
Long Non-Coding RNA LOC339059 Attenuates IL-6/STAT3-Signaling-Mediated PDL1 Expression and Macrophage M2 Polarization by Interacting with c-Myc in Gastric Cancer

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Established Cancer and Isogenic Cell Lines
2.2. Clinical Specimens
2.3. Total RNA Extraction, Nuclear and Cytoplasmic RNA Fractionation, Real-Time qPCR, and RNA Sequencing Analysis
2.4. RNA In Situ Hybridization (ISH)
2.5. Cell Viability
2.6. Plate Colony Formation Assay
2.7. Transwell Migration and Invasion Assay
2.8. In Vivo Tumor Growth and Metastasis
2.9. Flow Cytometer Analysis
2.10. Immunoblotting
2.11. RNA Pull-Down, RNA-Binding Protein Immunoprecipitation (RIP), and Chromatin Immunoprecipitation (ChIP) Assay
2.12. Promoter-Driven Reporter Gene Assay
2.13. Statistical Analysis
3. Results
3.1. LOC339059 Is Down-Regulated in Gastric Cancer, and Low Expression Predicts Poor Survival
3.2. LOC339059 Is Associated with the Expression of Immune Genes and Macrophage Biomarkers
3.3. Low LOC339059 Is Associated with Poor Differentiation of GC
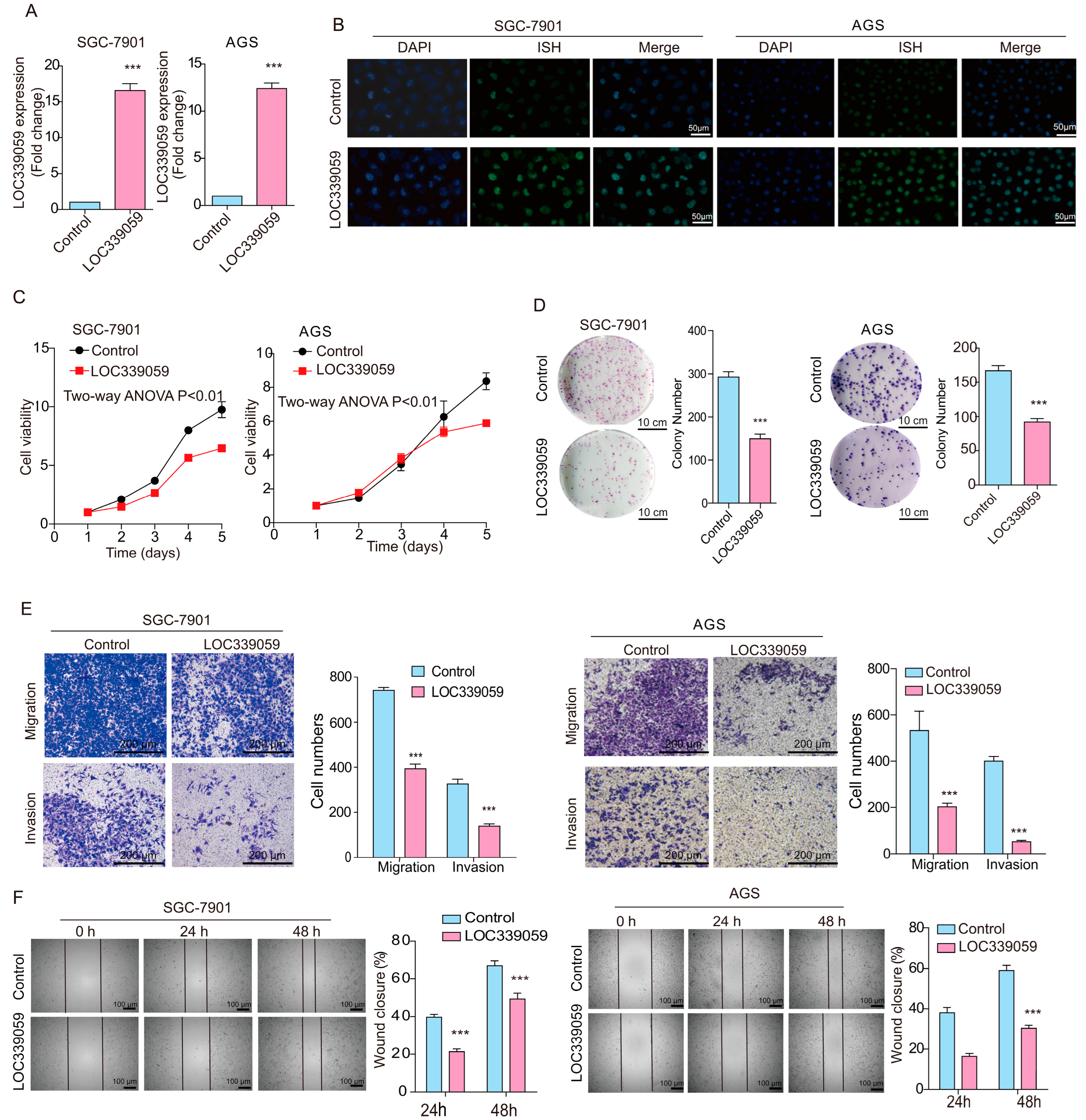
3.4. LOC339059 Suppresses Malignant Phenotypes in GC Cells
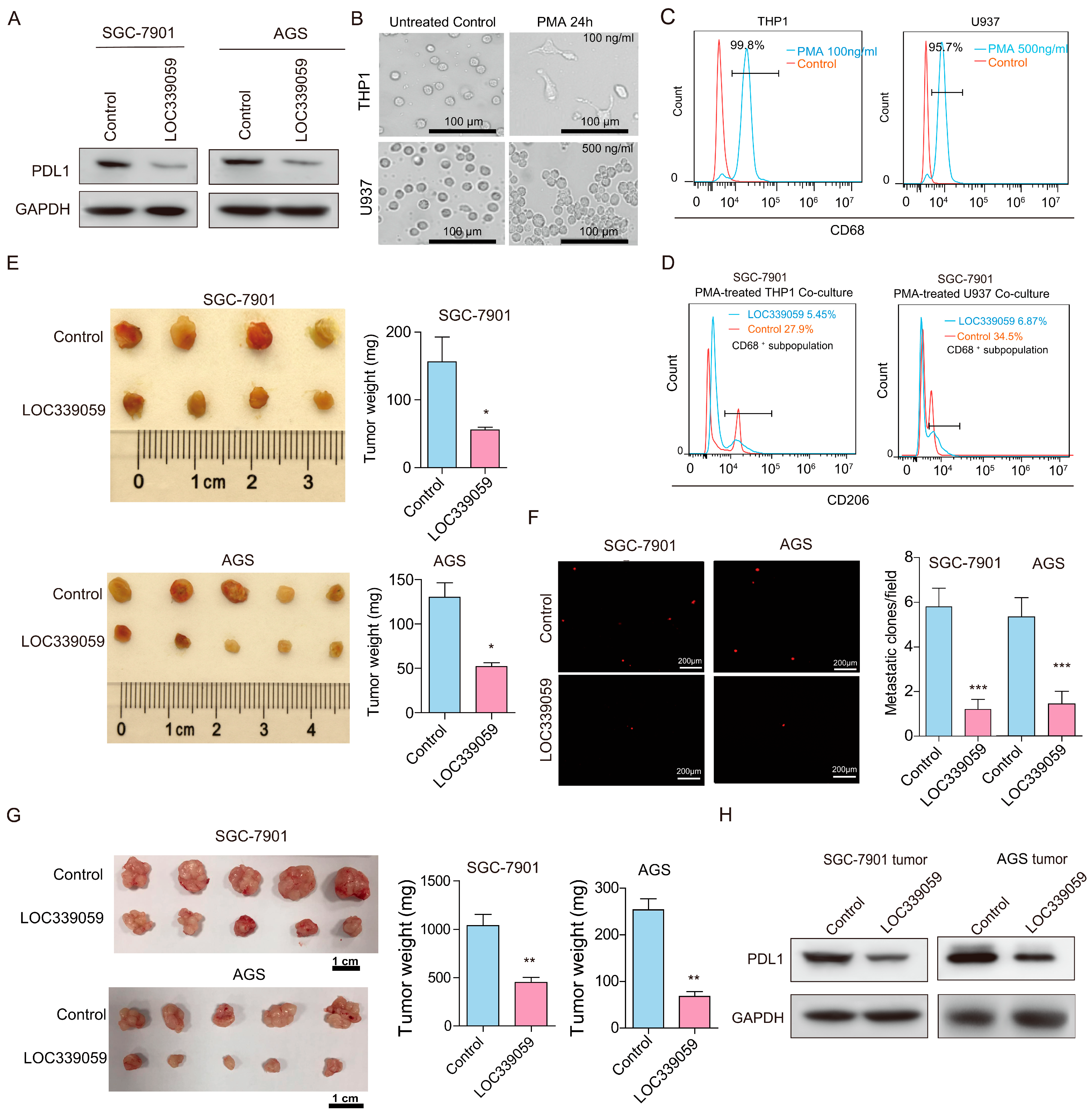
3.5. LOC339059 Inhibits PDL1 and M2 Macrophage Polarization
3.6. LOC339059 Inhibits Tumor Growth, PDL1 Expression, and Lung Metastasis In Vivo
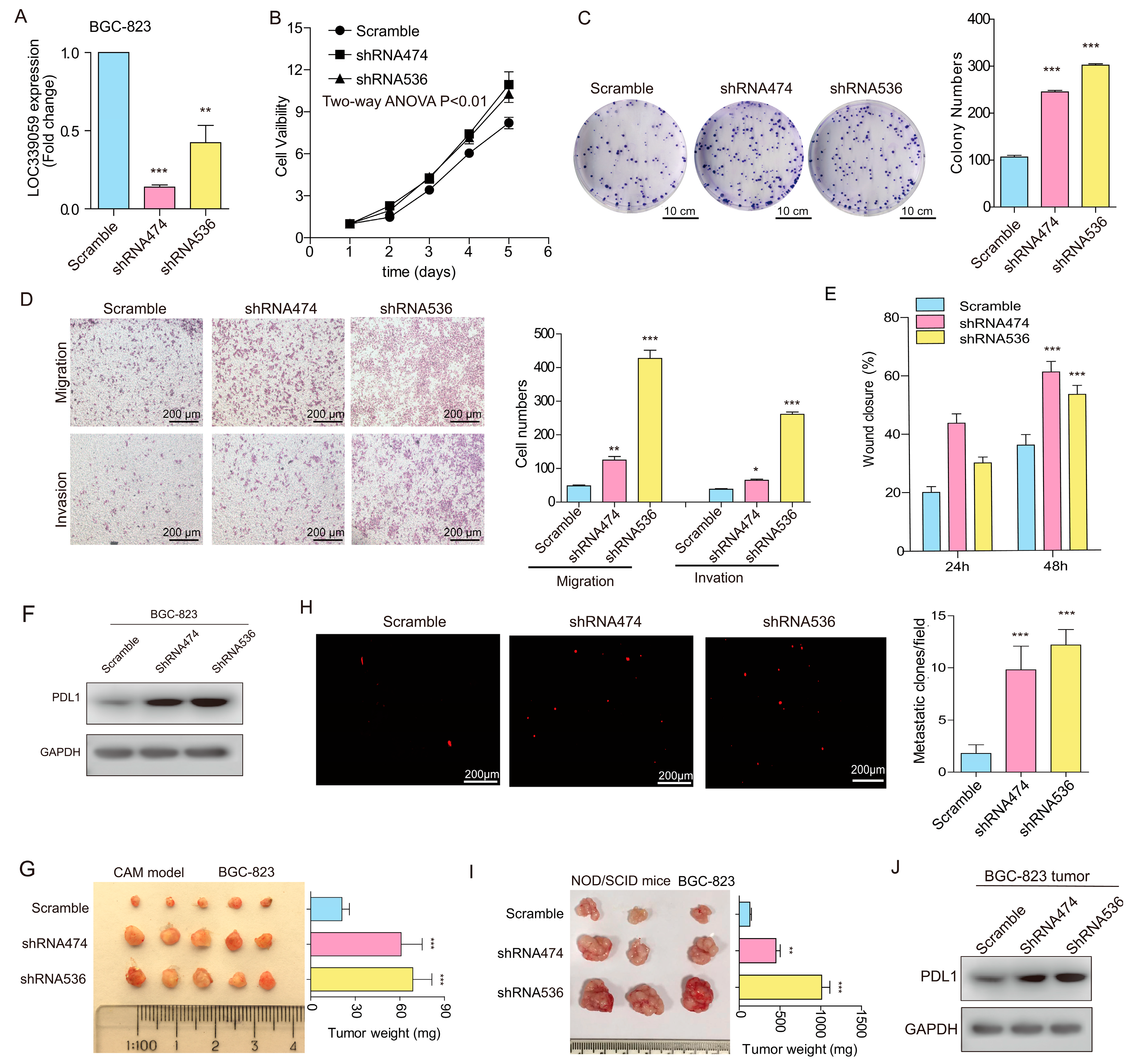
3.7. Interference with LOC339059 Enhanced Malignant Phenotypes, PDL1 Expression, M2 Macrophage Polarization, Tumor Growth, and Metastasis
