
High Tumoral STMN1 Expression Is Associated with Malignant Potential and Poor Prognosis in Patients with Neuroblastoma

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Immunohistochemistry
2.3. Evaluation of Immunostaining
2.4. siRNA Transfection
2.5. Establishment of MYCN-Overexpressing NB Cell Lines
2.6. Western Blot Analysis
2.7. Cell Proliferation Assay
2.8. Statistical Analysis
3. Results
3.1. Immunohistochemical Staining of STMN1 in Clinical NB Tissues
3.2. Association of STMN1 Expression with the Clinicopathological Features of Clinical Neuroblastoma Patients
3.3. Prognostic Significance of STMN1 Expression in NB Patients with and without MYCN Amplification
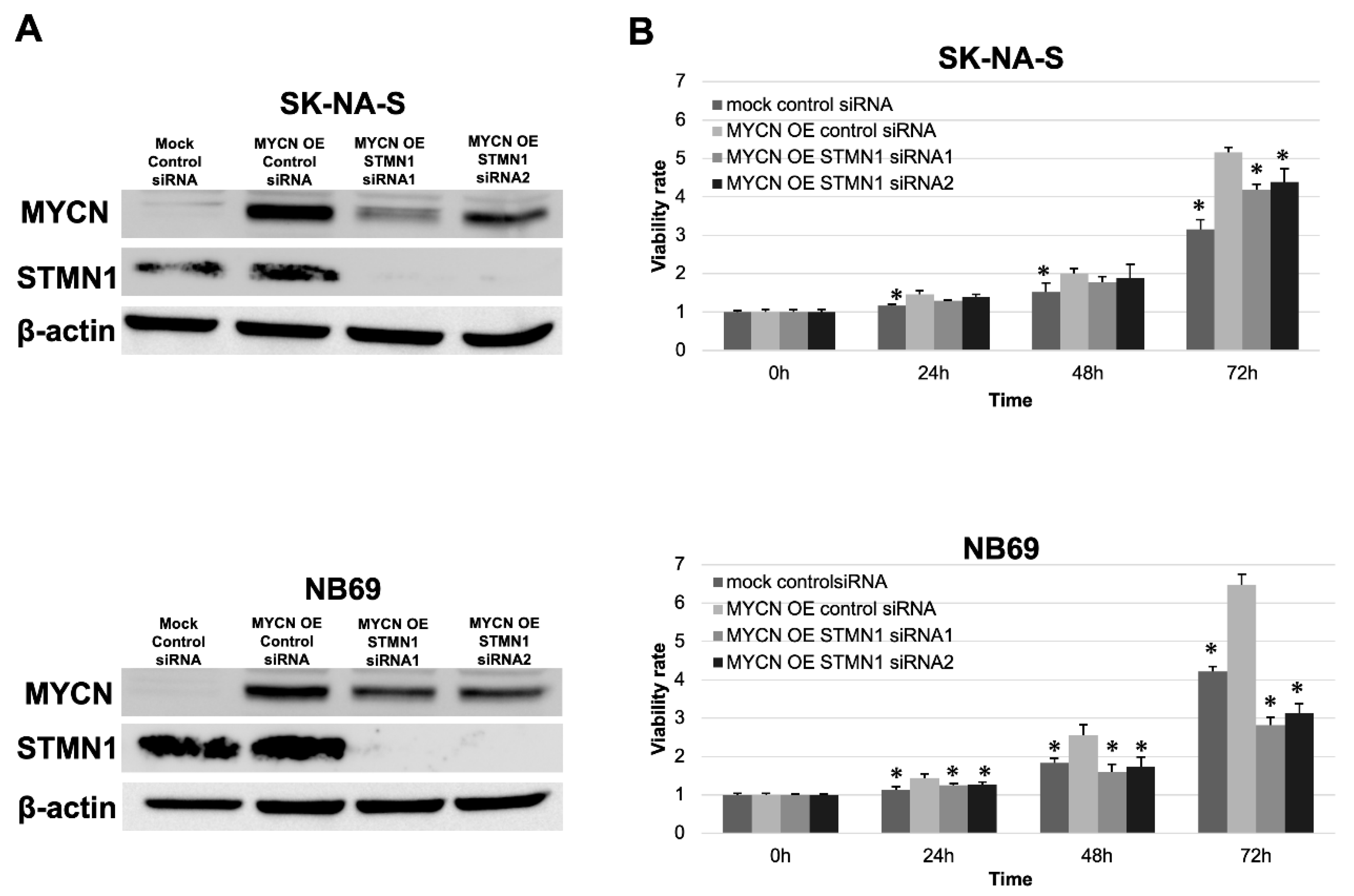
3.4. STMN1 Suppression Inhibited Cellular Viability in NB Cell Lines with and without MYCN Amplification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakagawara, A.; Li, Y.; Izumi, H.; Muramori, K.; Inada, H.; Nishi, M. Neuroblastoma. Jpn. J. Clin. Oncol. 2018, 48, 214–241. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Boterberg, T.; Lucas, J.; Panoff, J.; Valteau-Couanet, D.; Hero, B.; Bagatell, R.; Hill-Kayser, C.E. Neuroblastoma. Pediatr. Blood Cancer 2021, 68 (Suppl. S2), e28473. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.; Chan, C.M. Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules 2022, 12, 358. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A.; Abdessalam, S.; Aldrink, J.H.; Austin, M.; Heaton, T.E.; Bruny, J.; Ehrlich, P.; Dasgupta, R.; Baertschiger, R.M.; Lautz, T.B.; et al. Update on neuroblastoma. J. Pediatr. Surg. 2019, 54, 383–389. [Google Scholar] [CrossRef]
- Cohn, S.L.; Pearson, A.D.; London, W.B.; Monclair, T.; Ambros, P.F.; Brodeur, G.M.; Faldum, A.; Hero, B.; Iehara, T.; Machin, D.; et al. The International Neuroblastoma Risk Group (INRG) classification system: An INRG Task Force report. J. Clin. Oncol. 2009, 27, 289–297. [Google Scholar] [CrossRef]
- Andersen, S.S. Balanced regulation of microtubule dynamics during the cell cycle: A contemporary view. Bioessays 1999, 21, 53–60. [Google Scholar] [CrossRef]
- Wang, W.; Wang, M.; Yang, M.; Zeng, B.; Qiu, W.; Ma, Q.; Jing, X.; Zhang, Q.; Wang, B.; Yin, C.; et al. Transcriptome dynamics of hippocampal neurogenesis in macaques across the lifespan and aged humans. Cell Res. 2022, 32, 729–743. [Google Scholar] [CrossRef]
- Takekoshi, K.; Nomura, F.; Isobe, K.; Motooka, M.; Nammoku, T.; Nakai, T. Identification and initial characterization of stathmin by the differential display method in nerve growth factor-treated PC12 cells. Eur. J. Endocrinol. 1998, 138, 707–712. [Google Scholar] [CrossRef]
