1. Introduction
Immunotherapy is a recently discovered treatment that stimulates or inhibits the immune system from intensifying or decreasing an immune response and has rapidly become a significant treatment for multiple types of solid cancers [
1].
In colorectal cancer, immune checkpoint therapy is approved for treating tumours with a mismatch repair deficiency (MMR-D) and microsatellite instability-high (MSI-H) phenotype [
2]. MSI results from mutations in the DNA mismatch repair genes and is found in 15% of sporadic colorectal cancers [
3].
Present immune checkpoint inhibitors (ICIs) are ineffective in tumours that are microsatellite-stable (MSS) or have low levels of microsatellite instability (MSI-L) [
4]. Research on new inhibitory receptors for immunotherapy is essential to expand the patient group that benefits from immunotherapy.
The B7 family molecules have received growing attention in recent years and are essential for regulating T-cell responses. The members of the B7 family function as immune regulatory ligands that exhibit interaction with the CD28 receptors family [
5,
6]. B7H3 (CD276) is a newly found immune checkpoint of the B7 family, representing a good target for cancer immunotherapy. The human B7-H3 is a type 1 transmembrane glycoprotein with two isoforms: 2IgB7-H3 (B7-H3 VC) and 4IgB7-H3 (B7-H3 VCVC) [
7,
8]. The 2IgB7-H3 structure incorporates single extracellular V- and C-like Ig domains, a transmembrane region, and a 45-aa cytoplasmic tail. The 4IgB7-H3 isoform contains two identical pairs of IgV-like and IgC-like domains (four Ig-like domains), so this isoform is mostly expressed on the cell surface in human mononuclear cells and cancer cells [
9,
10]. The FG loop of the IgV domain of B7H3 [
11] is critical for inhibiting T-cell proliferation. B7H3, as an immune checkpoint, is overexpressed on malignant cells with limited heterogeneity [
12]. It was reported that B7H3 has a dual role in regulating the immune system, playing both costimulatory and coinhibitory functions. As a costimulator of the immune system, B7H3 promotes cellular immunity and upregulates IFNγ production in the presence of TCR signalling. The inhibitory functions of the studied immune checkpoint are primarily associated with suppressing T-cell activation and proliferation through NFAT, NFκB and AP-1 factors, the primary regulators of TCR gene transcription [
13]. The knowledge about the role of B7H3 in colorectal cancer is insufficient and needs more investigation. Importantly, previous studies demonstrated that B7H3 has a crucial role in promoting epithelial-to-mesenchymal transition (EMT), invasion, metastasis and chemotherapy resistance in CRC [
14,
15,
16].
Here, we aimed to assess the expression of B7H3 regarding clinicopathological parameters, including MSI/MSS status, CD8+ T-cells, histopathological features: budding, tumour-infiltrating lymphocytes (TILs), TNM scale, and grading. Moreover, we examined the B7H3-related pathways using available online datasets. Our research also aimed to investigate the immunological context of B7H3 expression through the 48-cytokine screening panel, immunogenic elements, and immune landscape.