3.8. High Expression of LOC339059 Decreases Activity of IL-6/STAT3 Signaling Axis
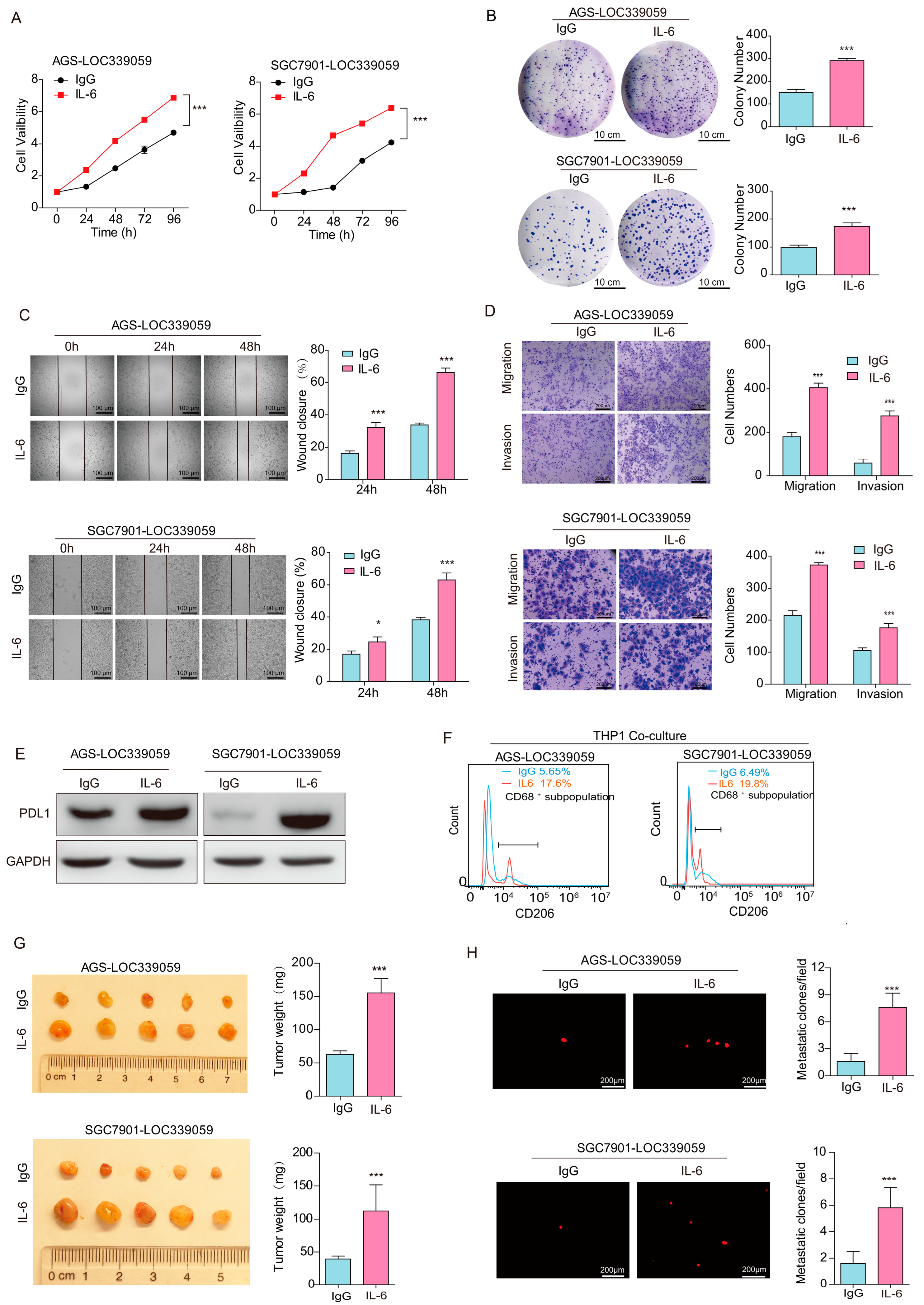
3.9. IL-6 Rescue Partly Reversed the Inhibitory Effect of LOC339059 on the Malignant Phenotype, PDL1 Expression, and M2 Macrophage Polarization
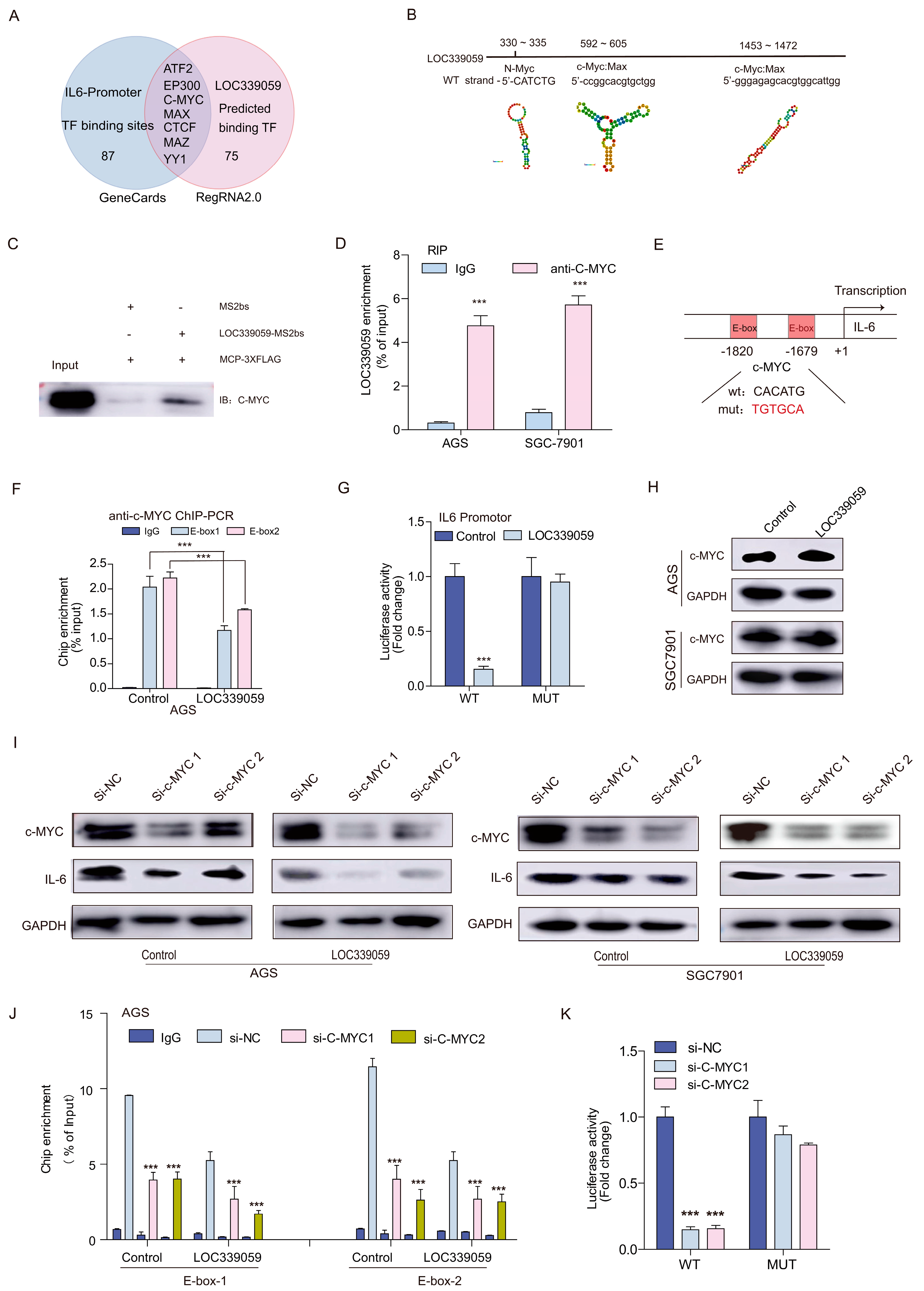
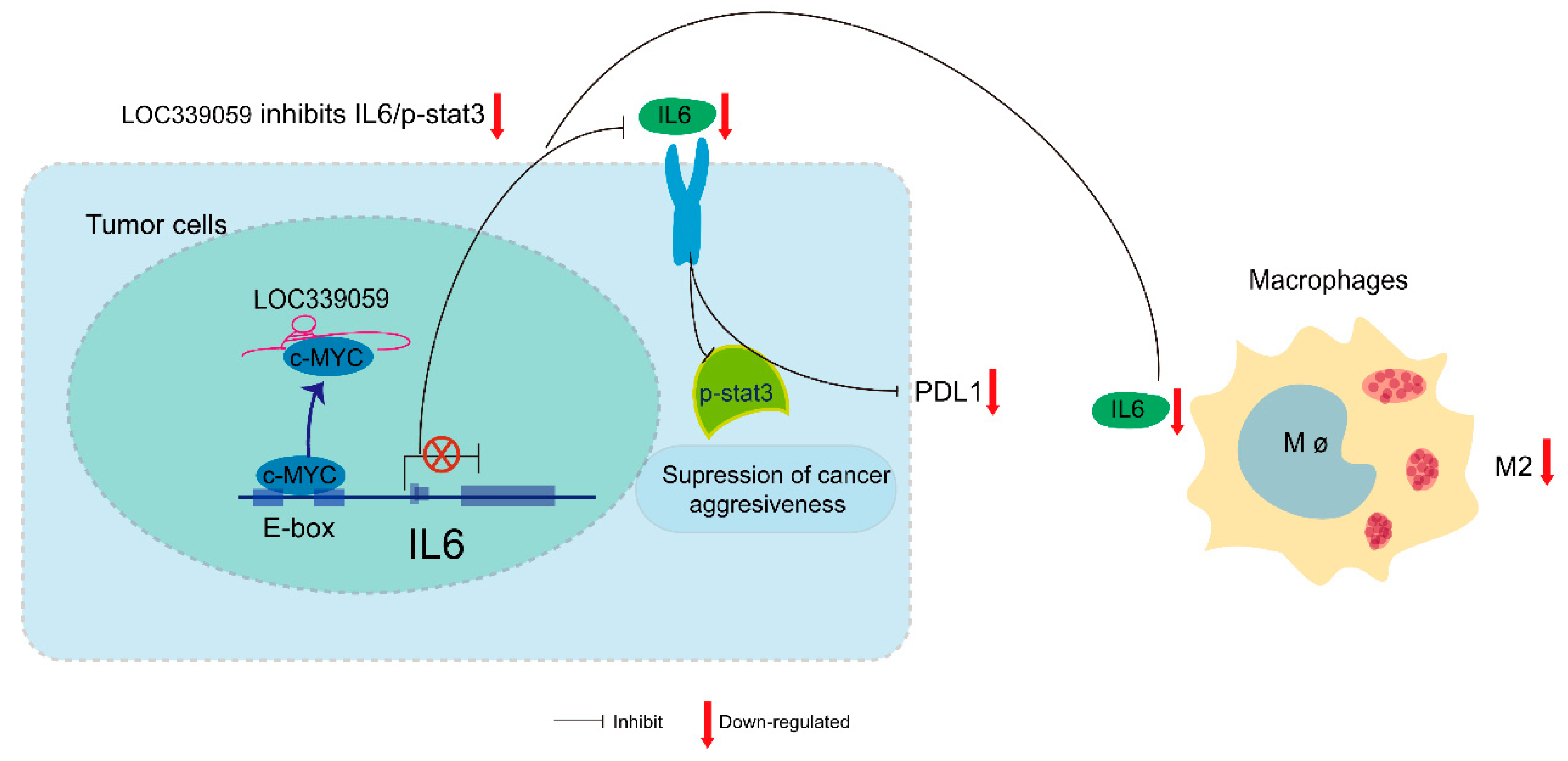
3.10. LOC339059 Competitively Interacted with c-Myc, Resulting in Reduced Transcriptional Activation of IL-6
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.M.; Zong, Y.N.; Cao, S.M.; Xu, R.H. Current cancer situation in China: Good or bad news from the 2018 Global Cancer Statistics? Cancer Commun. 2019, 39, 22. [Google Scholar] [CrossRef] [PubMed]
- Zong, L.; Abe, M.; Seto, Y.; Ji, J. The challenge of screening for early gastric cancer in China. Lancet 2016, 388, 2606. [Google Scholar] [CrossRef] [PubMed]
- Christofides, A.; Strauss, L.; Yeo, A.; Cao, C.; Charest, A.; Boussiotis, V.A. The complex role of tumor-infiltrating macrophages. Nat. Immunol. 2022, 23, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef]
- Duan, Z.; Luo, Y. Targeting macrophages in cancer immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 127. [Google Scholar] [CrossRef]
- Liu, J.; Geng, X.; Hou, J.; Wu, G. New insights into M1/M2 macrophages: Key modulators in cancer progression. Cancer Cell Int. 2021, 21, 389. [Google Scholar] [CrossRef]
- Gao, J.; Liang, Y.; Wang, L. Shaping Polarization Of Tumor-Associated Macrophages In Cancer Immunotherapy. Front. Immunol. 2022, 13, 888713. [Google Scholar] [CrossRef]
