

Meta-Analysis of RNA-Seq Datasets Identifies Novel Players in Glioblastoma

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
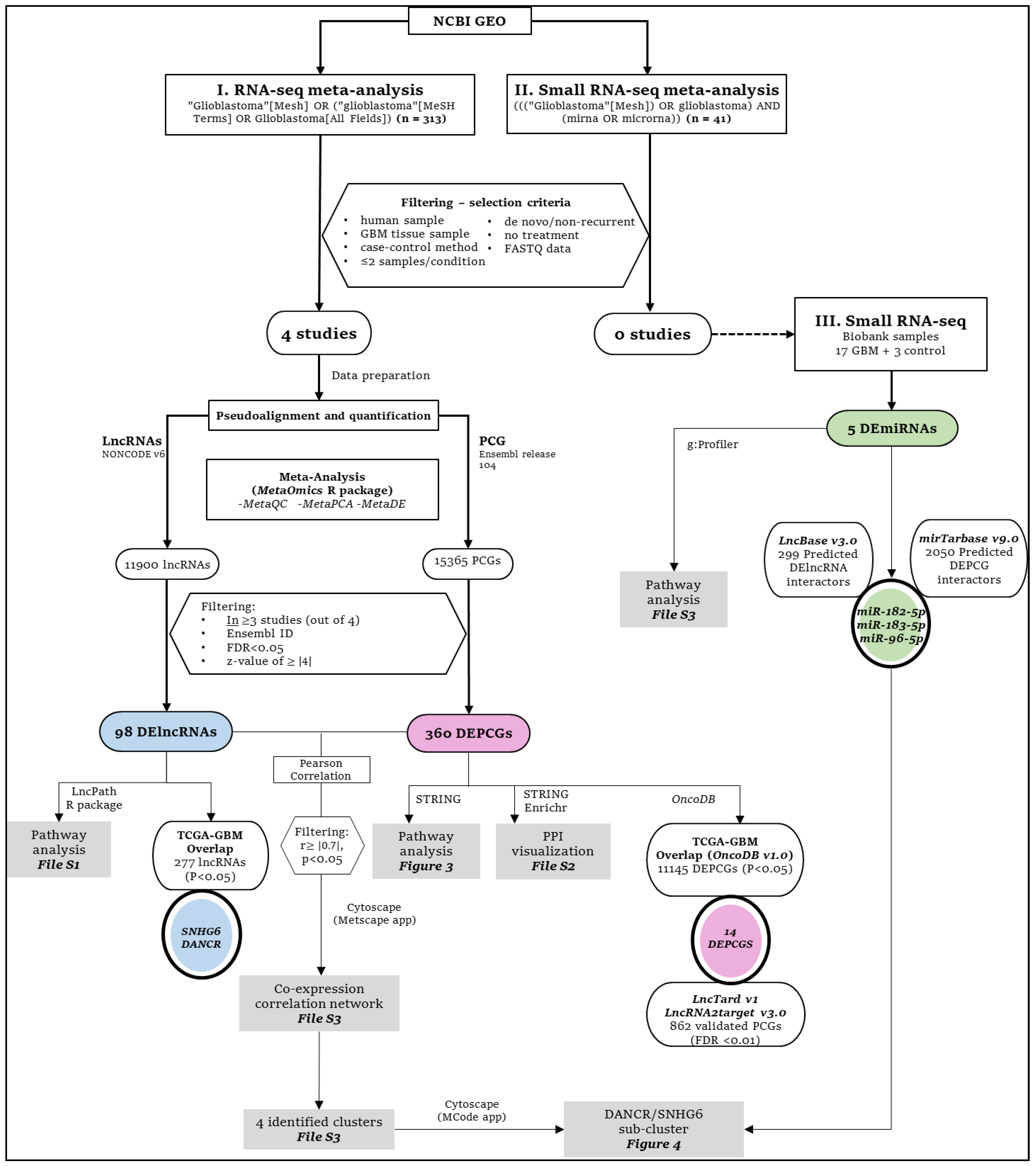
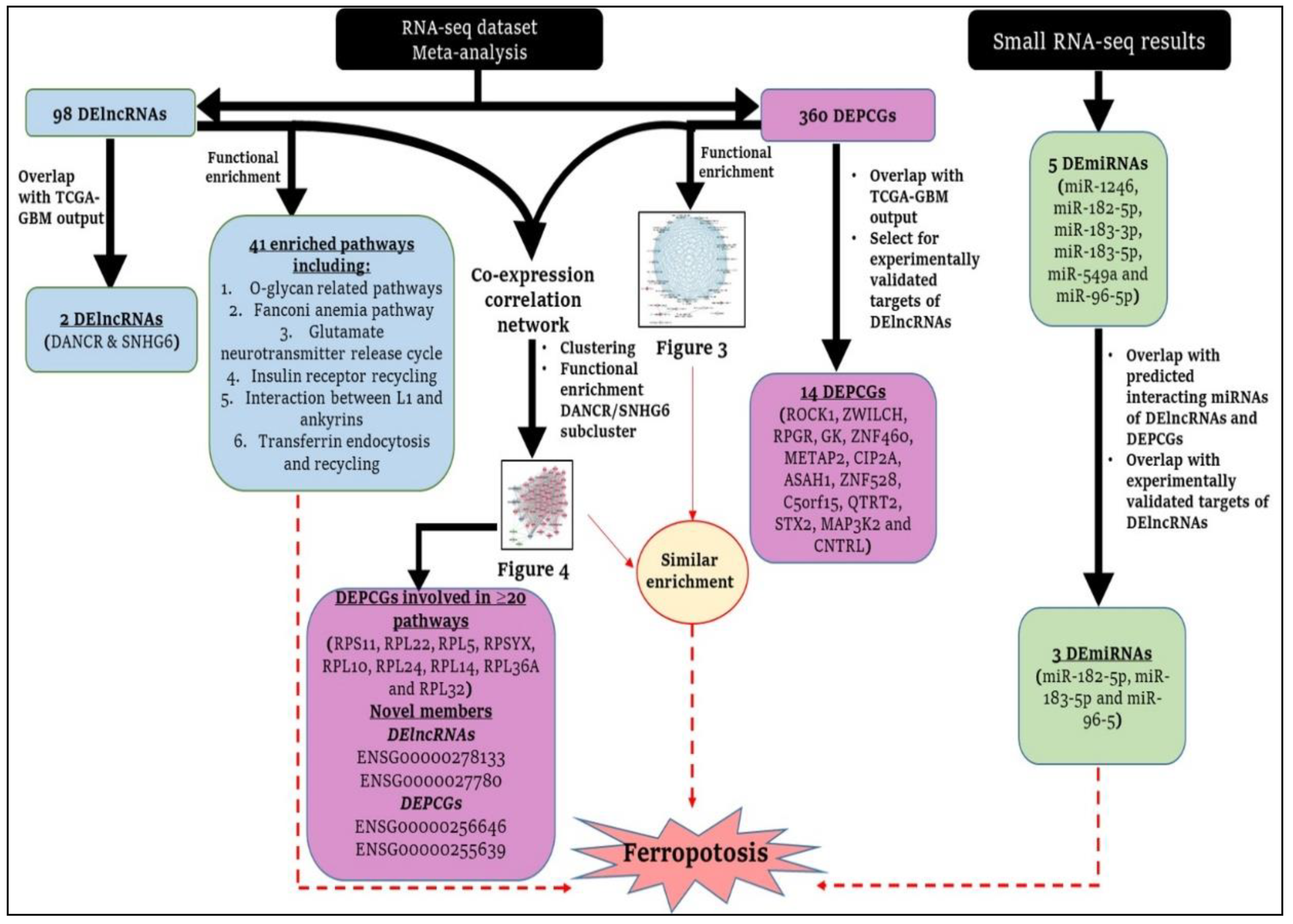
2. Materials and Methods
2.1. RNA-Seq and Small RNA-Seq Study Selection for Meta-Analysis
2.2. Quantification of Long Non-Coding RNA (lncRNA) and Protein Coding Gene (PCG) Sequencing Abundance Using RNA-Seq Data
2.3. Identification of Overlap between DElncRNAs/DEPCGs and lncRNAs/PCGs in Publicly Available Experimentally Verified Databases and TCGA-GBM Output
2.4. Pathway Analysis of DElncRNAs and DEPCGs
2.5. Co-Expression Analysis of DElncRNAs and DEPCGs and Identification of Highly Connected Nodes
2.6. Small RNA-Seq of Glioblastoma and Control Tissue Samples
2.7. Prediction of Interacting miRNAs of DElncRNAs and DEPCGs in Publicly Available Experimentally Verified Databases
3. Results
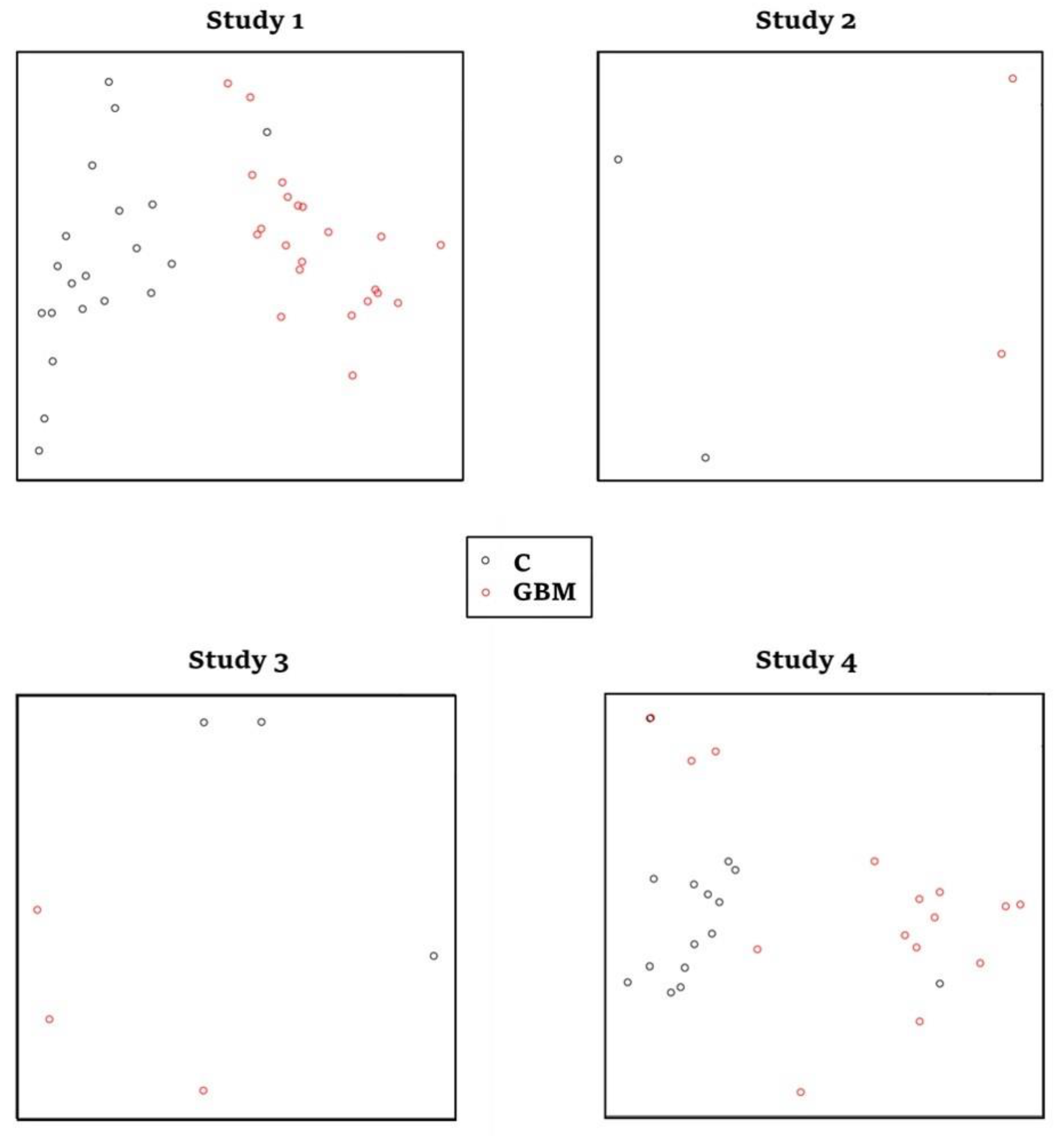
3.1. Four Glioblastoma RNA-Seq Datasets Were Selected for Meta-Analysis
3.2. Two DElncRNAs (DANCR and SNHG6) and 222 DEPCGs Were also Differentially Expressed in the TCGA-GBM Cohort
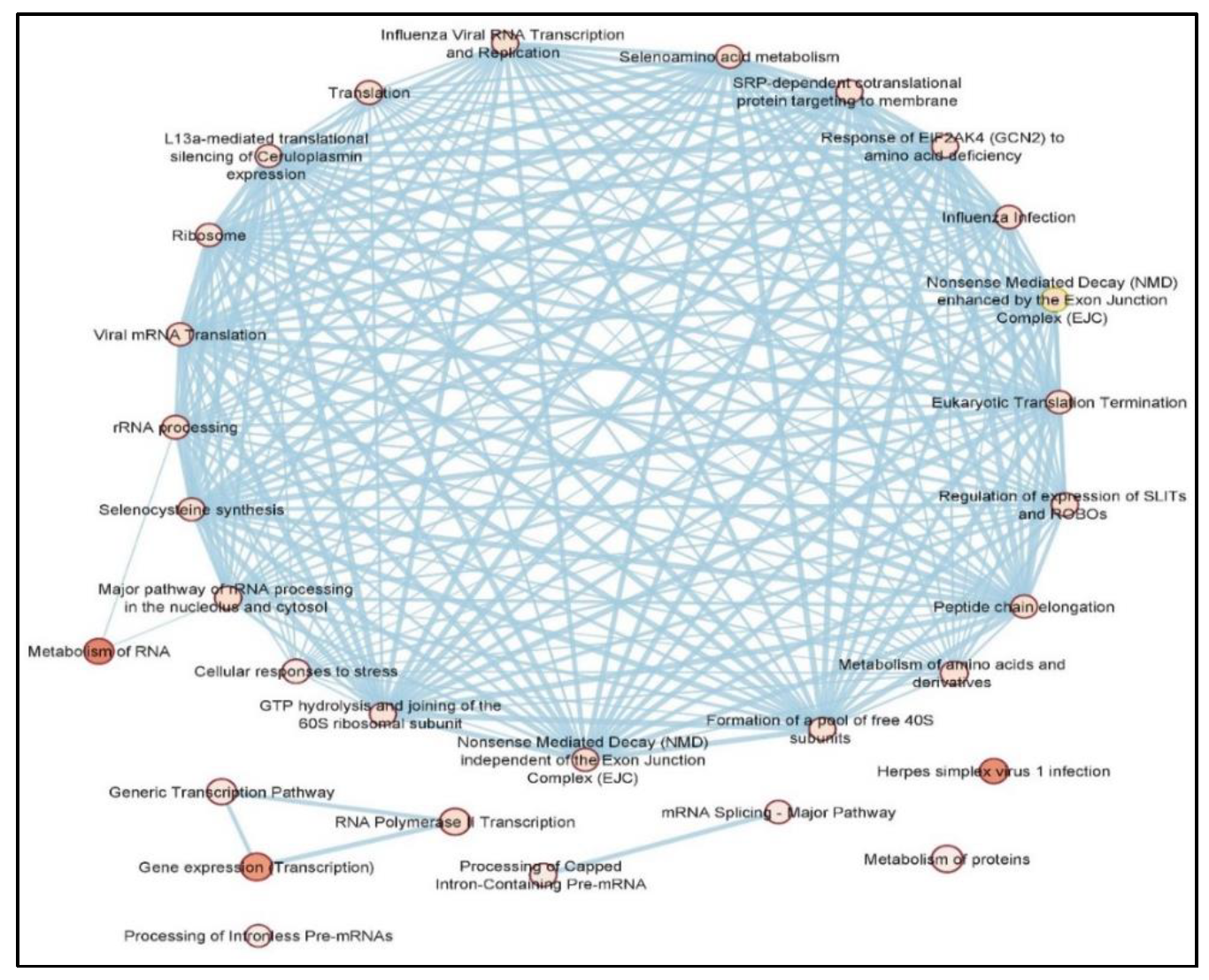
3.3. Pathway Analysis of DElncRNAs Reveals Several Glioblastoma-Associated Pathways
3.4. DEPCGs Show a Highly Connected PPI Network with Several Enriched Glioblastoma-Linked Pathways
3.5. Three DEmiRNAs Identified by Small RNA-Seq of Glioblastoma Tissue Overlap with Predicted DElncRNA and DEPCGs-Interacting miRNAs
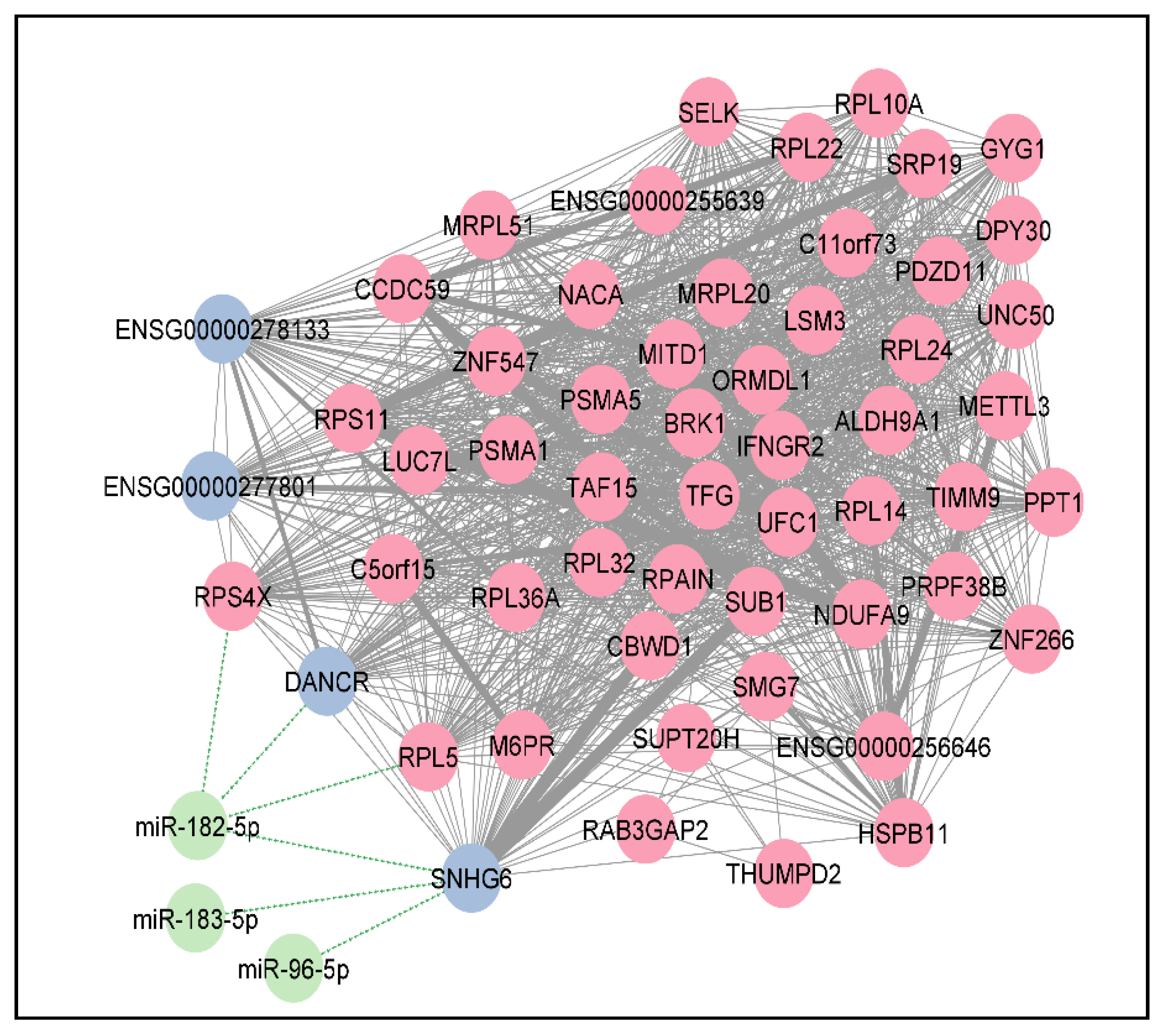
3.6. Co-Expression Analysis Identifies 4 Clusters of DElncRNAs/DEPCGs
4. Discussion
4.1. Meta-Analysis of Glioblastoma RNA-Seq Datasets
4.1.1. DElncRNAs
4.1.2. DEPCGs
4.2. Small RNA-Seq of Glioblastoma Tissues and Normal Controls
4.3. Overlap with Glioblastoma-Relevant Databases
4.4. Co-Expression Correlation Network Construction and Functional Enrichment
4.5. Literature-Based Associations of the Identified Pathways: Deducible Involvement of Ferroptosis?