- Rana, S.; Maples, P.B.; Senzer, N.; Nemunaitis, J. Stathmin 1: A novel therapeutic target for anticancer activity. Expert Rev. Anticancer Ther. 2008, 8, 1461–1470. [Google Scholar] [CrossRef]
- Zhang, D.; Dai, L.; Yang, Z.; Wang, X.; LanNing, Y. Association of STMN1 with survival in solid tumors: A systematic review and meta-analysis. Int. J. Biol. Markers 2019, 34, 108–116. [Google Scholar] [CrossRef]
- Suzuki, S.; Yokobori, T.; Altan, B.; Hara, K.; Ozawa, D.; Tanaka, N.; Sakai, M.; Sano, A.; Sohda, M.; Bao, H.; et al. High stathmin 1 expression is associated with poor prognosis and chemoradiation resistance in esophageal squamous cell carcinoma. Int. J. Oncol. 2017, 50, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Yokobori, T.; Altan, B.; Ide, M.; Mochiki, E.; Yanai, M.; Kimura, A.; Kogure, N.; Yanoma, T.; Suzuki, M.; et al. High STMN1 level is associated with chemo-resistance and poor prognosis in gastric cancer patients. Br. J. Cancer 2017, 116, 1177–1185. [Google Scholar] [CrossRef]
- Suzuki, K.; Watanabe, A.; Araki, K.; Yokobori, T.; Harimoto, N.; Gantumur, D.; Hagiwara, K.; Yamanaka, T.; Ishii, N.; Tsukagoshi, M.; et al. High STMN1 Expression Is Associated with Tumor Differentiation and Metastasis in Clinical Patients with Pancreatic Cancer. Anticancer Res. 2018, 38, 939–944. [Google Scholar] [CrossRef]
- Watanabe, A.; Suzuki, H.; Yokobori, T.; Tsukagoshi, M.; Altan, B.; Kubo, N.; Suzuki, S.; Araki, K.; Wada, S.; Kashiwabara, K.; et al. Stathmin1 regulates p27 expression, proliferation and drug resistance, resulting in poor clinical prognosis in cholangiocarcinoma. Cancer Sci. 2014, 105, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, S.; Horiguchi, J.; Higuchi, T.; Katayama, A.; Handa, T.; Altan, B.; Bai, T.; Bao, P.; Bao, H.; Yokobori, T.; et al. Stathmin1 expression is associated with aggressive phenotypes and cancer stem cell marker expression in breast cancer patients. Int. J. Oncol. 2017, 51, 781–790. [Google Scholar] [CrossRef]
- Bao, P.; Yokobori, T.; Altan, B.; Iijima, M.; Azuma, Y.; Onozato, R.; Yajima, T.; Watanabe, A.; Mogi, A.; Shimizu, K.; et al. High STMN1 Expression is Associated with Cancer Progression and Chemo-Resistance in Lung Squamous Cell Carcinoma. Ann. Surg. Oncol. 2017, 24, 4017–4024. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ohtaki, Y.; Altan, B.; Yokobori, T.; Nagashima, T.; Arai, M.; Mogi, A.; Kuwano, H. Prognostic impact of stathmin 1 expression in patients with lung adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2017, 154, 1406–1417.e1403. [Google Scholar] [CrossRef]
