diez-Fuertes, F.; Iglesias-Caballero, M.; Garcia-Perez, J.; Monzon, S.; Jimenez, P.; Varona, S.; Cuesta, I.; Zaballos, A.; Jimenez, M.; Checa, L.; et al. A Founder Effect Led Early SARS-CoV-2 Transmission in Spain. J. Virol. 2021, 95, e01583-20. [Google Scholar] [CrossRef]
- Torres, M.; Corona, M.; Rodríguez-Mora, S.; Casado-Fernández, G.; Zurdo-Castronuño, A.; Mateos, E.; Ramos-Martín, F.; Sánchez-Menéndez, C.; Murciano-Antón, M.A.; García-Pérez, J.; et al.; Hematology and Hemotherapy Service, Instituto Ramón y Cajal de Investigación Sanitaria (IRYCIS), Hospital Universitario Ramón y Cajal, Madrid, Spain Personal observation, 2022.
- Cao, Y.; Li, Q.; Chen, J.; Guo, X.; Miao, C.; Yang, H.; Chen, Z.; Li, C.; Li, L. Hospital Emergency Management Plan During the COVID-19 Epidemic. Acad. Emerg. Med. 2020, 27, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- Moor, M.B.; Suter-Riniker, F.; Horn, M.P.; Aeberli, D.; Amsler, J.; Moller, B.; Njue, L.M.; Medri, C.; Angelillo-Scherrer, A.; Borradori, L.; et al. Humoral and cellular responses to mRNA vaccines against SARS-CoV-2 in patients with a history of CD20 B-cell-depleting therapy (RituxiVac): An investigator-initiated, single-centre, open-label study. Lancet Rheumatol. 2021, 3, e789–e797. [Google Scholar] [CrossRef]
- Perry, C.; Luttwak, E.; Balaban, R.; Shefer, G.; Morales, M.M.; Aharon, A.; Tabib, Y.; Cohen, Y.C.; Benyamini, N.; Beyar-Katz, O.; et al. Efficacy of the BNT162b2 mRNA COVID-19 vaccine in patients with B-cell non-Hodgkin lymphoma. Blood Adv. 2021, 5, 3053–3061. [Google Scholar] [CrossRef]
- Tanguay, M.; Boutin, M.; Laumaea, A.; Salaciak, M.; Mendoza, A.; Cassis, C.; Ajjamada, L.; Assouline, S.; Patenaude, F.; Clark, M.W.; et al. B-cell cytopenia and time to last B-cell-depleting therapy predict response to SARS-COV-2 vaccines in patients with lymphoproliferative disorders. Vaccine 2022, 40, 1203–1207. [Google Scholar] [CrossRef]
- Jimenez, M.; Roldan, E.; Fernandez-Naval, C.; Villacampa, G.; Martinez-Gallo, M.; Medina-Gil, D.; Peralta-Garzon, S.; Pujadas, G.; Hernandez, C.; Pages, C.; et al. Cellular and humoral immunogenicity of the mRNA-1273 SARS-CoV-2 vaccine in patients with hematologic malignancies. Blood Adv. 2022, 6, 774–784. [Google Scholar] [CrossRef]
- Shields, A.M.; Venkatachalam, S.; Shafeek, S.; Paneesha, S.; Ford, M.; Sheeran, T.; Kelly, M.; Qureshi, I.; Salhan, B.; Karim, F.; et al. SARS-CoV-2 vaccine responses following CD20-depletion treatment in patients with haematological and rheumatological disease: A West Midlands Research Consortium study. Clin. Exp. Immunol. 2022, 207, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Gavriatopoulou, M.; Fotiou, D.; Giatra, C.; Asimakopoulos, I.; Dimou, M.; Sklirou, A.D.; Ntanasis-Stathopoulos, I.; Darmani, I.; Briasoulis, A.; et al. Poor Neutralizing Antibody Responses in 132 Patients with CLL, NHL and HL after Vaccination against SARS-CoV-2: A Prospective Study. Cancers 2021, 13, 4480. [Google Scholar] [CrossRef]
- Dulery, R.; Lamure, S.; Delord, M.; Di Blasi, R.; Chauchet, A.; Hueso, T.; Rossi, C.; Drenou, B.; Deau Fischer, B.; Soussain, C.; et al. Prolonged in-hospital stay and higher mortality after Covid-19 among patients with non-Hodgkin lymphoma treated with B-cell depleting immunotherapy. Am. J. Hematol. 2021, 96, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Barda, N.; Dagan, N.; Cohen, C.; Hernan, M.A.; Lipsitch, M.; Kohane, I.S.; Reis, B.Y.; Balicer, R.D. Effectiveness of a third dose of the BNT162b2 mRNA COVID-19 vaccine for preventing severe outcomes in Israel: An observational study. Lancet 2021, 398, 2093–2100. [Google Scholar] [CrossRef]