- Zhong, Q.; Fang, Y.; Lai, Q.; Wang, S.; He, C.; Li, A.; Liu, S.; Yan, Q. CPEB3 inhibits epithelial-mesenchymal transition by disrupting the crosstalk between colorectal cancer cells and tumor-associated macrophages via IL-6R/STAT3 signaling. J. Exp. Clin. Cancer Res. 2020, 39, 132. [Google Scholar] [CrossRef]
- Han, I.H.; Song, H.O.; Ryu, J.S. IL-6 produced by prostate epithelial cells stimulated with Trichomonas vaginalis promotes proliferation of prostate cancer cells by inducing M2 polarization of THP-1-derived macrophages. PLoS Negl. Trop. Dis. 2020, 14, e0008126. [Google Scholar] [CrossRef]
- Chen, L.; Wang, S.; Wang, Y.; Zhang, W.; Ma, K.; Hu, C.; Zhu, H.; Liang, S.; Liu, M.; Xu, N. IL-6 influences the polarization of macrophages and the formation and growth of colorectal tumor. Oncotarget 2018, 9, 17443–17454. [Google Scholar] [CrossRef] [PubMed]
- Vilgelm, A.E. Illuminating the mechanism of IL-6-mediated immunotherapy resistance. Cell Rep. Med. 2023, 4, 100901. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Cheng, W.; Zhang, G.; Wang, C.; Sun, B.; Zha, C.; Kong, F.; Jia, Y. Long Noncoding RNA: Shining Stars in the Immune Microenvironment of Gastric Cancer. Front. Oncol. 2022, 12, 862337. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, L.; Zhou, S.; Lyu, J.; Gao, Y.; Yang, R. Construction and Validation of a Novel Pyroptosis-Related Four-lncRNA Prognostic Signature Related to Gastric Cancer and Immune Infiltration. Front. Immunol. 2022, 13, 854785. [Google Scholar] [CrossRef]
