
The Association between the Gut Microbiome and Development and Progression of Cancer Treatment Adverse Effects


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Gut Microbiome
1.2. Changes in the Gut Microbiome as a Consequence of Cancer Treatment
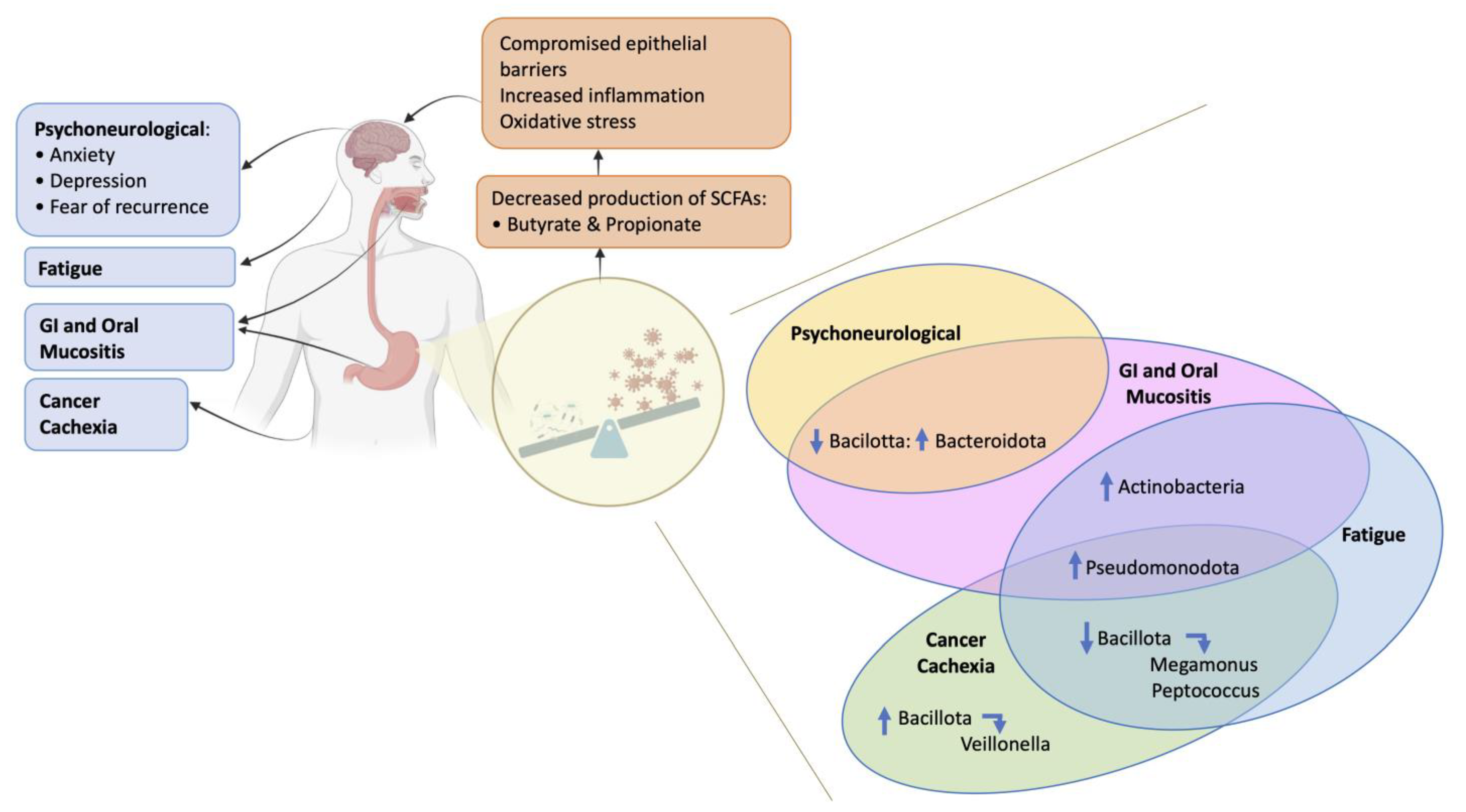
2. The Gut Microbiome in Supportive Care during and after Cancer Treatment
2.1. Gastrointestinal (GI) Mucositis Following Chemotherapy (CT)
2.2. Gastrointestinal (GI) Mucositis Following Radiotherapy (RT) or Chemoradiotherapy (CRT)
2.3. Oral Mucositis Following Cancer Treatment
3. Psychoneurological Disorders Following Cancer Treatments
4. Cancer Cachexia Following Cancer Treatments
5. Fatigue Following Cancer Treatments
6. Potential Interventions Targeting Gut Microbiome to Modulate Cancer Treatment Adverse Effects
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Simon, G.L.; Gorbach, S.L. Intestinal microflora. Med. Clin. N. Am. 1982, 66, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Stringer, A.M.; Gibson, R.J.; Bowen, J.M.; Logan, R.M.; Yeoh, A.S.; Keefe, D.M. Chemotherapy-induced mucositis: The role of gastrointestinal microflora and mucins in the luminal environment. J. Support Oncol. 2007, 5, 259–267. [Google Scholar] [PubMed]
- Gustafsson, B.E. The physiological importance of the colonic microflora. Scand. J. Gastroenterol. Suppl. 1982, 77, 117–131. [Google Scholar] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- Pistollato, F.; Sumalla Cano, S.; Elio, I.; Masias Vergara, M.; Giampieri, F.; Battino, M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr. Rev. 2016, 74, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Abu-Shanab, A.; Quigley, E.M. The role of the gut microbiota in nonalcoholic fatty liver disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Heller, F.; Duchmann, R. Intestinal flora and mucosal immune responses. Int. J. Med. Microbiol. 2003, 293, 77–86. [Google Scholar] [CrossRef]
- Zitvogel, L.; Daillère, R.; Roberti, M.P.; Routy, B.; Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Microbiol. 2017, 15, 465–478. [Google Scholar] [CrossRef]
- Ma, W.; Mao, Q.; Xia, W.; Dong, G.; Yu, C.; Jiang, F. Gut microbiota shapes the efficiency of cancer therapy. Front. Microbiol. 2019, 10, 1050. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Wu, C.-Y.; Yu, J. The role of gut microbiota in cancer treatment: Friend or foe? Gut 2020, 69, 1867–1876. [Google Scholar] [CrossRef]
- Ye, Y.; Carlsson, G.; Barr Agholme, M.; Wilson, J.A.L.; Roos, A.; Henriques-Normark, B.; Engstrand, L.; Modéer, T.; Pütsep, K. Oral bacterial community dynamics in paediatric patients with malignancies in relation to chemotherapy-related oral mucositis: A prospective study. Clin. Microbiol. Infect. 2013, 19, E559–E567. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Shen, L.; Shi, W.; Xia, F.; Zhang, H.; Wang, Y.; Zhang, J.; Wang, Y.; Sun, X.; Zhang, Z.; et al. Gut Microbiome Components Predict Response to Neoadjuvant Chemoradiotherapy in Patients with Locally Advanced Rectal Cancer: A Prospective, Longitudinal Study. Clin. Cancer Res. 2021, 27, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, D.; Andreyev, J. The Role of the Human Gut Microbiome in Inflammatory Bowel Disease and Radiation Enteropathy. Microorganisms 2022, 10, 1613. [Google Scholar] [CrossRef] [PubMed]