- Hall, V.G.; Ferreira, V.H.; Ku, T.; Ierullo, M.; Majchrzak-Kita, B.; Chaparro, C.; Selzner, N.; Schiff, J.; McDonald, M.; Tomlinson, G.; et al. Randomized Trial of a Third Dose of mRNA-1273 Vaccine in Transplant Recipients. N. Engl. J. Med. 2021, 385, 1244–1246. [Google Scholar] [CrossRef]
- Benucci, M.; Damiani, A.; Gobbi, F.L.; Lari, B.; Grossi, V.; Infantino, M.; Manfredi, M. Role of booster with BNT162b2 mRNA in SARS-CoV-2 vaccination in patients with rheumatoid arthritis. Immunol. Res. 2022, 70, 493–500. [Google Scholar] [CrossRef]
- Shapiro, L.C.; Thakkar, A.; Campbell, S.T.; Forest, S.K.; Pradhan, K.; Gonzalez-Lugo, J.D.; Quinn, R.; Bhagat, T.D.; Choudhary, G.S.; McCort, M.; et al. Efficacy of booster doses in augmenting waning immune responses to COVID-19 vaccine in patients with cancer. Cancer Cell 2022, 40, 3–5. [Google Scholar] [CrossRef]
- Lumley, S.F.; O’Donnell, D.; Stoesser, N.E.; Matthews, P.C.; Howarth, A.; Hatch, S.B.; Marsden, B.D.; Cox, S.; James, T.; Warren, F.; et al. Antibody Status and Incidence of SARS-CoV-2 Infection in Health Care Workers. N. Engl. J. Med 2021, 384, 533–540. [Google Scholar] [CrossRef]
- Marasco, V.; Carniti, C.; Guidetti, A.; Farina, L.; Magni, M.; Miceli, R.; Calabretta, L.; Verderio, P.; Ljevar, S.; Serpenti, F.; et al. T-cell immune response after mRNA SARS-CoV-2 vaccines is frequently detected also in the absence of seroconversion in patients with lymphoid malignancies. Br. J. Haematol. 2022, 196, 548–558. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: A literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef]
- Liu, Q.; Qin, C.; Liu, M.; Liu, J. Effectiveness and safety of SARS-CoV-2 vaccine in real-world studies: A systematic review and meta-analysis. Infect. Dis. Poverty 2021, 10, 132. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Gong, W. Will Mutations in the Spike Protein of SARS-CoV-2 Lead to the Failure of COVID-19 Vaccines? J. Korean Med. Sci. 2021, 36, e124. [Google Scholar] [CrossRef] [PubMed]
- Pormohammad, A.; Zarei, M.; Ghorbani, S.; Mohammadi, M.; Aghayari Sheikh Neshin, S.; Khatami, A.; Turner, D.L.; Djalalinia, S.; Mousavi, S.A.; Mardani-Fard, H.A.; et al. Effectiveness of COVID-19 Vaccines against Delta (B.1.617.2) Variant: A Systematic Review and Meta-Analysis of Clinical Studies. Vaccines 2021, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Chenchula, S.; Karunakaran, P.; Sharma, S.; Chavan, M. Current evidence on efficacy of COVID-19 booster dose vaccination against the Omicron variant: A systematic review. J. Med. Virol. 2022, 94, 2969–2976. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, H.K. Function of gammadelta T cells in tumor immunology and their application to cancer therapy. Exp. Mol. Med. 2021, 53, 318–327. [Google Scholar] [CrossRef]
- Raverdeau, M.; Cunningham, S.P.; Harmon, C.; Lynch, L. γδ T cells in cancer: A small population of lymphocytes with big implications. Clin. Transl. Immunol. 2019, 8, e01080. [Google Scholar] [CrossRef]
