
Circulating Nucleic Acids as Novel Biomarkers for Pancreatic Ductal Adenocarcinoma


Abstract
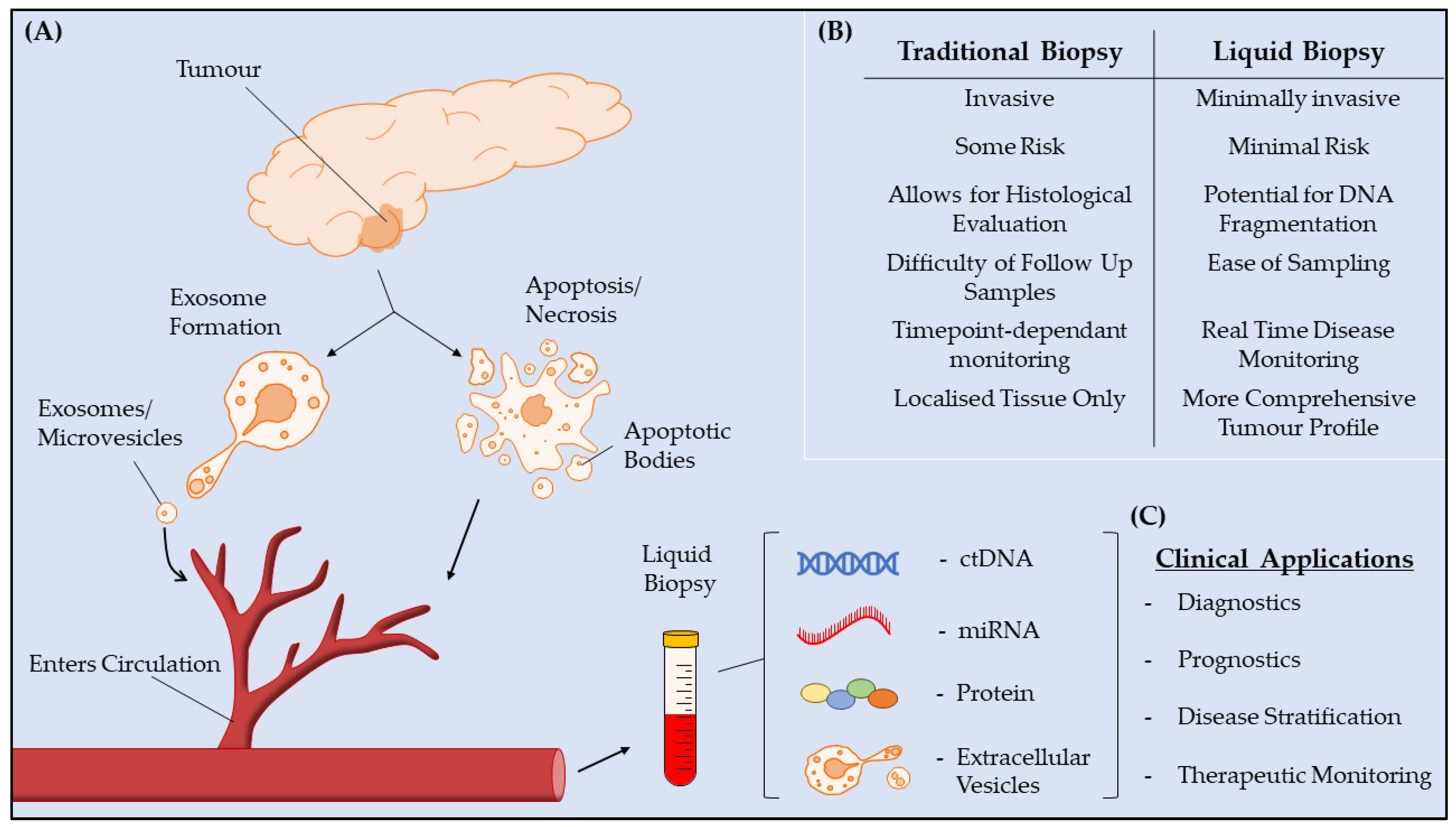
:Simple Summary
Abstract
1. Introduction
2. Protein Biomarkers in PDAC
3. KRAS Mutations in Cell Free DNA in Combination with Protein Biomarkers as a Diagnostic Panel
4. miRNA as a Diagnostic Biomarker in Early-Stage Disease
5. Potential of Exosomal miRNA as a PDAC Biomarker
6. Exosomal Derived KRAS Mutations as a Diagnostic Biomarker
7. Circulating Nucleic Acids as Prognostic Biomarkers
8. miRNA in Detecting Lymph Node Metastasis
9. Combined Diagnostic Panel
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Abbreviation | Description |
| AkT | Protein Kinase B |
| AUC | Area Under the Curve |
| CA 19-9 | Carbohydrate Antigen 19-9 |
| CEA | Carcinoembryonic Antigen |
| CI | Confidence Interval |
| CK18 | Cytokeratin-18 |
| CT | Computed Tomography |
| ctDNA | Circulating Tumor DNA |
| ctRNA | Circulating Tumor RNA |
| ddPCR | Droplet Digital PCR |
| DFS | Disease Free Survival |
| EMT | Epithelial–Mesenchymal Transition |
| ERK | Extracellular-Signal-Regulated Kinase |
| ERP | Endoscopic Retrograde Pancreatography |
| EUS-FNA | Endoscopic Ultrasound Fine Needle Aspiration |
| Ex-miRNA | Exosomal Micro RNA |
| GAPDH | Glyceraldehyde 3-Phosphate Dehydrogenase |
| GTP | Guanosine-5’-triphosphate |
| HGF | Hepatocyte Growth Factor |
| IPMNs | Intraductal Papillary Mucinous Neoplasm |
| JAK | Janus Kinase |
| KRAS | Kirsten Rat Sarcoma Viral Oncogene Homolog |
| LASSO | Least Absolute Shrinkage and Selection Operator |
| MAF | Mutant Allele Frequency |
| MAPK | Mitogen-Activated Protein Kinase |
| MET | Mesenchymal-Epithelial Transition |
| miRNA | Micro RNA |
| mRNA | Messenger RNA |
| NGS | Next-Generation Sequencing |
| OPN | Osteopontin |
| OS | Overall Survival |
| PanIN | Pancreatic Intraepithelial Neoplasia |
| PDAC | Pancreatic Ductal Adenocarcinoma |
| PFS | Progression Free Survival |
| PGM | Personal Genome Machine |
| PI3K | Phosphatidylinositol-3-Kinase |
| PJC | Pancreatic Juice Cytology |
| PNA | Peptide Nucleic Acid |
| PPV | Positive Predictive Value |
| qPCR | Real-Time Polymerase Chain Reaction |
| RAF | Rapidly Accelerated Fibrosarcoma |
| ROC | Receiver Operating Characteristic |
| STAT | Signal Transducer and Activator |
| TME | Tumor Microenvironment |
| UTR | Untranslated Region |
| ZEB-1 | Zinc Finger E-Box Binding Homeobox 1 |
References
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Bekkali, N.L.H.; Oppong, K.W. Pancreatic ductal adenocarcinoma epidemiology and risk assessment: Could we prevent? Possibility for an early diagnosis. Endosc. Ultrasound 2017, 6, S58–S61. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Laguna, I.; Hidalgo, M. Pancreatic cancer: From state-of-the-art treatments to promising novel therapies. Nat. Rev. Clin. Oncol. 2015, 12, 319–334. [Google Scholar] [CrossRef]
- Matsuno, S.; Egawa, S.; Fukuyama, S.; Motoi, F.; Sunamura, M.; Isaji, S.; Imaizumi, T.; Okada, S.; Kato, H.; Suda, K.; et al. Pancreatic Cancer Registry in Japan: 20 years of experience. Pancreas 2004, 28, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Ahmad, A.; Banerjee, S.; Azmi, A.S.; Kong, D.; Sarkar, F.H. Pancreatic cancer: Understanding and overcoming chemoresistance. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 27–33. [Google Scholar] [CrossRef]