4. Discussion
Immune Checkpoint Therapy (ICT) is a new approach to cancer treatment aimed at strengthening anti-tumour response mediated by T-cells and is characterized by high efficiency. B7 family and its most known member—PD-L1—are known for regulating immune response mediated by T-cells and exhibiting significant antitumour effect, which has been widely described in several malignancies. Due to the limited possibilities of therapies targeting the PD-1/PD-L1 axis in CRC, new members of the B7 family, including B7H3, are widely investigated. B7H3, a recently discovered protein belonging to the B7 family that acts as a T-cell inhibitor promoting tumour cell invasiveness rather than a T-cell stimulator as previously classified, has gained increasing attention in the anti-cancer battlefield [
27]. Numerous receptors have been reported to interact with different B7 family members, but the B7H3 receptor remains unexplored. Previously, there was some evidence that Trem-like transcript 2 (TREML2) could act as a putative B7H3 receptor, but the results were controversial [
28,
29]. Currently, the Interleukin receptor IL20RA is considered a possible B7H3 receptor, however, confirmation research is still ongoing [
30]. It seems that controversies about the costimulatory or coinhibitory role of B7H3 may be associated with an unidentified B7H3 receptor.
On the mRNA level, B7H3 is widely expressed in most normal tissues, but the presence of B7H3 protein is strictly limited, suggesting a significant role in post-transcriptional regulation of the B7H3 gene [
31]. On the contrary, B7H3 expression in tumour tissue is highly upregulated, which was also confirmed in CRC studies [
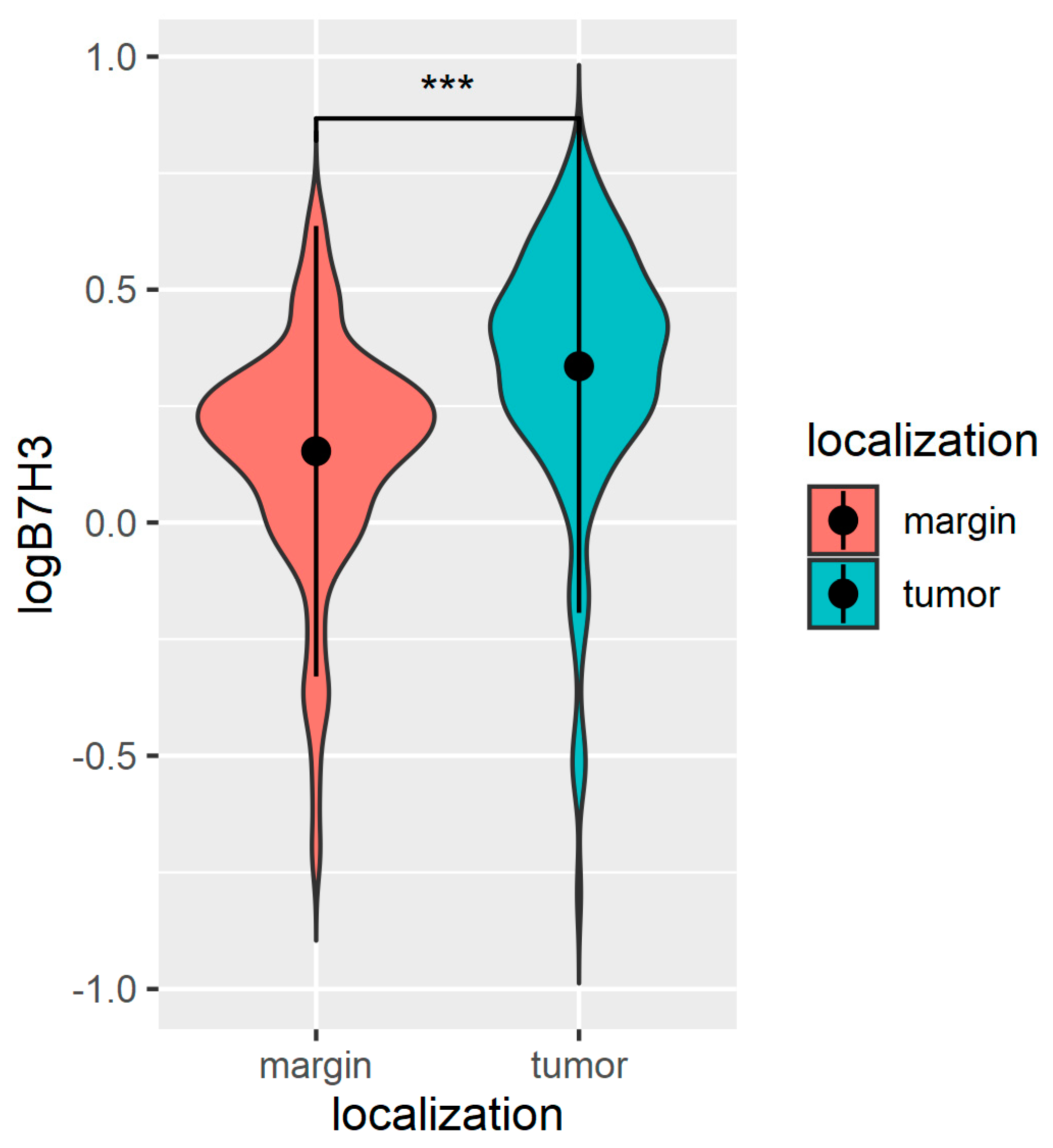
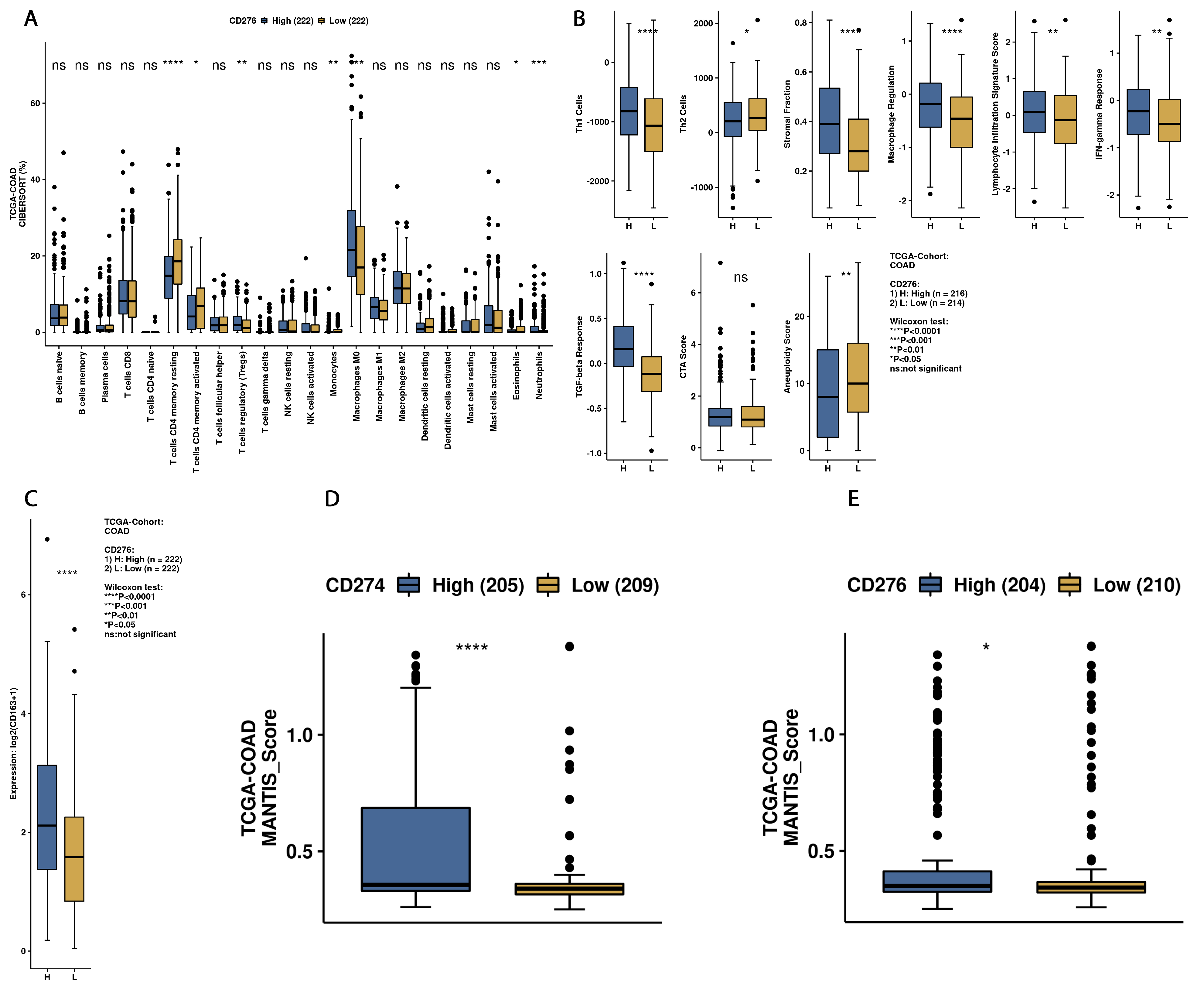
32]. Similarly to these previous findings, we also found significantly higher levels of B7H3 protein in CRC tumour tissue compared with normal adjacent mucosa, in contrast to the percentage of tumours that displayed positive staining for B7H3 in immunohistochemistry, which was 30% for all study cohorts. The percentage of B7H3-positive tumours differs considerably between particular studies. Sun et al. and Mao et al. reported 50% positive B7H3 tumours in their cohort, while according to other authors, B7H3 expression ranged to more than 80% CRC tumours [
32,
33]. Furthermore, we observed that B7H3 expression was independent of MSI/MSS status. These findings are consistent with the data obtained from CAMOIP, showing that differences in MANTIS score predicting MSS/MSI status of tumour are much lower for B7H3 compared with PD-L1, for which a close link with MSI status was well confirmed in CRC. Additionally, Zhao et al. did not report an association between B7H3 expression and MS/MSS status in the CRC cohort [
16].