- Yao, F.; Zhan, Y.; Pu, Z.; Lu, Y.; Chen, J.; Deng, J.; Wu, Z.; Chen, B.; Chen, J.; Tian, K.; et al. LncRNAs Target Ferroptosis-Related Genes and Impair Activation of CD4(+) T Cell in Gastric Cancer. Front. Cell Dev. Biol. 2021, 9, 797339. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Li, R.; Han, J.; Sun, D.; Shen, L.; Wu, G. Identification of an Immune-Related LncRNA Signature in Gastric Cancer to Predict Survival and Response to Immune Checkpoint Inhibitors. Front. Cell Dev. Biol. 2021, 9, 739583. [Google Scholar] [CrossRef]
- Huang, J.; Chen, W.; Chen, C.; Jie, Z.; Xiao, T. Comprehensive Analysis and Prognosis Prediction of N6-Methyladenosine-Related lncRNAs in Immune Microenvironment Infiltration of Gastric Cancer. Int. J. Gen. Med. 2022, 15, 2629–2643. [Google Scholar] [CrossRef]
- Wang, J.; Wang, B.; Zhou, B.; Chen, J.; Qi, J.; Shi, L.; Yu, S.; Chen, G.; Kang, M.; Jin, X.; et al. A novel immune-related lncRNA pair signature for prognostic prediction and immune response evaluation in gastric cancer: A bioinformatics and biological validation study. Cancer Cell Int. 2022, 22, 69. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, G.Q.; Tian, D.; Zhou, C.W.; Li, N.; Feng, Y.; Zeng, M.S. Comprehensive analysis of tumor immune microenvironment and prognosis of m6A-related lncRNAs in gastric cancer. BMC Cancer 2022, 22, 316. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhao, L.; Wu, Y.; Deng, S.; Cao, P.; Lei, X.; Yang, X. The Role of NcRNAs to Regulate Immune Checkpoints in Cancer. Front. Immunol. 2022, 13, 853480. