
NOTCH1 Intracellular Domain and the Tumor Microenvironment as Prognostic Markers in HNSCC

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfection
2.2. siRNA-Mediated Knockdown
2.3. Western Blot
2.4. DAPT and DLL4 Treatment
2.5. Migration Assay
2.6. Proliferation Assay
2.7. Patient Cohort
2.8. Immunohistochemistry
2.9. Immunoscore
2.10. RNA Isolation and cDNA Synthesis
2.11. Quantitative Real-Time PCR
2.12. Statistical Analysis
3. Results
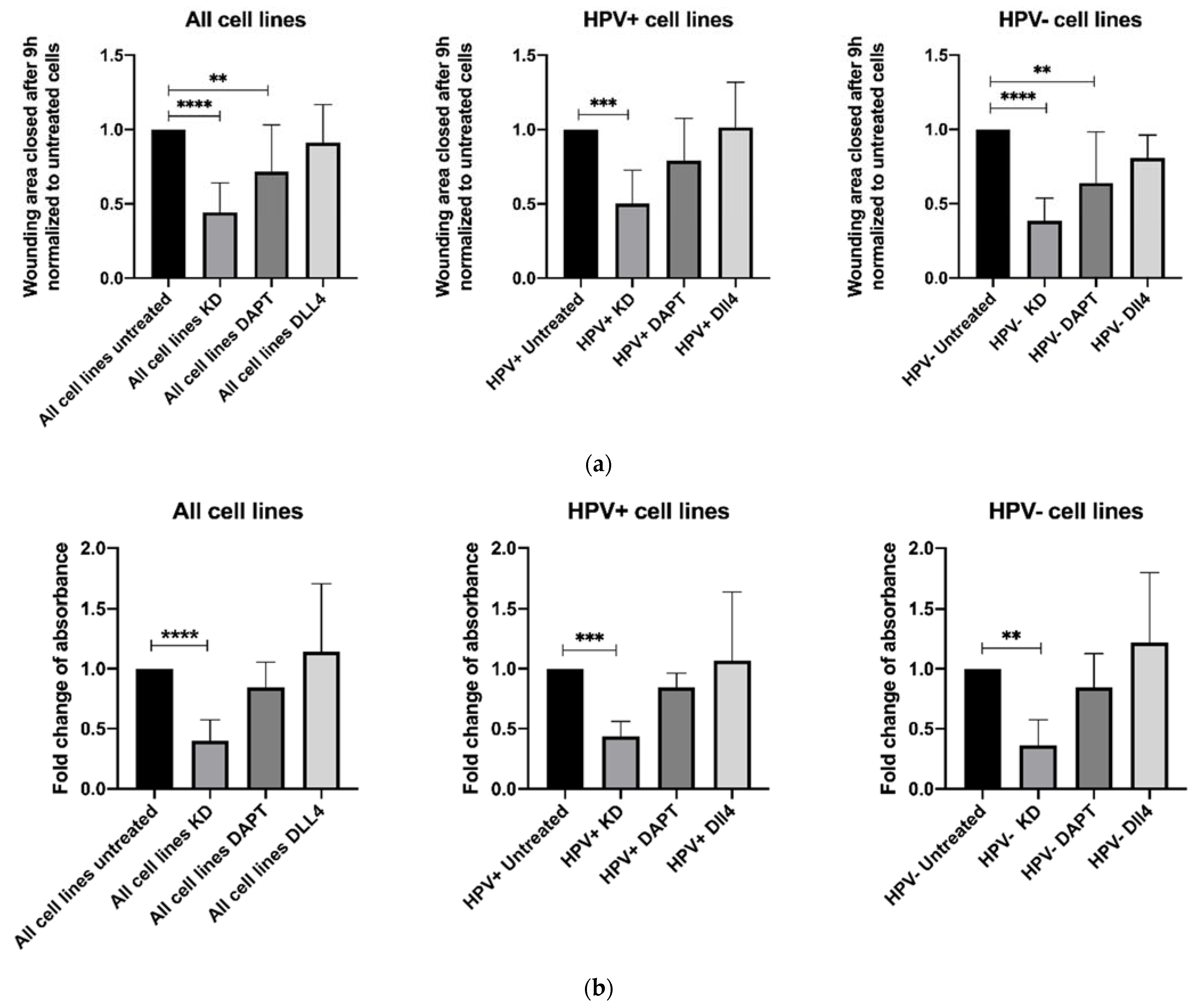
3.1. NOTCH1 Is Involved in Migration and Proliferation of HNSCC Cell Lines
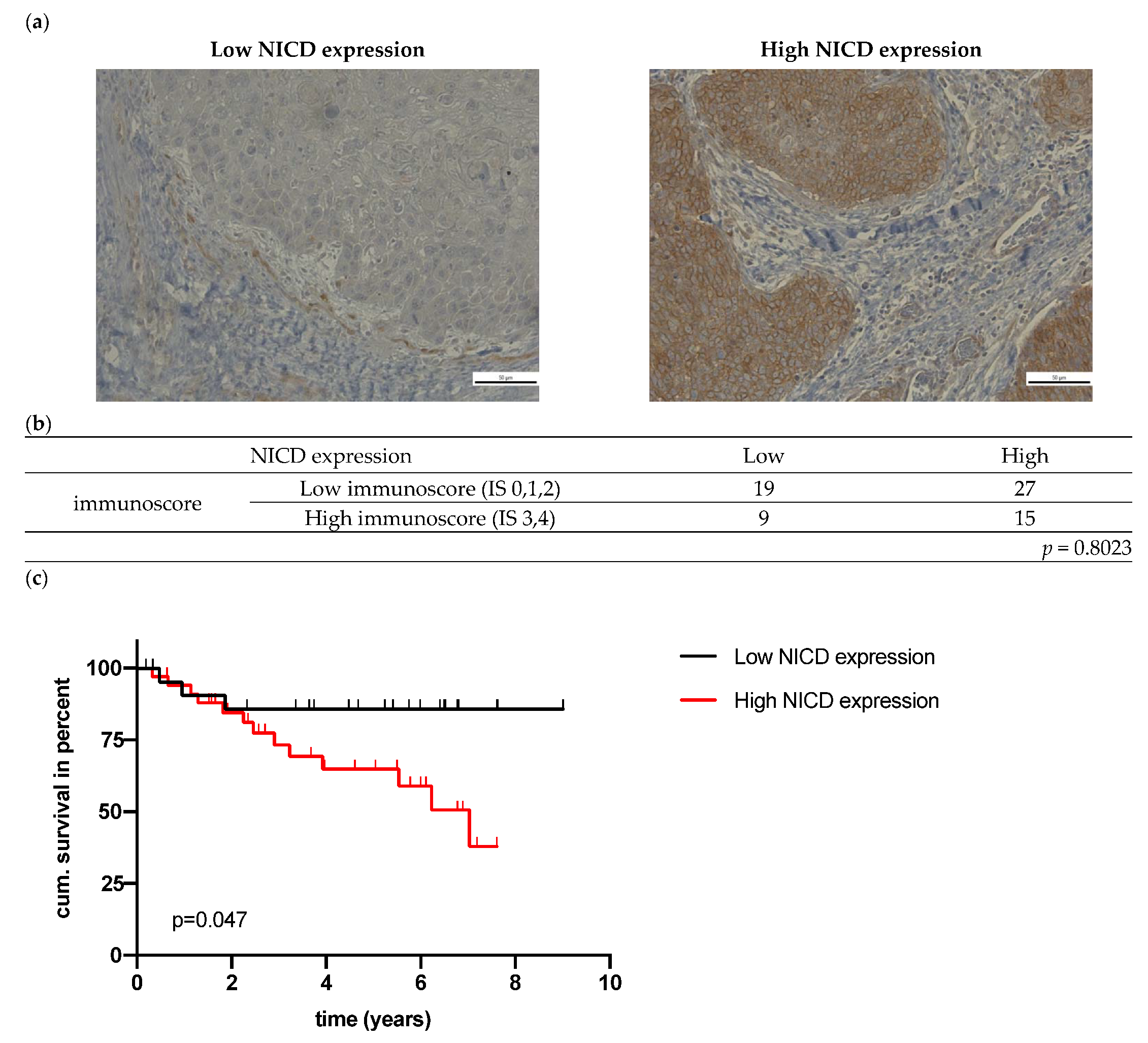
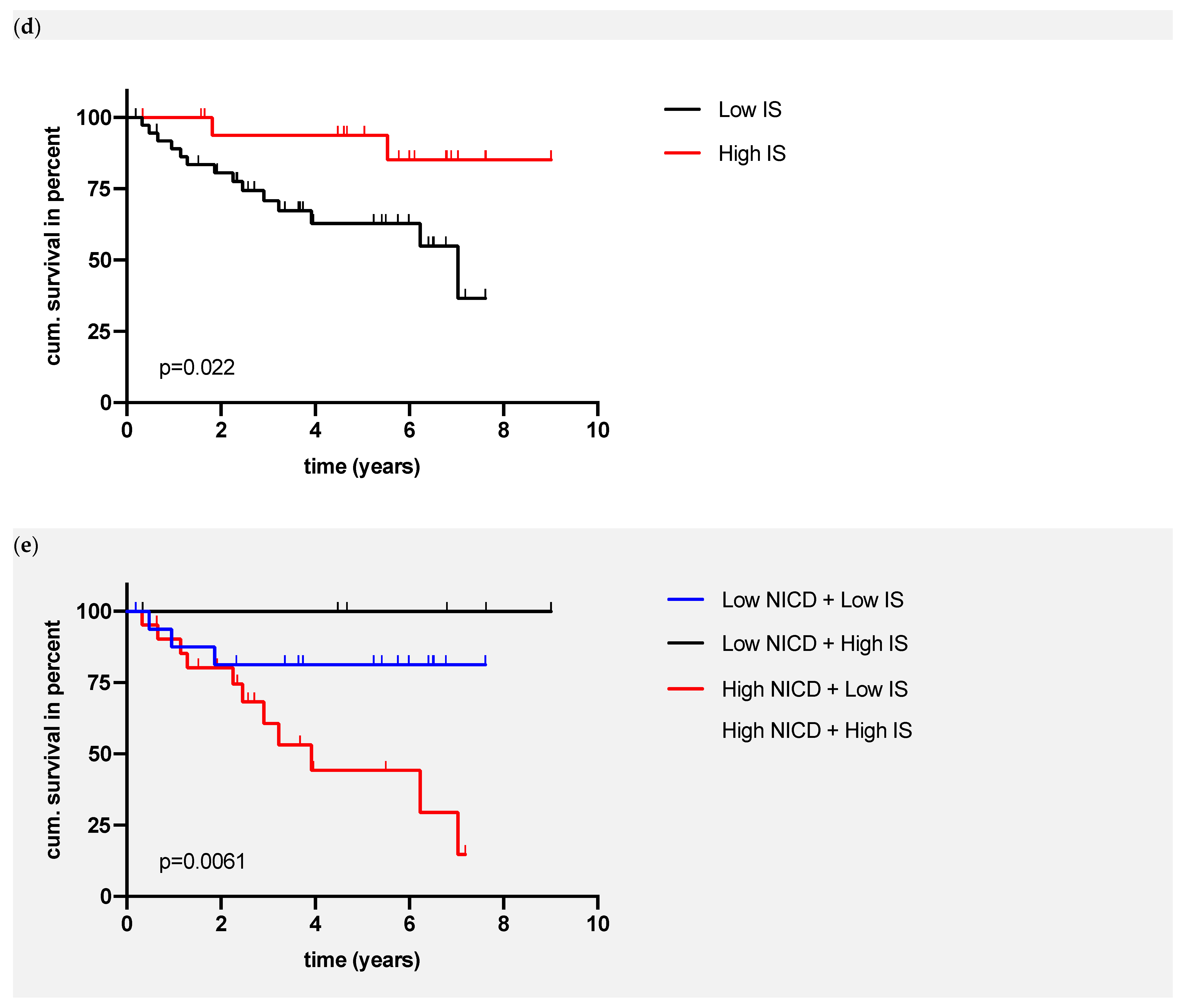
3.2. Association between NOTCH1 and NICD Expression and Clinical and Pathological Parameters
- High-risk patients: high NICD and low IS;
- Intermediate-risk patients: low NICD and high IS or high NICD and low IS;
- Low-risk patients: low NICD and high IS.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, T.L.; Stewart, P.A.; Davis, S.; Thomas, D.B. Work in dry cleaning and the incidence of cancer of the oral cavity, larynx, and oesophagus. Occup. Environ. Med. 1997, 54, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Becher, H.; Ramroth, H.; Ahrens, W.; Risch, A.; Schmezer, P.; Dietz, A. Occupation, exposure to polycyclic aromatic hydrocarbons and laryngeal cancer risk. Int. J. Cancer 2005, 116, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- Cohen, E.E.W.; Bell, R.B.; Bifulco, C.B.; Burtness, B.; Gillison, M.L.; Harrington, K.J.; Le, Q.-T.; Lee, N.Y.; Leidner, R.; Lewis, R.L.; et al. The Society for Immunotherapy of Cancer consensus statement on immunotherapy for the treatment of squamous cell carcinoma of the head and neck (HNSCC). J. Immunother. Cancer 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Puram, S.V.; Rocco, J.W. Molecular Aspects of Head and Neck Cancer Therapy. Hematol. Oncol. Clin. N. Am. 2015, 29, 971–992. [Google Scholar] [CrossRef] [Green Version]