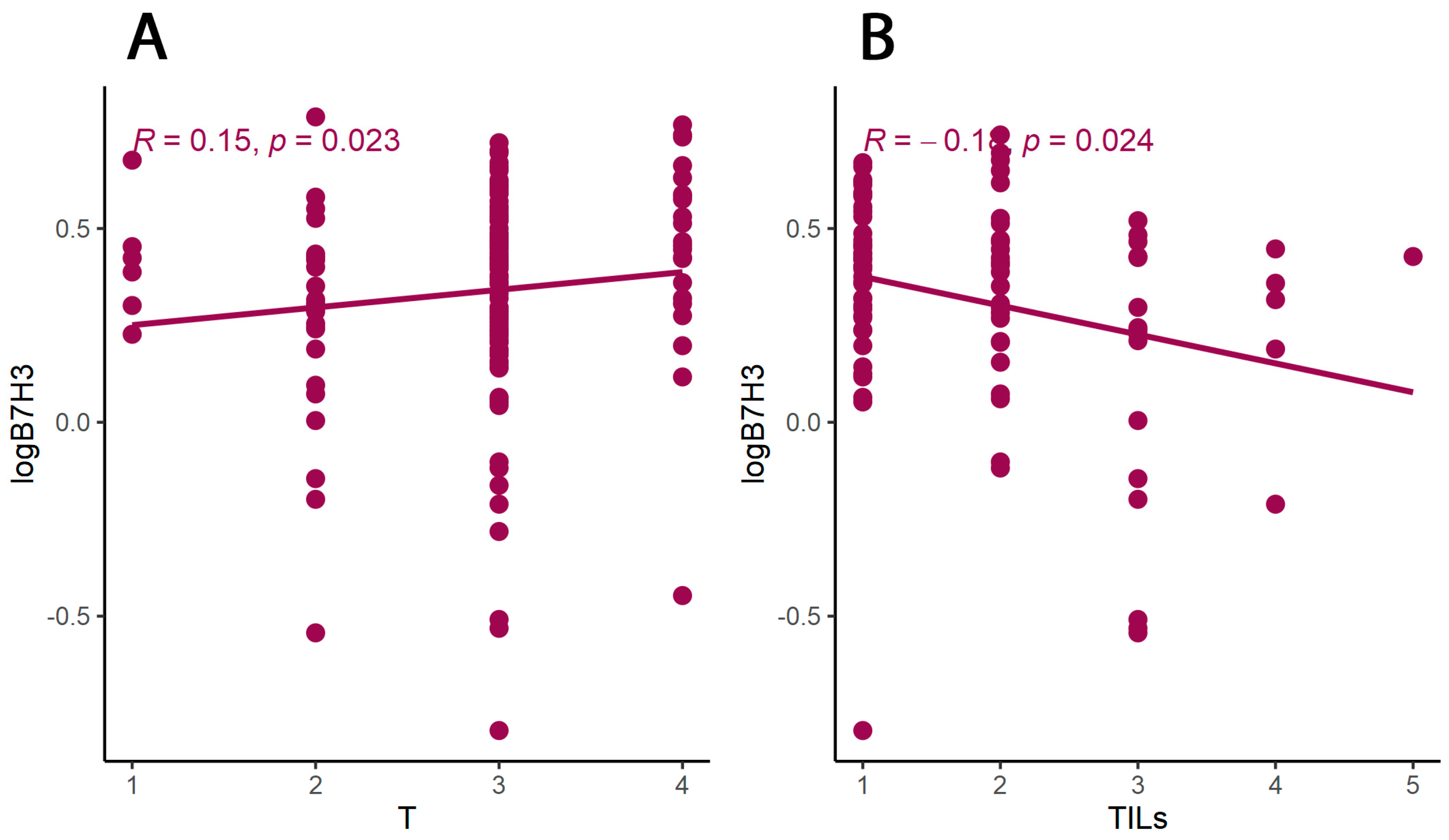
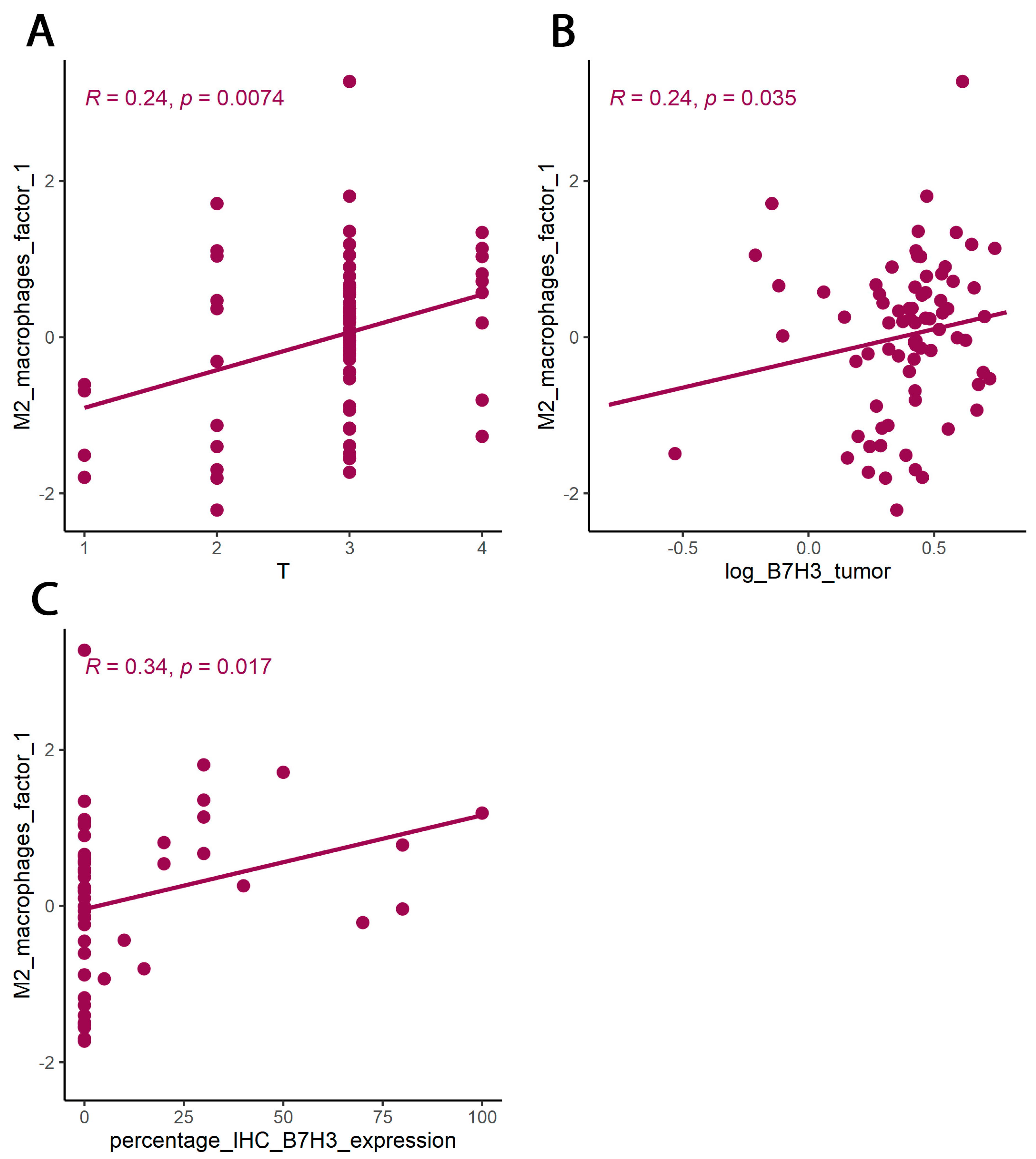
We explored the relationship between B7H3 expression and the clinicopathological parameters of the patients. The results showed a positive correlation between B7H3 tumour concentration and the T parameter, thus suggesting that B7H3 contributed to CRC progression. Similarly, Ingebrigtsen et al. reported that high B7H3 expression was associated with the advanced TNM stage in CRC [
15]. Furthermore, a positive relationship between B7H3 expression and TNM stage was confirmed in a variety of solid tumours, including breast, lung, and kidney tumours [
27]. In several studies, B7H3 was proven to play a crucial role not only in evasion from immune surveillance but also in non-immune processes promoting malignancy progression, such as epithelial-mesenchymal transition, chemoresistance, cell survival, migration and invasion, which makes B7H3 an even more appealing therapeutic target for CRC [
27,
34].