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, J. Role of non-coding RNA in immune microenvironment and anticancer therapy of gastric cancer. J. Mol. Med. 2022, 100, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, J.; Li, B.; Zhao, Z.; Liu, Y.; Zhao, Z.; Jing, S.; Wang, G. Identification of Potential lncRNAs and miRNAs as Diagnostic Biomarkers for Papillary Thyroid Carcinoma Based on Machine Learning. Int. J. Endocrinol. 2021, 2021, 3984463. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Park, H. Machine learning on thyroid disease: A review. Front. Biosci. 2022, 27, 101. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, S.; Meng, J.; Lyu, G.; Ding, G.; Hu, Y.; Wang, L.; Wu, L.; Yang, W.; Lv, Y.; et al. Long noncoding RNA PART1 restrains aggressive gastric cancer through the epigenetic silencing of PDGFB via the PLZF-mediated recruitment of EZH2. Oncogene 2020, 39, 6513–6528. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, X.; Li, S.; Sun, Y.; Zhai, H. Tumor Suppressor Gene XEDAR Promotes Differentiation and Suppresses Proliferation and Migration of Gastric Cancer Cells Through Upregulating the RELA/LXRalpha Axis and Deactivating the Wnt/beta-Catenin Pathway. Cell Transplant. 2021, 30, 963689721996346. [Google Scholar] [CrossRef]
- Moran, V.A.; Perera, R.J.; Khalil, A.M. Emerging functional and mechanistic paradigms of mammalian long non-coding RNAs. Nucleic Acids Res. 2012, 40, 6391–6400. [Google Scholar] [CrossRef]
