
KRAS p.G12C Mutation in Metastatic Colorectal Cancer: Prognostic Implications and Advancements in Targeted Therapies
 , , ,
, , ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. KRAS Mutations: Prognostic Divergences
3. KRAS Mutations: Linking Structure and Prognosis
4. Resurgence in KRAS Targeting: Overcoming Challenges
5. Sotorasib and Adagrasib: An Overview of Clinical Data
6. Pragmatic Considerations on the Clinical Use of KRAS Mutations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timar, J.; Kashofer, K. Molecular epidemiology and diagnostics of KRAS mutations in human cancer. Cancer Metastasis Rev. 2020, 39, 1029–1038. [Google Scholar] [CrossRef]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Biller, L.H.; Schrag, D. Diagnosis and Treatment of Metastatic Colorectal Cancer: A Review. JAMA 2021, 325, 669–685. [Google Scholar] [CrossRef]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Kim, H.J.; Na Lee, H.; Jeong, M.S.; Jang, S.B. Oncogenic KRAS: Signaling and Drug Resistance. Cancers 2021, 13, 5599. [Google Scholar] [CrossRef] [PubMed]
- László, L.; Kurilla, A.; Takács, T.; Kudlik, G.; Koprivanacz, K.; Buday, L.; Vas, V. Recent Updates on the Significance of KRAS Mutations in Colorectal Cancer Biology. Cells 2021, 10, 667. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shi, Q.; Wang, W.; Liu, J.; Ren, J.; Li, Q.; Hou, F. KRAS status and resistance to epidermal growth factor receptor tyrosine-kinase inhibitor treatment in patients with metastatic colorectal cancer: A meta-analysis. Color. Dis. 2014, 16, O370–O378. [Google Scholar] [CrossRef] [PubMed]
- Ottaiano, A.; Normanno, N.; Facchini, S.; Cassata, A.; Nappi, A.; Romano, C.; Silvestro, L.; De Stefano, A.; Rachiglio, A.M.; Roma, C.; et al. Study of Ras Mutations’ Prognostic Value in Metastatic Colorectal Cancer: STORIA Analysis. Cancers 2020, 12, 1919. [Google Scholar] [CrossRef]
- Schirripa, M.; Nappo, F.; Cremolini, C.; Salvatore, L.; Rossini, D.; Bensi, M.; Businello, G.; Pietrantonio, F.; Randon, G.; Fucà, G.; et al. KRAS G12C Metastatic Colorectal Cancer: Specific Features of a New Emerging Target Population. Clin. Color. Cancer 2020, 19, 219–225. [Google Scholar] [CrossRef]
- Chida, K.; Kotani, D.; Masuishi, T.; Kawakami, T.; Kawamoto, Y.; Kato, K.; Fushiki, K.; Sawada, K.; Kumanishi, R.; Shirasu, H.; et al. The Prognostic Impact of KRAS G12C Mutation in Patients with Metastatic Colorectal Cancer: A Multicenter Retrospective Observational Study. Oncologist 2021, 26, 845–853. [Google Scholar] [CrossRef]
- Fakih, M.; Tu, H.; Hsu, H.; Aggarwal, S.; Chan, E.; Rehn, M.; Chia, V.; Kopetz, S. Real-World Study of Characteristics and Treatment Outcomes among Patients with KRAS p.G12C-Mutated or Other KRAS Mutated Metastatic Colorectal Cancer. Oncologist 2022, 27, 663–674. [Google Scholar] [CrossRef]