As we suspected, B7H3 levels in the tumour were negatively correlated with tumour infiltrating lymphocytes (TILs) score. In CRC, tumour-infiltrating lymphocytes (TILs) consist of different populations of cells (T lymphocytes, B lymphocytes, NK cells) and are regarded as positive prognosis factors [
35]. B7H3 was negatively related to TILS score in lung cancer [
36]. At the same time, other members of the B7 family, such as PD-L1 and B7H4, were reported to correlate negatively with TILs in colorectal and gastric cancer [
20,
37]. While B7H3 tumour concentration was negatively associated with TILs score, there was no association between B7H3 expression and the density of CD8+ T-cells infiltrating a tumour. CD8 T-cells are the main effector of antitumour response partially controlled by immune checkpoints, which affect the activation of antigen-specific T-cells [
38]. During malignant development, coinhibitory immune checkpoints allow tumour cells to escape from immune surveillance by accelerating T-cells exhaustion, thus, immune checkpoint blockade was found to induce long-term antitumour response mediated by CD8+ T-cells [
38]. In addition, high expression of B7H3 was shown to be associated with a lower number of CD8+ T-cells in osteosarcoma, endometrial, and lung cancers [
36,
39,
40]. Still, other CRC studies did not confirm this link [
16,
41].
Tumour-associated macrophages (TAMs) that promote tumour invasion and metastasis elicit phenotypes of M2-polarized macrophages [
41]. Macrophage polarization is a process in which macrophages obtain distinct functional phenotypes (M1 and M2) in response to different microenvironmental signals and factors [
42]. Macrophages M1 induced by interferon-gamma and LPS can kill tumour cells and pathogens, whereas macrophages M2 induced by IL-4, IL-10, and IL-13 exhibit anti-inflammatory activity and contribute to tumour invasion and angiogenesis [