- Al-Qadami, G.H.; Secombe, K.R.; Subramaniam, C.B.; Wardill, H.R.; Bowen, J.M. Gut Microbiota-Derived Short-Chain Fatty Acids: Impact on Cancer Treatment Response and Toxicities. Microorganisms 2022, 10, 2048. [Google Scholar] [CrossRef]
- Li, Z.; Ke, X.; Zuo, D.; Wang, Z.; Fang, F.; Li, B. New Insights into the Relationship between Gut Microbiota and Radiotherapy for Cancer. Nutrients 2022, 15, 48. [Google Scholar] [CrossRef]
- Al-Qadami, G.; Bowen, J.; Van Sebille, Y.; Secombe, K.; Dorraki, M.; Verjans, J.; Wardill, H.; Le, H. Baseline gut microbiota composition is associated with oral mucositis and tumour recurrence in patients with head and neck cancer: A pilot study. Support. Care Cancer 2023, 31, 98. [Google Scholar] [CrossRef]
- Wang, A.; Ling, Z.; Yang, Z.; Kiela, P.R.; Wang, T.; Wang, C.; Cao, L.; Geng, F.; Shen, M.; Ran, X.; et al. Gut Microbial Dysbiosis May Predict Diarrhea and Fatigue in Patients Undergoing Pelvic Cancer Radiotherapy: A Pilot Study. PLoS ONE 2015, 10, e0126312. [Google Scholar] [CrossRef]
- González-Mercado, V.J.; Lim, J.; Marrero, S.; Pedro, E.; Saligan, L.N. Gut microbiota and fatigue in rectal cancer patients: A cross-sectional pilot study. Support. Care Cancer 2021, 29, 4615–4621. [Google Scholar] [CrossRef]
- Okubo, R.; Kinoshita, T.; Katsumata, N.; Uezono, Y.; Xiao, J.; Matsuoka, Y.J. Impact of chemotherapy on the association between fear of cancer recurrence and the gut microbiota in breast cancer survivors. Brain Behav. Immun. 2020, 85, 186–191. [Google Scholar] [CrossRef]
- Bilenduke, E.; Sterrett, J.D.; Ranby, K.W.; Borges, V.F.; Grigsby, J.; Carr, A.L.; Kilbourn, K.; Lowry, C.A. Impacts of breast cancer and chemotherapy on gut microbiome, cognitive functioning, and mood relative to healthy controls. Sci. Rep. 2022, 12, 19547. [Google Scholar] [CrossRef]
- Ubachs, J.; Ziemons, J.; Soons, Z.; Aarnoutse, R.; van Dijk, D.P.; Penders, J.; van Helvoort, A.; Smidt, M.L.; Kruitwagen, R.F.; Baade-Corpelijn, L.; et al. Gut microbiota and short-chain fatty acid alterations in cachectic cancer patients. J. Cachexia Sarcopenia Muscle 2021, 12, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Hakozaki, T.; Nolin-Lapalme, A.; Kogawa, M.; Okuma, Y.; Nakamura, S.; Moreau-Amaru, D.; Tamura, T.; Hosomi, Y.; Takeyama, H.; Richard, C.; et al. Cancer Cachexia among Patients with Advanced Non-Small-Cell Lung Cancer on Immunotherapy: An Observational Study with Exploratory Gut Microbiota Analysis. Cancers 2022, 14, 5405. [Google Scholar] [CrossRef] [PubMed]