- Modest, D.P.; Ricard, I.; Heinemann, V.; Hegewisch-Becker, S.; Schmiegel, W.; Porschen, R.; Stintzing, S.; Graeven, U.; Arnold, D.; von Weikersthal, L.F.; et al. Outcome according to KRAS-, NRAS- and BRAF-mutation as well as KRAS mutation variants: Pooled analysis of five randomized trials in metastatic colorectal cancer by the AIO colorectal cancer study group. Ann. Oncol. 2016, 27, 1746–1753. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.P.; Sutton, A.P.; Evans, J.P.; Clifford, R.; McAvoy, A.; Lewis, J.; Rousseau, A.; Mountford, R.; McWhirter, D.; Malik, H.Z. Specific mutations in KRAS codon 12 are associated with worse overall survival in patients with advanced and recurrent colorectal cancer. Br. J. Cancer 2017, 116, 923–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesweg, M.; Kasper, S.; Worm, K.; Herold, T.; Reis, H.; Sara, L.; Metzenmacher, M.; Abendroth, A.; Darwiche, K.; Aigner, C.; et al. Impact of RAS mutation subtype on clinical outcome—A cross-entity comparison of patients with advanced non-small cell lung cancer and colorectal cancer. Oncogene 2019, 38, 2953–2966. [Google Scholar] [CrossRef] [PubMed]
- Herdeis, L.; Gerlach, D.; McConnell, D.B.; Kessler, D. Stopping the beating heart of cancer: KRAS reviewed. Curr. Opin. Struct. Biol. 2021, 71, 136–147. [Google Scholar] [CrossRef]
- Pantsar, T. The current understanding of KRAS protein structure and dynamics. Comput. Struct. Biotechnol. J. 2019, 18, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Ostrem, J.M.; Peters, U.; Sos, M.L.; Wells, J.A.; Shokat, K.M. K-Ras(G12C) inhibitors allosterically control GTP affinity and effector interactions. Nature 2013, 503, 548–551. [Google Scholar] [CrossRef] [Green Version]
- O’brien, J.B.; Wilkinson, J.C.; Roman, D.L. Regulator of G-protein signaling (RGS) proteins as drug targets: Progress and future potentials. J. Biol. Chem. 2019, 294, 18571–18585. [Google Scholar] [CrossRef] [Green Version]
- Hallin, J.; Engstrom, L.D.; Hargis, L.; Calinisan, A.; Aranda, R.; Briere, D.M.; Sudhakar, N.; Bowcut, V.; Baer, B.R.; Ballard, J.A.; et al. The KRASG12C Inhibitor MRTX849 Provides Insight toward Therapeutic Susceptibility of KRAS-Mutant Cancers in Mouse Models and Patients. Cancer Discov. 2020, 10, 54–71. [Google Scholar] [CrossRef] [Green Version]
- Skoulidis, F.; Li, B.T.; Dy, G.K.; Price, T.J.; Falchook, G.S.; Wolf, J.; Italiano, A.; Schuler, M.; Borghaei, H.; Barlesi, F.; et al. Sotorasib for Lung Cancers with KRAS p.G12C Mutation. N. Engl. J. Med. 2021, 384, 2371–2381. [Google Scholar] [CrossRef]
- Hong, D.S.; Fakih, M.G.; Strickler, J.H.; Desai, J.; Durm, G.A.; Shapiro, G.I.; Falchook, G.S.; Price, T.J.; Sacher, A.; Denlinger, C.S.; et al. KRASG12C Inhibition with Sotorasib in Advanced Solid Tumors. N. Engl. J. Med. 2020, 383, 1207–1217. [Google Scholar] [CrossRef]