41,
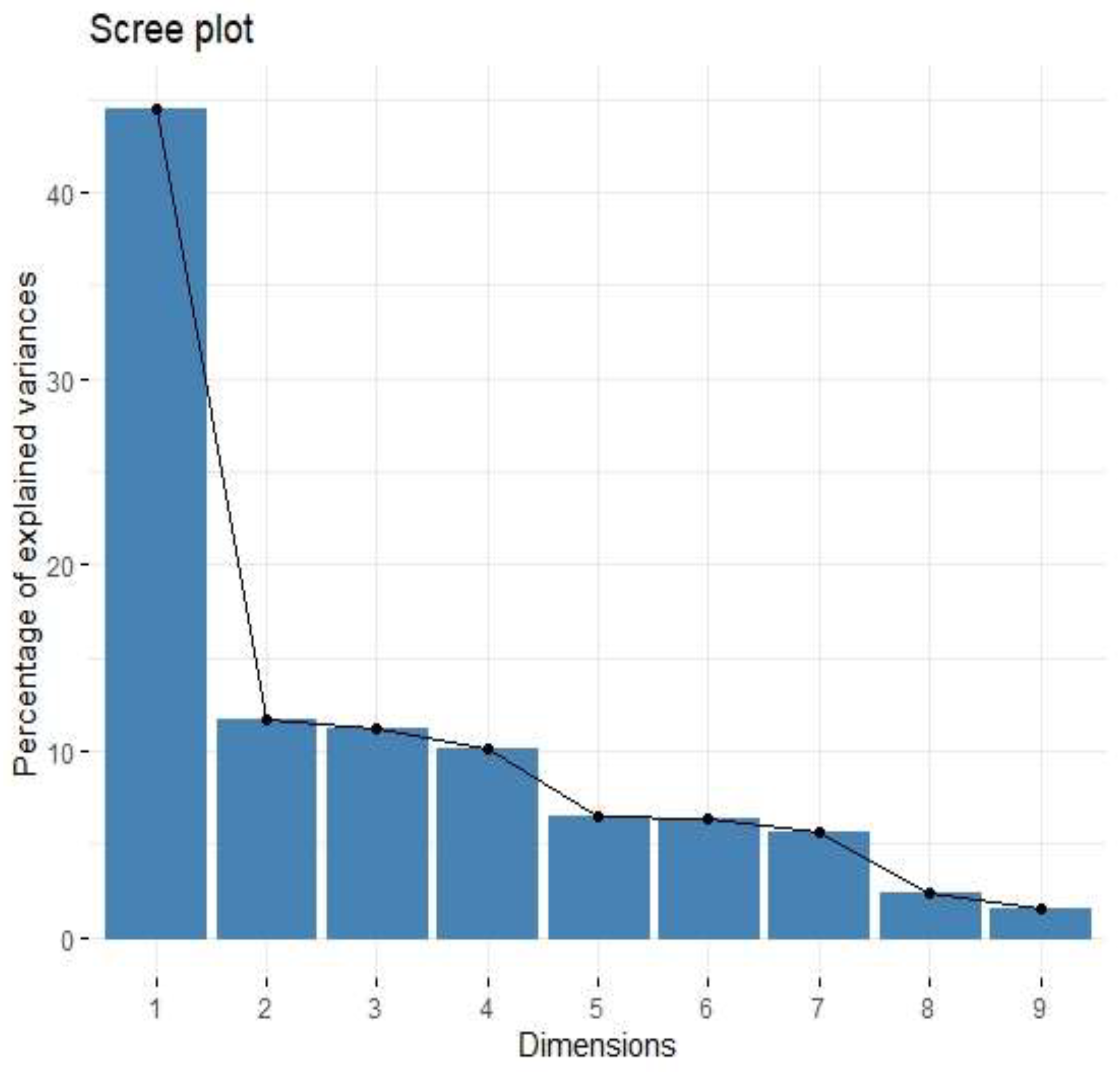
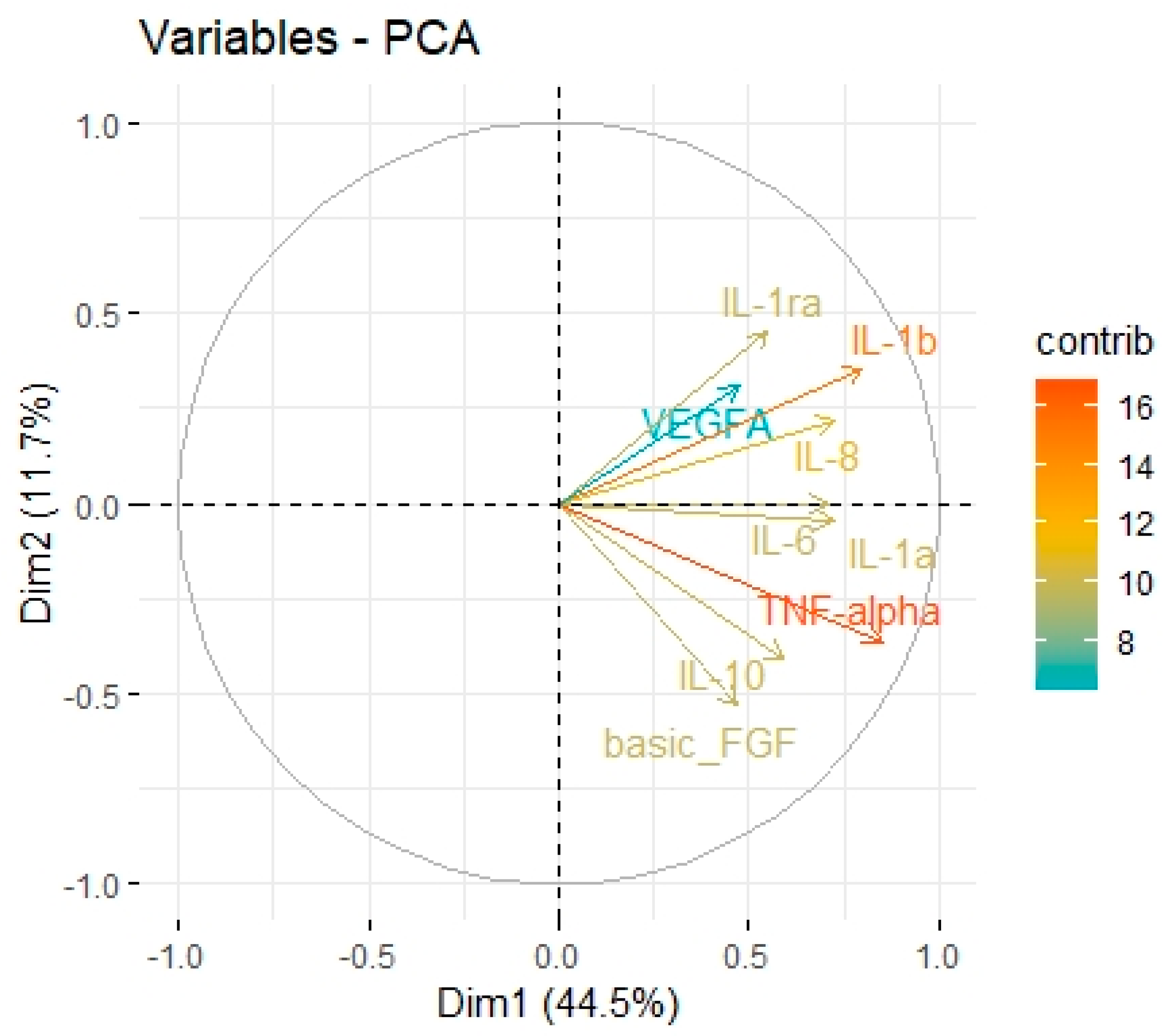
43]. Immune checkpoints, including B7 family members in the tumour microenvironment, are suspected of promoting TAMs polarization into M2 phenotypes. Performing Principal Component Analysis among molecules associated with M2-macrophages polarization (IL-1b, IL-1ra, IL-1a, TNF-alpha, IL-10, IL-6, Il-8, VEGF, FGF-basic), we found a positive correlation between Factor 1 obtained from PCA and B7H3 expression both in tissue homogenates and IHC staining. Factor 1 was also positively associated with the T parameter of patients. Similarly, CAMOIP analysis revealed that the M2 macrophage marker, CD163, is significantly higher in tumours with high B7H3 expression. However, a significant difference in immune cell scores between B7H3 high and low-expression tumours was found for M0 macrophage populations, but not M2 macrophages (CIBERSORT analysis). Previous studies have found an association between B7H3 expression and M2 polarization of macrophages in CRC, hepatocellular carcinoma and ovarian cancer [
41,
44,
45]. Furthermore, it has been demonstrated that M2 polarization induced by B7H3 is mediated by the JAK2-STAT3 pathway in multiple myeloma [
46]. These findings suggest that M2 TAMs may play a crucial role in mediating the immunosuppressive effects of B7H3.
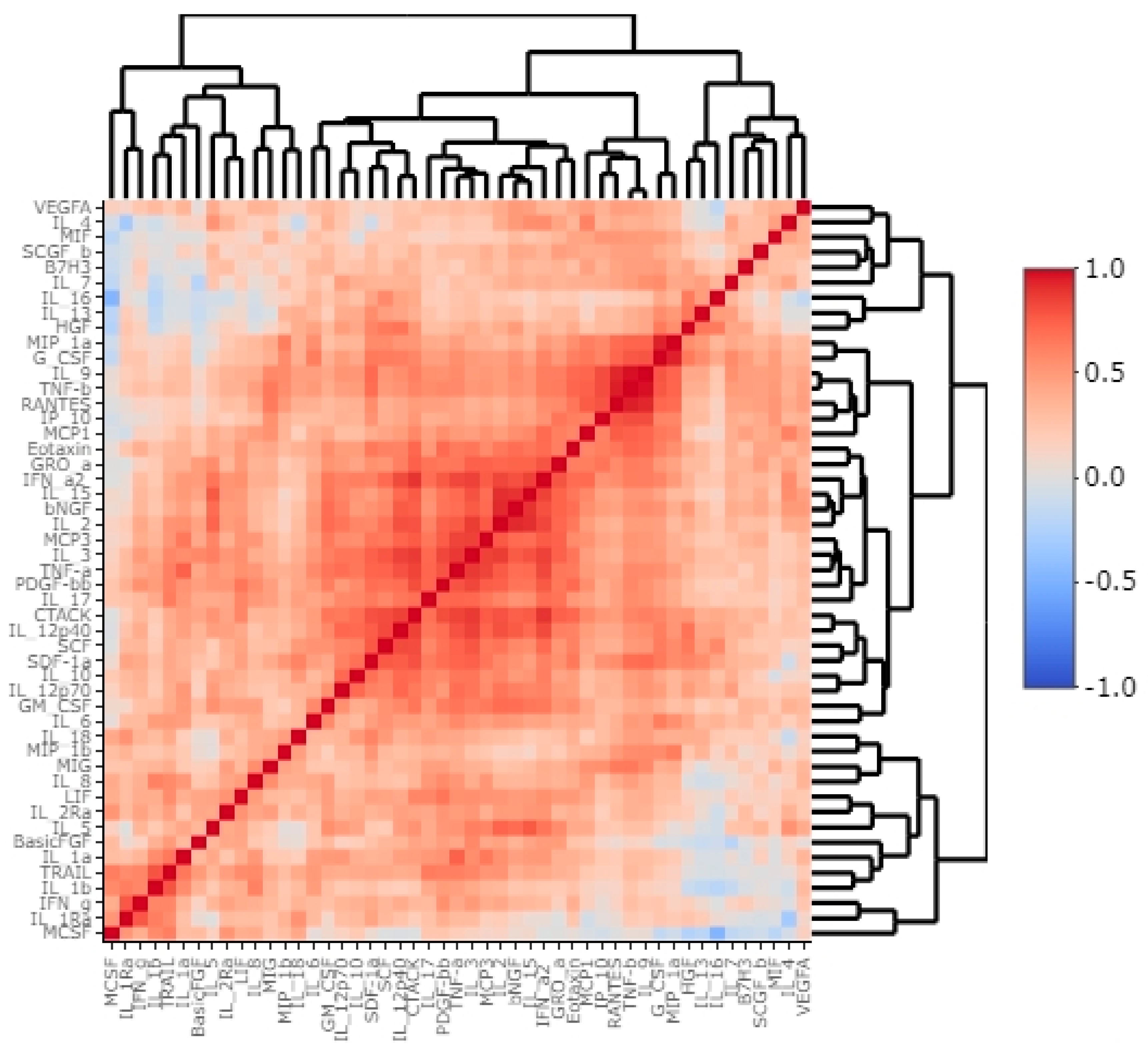
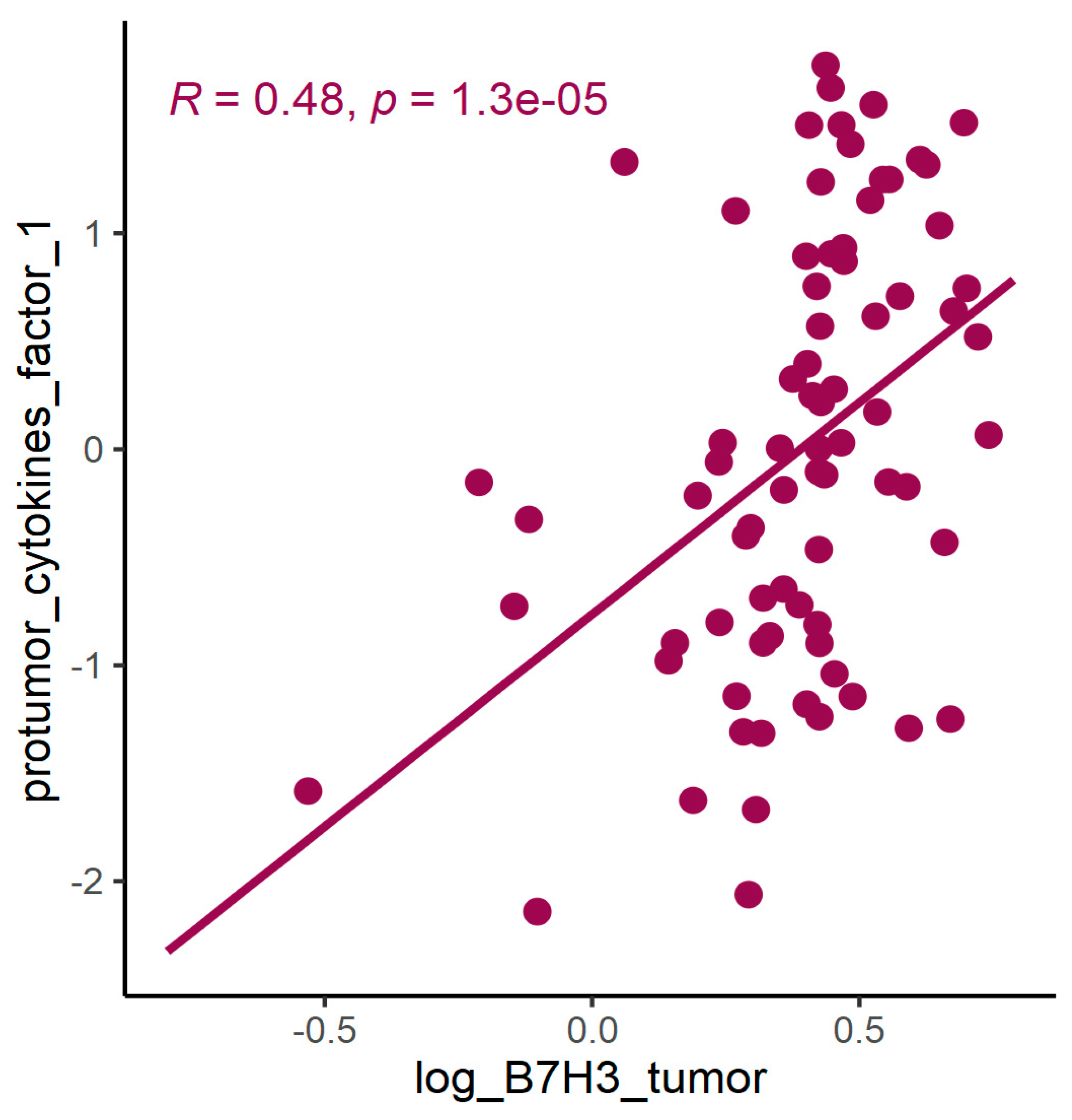
Exploring the cytokine network in the CRC tumour microenvironment may augment the current understanding of interrelationships between cytokines, chemokines, growth factors, and immune checkpoints and their common influence on TME modulation and tumour progression. As immune checkpoints blockade may