4.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, A.P.; Fisher, J.L.; Nichols, E.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; Abraha, H.N.; Agius, D.; Alahdab, F.; Alam, T.; et al. Global, Regional, and National Burden of Brain and Other CNS Cancer, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Tamimi, A.F.; Juweid, M. Epidemiology and Outcome of Glioblastoma. In Glioblastoma; Codon Publications: Brisbane, Australia, 2017; pp. 143–153. [Google Scholar] [CrossRef]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of Glioblastoma: State of the Art and Future Directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of Its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3. [Google Scholar] [CrossRef]
- Krieger, T.G.; Tirier, S.M.; Park, J.; Jechow, K.; Eisemann, T.; Peterziel, H.; Angel, P.; Eils, R.; Conrad, C. Modeling Glioblastoma Invasion Using Human Brain Organoids and Single-Cell Transcriptomics. Neuro Oncol. 2020, 22, 1138–1149. [Google Scholar] [CrossRef] [Green Version]
- Jovčevska, I.; Zottel, A.; Šamec, N.; Paska, A.V. Coding of Glioblastoma Progression and Therapy Resistance through Long Noncoding RNAs. Cancers 2020, 12, 1842. [Google Scholar] [CrossRef]
- Rezaei, O.; Honarmand, K.; Nateghinia, S.; Taheri, M.; Ghafouri-Fard, S. MiRNA Signature in Glioblastoma: Potential Biomarkers and Therapeutic Targets. Exp. Mol. Pathol. 2020, 117, 104550. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, C.; Wu, M. New Insights into Long Noncoding RNAs and Their Roles in Glioma. Mol. Cancer 2018, 17, 61. [Google Scholar] [CrossRef]
- Rutenberg-Schoenberg, M.; Sexton, A.N.; Simon, M.D. The Properties of Long Noncoding RNAs That Regulate Chromatin. Annu. Rev. Genomics Hum. Genet. 2016, 17, 69–94. [Google Scholar] [CrossRef]
- Balas, M.M.; Johnson, A.M. Exploring the Mechanisms behind Long Noncoding RNAs and Cancer. Non-Coding RNA Res. 2018, 3, 108–117. [Google Scholar] [CrossRef]
- Yao, Q.; Chen, Y.; Zhou, X. The Roles of MicroRNAs in Epigenetic Regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kui, L.; Tang, M.; Li, D.; Wei, K.; Chen, W.; Miao, J.; Dong, Y. High-Throughput Transcriptome Profiling in Drug and Biomarker Discovery. Front. Genet. 2020, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.; Weber, D.J.; Njar, V.C.O. Transcriptome Profiling Reveals That VNPP433-3β, the Lead next-Generation Galeterone Analog Inhibits Prostate Cancer Stem Cells by Downregulating Epithelial–Mesenchymal Transition and Stem Cell Markers. Mol. Carcinog. 