- Javadrashid, D.; Baghbanzadeh, A.; Derakhshani, A.; Leone, P.; Silvestris, N.; Racanelli, V.; Solimando, A.G.; Baradaran, B. Pancreatic Cancer Signaling Pathways, Genetic Alterations, and Tumor Microenvironment: The Barriers Affecting the Method of Treatment. Biomedicines 2021, 9, 373. [Google Scholar] [CrossRef]
- Karamitopoulou, E. Tumour microenvironment of pancreatic cancer: Immune landscape is dictated by molecular and histopathological features. Br. J. Cancer 2019, 121, 5–14. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. The biological functions of miRNAs: Lessons from in vivo studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Leva, G.; Croce, C.M. miRNA profiling of cancer. Curr. Opin. Genet. Dev. 2013, 23, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, A.A.; Azmi, A.S.; Philip, P.A. miRNA and Gene Expression in Pancreatic Ductal Adenocarcinoma. Am. J. Pathol. 2019, 189, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids—The mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-Throughput Droplet Digital PCR System for Absolute Quantitation of DNA Copy Number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.C.; Laperriere, G.; Germain, H. Droplet Digital PCR versus qPCR for gene expression analysis with low abundant targets: From variable nonsense to publication quality data. Sci. Rep. 2017, 7, 2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, D. Next-generation sequencing and its clinical application. Cancer Biol. Med. 2019, 16, 4–10. [Google Scholar] [CrossRef]
- Gonzaga-Jauregui, C.; Zepeda Mendoza, C.J. Chapter 4-Genomic sequencing of rare diseases. In Genomics of Rare Diseases; Gonzaga-Jauregui, C., Lupski, J.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 61–95. [Google Scholar]
- Meera Krishna, B.; Khan, M.A.; Khan, S.T. Next-Generation Sequencing (NGS) Platforms: An Exciting Era of Genome Sequence Analysis. In Microbial Genomics in Sustainable Agroecosystems: Volume 2; Tripathi, V., Kumar, P., Tripathi, P., Kishore, A., Kamle, M., Eds.; Springer: Singapore, 2019; pp. 89–109. [Google Scholar]
- Reuter, J.A.; Spacek, D.V.; Snyder, M.P. High-throughput sequencing technologies. Mol. Cell 2015, 58, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Sadakari, Y.; Ohtsuka, T.; Okayama, T.; Nakashima, Y.; Gotoh, Y.; Saeki, K.; Mori, Y.; Nakata, K.; Miyasaka, Y.; et al. Pancreatic Juice Exosomal MicroRNAs as Biomarkers for Detection of Pancreatic Ductal Adenocarcinoma. Ann. Surg. Oncol. 2019, 26, 2104–2111. [Google Scholar] [CrossRef]
- Xu, Y.-F.; Hannafon, B.N.; Zhao, Y.D.; Postier, R.G.; Ding, W.-Q. Plasma exosome miR-196a and miR-1246 are potential indicators of localized pancreatic cancer. Oncotarget 2017, 8, 77028–77040. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Kim, D.U.; San Lucas, F.A.; Castillo, J.; Allenson, K.; Mulu, F.C.; Stephens, B.M.; Huang, J.; Semaan, A.; Guerrero, P.A.; et al. Circulating Nucleic Acids Are Associated With Outcomes of Patients With Pancreatic Cancer. Gastroenterology 2019, 156, 108–118.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allenson, K.; Castillo, J.; San Lucas, F.A.; Scelo, G.; Kim, D.U.; Bernard, V.; Davis, G.; Kumar, T.; Katz, M.; Overman, M.J.; et al. High prevalence of mutantKRAS in circulating exosome-derived DNA from early-stage pancreatic cancer patients. Ann. Oncol. 2017, 28, 741–747. [Google Scholar] [CrossRef]