- Bauman, J.E.; Duvvuri, U.; Thomas, S.; Gooding, W.E.; Clump, D.A.; Karlovits, B.; Wehbe, A.; Miller, F.R.; Kim, S.; Sen, M.; et al. Phase 1 study of EGFR-antisense DNA, cetuximab, and radiotherapy in head and neck cancer with preclinical correlatives. Cancer 2018, 124, 3881–3889. [Google Scholar] [CrossRef] [Green Version]
- Penton, A.L.; Leonard, L.D.; Spinner, N.B. Notch signaling in human development and disease. Semin. Cell Dev. Biol. 2012, 23, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Struhl, G.; Greenwald, I. Presenilin is required for activity and nuclear access of Notch in Drosophila. Nature 1999, 398, 522–525. [Google Scholar] [CrossRef]
- Rettig, E.M.; Chung, C.H.; Bishop, J.A.; Howard, J.D.; Sharma, R.; Li, R.J.; Douville, C.; Karchin, R.; Izumchenko, E.; Sidransky, D.; et al. Cleaved NOTCH1 Expression Pattern in Head and Neck Squamous Cell Carcinoma Is Associated with NOTCH1 Mutation, HPV Status, and High-Risk Features. Cancer Prev. Res. 2015, 8, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Fukusumi, T.; Califano, J.A. The NOTCH Pathway in Head and Neck Squamous Cell Carcinoma. J. Dent. Res. 2018, 97, 645–653. [Google Scholar] [CrossRef]
- Ellisen, L.W.; Bird, J.; West, D.C.; Soreng, A.L.; Reynolds, T.C.; Smith, S.D.; Sklar, J. TAN-1, the human homolog of the Drosophila notch gene, is broken by chromosomal translocations in T lymphoblastic neoplasms. Cell 1991, 66, 649–661. [Google Scholar] [CrossRef]
- Stransky, N.; Egloff, A.M.; Tward, A.D.; Kostic, A.D.; Cibulskis, K.; Sivachenko, A.; Kryukov, G.V.; Lawrence, M.S.; Sougnez, C.; McKenna, A.; et al. The mutational landscape of head and neck squamous cell carcinoma. Science 2011, 333, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, N.; Frederick, M.J.; Pickering, C.R.; Bettegowda, C.; Chang, K.; Li, R.J.; Fakhry, C.; Xie, T.X.; Zhang, J.; Wang, J.; et al. Exome sequencing of head and neck squamous cell carcinoma reveals inactivating mutations in NOTCH1. Science 2011, 333, 1154–1157. [Google Scholar] [CrossRef] [Green Version]