2022, 61, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.; Tseng, G.C.; Sun, W. Statistical Methods in Integrative Genomics. Annu. Rev. Stat. Its Appl. 2016, 3, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.; Ke, H.; Chen, S.; Cruz-Cano, R.; He, X.; Zhang, J.; Dorgan, J.; Milton, D.K.; Ma, T. Biomarker Categorization in Transcriptomic Meta-Analysis by Concordant Patterns With Application to Pan-Cancer Studies. Front. Genet. 2021, 12, 651546. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets--Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [Green Version]
- Gill, B.J.; Pisapia, D.J.; Malone, H.R.; Goldstein, H.; Lei, L.; Sonabend, A.; Yun, J.; Samanamud, J.; Sims, J.S.; Banu, M.; et al. MRI-Localized Biopsies Reveal Subtype-Specific Differences in Molecular and Cellular Composition at the Margins of Glioblastoma. Proc. Natl. Acad. Sci. USA 2014, 111, 12550–12555. [Google Scholar] [CrossRef] [Green Version]
- Stathias, V.; Pastori, C.; Griffin, T.Z.; Komotar, R.; Clarke, J.; Zhang, M.; Ayad, N.G.A. Identifying Glioblastoma Gene Networks Based on Hypergeometric Test Analysis. PLoS ONE 2014, 9, e115842. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Jiaoming, L.; Xiang, W.; Yanhui, L.; Shu, J.; Maling, G.; Qing, M. Analyzing the Interactions of MRNAs, MiRNAs, LncRNAs and CircRNAs to Predict Competing Endogenous RNA Networks in Glioblastoma. J. Neurooncol. 2018, 137, 493–502. [Google Scholar] [CrossRef]
- Hwang, T.; Kim, S.; Chowdhury, T.; Yu, H.J.; Kim, K.M.; Kang, H.; Won, J.K.; Park, S.H.; Shin, J.H.; Park, C.K. Genome-Wide Perturbations of Alu Expression and Alu-Associated Post-Transcriptional Regulations Distinguish Oligodendroglioma from Other Gliomas. Commun. Biol. 2022, 5, 62. [Google Scholar] [CrossRef]
- Leinonen, R.; Sugawara, H.; Shumway, M. The Sequence Read Archive. Nucleic Acids Res. 2011, 39, D19–D21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Hua, J.; Xue, W.; Zhu, F. Integrating the Ribonucleic Acid Sequencing Data From Various Studies for Exploring the Multiple Sclerosis-Related Long Noncoding Ribonucleic Acids and Their Functions. Front. Genet. 2019, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Zhang, L.; Guo, J.; Niu, Y.; Wu, Y.; Li, H.; Zhao, L.; Li, X.; Teng, X.; Sun, X.; et al. NONCODEV5: A Comprehensive Annotation Database for Long Non-Coding RNAs. Nucleic Acids Res. 2018, 46, D308–D314. [Google Scholar] [CrossRef]