- Stringer, A.M.; Al-Dasooqi, N.; Bowen, J.M.; Tan, T.H.; Radzuan, M.; Logan, R.M.; Mayo, B.; Keefe, D.M.K.; Gibson, R.J. Biomarkers of chemotherapy-induced diarrhoea: A clinical study of intestinal microbiome alterations, inflammation and circulating matrix metalloproteinases. Support. Care Cancer 2013, 21, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Q.; Wang, X.; Zhu, L.; Chen, J.; Zhang, B.; Chen, Y.; Yuan, Z. Gut microbial dysbiosis is associated with development and progression of radiation enteritis during pelvic radiotherapy. J. Cell. Mol. Med. 2019, 23, 3747–3756. [Google Scholar] [CrossRef]
- Reis Ferreira, M.; Andreyev, H.J.N.; Mohammed, K.; Truelove, L.; Gowan, S.M.; Li, J.; Gulliford, S.L.; Marchesi, J.R.; Dearnaley, D.P. Microbiota-and radiotherapy-induced gastrointestinal side-effects (MARS) study: A large pilot study of the microbiome in acute and late-radiation enteropathy. Clin. Cancer Res. 2019, 25, 6487–6500. [Google Scholar] [CrossRef]
- Bowen, J.; Al-Dasooqi, N.; Bossi, P.; Wardill, H.; Van Sebille, Y.; Al-Azri, A.; Bateman, E.; Correa, M.E.; Raber-Durlacher, J.; Kandwal, A.; et al. The pathogenesis of mucositis: Updated perspectives and emerging targets. Support. Care Cancer 2019, 27, 4023–4033. [Google Scholar] [CrossRef]
- Clarke, G.; Sandhu, K.V.; Griffin, B.T.; Dinan, T.G.; Cryan, J.F.; Hyland, N.P. Gut Reactions: Breaking Down Xenobiotic–Microbiome Interactions. Pharmacol. Rev. 2019, 71, 198. [Google Scholar] [CrossRef]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef]
- Bidell, M.R.; Hobbs, A.L.V.; Lodise, T.P. Gut microbiome health and dysbiosis: A clinical primer. Pharmacotherapy 2022, 42, 849–857. [Google Scholar] [CrossRef]
- Montassier, E.; Gastinne, T.; Vangay, P.; Al-Ghalith, G.A.; Bruley des Varannes, S.; Massart, S.; Moreau, P.; Potel, G.; de La Cochetière, M.F.; Batard, E.; et al. Chemotherapy-driven dysbiosis in the intestinal microbiome. Aliment. Pharmacol. Ther. 2015, 42, 515–528. [Google Scholar] [CrossRef]
- Paulsen, J.A.; Ptacek, T.S.; Carter, S.J.; Liu, N.; Kumar, R.; Hyndman, L.; Lefkowitz, E.J.; Morrow, C.D.; Rogers, L.Q. Gut microbiota composition associated with alterations in cardiorespiratory fitness and psychosocial outcomes among breast cancer survivors. Support. Care Cancer 2017, 25, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Huang, W.; Guo, H.-L.; Deng, X.; Zhu, T.-T.; Xiong, J.-F.; Xu, Y.-H.; Xu, Y. Short-chain fatty acids inhibit oxidative stress and inflammation in mesangial cells induced by high glucose and lipopolysaccharide. Exp. Clin. Endocrinol. Diabetes 2017, 125, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, C.; Yue, J. Radiotherapy and the gut microbiome: Facts and fiction. Radiat. Oncol. 2021, 16, 9. [Google Scholar] [CrossRef]
- Liu, L.; Shah, K. The potential of the gut microbiome to reshape the cancer therapy paradigm: A review. JAMA Oncol. 2022, 8, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, W.; Kang, W.; He, Y.; Yang, R.; Mou, X.; Zhao, W. Effects of microbiota on anticancer drugs: Current knowledge and potentilal applications. EBioMedicine 2022, 83, 104197. [Google Scholar] [CrossRef]