- Don, S.; Verrills, N.M.; Liaw, T.Y.; Liu, M.L.; Norris, M.D.; Haber, M.; Kavallaris, M. Neuronal-associated microtubule proteins class III beta-tubulin and MAP2c in neuroblastoma: Role in resistance to microtubule-targeted drugs. Mol. Cancer Ther. 2004, 3, 1137–1146. [Google Scholar] [CrossRef]
- Po’uha, S.T.; Le Grand, M.; Brandl, M.B.; Gifford, A.J.; Goodall, G.J.; Khew-Goodall, Y.; Kavallaris, M. Stathmin levels alter PTPN14 expression and impact neuroblastoma cell migration. Br. J. Cancer 2020, 122, 434–444. [Google Scholar] [CrossRef]
- Byrne, F.L.; Yang, L.; Phillips, P.A.; Hansford, L.M.; Fletcher, J.I.; Ormandy, C.J.; McCarroll, J.A.; Kavallaris, M. RNAi-mediated stathmin suppression reduces lung metastasis in an orthotopic neuroblastoma mouse model. Oncogene 2014, 33, 882–890. [Google Scholar] [CrossRef]
- Hailat, N.; Strahler, J.; Melhem, R.; Zhu, X.X.; Brodeur, G.; Seeger, R.C.; Reynolds, C.P.; Hanash, S. N-myc gene amplification in neuroblastoma is associated with altered phosphorylation of a proliferation related polypeptide (Op18). Oncogene 1990, 5, 1615–1618. [Google Scholar]
- Hsu, H.P.; Li, C.F.; Lee, S.W.; Wu, W.R.; Chen, T.J.; Chang, K.Y.; Liang, S.S.; Tsai, C.J.; Shiue, Y.L. Overexpression of stathmin 1 confers an independent prognostic indicator in nasopharyngeal carcinoma. Tumour Biol. 2014, 35, 2619–2629. [Google Scholar] [CrossRef] [PubMed]
- San-Marina, S.; Han, Y.; Liu, J.; Minden, M.D. Suspected leukemia oncoproteins CREB1 and LYL1 regulate Op18/STMN1 expression. Biochim. Biophys. Acta 2012, 1819, 1164–1172. [Google Scholar] [CrossRef] [PubMed]
- Masciarelli, S.; Fontemaggi, G.; Di Agostino, S.; Donzelli, S.; Carcarino, E.; Strano, S.; Blandino, G. Gain-of-function mutant p53 downregulates miR-223 contributing to chemoresistance of cultured tumor cells. Oncogene 2014, 33, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Xun, G.; Hu, W.; Li, B. PTEN loss promotes oncogenic function of STMN1 via PI3K/AKT pathway in lung cancer. Sci. Rep. 2021, 11, 14318. [Google Scholar] [CrossRef]
- Shi, D.; Zhang, Z.; Kong, C. CARMA3 Transcriptional Regulation of STMN1 by NF-kappaB Promotes Renal Cell Carcinoma Proliferation and Invasion. Technol. Cancer Res. Treat. 2021, 20, 15330338211027915. [Google Scholar] [CrossRef]
- Chung, M.K.; Kim, H.J.; Lee, Y.S.; Han, M.E.; Yoon, S.; Baek, S.Y.; Kim, B.S.; Kim, J.B.; Oh, S.O. Hedgehog signaling regulates proliferation of prostate cancer cells via stathmin1. Clin. Exp. Med. 2010, 10, 51–57. [Google Scholar] [CrossRef]