- Meurette, O.; Mehlen, P. Notch Signaling in the Tumor Microenvironment. Cancer Cell 2018, 34, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Thomas Koch, D.; Pickhard, A.; Gebel, L.; Buchberger, A.M.S.; Bas, M.; Mogler, C.; Reiter, R.; Piontek, G.; Wirth, M. Epidermal growth factor receptor variant III in head and neck squamous cell carcinoma is not relevant for targeted therapy and irradiation. Oncotarget 2017, 8, 32668. [Google Scholar] [CrossRef] [Green Version]
- Galon, J.; Mlecnik, B.; Bindea, G.; Angell, H.K.; Berger, A.; Lagorce, C.; Lugli, A.; Zlobec, I.; Hartmann, A.; Bifulco, C.; et al. Towards the introduction of the ‘Immunoscore’ in the classification of malignant tumours. J. Pathol. 2014, 232, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Lechner, A.; Schlößer, H.; Rothschild, S.I.; Thelen, M.; Reuter, S.; Zentis, P.; Shimabukuro-Vornhagen, A.; Theurich, S.; Wennhold, K.; Garcia-Marquez, M.; et al. Characterization of tumor-associated T-lymphocyte subsets and immune checkpoint molecules in head and neck squamous cell carcinoma. Oncotarget 2017, 8, 44418–44433. [Google Scholar] [CrossRef] [Green Version]
- Galon, J.; Pagès, F.; Marincola, F.M.; Angell, H.K.; Thurin, M.; Lugli, A.; Zlobec, I.; Berger, A.; Bifulco, C.; Botti, G.; et al. Cancer classification using the Immunoscore: A worldwide task force. J. Transl. Med. 2012, 10, 205. [Google Scholar] [CrossRef]
- Jing, P.; Zhou, S.; Xu, P.; Cui, P.; Liu, X.; Liu, X.; Liu, X.; Wang, H.; Xu, W. PDK1 promotes metastasis by inducing epithelial-mesenchymal transition in hypopharyngeal carcinoma via the Notch1 signaling pathway. Exp. Cell Res. 2020, 386, 111746. [Google Scholar] [CrossRef]