- Viale, M.; Ferrini, S.; Bacigalupo, A. TCR gamma/delta positive lymphocytes after allogeneic bone marrow transplantation. Bone Marrow Transplant. 1992, 10, 249–253. [Google Scholar]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Velardi, E.; Tsai, J.J.; van den Brink, M.R.M. T cell regeneration after immunological injury. Nat. Rev. Immunol. 2021, 21, 277–291. [Google Scholar] [CrossRef]
- Gafter-Gvili, A.; Polliack, A. Bendamustine associated immune suppression and infections during therapy of hematological malignancies. Leuk. Lymphoma 2016, 57, 512–519. [Google Scholar] [CrossRef]
- Ito, K.; Okamoto, M.; Ando, M.; Kakumae, Y.; Okamoto, A.; Inaguma, Y.; Tokuda, M.; Yanada, M.; Yamada, S.; Emi, N. Influence of rituximab plus bendamustine chemotherapy on the immune system in patients with refractory or relapsed follicular lymphoma and mantle cell lymphoma. Leuk. Lymphoma 2015, 56, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Garcia Munoz, R.; Izquierdo-Gil, A.; Munoz, A.; Roldan-Galiacho, V.; Rabasa, P.; Panizo, C. Lymphocyte recovery is impaired in patients with chronic lymphocytic leukemia and indolent non-Hodgkin lymphomas treated with bendamustine plus rituximab. Ann. Hematol. 2014, 93, 1879–1887. [Google Scholar] [CrossRef] [PubMed]
- Yutaka, T.; Ito, S.; Ohigashi, H.; Naohiro, M.; Shimono, J.; Souichi, S.; Teshima, T. Sustained CD4 and CD8 lymphopenia after rituximab maintenance therapy following bendamustine and rituximab combination therapy for lymphoma. Leuk. Lymphoma 2015, 56, 3216–3218. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Maruyama, D.; Maeshima, A.M.; Makita, S.; Kitahara, H.; Miyamoto, K.; Fukuhara, S.; Munakata, W.; Suzuki, T.; Kobayashi, Y.; et al. Prolonged lymphocytopenia after bendamustine therapy in patients with relapsed or refractory indolent B-cell and mantle cell lymphoma. Blood Cancer J. 2015, 5, e362. [Google Scholar] [CrossRef] [Green Version]
- Kleinberg, L.; Sloan, L.; Grossman, S.; Lim, M. Radiotherapy, Lymphopenia, and Host Immune Capacity in Glioblastoma: A Potentially Actionable Toxicity Associated with Reduced Efficacy of Radiotherapy. Neurosurgery 2019, 85, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, P.; Zhao, Z.; Mao, Q.; Yu, J.; Li, M. A review of radiation-induced lymphopenia in patients with esophageal cancer: An immunological perspective for radiotherapy. Ther. Adv. Med. Oncol. 2020, 12, 1758835920926822. [Google Scholar] [CrossRef]
- Koukourakis, M.I.; Giatromanolaki, A. Lymphopenia and intratumoral lymphocytic balance in the era of cancer immuno-radiotherapy. Crit. Rev. Oncol. Hematol. 2021, 159, 103226. [Google Scholar] [CrossRef]
- Muhsen, K.; Maimon, N.; Mizrahi, A.; Bodenheimer, O.; Cohen, D.; Maimon, M.; Grotto, I.; Dagan, R. Effectiveness of BNT162b2 mRNA Coronavirus Disease 2019 (COVID-19) Vaccine Against Acquisition of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Among Healthcare Workers in Long-Term Care Facilities: A Prospective Cohort Study. Clin. Infect. Dis. 2022, 75, e755–e763. [Google Scholar] [CrossRef]
- Moncunill, G.; Aguilar, R.; Ribes, M.; Ortega, N.; Rubio, R.; Salmeron, G.; Molina, M.J.; Vidal, M.; Barrios, D.; Mitchell, R.A.; et al. Determinants of early antibody responses to COVID-19 mRNA vaccines in a cohort of exposed and naive healthcare workers. EBioMedicine 2022, 75, 103805. [Google Scholar] [CrossRef]