- Rajagopala, S.V.; Yooseph, S.; Harkins, D.M.; Moncera, K.J.; Zabokrtsky, K.B.; Torralba, M.G.; Tovchigrechko, A.; Highlander, S.K.; Pieper, R.; Sender, L.; et al. Gastrointestinal microbial populations can distinguish pediatric and adolescent Acute Lymphoblastic Leukemia (ALL) at the time of disease diagnosis. BMC Genom. 2016, 17, 635. [Google Scholar] [CrossRef]
- El Alam, M.B.; Sims, T.T.; Kouzy, R.; Biegert, G.W.; Jaoude, J.A.; Karpinets, T.V.; Yoshida-Court, K.; Wu, X.; Delgado-Medrano, A.Y.; Mezzari, M.P.; et al. A prospective study of the adaptive changes in the gut microbiome during standard-of-care chemoradiotherapy for gynecologic cancers. PLoS ONE 2021, 16, e0247905. [Google Scholar] [CrossRef]
- Stringer, A.M.; Logan, R.M. The role of oral flora in the development of chemotherapy-induced oral mucositis. J. Oral. Pathol. Med. 2015, 44, 81–87. [Google Scholar] [CrossRef]
- Bowen, J.M.; Gibson, R.J.; Coller, J.K.; Blijlevens, N.; Bossi, P.; Al-Dasooqi, N.; Bateman, E.H.; Chiang, K.; de Mooij, C.; Mayo, B.; et al. Systematic review of agents for the management of cancer treatment-related gastrointestinal mucositis and clinical practice guidelines. Support. Care Cancer 2019, 27, 4011–4022. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Varela, E.; Martinez, C.; Antolin, M.; Llopis, M.; Doré, J.; Giralt, J.; Guarner, F.; Malagelada, J.-R. The Gut Microbiota Predispose to the Pathophysiology of Acute Postradiotherapy Diarrhea. Am. J. Gastroenterol. 2008, 103, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Shen, L.; Zou, W.; Wang, J.; Yang, J.; Wang, Y.; Liu, B.; Xie, L.; Zhu, J.; Zhang, Z. The Gut Microbiome Is Associated With Therapeutic Responses and Toxicities of Neoadjuvant Chemoradiotherapy in Rectal Cancer Patients-A Pilot Study. Front. Cell. Infect. Microbiol. 2020, 10, 562463. [Google Scholar] [CrossRef] [PubMed]
- Aarnoutse, R.; Ziemons, J.; Hillege, L.E.; de Vos-Geelen, J.; de Boer, M.; Bisschop, S.M.; Vriens, B.; Vincent, J.; van de Wouw, A.J.; Le, G.N.; et al. Changes in intestinal microbiota in postmenopausal oestrogen receptor-positive breast cancer patients treated with (neo)adjuvant chemotherapy. NPJ Breast Cancer 2022, 8, 89. [Google Scholar] [CrossRef] [PubMed]
- Elad, S.; Cheng, K.K.; Lalla, R.V.; Yarom, N.; Hong, C.; Logan, R.M.; Bowen, J.; Gibson, R.; Saunders, D.P.; Zadik, Y.; et al. MASCC/ISOO clinical practice guidelines for the management of mucositis secondary to cancer therapy. Cancer 2020, 126, 4423–4431. [Google Scholar] [CrossRef] [PubMed]
- Napeñas, J.J.; Brennan, M.T.; Bahrani-Mougeot, F.K.; Fox, P.C.; Lockhart, P.B. Relationship between mucositis and changes in oral microflora during cancer chemotherapy. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. Endod. 2007, 103, 48–59. [Google Scholar] [CrossRef]
- Wardill, H.R.; Mander, K.A.; Van Sebille, Y.Z.A.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Sonis, S.T. Cytokine-mediated blood brain barrier disruption as a conduit for cancer/chemotherapy-associated neurotoxicity and cognitive dysfunction. Int. J. Cancer 2016, 139, 2635–2645. [Google Scholar] [CrossRef]
- Bajic, J.E.; Johnston, I.N.; Howarth, G.S.; Hutchinson, M.R. From the bottom-up: Chemotherapy and gut-brain axis dysregulation. Front. Behav. Neurosci. 2018, 12, 104. [Google Scholar] [CrossRef]