- Liao, L.; Zhang, Y.L.; Deng, L.; Chen, C.; Ma, X.Y.; Andriani, L.; Yang, S.Y.; Hu, S.Y.; Zhang, F.L.; Shao, Z.M.; et al. Protein Phosphatase 1 Subunit PPP1R14B Stabilizes STMN1 to Promote Progression and Paclitaxel Resistance in Triple-Negative Breast Cancer. Cancer Res. 2023, 83, 471–484. [Google Scholar] [CrossRef]
- Zheng, F.; Liao, Y.J.; Cai, M.Y.; Liu, T.H.; Chen, S.P.; Wu, P.H.; Wu, L.; Bian, X.W.; Guan, X.Y.; Zeng, Y.X.; et al. Systemic delivery of microRNA-101 potently inhibits hepatocellular carcinoma in vivo by repressing multiple targets. PLoS Genet. 2015, 11, e1004873. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.; Jia, L.T.; Hu, S.J.; Zhao, J.; Yang, J.D.; Wen, W.H.; Wang, Z.; Wang, T.; Zhao, J.; et al. c-Myc-mediated epigenetic silencing of MicroRNA-101 contributes to dysregulation of multiple pathways in hepatocellular carcinoma. Hepatology 2014, 59, 1850–1863. [Google Scholar] [CrossRef]
- Chen, J.; Abi-Daoud, M.; Wang, A.; Yang, X.; Zhang, X.; Feilotter, H.E.; Tron, V.A. Stathmin 1 is a potential novel oncogene in melanoma. Oncogene 2013, 32, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, Y.; Sang, Y.; Song, X.; Zhang, H.; Liu, Y.; Jiang, L.; Yang, Q. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Janoueix-Lerosey, I.; Novikov, E.; Monteiro, M.; Gruel, N.; Schleiermacher, G.; Loriod, B.; Nguyen, C.; Delattre, O. Gene expression profiling of 1p35-36 genes in neuroblastoma. Oncogene 2004, 23, 5912–5922. [Google Scholar] [CrossRef]
- Caron, H.; van Sluis, P.; de Kraker, J.; Bokkerink, J.; Egeler, M.; Laureys, G.; Slater, R.; Westerveld, A.; Voute, P.A.; Versteeg, R. Allelic loss of chromosome 1p as a predictor of unfavorable outcome in patients with neuroblastoma. N. Engl. J. Med. 1996, 334, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M.; Weiss, M.J.; Guo, C.; Gerbing, R.B.; Stram, D.O.; White, P.S.; Hogarty, M.D.; Sulman, E.P.; Thompson, P.M.; Lukens, J.N.; et al. Loss of heterozygosity at 1p36 independently predicts for disease progression but not decreased overall survival probability in neuroblastoma patients: A Children’s Cancer Group study. J. Clin. Oncol. 2000, 18, 1888–1899. [Google Scholar] [CrossRef]
- Jiang, W.; Huang, S.; Song, L.; Wang, Z. STMN1, a prognostic predictor of esophageal squamous cell carcinoma, is a marker of the activation of the PI3K pathway. Oncol. Rep. 2018, 39, 834–842. [Google Scholar] [CrossRef]
- Allen, M.M.; Douds, J.J.; Liang, S.X.; Desouki, M.M.; Parkash, V.; Fadare, O. An immunohistochemical analysis of stathmin 1 expression in uterine smooth muscle tumors: Differential expression in leiomyosarcomas and leiomyomas. Int. J. Clin. Exp. Pathol. 2015, 8, 2795–2801. [Google Scholar]