- Shrivastava, S.; Steele, R.; Sowadski, M.; Crawford, S.E.; Varvares, M.; Ray, R.B. Identification of molecular signature of head and neck cancer stem-like cells. Sci. Rep. 2015, 5, 7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangarajan, A.; Talora, C.; Okuyama, R.; Nicolas, M.; Mammucari, C.; Oh, H.; Aster, J.C.; Krishna, S.; Metzger, D.; Chambon, P.; et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001, 20, 3427–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.A.; Huang, C.; Li, Q.; Kazi, S.A.; Byers, L.A.; Wang, J.; Johnson, F.M.; Frederick, M.J. NOTCH1 Signaling in Head and Neck Squamous Cell Carcinoma. Cells 2020, 9, 2677. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Liu, C.; Lin, Y.; Fu, J.; Lu, G.; Lu, Z. pH sensitive peptide functionalized nanoparticles for co-delivery of erlotinib and DAPT to restrict the progress of triple negative breast cancer. Drug Deliv. 2019, 26, 470–480. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.-J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.Y.; Luan, X.; Huang, X.; Zhao, S.; Zhao, H. Relationship between the expression of Notch1 and EZH2 and the prognosis of breast invasive ductal carcinoma. Genet Mol. Res. 2016, 15, gmr.15017464. [Google Scholar] [CrossRef]
- Zhao, Z.L.; Zhang, L.; Huang, C.F.; Ma, S.R.; Bu, L.L.; Liu, J.F.; Yu, G.T.; Liu, B.; Gutkind, J.S.; Kulkarni, A.B.; et al. NOTCH1 inhibition enhances the efficacy of conventional chemotherapeutic agents by targeting head neck cancer stem cell. Sci. Rep. 2016, 6, 24704. [Google Scholar] [CrossRef] [Green Version]
- Zohny, S.F.; Zamzami, M.A.; Al-Malki, A.L.; Trabulsi, N.H. Highly Expressed DLL4 and JAG1: Their Role in Incidence of Breast Cancer Metastasis. Arch. Med. Res. 2020, 51, 145–152. [Google Scholar] [CrossRef]