reshape TME and restore antitumour response, investigating associations between B7H3 expression and functional groups of TME molecules could be extremely valuable for predicting the effects of B7H3 targeted therapy. Analysis of cytokines showed that B7H3 is related to protumour cytokines including IL-1β, IL-1Rα, IL-4, Il-5, IL-8, IL-9, IL-10, IL-13, IL-17A and TNFβ. Factor 1 obtained from PCA containing these molecules was positively related to B7H3 expression. IL-1β secreted by TAMs and neutrophils was found to promote cancer cell proliferation and recruit myeloid-derived suppressor cells (MDSCs) supporting tumour growth [
47]. IL-4 expressed by Th2 cells contributed to cancer cell proliferation, epithelial-mesenchymal transition and metastasis [
47]. Since IL-13 shares the same receptor with IL-4 (IL-4R alpha), its biological effects in CRC are similar to IL-4. IL-8 is involved in numerous processes supporting tumour progression, including proliferation, migration, invasion, survival of CRC cells and tumour angiogenesis [
48]. IL-10, produced mainly by M2 macrophages and Th2 cells, shapes immunosuppressive TME by reducing the production of proinflammatory cytokines and Th1 antitumour cytokines, inhibiting the proliferation of T-cells and decreasing the expression of MHC class II antigens [
48]. CD4+ Th17 cells are the main source of IL-17 which exhibits an immunosuppressive effect in TME by recruiting MDSCs, increasing CRC cell proliferation, upregulating VEGF production and activating oncogenic STAT-3 factor [
48,
49]. TNF-beta signalling was shown to be associated with tumour cell proliferation, epithelial-mesenchymal transition, invasion and metastasis formation [
50]. Finally, the role of IL-5 and IL-9 remains controversial due to their anti- and protumour effects in several studies [
51,
52].