- Aken, B.L.; Achuthan, P.; Akanni, W.; Amode, M.R.; Bernsdorff, F.; Bhai, J.; Billis, K.; Carvalho-Silva, D.; Cummins, C.; Clapham, P.; et al. Ensembl 2017. Nucleic Acids Res. 2017, 45, D635–D642. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.H.; Kim, S.Y.; Jo, D.; Park, H.J.; Kim, J.; Jung, S.; Kim, H.S.; Lee, K.Y. Meta-Analysis of Large-Scale Toxicogenomic Data Finds Neuronal Regeneration Related Protein and Cathepsin D to Be Novel Biomarkers of Drug-Induced Toxicity. PLoS ONE 2015, 10, e0136698. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H.; Kołodziej, M. Systematic Review with Meta-Analysis: Saccharomyces Boulardii in the Prevention of Antibiotic-Associated Diarrhoea. Aliment. Pharmacol. Ther. 2015, 42, 793–801. [Google Scholar] [CrossRef]
- Zhao, H.; Shi, J.; Zhang, Y.; Xie, A.; Yu, L.; Zhang, C.; Lei, J.; Xu, H.; Leng, Z.; Li, T.; et al. LncTarD: A Manually-Curated Database of Experimentally-Supported Functional LncRNA–Target Regulations in Human Diseases. Nucleic Acids Res. 2020, 48, D118–D126. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, P.; Tian, R.; Wang, S.; Guo, Q.; Luo, M.; Zhou, W.; Liu, G.; Jiang, H.; Jiang, Q. LncRNA2Target v2.0: A Comprehensive Database for Target Genes of LncRNAs in Human and Mouse. Nucleic Acids Res. 2019, 47, D140–D144. [Google Scholar] [CrossRef]
- Tang, G.; Cho, M.; Wang, X. OncoDB: An Interactive Online Database for Analysis of Gene Expression and Viral Infection in Cancer. Nucleic Acids Res. 2022, 50, D1334–D1339. [Google Scholar] [CrossRef]
- Han, J.; Liu, S.; Sun, Z.; Zhang, Y.; Zhang, F.; Zhang, C.; Shang, D.; Yang, H.; Su, F.; Xu, Y.; et al. LncRNAs2Pathways: Identifying the Pathways Influenced by a Set of LncRNAs of Interest Based on a Global Network Propagation Method. Sci. Rep. 2017, 7, 46566. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating Viruses and Cellular Organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Jensen, L.J.; et al. The STRING Database in 2021: Customizable Protein-Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2021, 49, 605–612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BE 71030031000; BBMR-ERIC, Belgian [BIORESOURCE]. Biobank Antwerp [BB190007]: Antwerp, Belgium, 2022.