- Cohen, J.D.; Javed, A.A.; Thoburn, C.; Wong, F.; Tie, J.; Gibbs, P.; Schmidt, C.M.; Yip-Schneider, M.T.; Allen, P.J.; Schattner, M.; et al. Combined circulating tumor DNA and protein biomarker-based liquid biopsy for the earlier detection of pancreatic cancers. Proc. Natl. Acad. Sci. USA 2017, 114, 10202–10207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perets, R.; Greenberg, O.; Shentzer, T.; Semenisty, V.; Epelbaum, R.; Bick, T.; Sarji, S.; Ben-Izhak, O.; Sabo, E.; Hershkovitz, D. Mutant KRAS Circulating Tumor DNA Is an Accurate Tool for Pancreatic Cancer Monitoring. Oncologist 2018, 23, 566–572. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Kitago, M.; Matsuda, S.; Nakamura, Y.; Fujita, Y.; Imai, S.; Shinoda, M.; Yagi, H.; Abe, Y.; Hibi, T.; et al. KRAS mutations in cell-free DNA from preoperative and postoperative sera as a pancreatic cancer marker: A retrospective study. Br. J. Cancer 2018, 118, 662–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Re, M.; Vivaldi, C.; Rofi, E.; Vasile, E.; Miccoli, M.; Caparello, C.; d’Arienzo, P.D.; Fornaro, L.; Falcone, A.; Danesi, R. Early changes in plasma DNA levels of mutant KRAS as a sensitive marker of response to chemotherapy in pancreatic cancer. Sci. Rep. 2017, 7, 7931. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Ayub, M.; Rothwell, D.G.; Gulati, S.; Kilerci, B.; Hollebecque, A.; Sun Leong, H.; Smith, N.K.; Sahoo, S.; Descamps, T.; et al. Analysis of circulating cell-free DNA identifies KRAS copy number gain and mutation as a novel prognostic marker in Pancreatic cancer. Sci. Rep. 2019, 9, 11610. [Google Scholar] [CrossRef] [PubMed]
- Lemberger, M.; Loewenstein, S.; Lubezky, N.; Nizri, E.; Pasmanik-Chor, M.; Barazovsky, E.; Klausner, J.M.; Lahat, G. MicroRNA profiling of pancreatic ductal adenocarcinoma (PDAC) reveals signature expression related to lymph node metastasis. Oncotarget 2019, 10, 2644–2656. [Google Scholar] [CrossRef]
- Vila-Navarro, E.; Vila-Casadesús, M.; Moreira, L.; Duran-Sanchon, S.; Sinha, R.; Ginés, À.; Fernández-Esparrach, G.; Miquel, R.; Cuatrecasas, M.; Castells, A.; et al. MicroRNAs for Detection of Pancreatic Neoplasia. Ann. Surg. 2016, 265, 1226–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aita, A.; Millino, C.; Sperti, C.; Pacchioni, B.; Plebani, M.; Pittà, C.D.; Basso, D. Serum miRNA Profiling for Early PDAC Diagnosis and Prognosis: A Retrospective Study. Biomedicines 2021, 9, 845. [Google Scholar] [CrossRef]
- Dittmar, R.L.; Liu, S.; Tai, M.C.; Rajapakshe, K.; Huang, Y.; Longton, G.; DeCapite, C.; Hurd, M.W.; Paris, P.L.; Kirkwood, K.S.; et al. Plasma miRNA Biomarkers in Limited Volume Samples for Detection of Early-stage Pancreatic Cancer. Cancer Prev. Res. 2021, 14, 729. [Google Scholar] [CrossRef]
- Selleck, M.J.; Senthil, M.; Wall, N.R. Making Meaningful Clinical Use of Biomarkers. Biomark. Insights 2017, 12, 1177271917715236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, W.; Strobel, O.; Hinz, U.; Fritz, S.; Hackert, T.; Roth, C.; Büchler, M.W.; Werner, J. CA19-9 in Potentially Resectable Pancreatic Cancer: Perspective to Adjust Surgical and Perioperative Therapy. Ann. Surg. Oncol. 2013, 20, 2188–2196. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, B.K.; Seo, J.H.; Choi, J.; Choi, J.W.; Lee, C.K.; Chung, J.B.; Park, Y.; Kim, D.W. Carbohydrate antigen 19-9 elevation without evidence of malignant or pancreatobiliary diseases. Sci. Rep. 2020, 10, 8820. [Google Scholar] [CrossRef] [PubMed]