- Tamiya, Y.; Matsumoto, S.; Zenke, Y.; Yoh, K.; Ikeda, T.; Shibata, Y.; Kato, T.; Nishino, K.; Nakamura, A.; Furuya, N.; et al. Large-scale clinico-genomic profile of non-small cell lung cancer with KRAS G12C: Results from LC-SCRUM-Asia study. Lung Cancer 2022, 176, 103–111. [Google Scholar] [CrossRef]
- Georgeson, P.; Harrison, T.A.; Pope, B.J.; Zaidi, S.H.; Qu, C.; Steinfelder, R.S.; Lin, Y.; Joo, J.E.; Mahmood, K.; Clendenning, M.; et al. Identifying colorectal cancer caused by biallelic MUTYH pathogenic variants using tumor mutational signatures. Nat. Commun. 2022, 13, 3254. [Google Scholar] [CrossRef]
- Ma, H.; Brosens, L.A.A.; Offerhaus, G.J.A.; Giardiello, F.M.; De Leng, W.W.J.; Montgomery, E.A. Pathology and genetics of hereditary colorectal cancer. Pathology 2018, 50, 49–59. [Google Scholar] [CrossRef]
- Maurer, T.; Garrenton, L.S.; Oh, A.; Pitts, K.; Anderson, D.J.; Skelton, N.J.; Fauber, B.P.; Pan, B.; Malek, S.; Stokoe, D.; et al. Small-molecule ligands bind to a distinct pocket in Ras and inhibit SOS-mediated nucleotide exchange activity. Proc. Natl. Acad. Sci. USA 2012, 109, 5299–5304. [Google Scholar] [CrossRef]
- Cruz-Migoni, A.; Canning, P.; Quevedo, C.E.; Bataille, C.J.R.; Bery, N.; Miller, A.; Russell, A.J.; Phillips, S.E.V.; Carr, S.B.; Rabbitts, T.H. Structure-based development of new RAS-effector inhibitors from a combination of active and inactive RAS-binding compounds. Proc. Natl. Acad. Sci. USA 2019, 116, 2545–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrem, J.M.L.; Shokat, K.M. Direct small-molecule inhibitors of KRAS: From structural insights to mechanism-based design. Nat. Rev. Drug Discov. 2016, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Fakih, M.G.; Kopetz, S.; Kuboki, Y.; Kim, T.W.; Munster, P.N.; Krauss, J.C.; Falchook, G.S.; Han, S.-W.; Heinemann, V.; Muro, K.; et al. Sotorasib for previously treated colorectal cancers with KRASG12C mutation (CodeBreaK100): A prespecified analysis of a single-arm, phase 2 trial. Lancet Oncol. 2022, 23, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.-H.I.; Jänne, P.A.; Leal, T.A.; Rybkin, I.I.; Sabari, J.K.; Barve, M.A.; Bazhenova, L.A.; Johnson, M.L.; Velastegui, K.L.; Cilliers, C.; et al. First-in-Human Phase I/IB Dose-Finding Study of Adagrasib (MRTX849) in Patients With Advanced KRASG12C Solid Tumors (KRYSTAL-1). J. Clin. Oncol. 2022, 40, 2530–2538. [Google Scholar] [CrossRef]