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Kull, M.; Peterson, H.; Hansen, J.; Vilo, J. G:Profiler—A Web-Based Toolset for Functional Profiling of Gene Lists from Large-Scale Experiments. Nucleic Acids Res. 2007, 35, W193–W200. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Tastsoglou, S.; Skoufos, G.; Karavangeli, A.; Pierros, V.; Zacharopoulou, E.; Hatzigeorgiou, A.G. DIANA-LncBase v3: Indexing Experimentally Supported MiRNA Targets on Non-Coding Transcripts. Nucleic Acids Res. 2020, 48, D101–D110. [Google Scholar] [CrossRef]
- Hsu, S.-D.; Lin, F.M.; Wu, W.Y.; Liang, C.; Huang, W.C.; Chan, W.L.; Tsai, W.T.; Chen, G.Z.; Lee, C.J.; Chiu, C.M.; et al. MiRTarBase: A Database Curates Experimentally Validated MicroRNA–Target Interactions. Nucleic Acids Res. 2011, 39, D163–D169. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Lin, Y.C.D.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. MiRTarBase Update 2022: An Informative Resource for Experimentally Validated MiRNA–Target Interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Tondepu, C.; Karumbaiah, L. Glycomaterials to Investigate the Functional Role of Aberrant Glycosylation in Glioblastoma. Adv. Healthc. Mater. 2022, 11, 2101956. [Google Scholar] [CrossRef] [PubMed]
- Dan, C.; Pei, H.; Zhang, B.; Zheng, X.; Ran, D.; Du, C. Fanconi Anemia Pathway and Its Relationship with Cancer. Genome Instab. Dis. 2021, 2, 175–183. [Google Scholar] [CrossRef]
- de Souza Fonseca, P.A.; Suárez-Vega, A.; Cánovas, A. Unrevealing Functional Candidate Genes for Bovine Fertility through RNA Sequencing Meta-Analysis and Regulatory Elements Networks of Co-Expressed Genes and LncRNAs. Funct. Integr. Genomics 2022. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.W.-L.; Vijayakumar, R. A Guide to Conducting a Meta-Analysis. Neuropsychol. Rev. 2016, 26, 121–128. [Google Scholar] [CrossRef]
- Liu, D.-H.; Yang, X.; Guo, J.-F.; Meng, H.; Shen, S.-H. Immune-Related LncRNAs, LINC01268 and CTB-31O20. 2, as Favorable Prognostic Markers for Glioma Inhibition. Transl. Cancer Res. 2022, 11, 823–834. [Google Scholar] [CrossRef]
- Brockhausen, I. Pathways of O-Glycan Biosynthesis in Cancer Cells. Biochim. Biophys. Acta-Gen. Subj. 1999, 1473, 67–95. [Google Scholar] [CrossRef]
- Cervoni, G.E.; Cheng, J.J.; Stackhouse, K.A.; Heimburg-Molinaro, J.; Cummings, R.D. O-Glycan Recognition and Function in Mice and Human Cancers. Biochem. J. 2020, 477, 1541–1564. [Google Scholar] [CrossRef]
- Kudelka, M.R.; Ju, T.; Heimburg-Molinaro, J.; Cummings, R.D. Simple Sugars to Complex Disease—Mucin-Type O-Glycans in Cancer. Adv. Cancer Res. 2015, 126, 53–135. [Google Scholar]
- Rominiyi, O.; Myers, K.; Gomez-Roman, N.; Lad, N.; Dar, D.; Jellinek, D.; Chalmers, A.; Carroll, T.; Chen, B.; Al-Tamimi, Y.; et al. RDNA-12. The fanconi anaemia (FA) pathway and glioblastoma: A new foundation for dna damage response targeted combinations. Neuro Oncol. 2019, 21, vi209. [Google Scholar] [CrossRef]
- de Groot, J.; Sontheimer, H. Glutamate and the Biology of Gliomas. Glia 2011, 59, 1181–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Yuan, X.; Li, H.; Zhao, Z.; Liao, Y.; Wang, X.; Su, J.; Sang, S.; Liu, Q. Anti-Cancer Effect of Metabotropic Glutamate Receptor 1 Inhibition in Human Glioma U87 Cells: Involvement of PI3K/Akt/MTOR Pathway. Cell Physiol. Biochem. 2015, 35, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Tirrò, E.; Massimino, M.; Romano, C.; Martorana, F.; Pennisi, M.S.; Stella, S.; Pavone, G.; Di Gregorio, S.; Puma, A.; Tomarchio, C.; et al. Prognostic and Therapeutic Roles of the Insulin Growth Factor System in Glioblastoma. Front. Oncol. 2021, 10, 612385. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Wu, Q.; Li, Z.; Sathornsumetee, S.; Wang, H.; McLendon, R.E.; Hjelmeland, A.B.; Rich, J.N. Targeting Cancer Stem Cells through L1CAM Suppresses Glioma Growth. Cancer Res. 2008, 68, 6043–6048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachowiak, R.; Krause, M.; Mayer, S.; Peukert, N.; Suttkus, A.; Müller, W.C.; Lacher, M.; Meixensberger, J.; Nestler, U. Increased L1CAM (CD171) Levels Are Associated with Glioblastoma and Metastatic Brain Tumors. Medicine 2018, 97, e12396. [Google Scholar] [CrossRef] [PubMed]
- Koneru, T.; McCord, E.; Pawar, S.; Tatiparti, K.; Sau, S.; Iyer, A.K. Transferrin: Biology and Use in Receptor-Targeted Nanotherapy of Gliomas. ACS Omega 2021, 6, 8727–8733. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Ye, L. The Interferon-γ Receptor Pathway: A New Way to Regulate CAR T Cell-Solid Tumor Cell Adhesion. Signal Transduct. Target. Ther. 2022, 7, 315. [Google Scholar] [CrossRef]
- Nayani, R.; Ashktorab, H.; Brim, H.; Laiyemo, A.O. Genetic Basis for Colorectal Cancer Disparities. Curr. Colorectal Cancer Rep. 2015, 11, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Scholz, B.A.; Sumida, N.; de Lima, C.D.M.; Chachoua, I.; Martino, M.; Tzelepis, I.; Nikoshkov, A.; Zhao, H.; Mehmood, R.; Sifakis, E.G.; et al. WNT Signaling and AHCTF1 Promote Oncogenic MYC Expression through Super-Enhancer-Mediated Gene Gating. Nat. Genet. 2019, 51, 1723–1731. [Google Scholar] [CrossRef]
- Ma, Z.-H.; Shi, P.-D.; Wan, B.-S. MiR-410-3p Activates the NF-ΚB Pathway by Targeting ZCCHC10 to Promote Migration, Invasion and EMT of Colorectal Cancer. Cytokine 2021, 140, 155433. [Google Scholar] [CrossRef]
- Mehmood, R.; Jibiki, K.; Alsafwani, Z.J.; Naseem, M.; Yasuhara, N. Systems Genomics of Nucleoporins Provides Prognostic Insights into Breast Cancer. Adv. Life Sci. 2022, 9, 98. [Google Scholar]
- Dadey, D.Y.A.; Kapoor, V.; Khudanyan, A.; Urano, F.; Kim, A.H.; Thotala, D.; Hallahan, D.E. The ATF6 Pathway of the ER Stress Response Contributes to Enhanced Viability in Glioblastoma. Oncotarget 2016, 7, 2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, M.W.; Maquat, L.E. Nonsense-Mediated MRNA Decay and Cancer. Curr. Opin. Genet. Dev. 2018, 48, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Gudikote, J.P.; Cascone, T.; Poteete, A.; Sitthideatphaiboon, P.; Wu, Q.; Morikawa, N.; Zhang, F.; Peng, S.; Tong, P.; Li, L.; et al. Inhibition of Nonsense-Mediated Decay Rescues P53β/γ Isoform Expression and Activates the P53 Pathway in MDM2-Overexpressing and Select P53-Mutant Cancers. J. Biol. Chem. 2021, 297, 101163. [Google Scholar] [CrossRef]
- McCarthy, R.C.; Kosman, D.J. Glial Cell Ceruloplasmin and Hepcidin Differentially Regulate Iron Efflux from Brain Microvascular Endothelial Cells. PLoS ONE 2014, 9, e89003. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, R.C.; Kosman, D.J. Activation of C6 Glioblastoma Cell Ceruloplasmin Expression by Neighboring Human Brain Endothelia-Derived Interleukins in an in Vitro Blood-Brain Barrier Model System. Cell Commun. Signal. 2014, 12, 65. [Google Scholar] [CrossRef]