- Jing, B.; Wang, T.; Sun, B.; Xu, J.; Xu, D.; Liao, Y.; Song, H.; Guo, W.; Li, K.; Hu, M.; et al. IL6/STAT3 Signaling Orchestrates Premetastatic Niche Formation and Immunosuppressive Traits in Lung. Cancer Res. 2020, 80, 784–797. [Google Scholar] [CrossRef]
- Yin, Z.; Ma, T.; Lin, Y.; Lu, X.; Zhang, C.; Chen, S.; Jian, Z. IL-6/STAT3 pathway intermediates M1/M2 macrophage polarization during the development of hepatocellular carcinoma. J. Cell. Biochem. 2018, 119, 9419–9432. [Google Scholar] [CrossRef]
- Wozniak, M.; Czyz, M. The Functional Role of Long Non-Coding RNAs in Melanoma. Cancers 2021, 13, 4848. [Google Scholar] [CrossRef] [PubMed]
- Pisani, G.; Baron, B. NEAT1 and Paraspeckles in Cancer Development and Chemoresistance. Noncoding RNA 2020, 6, 43. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Zhang, B.; An, R.; Qian, W.; Han, L.; Duan, W.; Wang, Z.; Ma, Q. Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer. Cancers 2022, 14, 4009. [Google Scholar] [CrossRef]
- Chang, T.H.; Huang, H.Y.; Hsu, J.B.; Weng, S.L.; Horng, J.T.; Huang, H.D. An enhanced computational platform for investigating the roles of regulatory RNA and for identifying functional RNA motifs. BMC Bioinform. 2013, 14 (Suppl. S2), S4. [Google Scholar] [CrossRef]
- Ren, Z.; Li, C.; Gan, Y.; Liu, X.; Liang, F. Long noncoding RNA taurine-up regulated gene 1 for the prognosis of osteosarcoma: A protocol for meta-analysis and bioinformatics analysis. Medicine 2021, 100, e26182. [Google Scholar] [CrossRef]
- Zhao, Y.; Teng, H.; Yao, F.; Yap, S.; Sun, Y.; Ma, L. Challenges and Strategies in Ascribing Functions to Long Noncoding RNAs. Cancers 2020, 12, 1458. [Google Scholar] [CrossRef] [PubMed]