- Barrio, C.; Arias-Sánchez, S.; Martín-Monzón, I. The gut microbiota-brain axis, psychobiotics and its influence on brain and behaviour: A systematic review. Psychoneuroendocrinology 2022, 137, 105640. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Flux, M.C.; Lowry, C.A. Finding intestinal fortitude: Integrating the microbiome into a holistic view of depression mechanisms, treatment, and resilience. Neurobiol. Dis. 2020, 135, 104578. [Google Scholar] [CrossRef]
- Deleemans, J.M.; Chleilat, F.; Baydoun, M.; Piedalue, K.A.; Lowry, D.E.; Henning, J.W.; Carlson, L.E. The Chemo-Gut Pilot Study: Associations between Gastrointestinal and Psychosocial Symptoms, Gut Microbiota and Cortisol in Young Adult Cancer Survivors. Curr. Oncol. 2022, 29, 2973–2994. [Google Scholar] [CrossRef]
- Smith, K.S.; Tissier, A.; Bail, J.R.; Novak, J.R.; Morrow, C.D.; Demark-Wahnefried, W.; Fruge, A.D. Health-related quality of life is associated with fecal microbial composition in breast cancer survivors. Support. Care Cancer 2023, 31, 10. [Google Scholar] [CrossRef]
- Cao, Z.; Scott, A.M.; Hoogenraad, N.J.; Osellame, L.D. Mediators and clinical treatment for cancer cachexia: A systematic review. JCSM Rapid. Commun. 2021, 4, 166–186. [Google Scholar] [CrossRef]
- Ni, Y.; Lohinai, Z.; Heshiki, Y.; Dome, B.; Moldvay, J.; Dulka, E.; Galffy, G.; Berta, J.; Weiss, G.J.; Sommer, M.O.; et al. Distinct composition and metabolic functions of human gut microbiota are associated with cachexia in lung cancer patients. ISME J 2021, 15, 3207–3220. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, J.; Mendoza, T.; Zhang, L.; Fu, S.; Piha-Paul, S.A.; Hong, D.S.; Janku, F.; Karp, D.D.; Ballhausen, A.; Gong, J.; et al. Associations between the gut microbiome and fatigue in cancer patients. Sci. Rep. 2021, 11, 5847. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Fedirko, V.; Beitler, J.; Bai, J.; Peng, G.; Zhou, C.; Gu, J.; Zhao, H.; Lin, I.-H.; Chico, C.E.; et al. The role of the gut microbiome in cancer-related fatigue: Pilot study on epigenetic mechanisms. Support. Care Cancer 2021, 29, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xie, L.; Zhao, Y.; He, J.; Zhu, J.; Li, M.; Sun, Y. Diverse gut microbiota pattern between mild and severe cancer-related fatigue in lung cancer patients treated with first-line chemotherapy: A pilot study. Thorac. Cancer 2023, 14, 309–319. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Deleemans, J.M.; Gajtani, Z.; Baydoun, M.; Reimer, R.A.; Piedalue, K.A.; Carlson, L.E. The Use of Prebiotic and Probiotic Interventions for Treating Gastrointestinal and Psychosocial Health Symptoms in Cancer Patients and Survivors: A Systematic Review. Integr. Cancer Ther. 2021, 20, 15347354211061733. [Google Scholar] [CrossRef]