- Fan, C.D.; Fu, X.Y.; Zhang, Z.Y.; Cao, M.Z.; Sun, J.Y.; Yang, M.F.; Fu, X.T.; Zhao, S.J.; Shao, L.R.; Zhang, H.F.; et al. Selenocysteine Induces Apoptosis in Human Glioma Cells: Evidence for TrxR1-Targeted Inhibition and Signaling Crosstalk. Sci. Rep. 2017, 7, 6465. [Google Scholar] [CrossRef] [Green Version]
- Carlisle, A.E.; Lee, N.; Matthew-Onabanjo, A.N.; Spears, M.E.; Park, S.J.; Youkana, D.; Doshi, M.B.; Peppers, A.; Li, R.; Joseph, A.B.; et al. Selenium Detoxification Is Required for Cancer Cell. Nat. Metab. 2020, 2, 603. [Google Scholar] [CrossRef]
- Gara, R.K.; Kumari, S.; Ganju, A.; Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Slit/Robo Pathway: A Promising Therapeutic Target for Cancer. Drug Discov. Today 2015, 20, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Yiin, J.-J.; Hu, B.; Jarzynka, M.J.; Feng, H.; Liu, K.-W.; Wu, J.Y.; Ma, H.-I.; Cheng, S.-Y. Slit2 Inhibits Glioma Cell Invasion in the Brain by Suppression of Cdc42 Activity. Neuro Oncol. 2009, 11, 779–789. [Google Scholar] [CrossRef]
- Geraldo, L.H.; Xu, Y.; Jacob, L.; Pibouin-Fragner, L.; Rao, R.; Maissa, N.; Verreault, M.; Lemaire, N.; Knosp, C.; Lesaffre, C.; et al. SLIT2/ROBO Signaling in Tumor-Associated Microglia and Macrophages Drives Glioblastoma Immunosuppression and Vascular Dysmorphia. J. Clin. Investig. 2021, 131, e141083. [Google Scholar] [CrossRef] [PubMed]
- Alasiri, G.; Jiramongkol, Y.; Trakansuebkul, S.; Ke, H.L.; Mahmud, Z.; Intuyod, K.; Lam, E.W.F. Reciprocal Regulation between GCN2 (EIF2AK4) and PERK (EIF2AK3) through the JNK-FOXO3 Axis to Modulate Cancer Drug Resistance and Clonal Survival. Mol. Cell Endocrinol. 2020, 515, 110932. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ning, Y.; Alam, G.N.; Jankowski, B.M.; Dong, Z.; Nör, J.E.; Polverini, P.J. Amino Acid Deprivation Promotes Tumor Angiogenesis through the GCN2/ATF4 Pathway. Neoplasia 2013, 15, 989–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; He, J.; Zhou, J.; Xiao, Z.; Ding, N.; Duan, Y.; Li, W.; Sun, L.Q. EIF2A Promotes Cell Survival during Paclitaxel Treatment in Vitro and in Vivo. J. Cell Mol. Med. 2019, 23, 6060. [Google Scholar] [CrossRef] [PubMed]
- Carballo, G.B.; Honorato, J.R.; de Lopes, G.P.F.; de Sampaio e Spohr, T.C.L. A Highlight on Sonic Hedgehog Pathway. Cell Commun. Signal. 2018, 16, 11. [Google Scholar] [CrossRef]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The Role of the Hedgehog Signaling Pathway in Cancer: A Comprehensive Review. Bosn. J. Basic Med. Sci. 2018, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Hanna, A.; Shevde, L.A. Hedgehog Signaling: Modulation of Cancer Properies and Tumor Mircroenvironment. Mol. Cancer 2016, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Hung, H.C.; Liu, C.C.; Chuang, J.Y.; Su, C.L.; Gean, P.W. Inhibition of Sonic Hedgehog Signaling Suppresses Glioma Stem-Like Cells Likely Through Inducing Autophagic Cell Death. Front. Oncol. 2020, 10, 1233. [Google Scholar] [CrossRef]
- Melamed, J.R.; Morgan, J.T.; Ioele, S.A.; Gleghorn, J.P.; Sims-Mourtada, J.; Day, E.S. Investigating the Role of Hedgehog/GLI1 Signaling in Glioblastoma Cell Response to Temozolomide. Oncotarget 2018, 9, 27000. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.; Guo, X.; Li, B.; Wang, J.; Qi, Y.; Chen, Z.; Zhao, R.; Deng, L.; Qian, M.; Wang, S.; et al. Exosomal MiR-1246 from Glioma Patient Body Fluids Drives the Differentiation and Activation of Myeloid-Derived Suppressor Cells. Mol. Ther. 2021, 29, 3449–3464. [Google Scholar] [CrossRef]
- Li, J.; Yuan, H.; Xu, H.; Zhao, H.; Xiong, N. Hypoxic Cancer-Secreted Exosomal MiR-182-5p Promotes Glioblastoma Angiogenesis by Targeting Kruppel-like Factor 2 and 4. Mol. Cancer Res. 2020, 18, 1218–1231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qin, W.; Yang, S.; Guan, N.; Sui, X.; Guo, W. Circular RNA SFMBT2 Inhibits the Proliferation and Metastasis of Glioma Cells Through Mir-182-5p/Mtss1 Pathway. Technol. Cancer Res. Treat. 2020, 19, 1533033820945799. [Google Scholar] [CrossRef] [PubMed]
- Donatelli, S.S.; Zhou, J.M.; Gilvary, D.L.; Eksioglu, E.A.; Chen, X.; Cress, W.D.; Haura, E.B.; Schabath, M.B.; Coppola, D.; Wei, S.; et al. TGF-β-Inducible MicroRNA-183 Silences Tumor-Associated Natural Killer Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 4203–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazi, B.; Felsani, A.; Grassi, L.; Moles, A.; D’Andrea, D.; Toschi, N.; Sicari, D.; De Bonis, P.D.; Anile, C.; Guerrisi, M.G.; et al. The Transcriptome and MiRNome Profiling of Glioblastoma Tissues and Peritumoral Regions Highlights Molecular Pathways Shared by Tumors and Surrounding Areas and Reveals Differences between Short-Term and Long-Term Survivors. Oncotarget 2015, 6, 22526–22552. [Google Scholar] [CrossRef] [Green Version]
- Pavlakis, E.; Tonchev, A.B.; Kaprelyan, A.; Enchev, Y.; Stoykova, A. Interaction between Transcription Factors PAX6/PAX6-5a and Specific Members of MiR-183-96-182 Cluster, May Contribute to Glioma Progression in Glioblastoma Cell Lines. Oncol. Rep. 2017, 37, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Guo, J.; Liu, W.; Hu, S.; Hu, X.; Wang, Q.; Jiang, X. Circ-EGFR Functions as an Inhibitory Factor in the Malignant Progression of Glioma by Regulating the MiR-183-5p/TUSC2 Axis. Cell Mol. Neurobiol. 2021, 42, 2245–2256. [Google Scholar] [CrossRef]
- Drusco, A.; Fadda, P.; Nigita, G.; Fassan, M.; Bottoni, A.; MP, G.; Sacchi, D.; Calore, F.; Carosi, M.; Antenucci, A.; et al. Circulating Micrornas Predict Survival of Patients with Tumors of Glial Origin. EBioMedicine 2018, 30, 105–112. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, W. Long Non-Coding RNA MEG3 Suppresses the Growth of Glioma Cells by Regulating the MiR-96-5p/MTSS1 Signaling Pathway. Mol. Med. Rep. 2019, 20, 4215–4225. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Yu, Y.; Tian, Z.; Lin, Y.; Qiu, Y.; Yao, W.; Zhang, L. Upregulation of MiR-96 Promotes Radioresistance in Glioblastoma Cells via Targeting PDCD4. Int. J. Oncol. 2018, 53, 1591–1600. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Bian, Y.; Tu, C.; Wang, Z.; Yu, Z.; Liu, Q.; Xu, G.; Wu, M.; Li, G. The MiR-183/96/182 Cluster Regulates Oxidative Apoptosis and Sensitizes Cells to Chemotherapy in Gliomas. Curr. Cancer Drug Targets 2013, 13, 221–231. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Mukhanbetzhanovna, A.A.; Yilmaz, S.; Karasholakova, L.; Yulaevna, I.M. Mechanistic Role of DANCR in the Choreography of Signaling Pathways in Different Cancers: Spotlight on Regulation of Wnt/β-Catenin and JAK/STAT Pathways by Oncogenic Long Non-Coding RNA. Non-Coding RNA Res. 2021, 6, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.J.; Jin, M.Z.; Xia, B.R.; Jin, W.L. Long Non-Coding RNA DANCR as an Emerging Therapeutic Target in Human Cancers. Front. Oncol. 2019, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Thin, K.Z.; Liu, X.; Feng, X.; Raveendran, S.; Tu, J.C. LncRNA-DANCR: A Valuable Cancer Related Long Non-Coding RNA for Human Cancers. Pathol.-Res. Pract. 2018, 214, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, L. Overexpression of LncRNA DANCR Positively Affects Progression of Glioma via Activating Wnt/β-Catenin Signaling. Biomed. Pharmacother. 2018, 102, 602–607. [Google Scholar] [CrossRef]