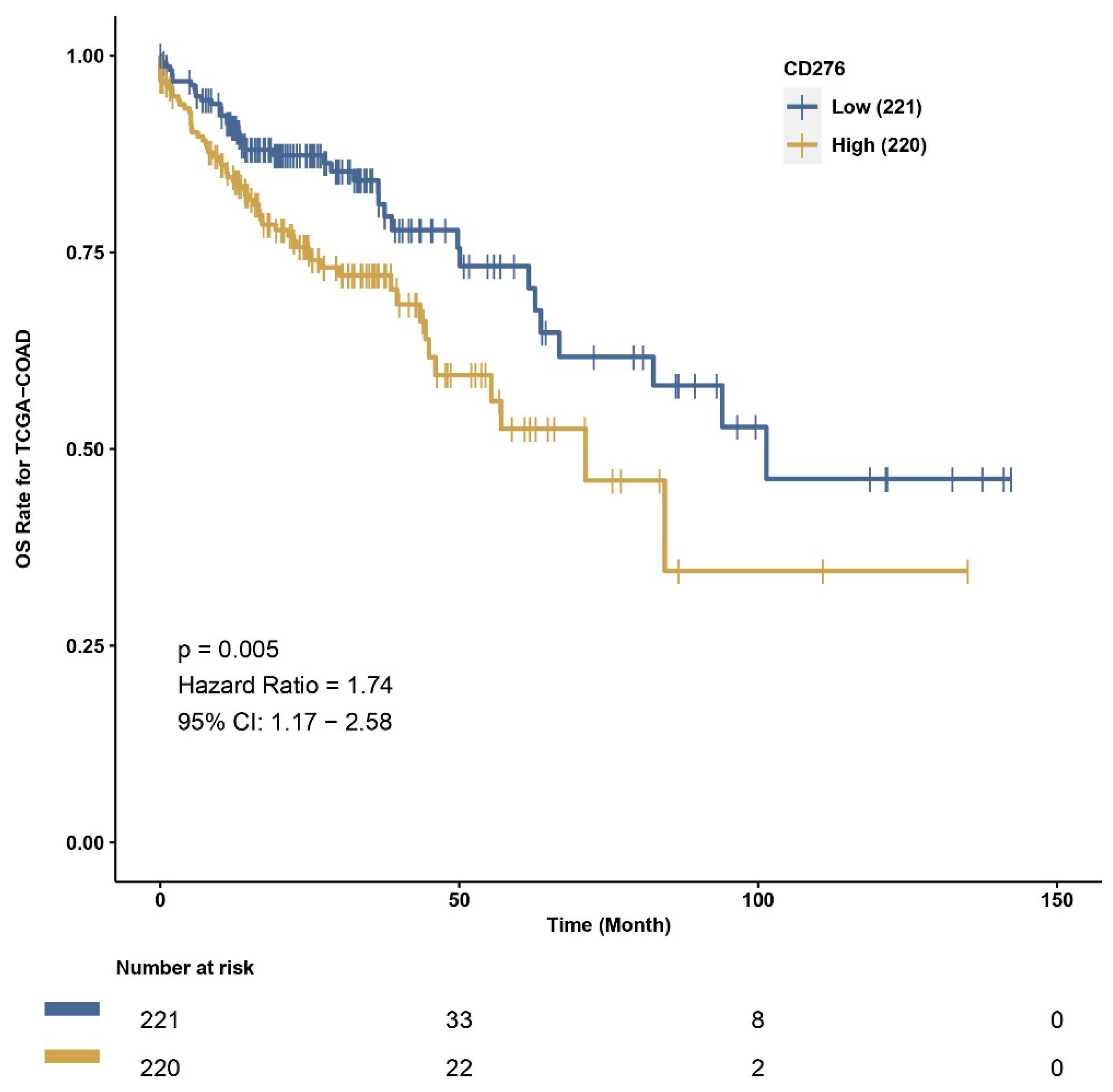
We also used CAMOIP, a web-based tool analyzing the TCGA-COAD dataset, to explore the immune composition of CRC according to high and low B7H3 expression. We found that upregulation of B7H3 affects only a few populations of immune cells, including CD4+ memory resting T-cells, CD4+ memory activated T-cells, regulatory T-cells, monocytes, macrophages M0, eosinophils and neutrophils. However, the immune score for Th1 cells, stromal fraction, macrophages regulation, lymphocytes infiltration, IFN gamma response and TGF beta was higher in a group with high B7H3 expression, indicating B7H3 contribution to immunological processes associated with tumourigenesis. Additionally, high B7H3 expression was associated with decreased survival in the TCGA-COAD dataset. This finding is consistent with clinical studies in which B7H3 was demonstrated to predict shorter overall survival and disease-free survival in CRC cohorts [
38].
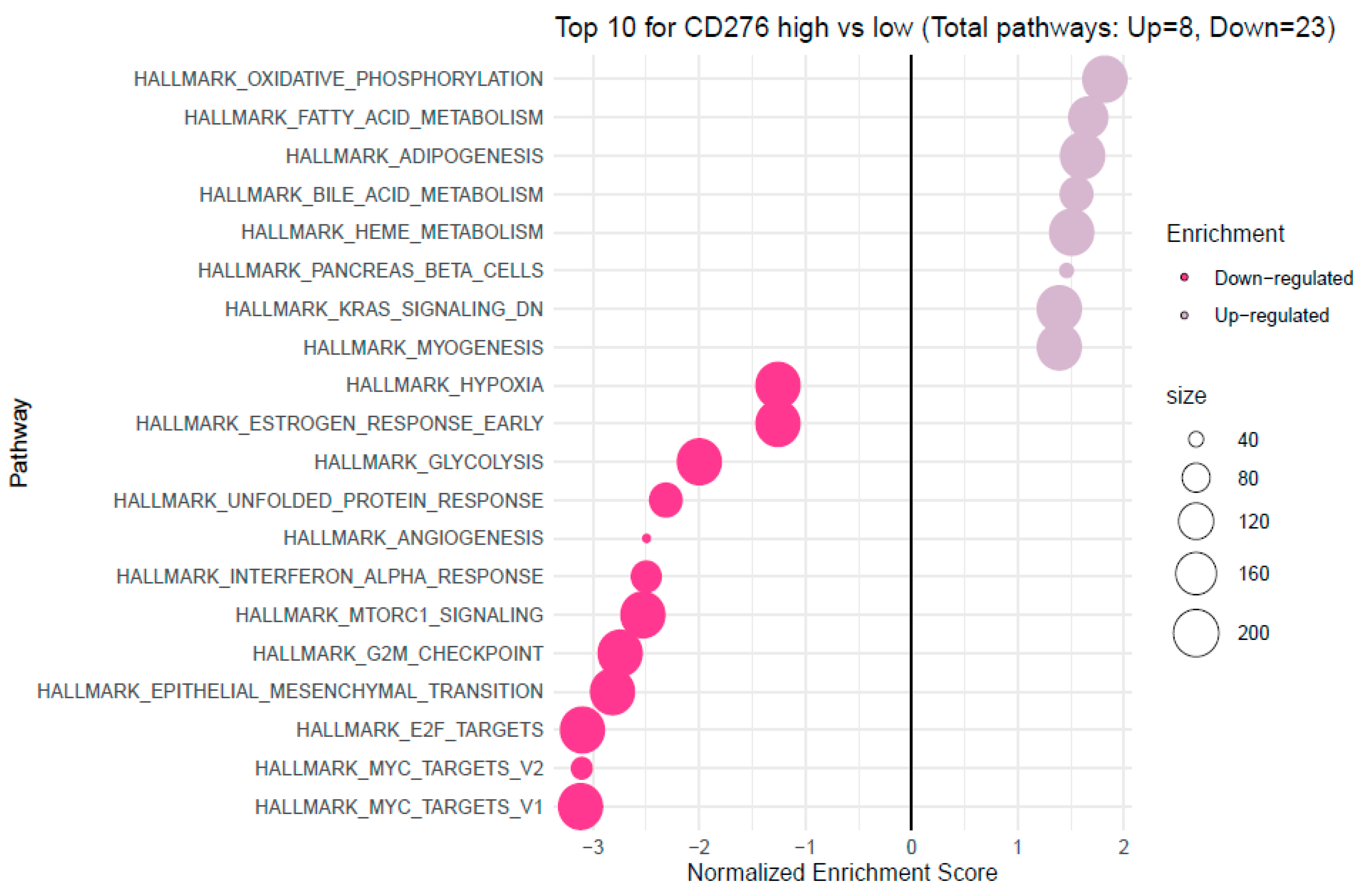
Analyzing the FieldEffectCrc dataset, we explored functions, processes and upregulated pathways associated with high B7H3 expression using GSEA and GO enrichment analysis.