- Ferrone, C.R.; Finkelstein, D.M.; Thayer, S.P.; Muzikansky, A.; Fernandez-delCastillo, C.; Warshaw, A.L. Perioperative CA19-9 levels can predict stage and survival in patients with resectable pancreatic adenocarcinoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 2897–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Comprehensive Cancer Network. Clinical Practice Guidelines in Oncology-Pancreatic Adenocarcinoma; National Comprehensive Cancer Network: Plymouth Meeting, PA, USA, 2022. [Google Scholar]
- Ballehaninna, U.K.; Chamberlain, R.S. The clinical utility of serum CA 19-9 in the diagnosis, prognosis and management of pancreatic adenocarcinoma: An evidence based appraisal. J. Gastrointest. Oncol. 2012, 3, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Salomaa, V.; Pankow, J.; Heiss, G.; Cakir, B.; Eckfeldt, J.H.; Ellison, R.C.; Myers, R.H.; Hiller, K.M.; Brantley, K.R.; Morris, T.L.; et al. Genetic background of Lewis negative blood group phenotype and its association with atherosclerotic disease in the NHLBI Family Heart Study. J. Intern. Med. 2000, 247, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, K.T.; Lee, J.K.; Paik, S.W.; Rhee, J.C.; Choi, K.W. Clinical usefulness of carbohydrate antigen 19-9 as a screening test for pancreatic cancer in an asymptomatic population. J. Gastroenterol. Hepatol. 2004, 19, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Bedi, M.M.; Gandhi, M.D.; Jacob, G.; Lekha, V.; Venugopal, A.; Ramesh, H. CA 19-9 to differentiate benign and malignant masses in chronic pancreatitis: Is there any benefit? Indian J. Gastroenterol. 2009, 28, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Kuntz, A.I.; Wadleigh, R.G. CA 19-9 tumor marker: Is it reliable? A case report in a patient with pancreatic cancer. Clin. Adv. Hematol. Oncol. 2013, 11, 50–52. [Google Scholar] [PubMed]
- Swords, D.S.; Firpo, M.A.; Scaife, C.L.; Mulvihill, S.J. Biomarkers in pancreatic adenocarcinoma: Current perspectives. OncoTargets Ther. 2016, 9, 7459–7467. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.; Clarke, L.; Pal, A.; Buchwald, P.; Eglinton, T.; Wakeman, C.; Frizelle, F. A Review of the Role of Carcinoembryonic Antigen in Clinical Practice. Ann. Coloproctol. 2019, 35, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Goonetilleke, K.S.; Siriwardena, A.K. Systematic review of carbohydrate antigen (CA 19-9) as a biochemical marker in the diagnosis of pancreatic cancer. Eur. J. Surg. Oncol. (EJSO) 2007, 33, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Shimoda, M.; Shimazaki, J.; Maruyama, T.; Oshiro, Y.; Nishida, K.; Sahara, Y.; Nagakawa, Y.; Tsuchida, A. Predictive Early Recurrence Factors of Preoperative Clinicophysiological Findings in Pancreatic Cancer. Eur. Surg. Res. 2018, 59, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Scholler, N.; Urban, N. CA125 in ovarian cancer. Biomark. Med. 2007, 1, 513–523. [Google Scholar] [CrossRef]
- Jiang, K.; Tan, E.; Sayegh, Z.; Centeno, B.; Malafa, M.; Coppola, D. Cancer Antigen 125 (CA125, MUC16) Protein Expression in the Diagnosis and Progression of Pancreatic Ductal Adenocarcinoma. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 620–623. [Google Scholar] [CrossRef]
- Luo, G.; Xiao, Z.; Long, J.; Liu, Z.; Liu, L.; Liu, C.; Xu, J.; Ni, Q.; Yu, X. CA125 is Superior to CA19-9 in Predicting the Resectability of Pancreatic Cancer. J. Gastrointest. Surg. 2013, 17, 2092–2098. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, M.-G.; Xu, H.-X.; Wang, W.-Q.; Liu, L.; Yu, X.-J. Preoperative Serum CA125 Levels Predict the Prognosis in Hyperbilirubinemia Patients With Resectable Pancreatic Ductal Adenocarcinoma. Medicine 2015, 94, e751. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.M.; Der, C.J. KRAS: The Critical Driver and Therapeutic Target for Pancreatic Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a031435. [Google Scholar] [CrossRef] [PubMed]