- Tan, Z.; Zhao, J.; Jiang, Y. MiR-634 Sensitizes Glioma Cells to Temozolomide by Targeting CYR61 through Raf-ERK Signaling Pathway. Cancer Med. 2018, 7, 913. [Google Scholar] [CrossRef]
- Xu, D.; Yu, J.; Gao, G.; Lu, G.; Zhang, Y.; Ma, P. LncRNA DANCR Functions as a Competing Endogenous RNA to Regulate RAB1A Expression by Sponging MIR-634 in Glioma. Biosci. Rep. 2018, 38, 20171664. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Zhou, G.; Li, M.; Hu, D.; Zhang, L.; Liu, P.; Lin, K. Long Noncoding RNA DANCR Mediates Cisplatin Resistance in Glioma Cells via Activating AXL/PI3K/Akt/NF-ΚB Signaling Pathway. Neurochem. Int. 2018, 118, 233–241. [Google Scholar] [CrossRef]
- Zhang, Y.; An, J.; Pei, Y. LncRNA SNHG6 Promotes LMO3 Expression by Sponging MiR-543 in Glioma. Mol. Cell Biochem. 2020, 472, 9–17. [Google Scholar] [CrossRef]
- Meng, Q.; Yang, B.Y.; Liu, B.; Yang, J.X.; Sun, Y. Long Non-Coding RNA SNHG6 Promotes Glioma Tumorigenesis by Sponging MiR-101-3p. Int. J. Biol. Markers 2018, 33, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, F.; Ma, J.; Ruan, X.; Liu, X.; Zheng, J.; Liu, Y.; Cao, S.; Shen, S.; Shao, L.; et al. NCBP3/SNHG6 Inhibits GBX2 Transcription in a Histone Modification Manner to Facilitate the Malignant Biological Behavior of Glioma Cells. RNA Biol. 2020, 18, 47–63. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, R.; Zhang, Y.; Matuszek, Ż.; Zhang, W.; Xia, Y.; Pan, T.; Sun, J. TRNA Queuosine Modification Enzyme Modulates the Growth and Microbiome Recruitment to Breast Tumors. Cancers 2020, 12, 628. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fang, Y.; Ma, L.; Xu, J.; Lv, C.; Deng, L.; Zhu, G. LINC00857 Regulated by ZNF460 Enhances the Expression of CLDN12 by Sponging MiR-150-5p and Recruiting SRSF1 for Alternative Splicing to Promote Epithelial-Mesenchymal Transformation of Pancreatic Adenocarcinoma Cells. RNA Biol. 2022, 19, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.; Xu, J.; Fang, S.; Jiang, J.; Chen, X.; Wu, W.; Li, L.; Li, M.; Zhang, C.; He, Y. Overexpression of ZNF460 Predicts Worse Survival and Promotes Metastasis through JAK2/STAT3 Signaling Pathway in Patient with Colon Cancer. J. Cancer 2021, 12, 3198. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Li, X.; Lv, X.; Wu, S.; Song, J.; Tang, Z.; Luo, H.; Wei, H.; Ai, Y. Circular RNA Mitochondrial Translation Optimization 1 Homologue (CircMTO1) Induced by Zinc Finger Protein 460 (ZNF460) Promotes Oral Squamous Cell Carcinoma Progression through the MicroRNA MiR-320a / Alpha Thalassemia/Mental Retardation, X-Linked (ATRX) Axis. Bioengineered 2021, 12, 9585–9597. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liyanarachchi, S.; Miller, K.E.; Nieminen, T.T.; Comiskey, D.F.; Li, W.; Brock, P.; Symer, D.E.; Akagi, K.; Delap, K.E.; et al. Identification of Rare Variants Predisposing to Thyroid Cancer. Thyroid 2019, 29, 946. [Google Scholar] [CrossRef]
- Puchalski, R.B.; Shah, N.; Miller, J.; Dalley, R.; Nomura, S.R.; Yoon, J.-G.; Smith, K.A.; Lankerovich, M.; Bertagnolli, D.; Bickley, K. An Anatomic Transcriptional Atlas of Human Glioblastoma. Science 2018, 360, 660–663. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, K.-N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genomics Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Awah, C.U.; Chen, L.; Bansal, M.; Mahajan, A.; Winter, J.; Lad, M.; Warnke, L.; Gonzalez-Buendia, E.; Park, C.; Zhang, D. Ribosomal Protein S11 Influences Glioma Response to TOP2 Poisons. Oncogene 2020, 39, 5068–5081. [Google Scholar] [CrossRef]
- Yong, W.H.; Shabihkhani, M.; Telesca, D.; Yang, S.; Tso, J.L.; Menjivar, J.C.; Wei, B.; Lucey, G.M.; Mareninov, S.; Chen, Z.; et al. Ribosomal Proteins RPS11 and RPS20, Two Stress-Response Markers of Glioblastoma Stem Cells, Are Novel Predictors of Poor Prognosis in Glioblastoma Patients. PLoS ONE 2015, 10, e0141334. [Google Scholar] [CrossRef]
- Zhao, S.; Ji, W.; Shen, Y.; Fan, Y.; Huang, J.; Huang, H.; Cheng, C.; Shao, J. Expression of Hub Genes of Endothelial Cells in Glioblastoma—A Prognostic Model for GBM Patients Integrating Single Cell RNA Sequencing and Bulk RNA Sequencing; Research Square: Durham, NC, USA, 2022. [Google Scholar]
- Li, R.; Jiang, Q.; Tang, C.; Chen, L.; Kong, D.; Zou, C.; Lin, Y.; Luo, J.; Zou, D. Identification of Candidate Genes Associated With Prognosis in Glioblastoma. Front. Mol. Neurosci. 2022, 15, 913328. [Google Scholar] [CrossRef]
- Li, S.; Shi, J.; Gao, H.; Yuan, Y.; Chen, Q.; Zhao, Z.; Wang, X.; Li, B.; Ming, L.; Zhong, J.; et al. Identification of a Gene Signature Associated with Radiotherapy and Prognosis in Gliomas. Oncotarget 2017, 8, 88974–88987. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, C.; Peng, Y.; Liu, S. Ferroptosis Biology and Implication in Cancers. Front. Mol. Biosci. 2022, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in Cancer Therapy: The Bright Side of the Moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- De Souza, I.; Carolina, M.; Ramalho, C.; Guedes, C.B.; Yumi, I.; Osawa, A.; Karolynne, L.; Monteiro, S.; Gomes, L.R.; Ribeiro, C.; et al. Ferroptosis Modulation: Potential Therapeutic Target for Glioblastoma Treatment. Int. J. Mol. Sci. 2022, 23, 6879. [Google Scholar] [CrossRef]
- Yuan, F.; Sun, Q.; Zhang, S.; Ye, L.; Xu, Y.; Xu, Z.; Liu, B.; Zhang, S.; Chen, Q. HSP27 Protects against Ferroptosis of Glioblastoma Cells. Hum. Cell 2022, 35, 238–249. [Google Scholar] [CrossRef]
- Song, Q.; Peng, S.; Sun, Z.; Heng, X.; Zhu, X. Temozolomide Drives Ferroptosis via a DMT1-Dependent Pathway in Glioblastoma Cells. Yonsei Med. J. 2021, 62, 843. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Q.; Liu, H.; Liu, J.; Yang, S.; Luo, X.; Liu, W.; Zheng, H.; Liu, Q.; Cui, Y.; et al. Dynamic O-GlcNAcylation Coordinates Ferritinophagy and Mitophagy to Activate Ferroptosis. Cell Discov. 2022, 8, 40. [Google Scholar] [CrossRef]