- Yaeger, R.; Weiss, J.; Pelster, M.S.; Spira, A.I.; Barve, M.; Ou, S.-H.I.; Leal, T.A.; Bekaii-Saab, T.S.; Paweletz, C.P.; Heavey, G.A.; et al. Adagrasib with or without Cetuximab in Colorectal Cancer with Mutated KRAS G12C. N. Engl. J. Med. 2023, 388, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Cervantes, A.; Adam, R.; Sobrero, A.; van Krieken, J.H.; Aderka, D.; Aguilar, E.A.; Bardelli, A.; Benson, A.; Bodoky, G.; et al. ESMO consensus guidelines for the management of patients with metastatic colorectal cancer. Ann. Oncol. 2016, 27, 1386–1422. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | WT | Mutated | p | Mutation | p | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| p.G12D | p.G12V | p.G13D | p.G12A | p.G12C | p.G12S | |||||
| Age | ||||||||||
| <65 y | 50 | 40 | 9 | 9 | 11 | 3 | 5 | 3 | ||
| ≥65 y | 46 | 38 | 0.91 | 15 | 5 | 5 | 4 | 5 | 4 | 0.40 |
| Gender | ||||||||||
| M | 49 | 35 | 10 | 8 | 5 | 5 | 4 | 3 | ||
| F | 47 | 43 | 0.65 | 14 | 6 | 11 | 2 | 6 | 4 | 0.51 |
| Grading | ||||||||||
| G1/G2 | 20 | 17 | 4 | 3 | 5 | 2 | 1 | 2 | ||
| G3 | 76 | 61 | 0.87 | 20 | 11 | 11 | 5 | 9 | 5 | 0.79 |
| Side of primary tumor | ||||||||||
| Left | 45 | 32 | 9 | 8 | 8 | 1 | 3 | 3 | ||
| Right | 51 | 46 | 0.44 | 15 | 6 | 8 | 6 | 7 | 4 | 0.44 |
| pT | ||||||||||
| pT1/pT2 | 13 | 11 | 4 | 1 | 3 | 2 | 1 | 0 | ||
| pT3 | 48 | 45 | 12 | 11 | 8 | 2 | 7 | 5 | ||
| pT4 | 35 | 22 | 0.50 | 8 | 2 | 5 | 3 | 2 | 2 | 0.63 |
| pN | ||||||||||
| Not Involved | 21 | 23 | 8 | 4 | 6 | 3 | 0 | 2 | ||
| Involved | 75 | 55 | 0.25 | 16 | 10 | 10 | 4 | 10 | 5 | 0.36 |
| Metastatic involvement | ||||||||||
| One site | 23 | 25 | 9 | 6 | 4 | 3 | 1 | 2 | ||
| More than one | 73 | 53 | 0.23 | 15 | 8 | 12 | 4 | 9 | 5 | 0.53 |
| No. of chemotherapy lines | ||||||||||
| Two | 10 | 16 | 3 | 3 | 3 | 2 | 2 | 3 | ||
| More than two | 86 | 62 | 0.06 | 21 | 11 | 13 | 5 | 8 | 4 | 0.63 |
| Characteristics | Risk Factor (vs. Comparator) | mOS (Months) | No. of Events/Patients | p at Univariate | HR | 95% CI | p at Multivariate |
|---|---|---|---|---|---|---|---|
| Age | <65 y (vs. ≥65 y) | 15.5 (vs. 19.5) | 87/90 (vs. 77/84) | 0.1310 | 0.79 | 0.59–1.07 | 0.0965 |
| Gender | male (vs. female) | 19.2 (vs. 15.4) | 81/84 (vs. 83/90) | 0.7870 | 1.04 | 0.77–1.39 | 0.1181 |
| Metastatic involvement | >1 site (vs. 1 site) | 13.2 (vs. 23.6) | 132/138 (vs. 32/36) | 0.0005 | 1.77 | 1.28–2.45 | <0.0001 |
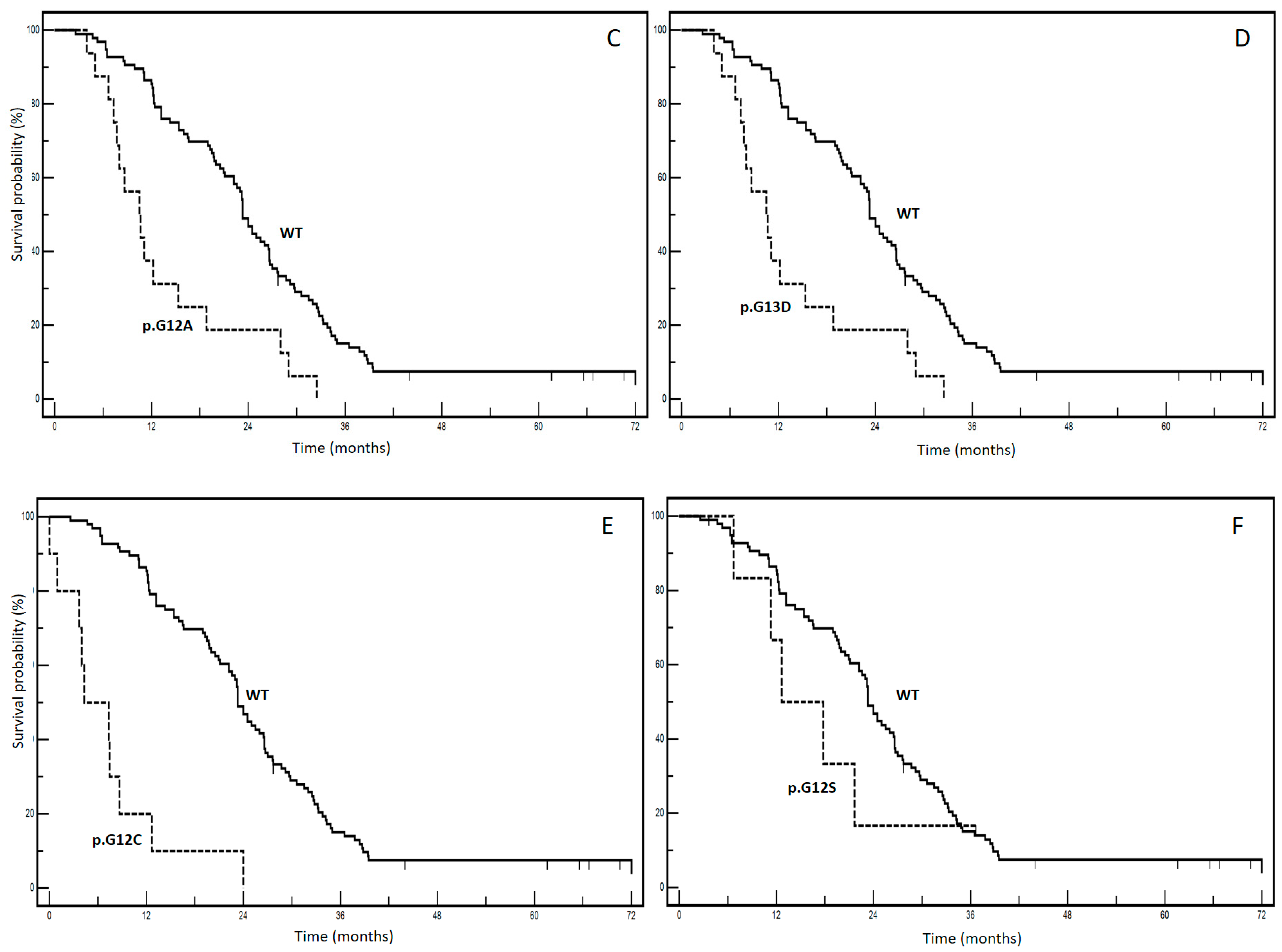
| KRAS mutations | p.G12D (vs. WT) | 12.0 (vs. 23.3) | 24/24 (vs. 89/96) | <0.0001 | 4.88 | 2.52–9.46 | <0.0001 |
| p.G12V (vs. WT) | 12.2 (vs. 23.3) | 14/14 (vs. 89/96) | 0.0508 | 2.01 | 0.99–4.08 | 0.0745 | |
| p.G13D (vs. WT) | 10.5 (vs. 23.3) | 16/16 (vs. 89/96) | 0.0253 | 5.49 | 2.41–12.4 | 0.0927 | |
| p.G12A (vs. WT) | 12.5 (vs. 23.3) | 6/7 (vs. 89/96) | 0.4127 | 1.50 | 0.56–4.00 | 0.2656 | |
| p.G12C (vs. WT) | 4.3 (vs. 23.3) | 10/10 vs. (89/96) | <0.0001 | 13.6 | 3.9–17.16 | <0.0001 | |
| p.G12S (vs. WT) | 12.6 (vs. 23.3) | 5/7 vs. (89/96) | 0.3541 | 1.68 | 0.56–5.05 | 0.1748 |