- Buscail, L.; Bournet, B.; Cordelier, P. Role of oncogenic KRAS in the diagnosis, prognosis and treatment of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Downward, J. Targeting RAS signalling pathways in cancer therapy. Nat. Rev. Cancer 2003, 3, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Su, L.; Qian, C. Circulating tumor DNA: A promising biomarker in the liquid biopsy of cancer. Oncotarget 2016, 7, 48832–48841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.L.; Wang, Y.; Gang, Q.; Zhang, Y.; Sun, Y. Plasma level of miR-93 is associated with higher risk to develop type 2 diabetic retinopathy. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Vila-Navarro, E.; Duran-Sanchon, S.; Vila-Casadesús, M.; Moreira, L.; Ginès, À.; Cuatrecasas, M.; Lozano, J.J.; Bujanda, L.; Castells, A.; Gironella, M. Novel Circulating miRNA Signatures for Early Detection of Pancreatic Neoplasia. Clin. Transl. Gastroenterol. 2019, 10, e00029. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; Zhang, M.; Wang, B.; Ye, J.; Zhang, Y.; Tang, D.; Ma, D.; Jin, W.; Li, X.; et al. Circ-ASH2L promotes tumor progression by sponging miR-34a to regulate Notch1 in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 466. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Wang, J.; Zhang, L.; Li, J.; Jin, Y. LncRNA HOTAIR Promotes Cancer Stem-Like Cells Properties by Sponging miR-34a to Activate the JAK2/STAT3 Pathway in Pancreatic Ductal Adenocarcinoma. OncoTargets Ther. 2021, 14, 1883–1893. [Google Scholar] [CrossRef]
- Yao, L.-C.; Jiang, X.-H.; Yan, S.-S.; Wang, W.; Wu, L.; Zhai, L.-L.; Xiang, F.; Ji, T.; Ye, L.; Tang, Z.-G. Four potential microRNAs affect the progression of pancreatic ductal adenocarcinoma by targeting MET via the PI3K/AKT signaling pathway. Oncol. Lett. 2021, 21, 326. [Google Scholar] [CrossRef]
- Brabletz, T. EMT and MET in Metastasis: Where Are the Cancer Stem Cells? Cancer Cell 2012, 22, 699–701. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tao, Y.; Wang, X.; Jiang, P.; Li, J.; Peng, M.; Zhang, X.; Chen, K.; Liu, H.; Zhen, P.; et al. Tumor-Secreted Exosomal miR-222 Promotes Tumor Progression via Regulating P27 Expression and Re-Localization in Pancreatic Cancer. Cell. Physiol. Biochem. 2018, 51, 610–629. [Google Scholar] [CrossRef]
- Sharma, S.S.; Pledger, W.J. The non-canonical functions of p27(Kip1) in normal and tumor biology. Cell Cycle 2016, 15, 1189–1201. [Google Scholar] [CrossRef] [Green Version]
- Tai, Y.-L.; Chen, K.-C.; Hsieh, J.-T.; Shen, T.-L. Exosomes in cancer development and clinical applications. Cancer Sci. 2018, 109, 2364–2374. [Google Scholar] [CrossRef] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAndrews, K.M.; Kalluri, R. Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 2019, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Bor, R.; Madácsy, L.; Fábián, A.; Szepes, A.; Szepes, Z. Endoscopic retrograde pancreatography: When should we do it? World J. Gastrointest. Endosc. 2015, 7, 1023–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, M.; Zhong, K.; Jiang, T.; Liu, Z.; Kwan, H.Y.; Su, T. The current understanding on the impact of KRAS on colorectal cancer. Biomed. Pharmacother. 2021, 140, 111717. [Google Scholar] [CrossRef] [PubMed]