- Huang, J.; Hu, W.; Hu, L.; Previs, R.A.; Dalton, H.J.; Yang, X.Y.; Sun, Y.; McGuire, M.; Rupaimoole, R.; Nagaraja, A.S.; et al. Dll4 Inhibition plus Aflibercept Markedly Reduces Ovarian Tumor Growth. Mol. Cancer Ther. 2016, 15, 1344–1352. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, F.; Osborne, B.A. Non-canonical notch signaling in cancer and immunity. Front. Oncol. 2014, 4, 345. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, R.; Ito, T.; Hassan, W.A.; Nakayama, H. Notch1 in oral squamous cell carcinoma. Histol. Histopathol. 2017, 32, 315–323. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, Z.; Zhang, M.; Gross, N.; Gong, L.; Zhang, S.; Lei, D.; Zeng, Q.; Luo, X.; Li, G.; et al. High Notch1 expression affects chemosensitivity of head and neck squamous cell carcinoma to paclitaxel and cisplatin treatment. Biomed. Pharmacother. 2019, 118, 109306. [Google Scholar] [CrossRef]
- Covelo-Molares, H.; Bartosovic, M.; Vanacova, S. RNA methylation in nuclear pre-mRNA processing. Wiley Interdiscip. Rev. RNA 2018, 9, e1489. [Google Scholar] [CrossRef] [Green Version]
- Kaka, A.S.; Nowacki, N.B.; Kumar, B.; Zhao, S.; Old, M.O.; Agrawal, A.; Ozer, E.; Carrau, R.L.; Schuller, D.E.; Kumar, P.; et al. Notch1 Overexpression Correlates to Improved Survival in Cancer of the Oropharynx. Otolaryngol. Head Neck Surg. 2017, 156, 652–659. [Google Scholar] [CrossRef]
- Tian, J.; Liu, X.; Liu, X.; Jing, P.; Sa, N.; Wang, H.; Xu, W. Notch1 serves as a prognostic factor and regulates metastasis via regulating EGFR expression in hypopharyngeal squamous cell carcinoma. Onco Targets Ther. 2018, 11, 7395–7405. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Tang, P.; Li, S.; Qin, X.; Yang, H.; Wu, C.; Liu, Y. Notch signaling pathway networks in cancer metastasis: A new target for cancer therapy. Med. Oncol. 2017, 34, 180. [Google Scholar] [CrossRef]