- Demers, M.; Dagnault, A.; Desjardins, J. A randomized double-blind controlled trial: Impact of probiotics on diarrhea in patients treated with pelvic radiation. Clin. Nutr. 2014, 33, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Linn, Y.H.; Thu, K.K.; Win, N.H.H. Effect of Probiotics for the Prevention of Acute Radiation-Induced Diarrhoea Among Cervical Cancer Patients: A Randomized Double-Blind Placebo-Controlled Study. Probiotics. Antimicrob. Proteins. 2019, 11, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Rosli, D.; Shahar, S.; Manaf, Z.A.; Lau, H.J.; Yusof, N.Y.M.; Haron, M.R.; Majid, H.A. Randomized Controlled Trial on the Effect of Partially Hydrolyzed Guar Gum Supplementation on Diarrhea Frequency and Gut Microbiome Count Among Pelvic Radiation Patients. JPEN J. Parenter. Enter. Nutr. 2021, 45, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, B.K.; Ramadass, B.; Viswanathan, P.N.; Samuel, P.; Gowri, M.; Pugazhendhi, S.; Ramakrishna, B.S. A phase 2 randomized controlled trial of oral resistant starch supplements in the prevention of acute radiation proctitis in patients treated for cervical cancer. J. Cancer Res. Ther. 2019, 15, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Wierdsma, N.J.; van Bodegraven, A.A.; Uitdehaag, B.M.J.; Arjaans, W.; Savelkoul, P.H.M.; Kruizenga, H.M.; van Bokhorst-de van der Schueren, M.A.E. Fructo-oligosaccharides and fibre in enteral nutrition has a beneficial influence on microbiota and gastrointestinal quality of life. Scand. J. Gastroenterol. 2009, 44, 804–812. [Google Scholar] [CrossRef]
- Ohigashi, S.; Hoshino, Y.; Ohde, S.; Onodera, H. Functional outcome, quality of life, and efficacy of probiotics in postoperative patients with colorectal cancer. Surg. Today 2011, 41, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Chu, S.-H.; Jeon, J.Y.; Lee, M.-K.; Park, J.-H.; Lee, D.-C.; Lee, J.-W.; Kim, N.-K. Effects of 12 weeks of probiotic supplementation on quality of life in colorectal cancer survivors: A double-blind, randomized, placebo-controlled trial. Dig Liver Dis. 2014, 46, 1126–1132. [Google Scholar] [CrossRef]
- Yoon, B.J.; Oh, H.-K.; Lee, J.; Cho, J.R.; Kim, M.J.; Kim, D.-W.; Kang, S.-B. Effects of probiotics on bowel function restoration following ileostomy closure in rectal cancer patients: A randomized controlled trial. Colorectal. Dis. 2021, 23, 901–910. [Google Scholar] [CrossRef]
- Juan, Z.; Chen, J.; Ding, B.; Yongping, L.; Liu, K.; Wang, L.; Le, Y.; Liao, Q.; Shi, J.; Huang, J.; et al. Probiotic supplement attenuates chemotherapy-related cognitive impairment in patients with breast cancer: A randomised, double-blind, and placebo-controlled trial. Eur. J. Cancer 2022, 161, 10–22. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Knisely, A.; Seo, Y.D.; Wargo, J.A.; Chelvanambi, M. Monitoring and Modulating Diet and Gut Microbes to Enhance Response and Reduce Toxicity to Cancer Treatment. Cancers 2023, 15, 777. [Google Scholar] [CrossRef]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut microbiota-derived short-chain fatty acids and colorectal cancer: Ready for clinical translation? Cancer Lett. 2022, 526, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-Y.; Mei, J.-X.; Yu, G.; Lei, L.; Zhang, W.-H.; Liu, K.; Chen, X.-L.; Kolat, D.; Yang, K.; Hu, J.-K. Role of the gut microbiota in anticancer therapy: From molecular mechanisms to clinical applications. Signal. Transduct. Target. Ther. 2023, 8, 201. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| GI Mucositis Following CT | |||