- Westcott, P.M.K.; To, M.D. The genetics and biology of KRAS in lung cancer. Chin. J. Cancer 2013, 32, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principe, D.R.; Underwood, P.W.; Korc, M.; Trevino, J.G.; Munshi, H.G.; Rana, A. The Current Treatment Paradigm for Pancreatic Ductal Adenocarcinoma and Barriers to Therapeutic Efficacy. Front. Oncol. 2021, 11, 2773. [Google Scholar] [CrossRef] [PubMed]
- Gall, T.M.; Jacob, J.; Frampton, A.E.; Krell, J.; Kyriakides, C.; Castellano, L.; Stebbing, J.; Jiao, L.R. Reduced dissemination of circulating tumor cells with no-touch isolation surgical technique in patients with pancreatic cancer. JAMA Surg. 2014, 149, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimasu, T.; Maebeya, S.; Suzuma, T.; Bessho, T.; Tanino, H.; Arimoto, J.; Sakurai, T.; Naito, Y. Disappearance Curves for Tumor Markers after Resection of Intrathoracic Malignancies. Int. J. Biol. Markers 1999, 14, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, L.A., Jr.; Bardelli, A. Liquid biopsies: Genotyping circulating tumor DNA. J. Clin. Oncol. 2014, 32, 579–586. [Google Scholar] [CrossRef]
- Pai, R.K.; Beck, A.H.; Mitchem, J.; Linehan, D.C.; Chang, D.T.; Norton, J.A. Pattern of lymph node involvement and prognosis in pancreatic adenocarcinoma: Direct lymph node invasion has similar survival to node-negative disease. Am. J. Surg. Pathol. 2011, 35, 228–234. [Google Scholar] [CrossRef]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Kawamoto, K.; et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 42339. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Du, Y.; Wang, G.; Gao, J.; Gong, Y.; Li, L.; Zhang, Z.; Zhu, J.; Jing, Q.; Qin, Y.; et al. Detection of differentially expressed microRNAs in serum of pancreatic ductal adenocarcinoma patients: miR-196a could be a potential marker for poor prognosis. Dig. Dis. Sci. 2011, 56, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-N.; Chen, Z.-H.; Chen, W.-C. Novel circulating microRNAs expression profile in colon cancer: A pilot study. Eur. J. Med. Res. 2017, 22, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Yan, Z.; Yang, H.; Ni, H.; Zhang, L.; Wang, Y. Clinical value of combined detection of miR-1202 and miR-195 in early diagnosis of cervical cancer. Oncol. Lett. 2019, 17, 3387–3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Y.; Song, Q.; Wang, J.; Zhao, L.; Lv, J.; Gong, S. MiR-1202 functions as a tumor suppressor in glioma cells by targeting Rab1A. Tumor Biol. 2017, 39, 1010428317697565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yoo, Y.; Fan, H.; Kim, E.; Guan, K.-L.; Guan, J.-L. Regulation of Integrin β1 Recycling to Lipid Rafts by Rab1a to Promote Cell Migration*. J. Biol. Chem. 2010, 285, 29398–29405. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; LaRiviere, M.J.; Ko, J.; Till, J.E.; Christensen, T.; Yee, S.S.; Black, T.A.; Tien, K.; Lin, A.; Shen, H.; et al. A Multianalyte Panel Consisting of Extracellular Vesicle miRNAs and mRNAs, cfDNA, and CA19-9 Shows Utility for Diagnosis and Staging of Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. 2020, 26, 3248–3258. [Google Scholar] [CrossRef] [Green Version]