- Ward, M.J.; Thirdborough, S.M.; Mellows, T.; Riley, C.; Harris, S.; Suchak, K.; Webb, A.; Hampton, C.; Patel, N.N.; Randall, C.J.; et al. Tumour-infiltrating lymphocytes predict for outcome in HPV-positive oropharyngeal cancer. Br. J. Cancer 2014, 110, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, H.; Zeng, Q.; Guo, X.-J.; Wang, H.; Liu, H.-H.; Dong, Z.-Y. HPV-positive status associated with inflamed immune microenvironment and improved response to anti-PD-1 therapy in head and neck squamous cell carcinoma. Sci. Rep. 2019, 9, 13404. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Ulrich, P.; Wilson, E.; Parikh, F.; Narang, P.; Yang, S.; Read, A.K.; Kim-Schulze, S.; Park, J.G.; Posner, M.; et al. Human Papilloma Virus Specific Immunogenicity and Dysfunction of CD8(+) T Cells in Head and Neck Cancer. Cancer Res. 2018, 78, 6159–6170. [Google Scholar] [CrossRef] [Green Version]
- Angell, H.K.; Bruni, D.; Barrett, J.C.; Herbst, R.; Galon, J. The Immunoscore: Colon Cancer and Beyond. Clin. Cancer Res. 2020, 26, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Kümpers, C.; Jokic, M.; Haase, O.; Offermann, A.; Vogel, W.; Grätz, V.; Langan, E.A.; Perner, S.; Terheyden, P. Immune Cell Infiltration of the Primary Tumor, Not PD-L1 Status, Is Associated With Improved Response to Checkpoint Inhibition in Metastatic Melanoma. Front. Med. 2019, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Annealing Temperature |
|---|---|---|
| NOTCH1 forward NOTCH1 reverse | TGAATGGCGGGAAGTGTGAAG GGTTGGGGTCCTGGCATCG | 62 °C |
| GAPDH forward GAPDH reverse | AGCCACATCGCTCAGACA GCCCAATACGACCAAATCC | 56 °C |
| Variate | Overall | NICD IHC Expression | p Value (Fisher’s Exact) | Overall | NOTCH1 mRNA Expression | p Value (Fisher’s Exact) | ||
|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | |||||
| Tumor stage (pathological) | ||||||||
| T1/T2 | 48 (68.6%) | 23 | 25 | 43 (71.7%) | 26 | 17 | ||
| T3/T4 | 22 (31.4%) | 5 | 17 | 0.0659 | 17 (28.3%) | 4 | 13 | 0.0204 |