- Shu, F.; Zou, X.; Tuo, H.; She, S.; Huang, J.; Ren, H.; Hu, H.; Peng, S.; Wang, J.; Yang, Y. Stathmin gene silencing suppresses proliferation, migration and invasion of gastric cancer cells via AKT/sCLU and STAT3 signaling. Int. J. Oncol. 2019, 54, 1086–1098. [Google Scholar] [CrossRef]
- Yoshie, M.; Miyajima, E.; Kyo, S.; Tamura, K. Stathmin, a microtubule regulatory protein, is associated with hypoxia-inducible factor-1alpha levels in human endometrial and endothelial cells. Endocrinology 2009, 150, 2413–2418. [Google Scholar] [CrossRef]
- Elhasasna, H.; Khan, R.; Bhanumathy, K.K.; Vizeacoumar, F.S.; Walke, P.; Bautista, M.; Dahiya, D.K.; Maranda, V.; Patel, H.; Balagopal, A.; et al. A Drug Repurposing Screen Identifies Fludarabine Phosphate as a Potential Therapeutic Agent for N-MYC Overexpressing Neuroendocrine Prostate Cancers. Cells 2022, 11, 2246. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Total NB Cohort (n = 81) | p Value | MYCN Not-Amplified (n = 69) | p Value | MYCN Amplified (n = 8) | p Value | |||
|---|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | Low | High | ||||
| n = 62 | n = 19 | n = 54 | n = 15 | n = 5 | n = 3 | ||||
| Gender | |||||||||
| Male | 27 | 10 | 0.6 | 23 | 8 | 0.561 | 3 | 1 | 1 |
| Female | 35 | 9 | 31 | 7 | 2 | 2 | |||
| Age, month | |||||||||
| <18 m | 56 | 11 | 0.002 * | 52 | 10 | 0.004 * | 1 | 1 | 1 |
| 18 m≤ | 6 | 8 | 2 | 5 | 4 | 2 | |||
| Distant metastasis | |||||||||
| Absent | 26 | 4 | 0.113 | 26 | 4 | 0.156 | 0 | 0 | - |
| Present | 36 | 15 | 28 | 11 | 5 | 3 | |||
| INRGSS | |||||||||
| L1, L2, MS | 50 | 7 | 0.001 * | 46 | 7 | 0.004 * | 1 | 0 | 1 |
| M | 12 | 12 | 8 | 8 | 4 | 3 | |||
| Radical resection | |||||||||
| No | 21 | 16 | <0.001 * | 16 | 12 | <0.001 * | 5 | 3 | - |
| Yes | 41 | 3 | 38 | 3 | 0 | 0 | |||
| INPC | |||||||||
| Favorable | 52 | 7 | <0.001 * | 48 | 7 | 0.001 * | 1 | 0 | 1 |
| Unfavorable | 10 | 12 | 6 | 8 | 4 | 3 | |||
| MYCN status | |||||||||
| Not amplified | 54 | 15 | 0.38 | ||||||
| Amplified | 5 | 3 | |||||||
| INRG risk group | |||||||||
| Not high | 55 | 12 | 0.016 * | 52 | 11 | 0.017 * | 0 | 0 | - |
| High | 7 | 7 | 2 | 4 | 5 | 3 | |||
| Chemotheraphy | |||||||||
| No | 23 | 3 | 0.098 | 23 | 3 | 0.139 | 0 | 0 | - |
| Yes | 39 | 16 | 31 | 12 | 5 | 3 | |||
| Ki67 expression | |||||||||
| Low | 42 | 6 | 0.007 * | 37 | 5 | 0.018 * | 4 | 1 | 0.464 |
| High | 20 | 13 | 17 | 10 | 1 | 2 | |||
| Factors | Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|---|
| Hazard Ratio [95% Confidence Interval] | p Value | Hazard Ratio [95% Confidence Interval] | p Value | ||
| Gender | Female | 1 | 0.196 | 1 | 0.015 * |
| Male | 2.247 [0.657–7.678] | 21.56 [1.814–256.2] | |||