- Weng, Y.R.; Cui, Y.; Fang, J.Y. Biological functions of cytokeratin 18 in cancer. Mol. Cancer Res. 2012, 10, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Khushman, M.; Patel, G.K.; Laurini, J.A.; Bhardwaj, A.; Roveda, K.; Donnell, R.; Sherling, K.; Case, B.; Frankel, A.E.; Pai, S.; et al. Exosomal markers (CD63 and CD9) expression and their prognostic significance using immunohistochemistry in patients with pancreatic ductal adenocarcinoma. J. Gastrointest. Oncol. 2019, 10, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, A.; Yuan, X.; Zhang, Q.; Fang, T.; Wang, W.; Li, C. Downregulation of microRNA-409-3p promotes aggressiveness and metastasis in colorectal cancer: An indication for personalized medicine. J. Transl. Med. 2015, 13, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Ji, Q.; Xiao, J.; Li, F.; Wang, L.; Chen, Y.; Xu, Y.; Jiao, S. miR-409 Inhibits Human Non-Small-Cell Lung Cancer Progression by Directly Targeting SPIN1. Mol. Ther. Nucleic Acids 2018, 13, 154–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhang, Y.; Huang, Z.; Gu, H.; Zhou, K.; Yin, X.; Xu, J. MiR-409-3p Inhibits Cell Proliferation and Invasion of Osteosarcoma by Targeting Zinc-Finger E-Box-Binding Homeobox-1. Front. Pharmacol. 2019, 10, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.B.; Du, Y.; Tian, Y.; Ji, Z.G.; Yang, P.Q. MiR-1299 functions as a tumor suppressor to inhibit the proliferation and metastasis of prostate cancer by targeting NEK2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 530–538. [Google Scholar] [CrossRef]
- Vicentini, C.; Calore, F.; Nigita, G.; Fadda, P.; Simbolo, M.; Sperandio, N.; Luchini, C.; Lawlor, R.T.; Croce, C.M.; Corbo, V.; et al. Exosomal miRNA signatures of pancreatic lesions. BMC Gastroenterol. 2020, 20, 137. [Google Scholar] [CrossRef]
- Butera, G.; Pacchiana, R.; Mullappilly, N.; Margiotta, M.; Bruno, S.; Conti, P.; Riganti, C.; Donadelli, M. Mutant p53 prevents GAPDH nuclear translocation in pancreatic cancer cells favoring glycolysis and 2-deoxyglucose sensitivity. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2018, 1865, 1914–1923. [Google Scholar] [CrossRef]

{kind=link}
| Biomarker | Patient Cohort | Method | Significant Comments | Ref. |
|---|---|---|---|---|
| Exosomal miR-21, -155 | PDAC (n = 27), Chronic Pancreatitis (n = 8) | qPCR | Exosomal miR-21 and Exosomal miR-155 in pancreatic juice samples could differentiate PDAC from CP with significant AUC. | [20] |
| Exosomal miR-196a, -1246 | Localized PDAC (n = 15), HC (n = 15) | qPCR | Exosomal miR-196a and miR-1246 could discriminate between PDAC and controls with AUCs of 0.81 and 0.73 respectively. miR-196a was a better indicator of PDAC, whereas miR-1246 was a better indicator of IPMNs (p = 0.0053, and p < 0.0001 respectively). | [21] |
| Exosomal KRAS codon 12/13 mutations | PDAC (n = 194), Disease Controls (n = 37) | ddPCR | ExoKRAS level significantly correlated with candidates for surgical resection (OR = 38.4), and is an independent prognostic marker for PFS and OS (HR = 2.28 and 3.46 respectively). ExoKRAS could predict disease progression at significantly earlier times than both CA 19-9 and radiological imaging (sensitivity & specificity 79% & 100% respectively). | [22] |
| Exosomal KRAS codon 12/13 mutations | PDAC (n = 127), HC (n = 136) | ddPCR | ExoKRAS detected at a higher rate than ctDNA KRAS across all PDAC stages. ExoKRAS could predict PDAC with a sensitivity and specificity of 75.4% and 92.6% respectively. Pre-surgery exoKRAS MAF of <1% was associated with disease-free survival post resection. | [23] |
| ctDNA KRAS codon 12/61 mutations | Resectable PDAC (n = 221), HC (n = 182) | PCR-based SafeSeqS | KRAS mutations were identified in 30% of PDAC cases, more frequently in stage II and larger tumors than smaller stage I tumors. Agreement between mutations identified in ctDNA and in the primary tumor was 100%. | [24] |