- Dixon, S.J.; Patel, D.N.; Welsch, M.; Skouta, R.; Lee, E.D.; Hayano, M.; Thomas, A.G.; Gleason, C.E.; Tatonetti, N.P.; Slusher, B.S. Pharmacological Inhibition of Cystine–Glutamate Exchange Induces Endoplasmic Reticulum Stress and Ferroptosis. Elife 2014, 3, e02523. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, K.; Ma, L.; Qian, Z.; Tian, X.; Miao, Y.; Niu, Y.; Xu, X.; Guo, S.; Yang, Y. Endogenous Glutamate Determines Ferroptosis Sensitivity via ADCY10-Dependent YAP Suppression in Lung Adenocarcinoma. Theranostics 2021, 11, 5650. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Quadri, N.; Ramasamy, R.; Jiang, X. Glutaminolysis and Transferrin Regulate Ferroptosis. Mol. Cell 2015, 59, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in Cancer Therapy: A Novel Approach to Reversing Drug Resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Luo, M.; Yao, F.; Wang, S.; Yuan, Z.; Yang, Y. Ceruloplasmin Suppresses Ferroptosis by Regulating Iron Homeostasis in Hepatocellular Carcinoma Cells. Cell Signal. 2020, 72, 109633. [Google Scholar] [CrossRef] [PubMed]
- Ryan, F.; Zarruk, J.G.; Lößlein, L.; David, S. Ceruloplasmin Plays a Neuroprotective Role in Cerebral Ischemia. Front. Neurosci. 2019, 13, 988. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Bai, L.; Fang, X.; Liu, J.; Kang, R.; Zhou, D.; Tang, D.; Dai, E. SMG9 Drives Ferroptosis by Directly Inhibiting GPX4 Degradation. Biochem. Biophys. Res. Commun. 2021, 567, 92–98. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.-L.; Huang, Z.-J.; Lin, Z.-T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, Present and Future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Poltorack, C.D.; Dixon, S.J. Understanding the Role of Cysteine in Ferroptosis: Progress & Paradoxes. FEBS J. 2022, 289, 374–385. [Google Scholar] [CrossRef]
- Nathaniel Roybal, C.; Hunsaker, L.A.; Barbash, O.; Vander Jagt, D.L.; Abcouwer, S.F. The Oxidative Stressor Arsenite Activates Vascular Endothelial Growth Factor MRNA Transcription by an ATF4-Dependent Mechanism. J. Biol. Chem. 2005, 280, 20331–20339. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Yan, J.; Ma, H.; Wu, J.; Zhang, Y. Autophagy-Dependent Ferroptosis-Related Signature Is Closely Associated with the Prognosis and Tumor Immune Escape of Patients with Glioma. Int. J. Gen. Med. 2022, 15, 253. [Google Scholar] [CrossRef]
- López-Janeiro, Á.; Ruz-Caracuel, I.; Ramón-Patino, J.L.; Ríos, V.D.L.; Esparza, M.V.; Berjón, A.; Yébenes, L.; Hernández, A.; Masetto, I.; Kadioglu, E.; et al. Proteomic Analysis of Low-Grade, Early-Stage Endometrial Carcinoma Reveals New Dysregulated Pathways Associated with Cell Death and Cell Signaling. Cancers 2021, 13, 794. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Y.; Fan, M.; Ren, H.; Chen, M.; Shi, P. Identification of the Ferroptosis-Related Long Non-Coding RNAs Signature to Improve the Prognosis Prediction and Immunotherapy Response in Patients with NSCLC. BMC Med. Genomics 2021, 14, 286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, R.; Lei, X.; Mao, L.; Yin, Z.; Zhong, X.; Cao, W.; Zheng, Q.; Li, D. Comprehensive Analysis of a Ferroptosis-Related LncRNA Signature for Predicting Prognosis and Immune Landscape in Osteosarcoma. Front. Oncol. 2022, 12, 880459. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Ding, X.; Zheng, J.; Wang, B.; Li, Y.; Xiang, H.; Dou, M.; Qiao, Y.; Tian, P.; Xue, W. MiR-182-5p and MiR-378a-3p Regulate Ferroptosis in I/R-Induced Renal Injury. Cell Death Dis. 2020, 11, 929. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Sun, X. Construction of a Ferroptosis-Associated CircRNA-MiRNA-MRNA Network in Age-Related Macular Degeneration. Exp. Eye Res. 2022, 224, 109234. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.-J.; Zhou, Y.-J.; Xiong, X.-M.; Li, N.-S.; Zhang, J.-J.; Luo, X.-J.; Peng, J. Ubiquitin-Specific Protease 7 Promotes Ferroptosis via Activation of the P53/TfR1 Pathway in the Rat Hearts after Ischemia/Reperfusion. Free Radic. Biol. Med. 2021, 162, 339–352. [Google Scholar] [CrossRef]
- Couturier, C.P.; Ayyadhury, S.; Le, P.U.; Nadaf, J.; Monlong, J.; Riva, G.; Allache, R.; Baig, S.; Yan, X.; Bourgey, M.; et al. Single-Cell RNA-Seq Reveals That Glioblastoma Recapitulates a Normal Neurodevelopmental Hierarchy. Nat. Commun. 2020, 11, 3406. [Google Scholar] [CrossRef]
- Lee, E.; Yong, R.L.; Paddison, P.; Zhu, J. Comparison of Glioblastoma (GBM) Molecular Classification Methods. Semin. Cancer Biol. 2018, 53, 201–211. [Google Scholar] [CrossRef]
- Barili, F.; Parolari, A.; Kappetein, P.A.; Freemantle, N. Statistical Primer: Heterogeneity, Random- or Fixed-Effects Model Analyses? Interact. Cardiovasc. Thorac. Surg. 2018, 27, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. A Basic Introduction to Fixed-Effect and Random-Effects Models for Meta-Analysis. Res. Synth. Methods 2010, 1, 97–111. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Senhaji, N.; Squalli Houssaini, A.; Lamrabet, S.; Louati, S.; Bennis, S. Molecular and Circulating Biomarkers in Patients with Glioblastoma. Int. J. Mol. Sci. 2022, 23, 7474. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Dataset | Platform | Sample Size | IQC | AQCg | CQCg | SMR | Reference |
|---|---|---|---|---|---|---|---|---|
| 1 | GSE59612 | Illumina HiSeq 2000 | 22 glioblastoma tumor tissue, 22 controls | 5.6 | 61.25 | 145.18 | 1.67 | [18] |
| 2 | GSE62731 | Illumina HiSeq 2000 | 2 glioblastoma tumor tissue, 2 controls | 3.3 | 2.49 | 52.78 | 3.33 | [19] |
| 3 | GSE86202 | Illumina HiSeq 2500 | 3 glioblastoma tumor tissue, 3 controls | 1.3 | 2.66 | 17.74 | 3.67 | [20] |
| 4 | GSE165595 | Illumina HiSeq 4000 | 15 glioblastoma tumor tissue, 15 controls | 5.6 | 23.08 | 240.99 | 1.33 | [21] |
| Database | Overlapping DElncRNA/DEPCGs Associated Pathways (FDR < 0.05) |
|---|---|
| KEGG | Ribosome |
| Reactome | Translation Peptide Chain Elongation Influenza Viral RNA Transcription And Replication Nonsense-Mediated Decay Enhanced By The Exon Junction Complex SRP-dependent co-translational protein targeting to membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sallam, M.; Mysara, M.; Baatout, S.; Guns, P.-J.; Ramadan, R.; Benotmane, M.A. Meta-Analysis of RNA-Seq Datasets Identifies Novel Players in Glioblastoma. Cancers 2022, 14, 5788. https://doi.org/10.3390/cancers14235788
Sallam M, Mysara M, Baatout S, Guns P-J, Ramadan R, Benotmane MA. Meta-Analysis of RNA-Seq Datasets Identifies Novel Players in Glioblastoma. Cancers. 2022; 14(23):5788. https://doi.org/10.3390/cancers14235788
Chicago/Turabian StyleSallam, Magy, Mohamed Mysara, Sarah Baatout, Pieter-Jan Guns, Raghda Ramadan, and Mohammed Abderrafi Benotmane. 2022. "Meta-Analysis of RNA-Seq Datasets Identifies Novel Players in Glioblastoma" Cancers 14, no. 23: 5788. https://doi.org/10.3390/cancers14235788
APA StyleSallam, M., Mysara, M., Baatout, S., Guns, P.-J., Ramadan, R., & Benotmane, M. A. (2022). Meta-Analysis of RNA-Seq Datasets Identifies Novel Players in Glioblastoma. Cancers, 14(23), 5788. https://doi.org/10.3390/cancers14235788