Enrichment analysis showed that B7H3 is related to upregulated pathways involved in oxidative phosphorylation, fatty acid metabolism and downregulation of KRAS signalling. Recently, Picarda et al. demonstrated that B7H3 may play a pivotal role in regulating adipose tissue metabolism. B7H3 expression was upregulated in adipose tissue, with the highest levels in adipocyte progenitor cells and lower levels with the initiation of adipocyte differentiation. Additionally, the knockout of the B7H3 gene in progenitor cells led to impairment in aerobic metabolism and accumulation of fatty acids [
53]. Mutation in the KRAS gene was found to be more frequent in B7H3-positive lung adenocarcinoma, however, data regarding CRC are missing, and the link between B7H3 and KRAS needs further research and explanation.
Furthermore, high B7H3 expression is associated with downregulating pathways related to protooncogene MYC and cell-cycle control systems, such as G2/M Checkpoint and E2F target. Downregulation of the G2/M Checkpoint and E2F target indicates that one of the effects of B7H3 could be impaired G2/M phase cell cycle arrest. However, literature data about the association of B7H3 and the MYC gene are limited and concern only malignant gliomas in which a knockout of the B7H3 gene inhibits MYC expression [
54].
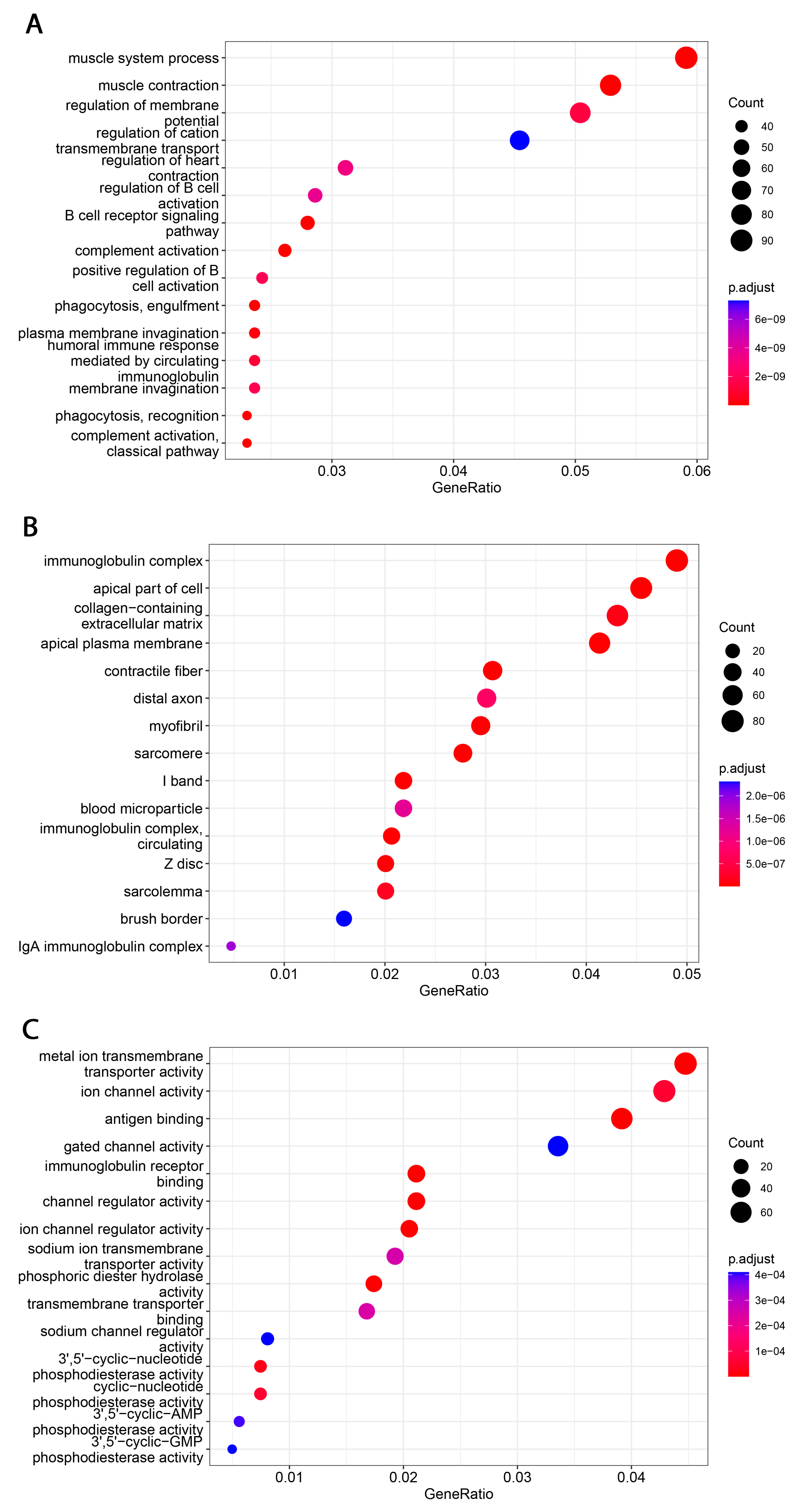
Functional analyses of B7H3 revealed its involvement in immunological-associated processes. Significantly enriched GO terms were related to various immunological functions, including innate and adaptive immunity, such as phagocytosis, complement activation, antigen binding, activation of B-cells and their receptors, and regulation of immunoglobulin complex.
Our findings improved our understanding of B7H3′s role in cancer immunity. In addition, the studies on colorectal tumour tissues allowed us to estimate clinical, pathological, and immunological parameters. We assessed B7H3 concentrations by ELISA, and IHC, determined MSI/MSS status, and estimated the immune landscape by cytokines screening panel and tumour-infiltrating CD8+ T-cells. The analysis based on mRNA expression profiles has significantly expanded the scope of research by reporting many significant molecular roles of B7H3. CAMOIP, a tool for comprehensive analysis of potential immunotherapy targets, allowed us to perform survival analysis according to low or high B7H3 expression in the TCGA cohort and determined the link between B7H3 expression, immune cell infiltration, Mantis score, and patients’ survival in CRC tumours. We studied the pathways activated in tumours with upregulated B7H3 expression with gene set enrichment analysis. Our future research direction focuses on further studies of B7H3, as a potential target for immunotherapy which requires extending research to cellular, animal, and clinical models. The other cancer therapy trends require studying the diagnostic and therapeutic potential of B7-H3 thoroughly, its cellular and molecular mechanisms, B7H3-mediated metastases, tumour-associated vasculature, and recurrence. Clinical data with a long-term follow-up are essential to understand the relevance of B7H3 expression.


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}