| ctDNA KRAS codon 12 mutations | Metastatic PDAC (n = 17) | Ion Torrent PGM sequencer | Significant and rapid increase in ctDNA levels was associated with poor prognosis, and a sudden significant decrease in ctDNA was associated with an improved prognosis (r = −0.76, p = 0.03). | [25] |
| ctDNA KRAS codon 12/13 mutations | PDAC (n = 45) | qPCR with PNA Clamping | Postoperative detection of ctDNA KRAS is a prognostic marker for DFS (HR = 2.919). Transformation from pre-op KRAS negative to post-op KRAS positive indicated poor OS (HR = 9.419). | [26] |
| ctDNA KRAS codon 12/13 mutations | PDAC (n = 27) | ddPCR | Increase in ctDNA KRAS level during treatment was associated with decreased PFS and OS (median PFS 2.5 vs. 7.5 months, median OS 6.5 vs. 11.5 months). | [27] |
| ctDNA KRAS codon 12/13 mutations | Metastatic PDAC (n = 31), Locally Advanced PDAC (n = 24) | ddPCR | Best prognosis was identified in patients with wild-type KRAS, followed by KRAS mutation with no copy number gain, worst prognosis was associated with increasing KRAS mutation copy number (median survival 10.6, 5.5, and 2.5 months respectively). | [28] |
| miR-155, -196a, -720, -141 | Nodal Metastasis PDAC (n = 10), No Metastasis PDAC (n = 10), HC (n = 10) | qPCR | Significantly higher expression of miR-155, miR-196a and lower expression of miR-720, miR-141 in PDAC with nodal metastasis versus without. Upregulation of miR-720, miR-141 resulted in decreased cellular aggressiveness and increased chemosensitivity in PDAC cell lines. | [29] |
| miR-93, -16, -548d-3p, -320a, -4468, -3120–3p, -4713–5p, -103a, -155, -4770, -181a, -221, -151b | PDAC (n = 60), HC (n = 26) | qPCR | 13 miRNAs in EUS FNA samples could distinguish PDAC from controls with high accuracy (AUC > 0.9). | [30] |
| miR-103a, -155, -181a, -181b, -93 | IPMN (n = 9), HC (n = 26) | qPCR | 5 miRNAs in EUS FNA samples could distinguish IPMNs from controls with high accuracy (AUC > 0.9). | [30] |
| miR-1915-3p, -371b-5p, -1202, -4669, -3679-5p, -6088, -4499, -7107-5p | Stage I-III PDAC (n = 15), HC (n = 4) | Agilent Microarray | 8 miRNAs significantly associated with lymph node metastasis. Of note miR-4669 and miR-1202, displayed decreased expression in cases where lymph node metastasis occurred. | [31] |
| miR-34a-5p, -130a-3p, -222-3p | Stage II PDAC (n = 136), HC (n = 73) | Abcam Fireplex-Oncology Panel | Combination miRNA with CA 19-9 improved upon CA 19-9s diagnostic ability. All 3 miRNAs identified are associated with metastasis. | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGowan, R.; Sally, Á.; McCabe, A.; Moran, B.M.; Finn, K. Circulating Nucleic Acids as Novel Biomarkers for Pancreatic Ductal Adenocarcinoma. Cancers 2022, 14, 2027. https://doi.org/10.3390/cancers14082027
McGowan R, Sally Á, McCabe A, Moran BM, Finn K. Circulating Nucleic Acids as Novel Biomarkers for Pancreatic Ductal Adenocarcinoma. Cancers. 2022; 14(8):2027. https://doi.org/10.3390/cancers14082027
Chicago/Turabian StyleMcGowan, Ryan, Áine Sally, Anthony McCabe, Brian Michael Moran, and Karen Finn. 2022. "Circulating Nucleic Acids as Novel Biomarkers for Pancreatic Ductal Adenocarcinoma" Cancers 14, no. 8: 2027. https://doi.org/10.3390/cancers14082027
APA StyleMcGowan, R., Sally, Á., McCabe, A., Moran, B. M., & Finn, K. (2022). Circulating Nucleic Acids as Novel Biomarkers for Pancreatic Ductal Adenocarcinoma. Cancers, 14(8), 2027. https://doi.org/10.3390/cancers14082027