- Piddock, R.E.; Marlein, C.R.; Abdul-Aziz, A.; Shafat, M.S.; Auger, M.J.; Bowles, K.M.; Rushworth, S.A. Myeloma-derived macrophage inhibitory factor regulates bone marrow stromal cell-derived IL-6 via c-MYC. J. Hematol. Oncol. 2018, 11, 66. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; McClusky, W.G.; Abdelmohsen, K.; Gorospe, M. Cytoplasmic functions of long noncoding RNAs. Wiley Interdiscip. Rev. RNA 2018, 9, e1471. [Google Scholar] [CrossRef]
- Zhou, C.; Duan, S. The Role of Long Non-Coding RNA NNT-AS1 in Neoplastic Disease. Cancers 2020, 12, 3086. [Google Scholar] [CrossRef]
- Elguindy, M.M.; Mendell, J.T. NORAD-induced Pumilio phase separation is required for genome stability. Nature 2021, 595, 303–308. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | High Expression | Low Expression | p Value |
|---|---|---|---|
| n | n | ||
| Age (years) | 0.728 | ||
| >60 | 26 (36.10%) | 46 (63.9%) | |
| <60 | 24 (32.4%) | 50 (67.60%) | |
| Gender | 1.000 | ||
| Male | 36 (34.0%) | 70 (66.0%) | |
| Female | 14 (35.0%) | 26 (65.0%) | |
| Size | 0.298 | ||
| <5 cm | 29 (37.2%) | 49 (62.8%) | |
| >5 cm | 19 (29.2%) | 46 (70.8%) | |
| Differentiation | 0.029 * | ||
| Good | 47 (38.2%) | 76 (61.8%) | |
| Poor | 3 (13.0%) | 20 (87.0%) | |
| Invasion depth | 0.051 | ||
| T1/T2 | 3 (15.0%) | 17 (85.0%) | |
| T3/T4 | 47 (37.3%) | 79 (62.7%) | |
| Tumor embolus | 0.723 | ||
| Yes | 30 (61.2%) | 55 (57.3%) | |
| No | 19 (38.8%) | 41 (42.7%) | |
| Lymphatic metastasis | 0.126 | ||