- Katz, M.A.; Harlev, E.B.; Chazan, B.; Chowers, M.; Greenberg, D.; Peretz, A.; Tshori, S.; Levy, J.; Yacobi, M.; Hirsch, A.; et al. Early effectiveness of BNT162b2 Covid-19 vaccine in preventing SARS-CoV-2 infection in healthcare personnel in six Israeli hospitals (CoVEHPI). Vaccine 2022, 40, 512–520. [Google Scholar] [CrossRef]
- Novazzi, F.; Taborelli, S.; Baj, A.; Focosi, D.; Maggi, F. Asymptomatic SARS-CoV-2 Vaccine Breakthrough Infections in Health Care Workers Identified Through Routine Universal Surveillance Testing. Ann. Intern. Med. 2021, 174, 1770–1772. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n = 18) | Healthy Controls (n = 15) | |
|---|---|---|
| Age, median (IQR) | 61 (53.2–72.8) | 77 (72.0–79.0) |
| Sex: Female, n (%) | 12 (66.66) | 12 (80%) |
| Diagnosis | ||
| 16 (88.89) 1 (5.56) 1 (5.56) | - - - |
| Number of doses of rituximab, n (%) | ||
| 4 (22.22) 9 (50) 5 (27.78) | - - - |
| Concomitant chemotherapy within 18 months prior to booster, n (%) | ||
| 4 (22.22) 2 (11.11) 10 (55.56) 2 (11.11) 1 (5.56) 2 (11.11) 1 (5.56) | - - - - - - - |
| Months from last treatment with rituximab to second vaccine dose, median (IQR) | 9.6 (6–15) | - |
| Months from last treatment with rituximab to third vaccine dose, median (IQR) | 13.8 (9.4–19) | - |
| Days from second dose to first sample, median (IQR) | 124.5 (122.75–126) | 189.3 (184–189) |
| Days from booster dose to second sample, median (IQR) | 28 (28–33.5) | 29 (29.0–42.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, M.; Corona, M.; Rodríguez-Mora, S.; Casado-Fernández, G.; Zurdo-Castronuño, A.; Mateos, E.; Ramos-Martín, F.; Sánchez-Menéndez, C.; Murciano-Antón, M.A.; García-Pérez, J.; et al. Strong Humoral but Not Cellular Immune Responses against SARS-CoV-2 in Individuals with Oncohematological Disease Who Were Treated with Rituximab before Receiving a Vaccine Booster. Cancers 2022, 14, 5537. https://doi.org/10.3390/cancers14225537
Torres M, Corona M, Rodríguez-Mora S, Casado-Fernández G, Zurdo-Castronuño A, Mateos E, Ramos-Martín F, Sánchez-Menéndez C, Murciano-Antón MA, García-Pérez J, et al. Strong Humoral but Not Cellular Immune Responses against SARS-CoV-2 in Individuals with Oncohematological Disease Who Were Treated with Rituximab before Receiving a Vaccine Booster. Cancers. 2022; 14(22):5537. https://doi.org/10.3390/cancers14225537
Chicago/Turabian StyleTorres, Montserrat, Magdalena Corona, Sara Rodríguez-Mora, Guiomar Casado-Fernández, Alejandro Zurdo-Castronuño, Elena Mateos, Fernando Ramos-Martín, Clara Sánchez-Menéndez, María Aranzazú Murciano-Antón, Javier García-Pérez, and et al. 2022. "Strong Humoral but Not Cellular Immune Responses against SARS-CoV-2 in Individuals with Oncohematological Disease Who Were Treated with Rituximab before Receiving a Vaccine Booster" Cancers 14, no. 22: 5537. https://doi.org/10.3390/cancers14225537
APA StyleTorres, M., Corona, M., Rodríguez-Mora, S., Casado-Fernández, G., Zurdo-Castronuño, A., Mateos, E., Ramos-Martín, F., Sánchez-Menéndez, C., Murciano-Antón, M. A., García-Pérez, J., Alcamí, J., Pérez-Olmeda, M., Coiras, M., López-Jiménez, J., García-Gutiérrez, V., & on behalf of the Multidisciplinary Group of Study of COVID-19 (MGS-COVID). (2022). Strong Humoral but Not Cellular Immune Responses against SARS-CoV-2 in Individuals with Oncohematological Disease Who Were Treated with Rituximab before Receiving a Vaccine Booster. Cancers, 14(22), 5537. https://doi.org/10.3390/cancers14225537