| Author, Year | Type of Study | Method | No. of Patients | % of p.G12C Variant | Comparison | mOSs | Co-Variates | p * |
|---|---|---|---|---|---|---|---|---|
| Schirripa M. et al., 2020 [9] | Retrospective | Sanger sequencing. | 839 | 17.0 | p.G12C vs. non-p.G12C | 29.0 vs. 36.7 | Gender, ECOG PS, primary tumor surgery, pT, pN, time of first metastasis, grading, number of metastatic sites. | 0.004 |
| Chida K. et al., 2021 [10] | Retrospective | Exon 2 through PCR-based kits. | 1632 | 2.8 | p.G12C vs. non-p.G12C | 21.2 vs. 27.3 | Age, gender, ECOG PS, primary tumor site, surgery on the primary tumor, time of first metastasis, histology, white blood cell count, serum albumin level, LDH level, serum C-reactive protein level, metastatic tumor site, number of metastatic sites. | 0.030 |
| Fakih M. et al., 2022 [11] | Retrospective | NGS. | 6477 | 3.7 | p.G12C vs. non-p.G12C vs. wt | 16.1 vs. 18.3 vs. 23.4 | None ** | None ** |
| Modest D.P. et al., 2016 [12] | Retrospective | Exon 2-4 through PCR-based kits. | 1239 | 2.2 | p.G12C vs. wt | 16.8 vs. 26.9 | Treatment, ECOG PS, gender, adjuvant chemotherapy, liver-limited disease and number of involved organs. | 0.001 |
| Jones R.P. et al., 2017 [13] | Retrospective | Pyrosequencing-based assay of codons 12, 13 and 61. | 392 | 3.8 | p.G12C vs. wt | 24.9 vs. 35.1 | None ** | None ** |
| Wiesweg M. et al., 2019 [14] | Retrospective | Exon 2, if KRAS wt proceeded to exon 3 through PCR-based kits. | 347 | 11.9 | p.G12C (included in an unfavorable cohort) vs. wt | 15.2 vs. 60.0 | Age, gender, stage, RAS prognostic cluster or RAS wild type, RAS DNA substitution category, grading, primary tumor location. | 0.087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottaiano, A.; Sabbatino, F.; Perri, F.; Cascella, M.; Sirica, R.; Patrone, R.; Capuozzo, M.; Savarese, G.; Ianniello, M.; Petrillo, N.; et al. KRAS p.G12C Mutation in Metastatic Colorectal Cancer: Prognostic Implications and Advancements in Targeted Therapies. Cancers 2023, 15, 3579. https://doi.org/10.3390/cancers15143579
Ottaiano A, Sabbatino F, Perri F, Cascella M, Sirica R, Patrone R, Capuozzo M, Savarese G, Ianniello M, Petrillo N, et al. KRAS p.G12C Mutation in Metastatic Colorectal Cancer: Prognostic Implications and Advancements in Targeted Therapies. Cancers. 2023; 15(14):3579. https://doi.org/10.3390/cancers15143579
Chicago/Turabian StyleOttaiano, Alessandro, Francesco Sabbatino, Francesco Perri, Marco Cascella, Roberto Sirica, Renato Patrone, Maurizio Capuozzo, Giovanni Savarese, Monica Ianniello, Nadia Petrillo, and et al. 2023. "KRAS p.G12C Mutation in Metastatic Colorectal Cancer: Prognostic Implications and Advancements in Targeted Therapies" Cancers 15, no. 14: 3579. https://doi.org/10.3390/cancers15143579