| Month at diagnosis | <18 | 1 | <0.001 * | 1 | 0.158 |
| ≥18 | 17.92 [4.689–68.51] | 0.074 [0.001–2.766] | |||
| Distant metastasis | Absent | 1 | 0.999 | 1 | 0.999 |
| Present | 3.06 × 109 | 4.63 × 109 | |||
| INRGSS | L1, L2, MS | 1 | 0.013 * | 1 | 0.562 |
| M | 29.26 [3.738–229] | 0.372 [0.013–10.5] | |||
| Radical resection | No | 1 | 0.999 | 1 | 0.999 |
| Yes | 1.53 × 10−10 | 6.06 × 10−9 | |||
| INPC | FH | 1 | 0.006 * | 1 | 0.46 |
| UH | 14.51 [3.12–67.45] | 0.31 [0.014–6.9] | |||
| MYCN status | Not amplified | 1 | 0.0029 * | 1 | 0.711 |
| Amplified | 6.937 [1.937–24.83] | 0.714 [0.12–4.242] | |||
| INRG risk group | Not high | 1 | <0.001 * | 1 | 0.031 * |
| High | 18.19 [4.757–69.62] | 199 [1.603–24726] | |||
| Chemotherapy | No | 1 | 0.999 | 1 | 1 |
| Yes | 2.62 × 109 | 0.477 | |||
| Complication | Absent | 1 | 0.006 * | 1 | 0.022 * |
| Present | 5.24 [1.595–17.21] | 9.985 [1.383–72.04] | |||
| Ki67 expression | Low | 1 | 0.046 * | 1 | 0.035 * |
| High | 3.491 [1.021–11.936] | 9.402 [1.167–75.74] | |||
| STMN1 expression | Low | 1 | 0.003 * | 1 | 0.019 * |
| High | 6.439 [1.882–22.03] | 16.95 [1.567–183.4] | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogushi, K.; Yokobori, T.; Nobusawa, S.; Shirakura, T.; Hirato, J.; Erkhem-Ochir, B.; Okami, H.; Dorjkhorloo, G.; Nishi, A.; Suzuki, M.; et al. High Tumoral STMN1 Expression Is Associated with Malignant Potential and Poor Prognosis in Patients with Neuroblastoma. Cancers 2023, 15, 4482. https://doi.org/10.3390/cancers15184482
Ogushi K, Yokobori T, Nobusawa S, Shirakura T, Hirato J, Erkhem-Ochir B, Okami H, Dorjkhorloo G, Nishi A, Suzuki M, et al. High Tumoral STMN1 Expression Is Associated with Malignant Potential and Poor Prognosis in Patients with Neuroblastoma. Cancers. 2023; 15(18):4482. https://doi.org/10.3390/cancers15184482
Chicago/Turabian StyleOgushi, Kenjiro, Takehiko Yokobori, Sumihito Nobusawa, Takahiro Shirakura, Junko Hirato, Bilguun Erkhem-Ochir, Haruka Okami, Gendensuren Dorjkhorloo, Akira Nishi, Makoto Suzuki, and et al. 2023. "High Tumoral STMN1 Expression Is Associated with Malignant Potential and Poor Prognosis in Patients with Neuroblastoma" Cancers 15, no. 18: 4482. https://doi.org/10.3390/cancers15184482
APA StyleOgushi, K., Yokobori, T., Nobusawa, S., Shirakura, T., Hirato, J., Erkhem-Ochir, B., Okami, H., Dorjkhorloo, G., Nishi, A., Suzuki, M., Otake, S., Saeki, H., & Shirabe, K. (2023). High Tumoral STMN1 Expression Is Associated with Malignant Potential and Poor Prognosis in Patients with Neuroblastoma. Cancers, 15(18), 4482. https://doi.org/10.3390/cancers15184482