|---|---|---|---|
| Participants | Study Protocol | Study Outcomes | Reference |
Group 1: mixed Ca
|
|
| Stringer et al., 2013 [23] |
| n = 28 (18 M, 10 F), non-Hodgkin’s lymphoma |
|
| Montassier et al., 2015 [30] |
Total n = 44 F, breast Ca
|
|
| Aarnoutse et al., 2022 [44] |
| GI mucositis following RT | |||
|
|
| Manichanh et al., 2008 [42] |
|
|
| Wang et al., 2015 [17] |
Total n = 18 F, cervical Ca, RT
|
|
| Wang et al., 2019 [24] |
Total n = 134 M, prostate Ca with RE and without RE
|
|
| Reis Ferreira et al., 2019 [25] |
| GI mucositis following CRT | |||
Total CRT n = 22 (6 M, 16 F), rectal Ca
|
|
| Shi et al., 2020 [43] |
| Oral mucositis following CT | |||
Total n = 37 CT, mixed paediatric Ca
|
|
| Ye et al., 2013 [11] |
| Oral mucositis following RT or CRT | |||
Total n = 17 (13 M, 4 F), head/neck Ca
|
|
| Al-Qadami et al., 2023 [16] |
| Participants | Study Protocol | Study Outcomes | Reference |
|---|---|---|---|
Total n = 12 F, breast Ca
|
|
| Paulsen et al., 2017 [31] * |
Total n = 126 (1 M, 125 F), breast Ca
|
|
| Okubo et al., 2020 [19] |
Total n = 35
|
|
| Deleemans et al., 2022 [52] * |
Total n = 35 F, breast Ca
|
|
| Bilenduke et al., 2022 [20] |
Total n = 70 F, breast Ca
|
|
| Smith et al., 2023 [53] |
| Participants | Study Protocol | Study Outcomes | Reference |
|---|---|---|---|
Total n = 31 (19 M, 12 F), lung Ca
|
|
| Ni et al., 2021 [55] |
Total n = 183 (80 M, 103 F)
|
|
| Ubachs et al., 2021 [21] |
Total n = 113 (72 M, 41 F), lung Ca
|
|
| Hakozaki et al., 2022 [22] |
| Participants | Study Protocol | Study Outcomes | Reference |
|---|---|---|---|
Total n = 12 F, breast Ca
|
|
| Paulsen et al., 2017 [31] * |
Total n = 88 (43 M, 45 F), mixed Ca
|
|
| Hajjar et al., 2021 [56] |
Total n = 13 (11 M, 2 F), head/neck Ca
|
|
| Xiao et al., 2021 [57] |
Total n = 50 (28 M, 22 F), rectal Ca
|
|
| González-Mercado et al., 2021 [18] |
Total n = 35
|
|
| Deleemans et al., 2022 [52] * |
Total n = 20 (15 M, 5 F), lung Ca, CT
|
|
| Wei et al., 2023 [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maddern, A.S.; Coller, J.K.; Bowen, J.M.; Gibson, R.J. The Association between the Gut Microbiome and Development and Progression of Cancer Treatment Adverse Effects. Cancers 2023, 15, 4301. https://doi.org/10.3390/cancers15174301
Maddern AS, Coller JK, Bowen JM, Gibson RJ. The Association between the Gut Microbiome and Development and Progression of Cancer Treatment Adverse Effects. Cancers. 2023; 15(17):4301. https://doi.org/10.3390/cancers15174301
Chicago/Turabian StyleMaddern, Amanda S., Janet K. Coller, Joanne M. Bowen, and Rachel J. Gibson. 2023. "The Association between the Gut Microbiome and Development and Progression of Cancer Treatment Adverse Effects" Cancers 15, no. 17: 4301. https://doi.org/10.3390/cancers15174301