| Nodal stage (pathological) | ||||||||
| N0 | 20 (28.6%) | 12 | 8 | 17 (28.3%) | 10 | 7 | ||
| N1-3 | 50 (71.4%) | 16 | 34 | 0.0571 | 43 (71.7%) | 20 | 23 | 0.5676 |
| Metastasis (initial stage) | ||||||||
| M0 | 67 (95.7%) | 28 | 39 | 57 (95%) | 29 | 28 | ||
| M1 | 3 (4.3%) | 0 | 3 | 0.2696 | 3 (5%) | 1 | 2 | >0.9999 |
| p16 Status | ||||||||
| p16 positive | 26 (37.1%) | 6 | 20 | 23 (38.3%) | 8 | 15 | ||
| p16 negative | 44 (62.9%) | 22 | 22 | 0.0425 | 37 (61.7%) | 22 | 15 | 0.1102 |
| Gender | ||||||||
| Male | 56 (80%) | 24 | 31 | 47 (78.3%) | 24 | 23 | ||
| Female | 14 (20%) | 4 | 11 | 0.3730 | 13 (21.7%) | 6 | 7 | >0.9999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidl, B.; Siegl, M.; Boxberg, M.; Stögbauer, F.; Jira, D.; Winter, C.; Stark, L.; Pickhard, A.; Wollenberg, B.; Wirth, M. NOTCH1 Intracellular Domain and the Tumor Microenvironment as Prognostic Markers in HNSCC. Cancers 2022, 14, 1080. https://doi.org/10.3390/cancers14041080
Schmidl B, Siegl M, Boxberg M, Stögbauer F, Jira D, Winter C, Stark L, Pickhard A, Wollenberg B, Wirth M. NOTCH1 Intracellular Domain and the Tumor Microenvironment as Prognostic Markers in HNSCC. Cancers. 2022; 14(4):1080. https://doi.org/10.3390/cancers14041080
Chicago/Turabian StyleSchmidl, Benedikt, Michael Siegl, Melanie Boxberg, Fabian Stögbauer, Daniel Jira, Christof Winter, Leonhard Stark, Anja Pickhard, Barbara Wollenberg, and Markus Wirth. 2022. "NOTCH1 Intracellular Domain and the Tumor Microenvironment as Prognostic Markers in HNSCC" Cancers 14, no. 4: 1080. https://doi.org/10.3390/cancers14041080
APA StyleSchmidl, B., Siegl, M., Boxberg, M., Stögbauer, F., Jira, D., Winter, C., Stark, L., Pickhard, A., Wollenberg, B., & Wirth, M. (2022). NOTCH1 Intracellular Domain and the Tumor Microenvironment as Prognostic Markers in HNSCC. Cancers, 14(4), 1080. https://doi.org/10.3390/cancers14041080