| Negative | 6 (22.2%) | 21 (77.8%) | |
| Positive | 44 (37.0%) | 75 (63.0%) | |
| Distant metastasis | 0.445 | ||
| Yes | 17 (39.5%) | 26 (60.5%) | |
| No | 33 (32.0%) | 70 (68.0%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Ding, G.; Wang, S.; Meng, J.; Lv, Y.; Yang, W.; Zhang, H.; Wen, X.; Zhao, W. Long Non-Coding RNA LOC339059 Attenuates IL-6/STAT3-Signaling-Mediated PDL1 Expression and Macrophage M2 Polarization by Interacting with c-Myc in Gastric Cancer. Cancers 2023, 15, 5313. https://doi.org/10.3390/cancers15225313
Han H, Ding G, Wang S, Meng J, Lv Y, Yang W, Zhang H, Wen X, Zhao W. Long Non-Coding RNA LOC339059 Attenuates IL-6/STAT3-Signaling-Mediated PDL1 Expression and Macrophage M2 Polarization by Interacting with c-Myc in Gastric Cancer. Cancers. 2023; 15(22):5313. https://doi.org/10.3390/cancers15225313
Chicago/Turabian StyleHan, Haibo, Guangyu Ding, Shanshan Wang, Junling Meng, Yunwei Lv, Wei Yang, Hong Zhang, Xianzi Wen, and Wei Zhao. 2023. "Long Non-Coding RNA LOC339059 Attenuates IL-6/STAT3-Signaling-Mediated PDL1 Expression and Macrophage M2 Polarization by Interacting with c-Myc in Gastric Cancer" Cancers 15, no. 22: 5313. https://doi.org/10.3390/cancers15225313
APA StyleHan, H., Ding, G., Wang, S., Meng, J., Lv, Y., Yang, W., Zhang, H., Wen, X., & Zhao, W. (2023). Long Non-Coding RNA LOC339059 Attenuates IL-6/STAT3-Signaling-Mediated PDL1 Expression and Macrophage M2 Polarization by Interacting with c-Myc in Gastric Cancer. Cancers, 15(22), 5313. https://doi.org/10.3390/cancers15225313