
LncRNA LINC01094 Promotes Cells Proliferation and Metastasis through the PTEN/AKT Pathway by Targeting AZGP1 in Gastric Cancer

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Total RNA Isolation, Reverse Transcription, and qRT-PCR
2.3. Subcellular Fractionation
2.4. RNA Fluorescence In Situ Hybridization (FISH)
2.5. Next-Generation Transcriptome Sequencing
2.6. Cell Lines and Culture Conditions
2.7. Short Tandem Repeat (STR) Profiling for Cell Lines
2.8. Western Blot
2.9. Transfection and Lentivirus Infection
2.10. Transwell Migration Assay
2.11. Proliferation and Colony-Formation Assays
2.12. In Vitro Sphere-Forming Assay
2.13. Transplantable Xenograft Mouse Model
2.14. HE Staining
2.15. Immunohistochemistry
2.16. RNA Pulldown Assay
2.17. Mass Spectrometry
2.18. RNA-Binding Protein Immunoprecipitation (RIP) Assay
2.19. Statistical Analysis
3. Results
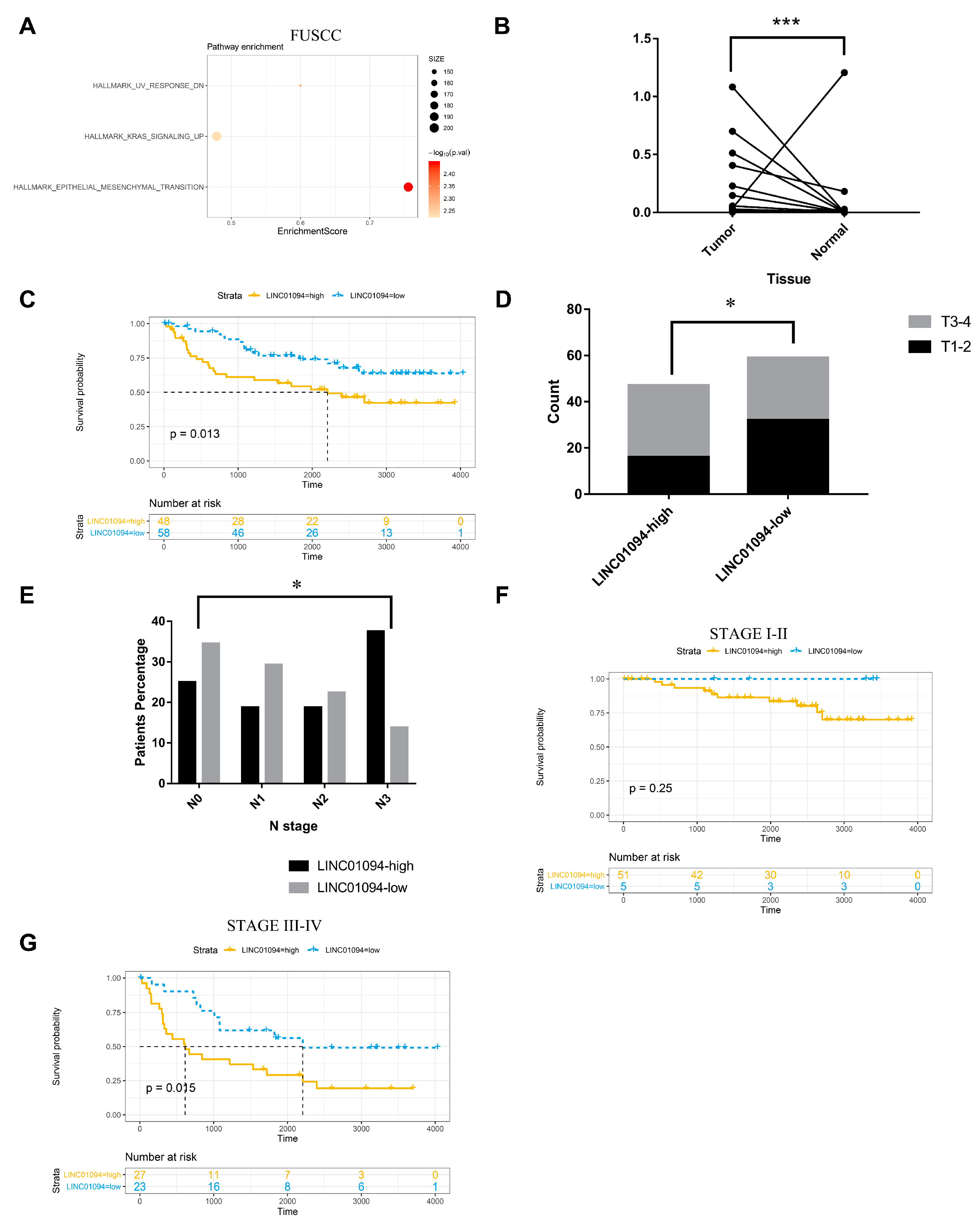
3.1. LINC01094 Was Correlated with Early Metastasis and Prognosis in GC
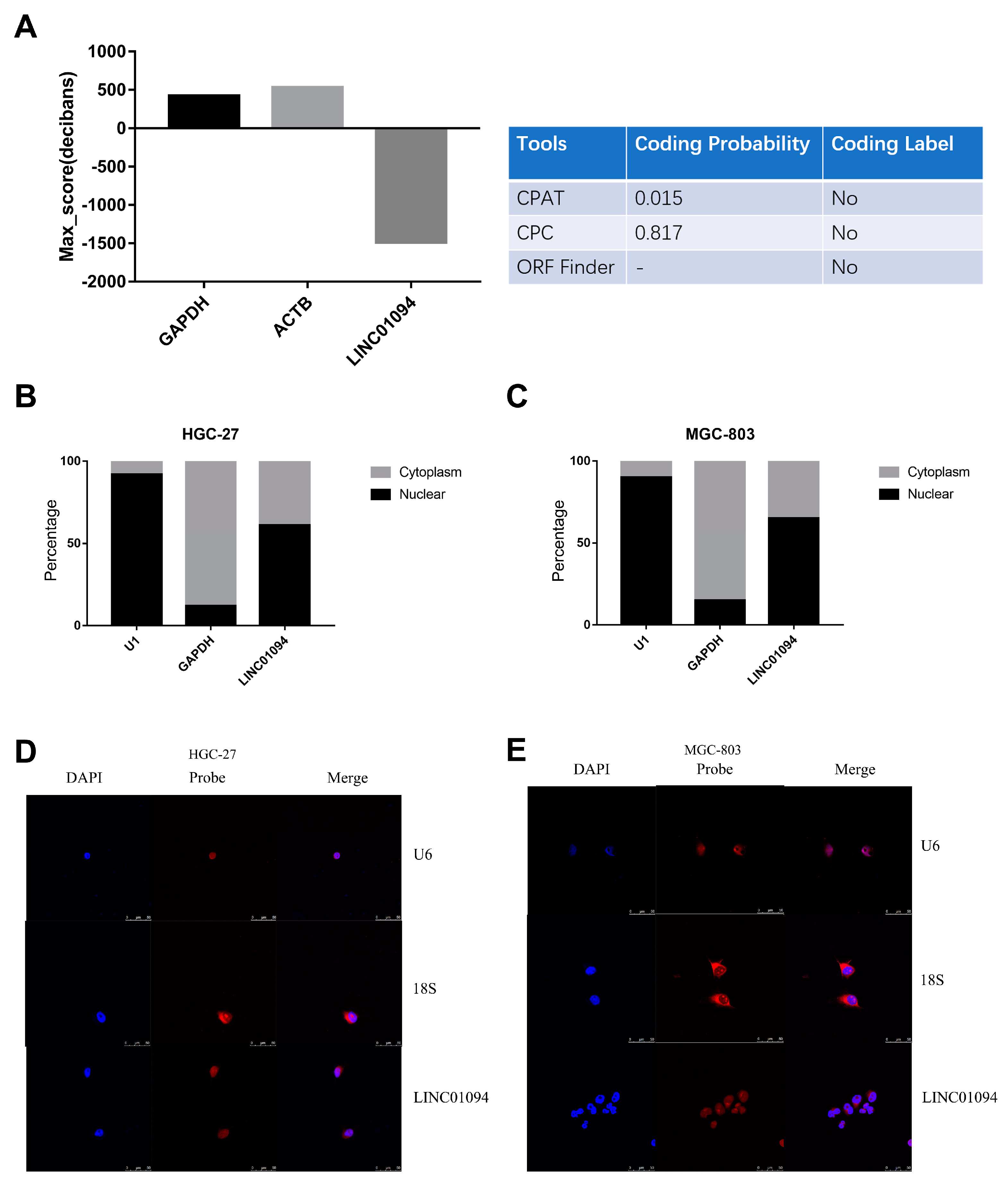
3.2. The Characteristics of LINC01094 in GC
3.3. LINC01094 Promotes Cells Proliferation and Metastasis in GC
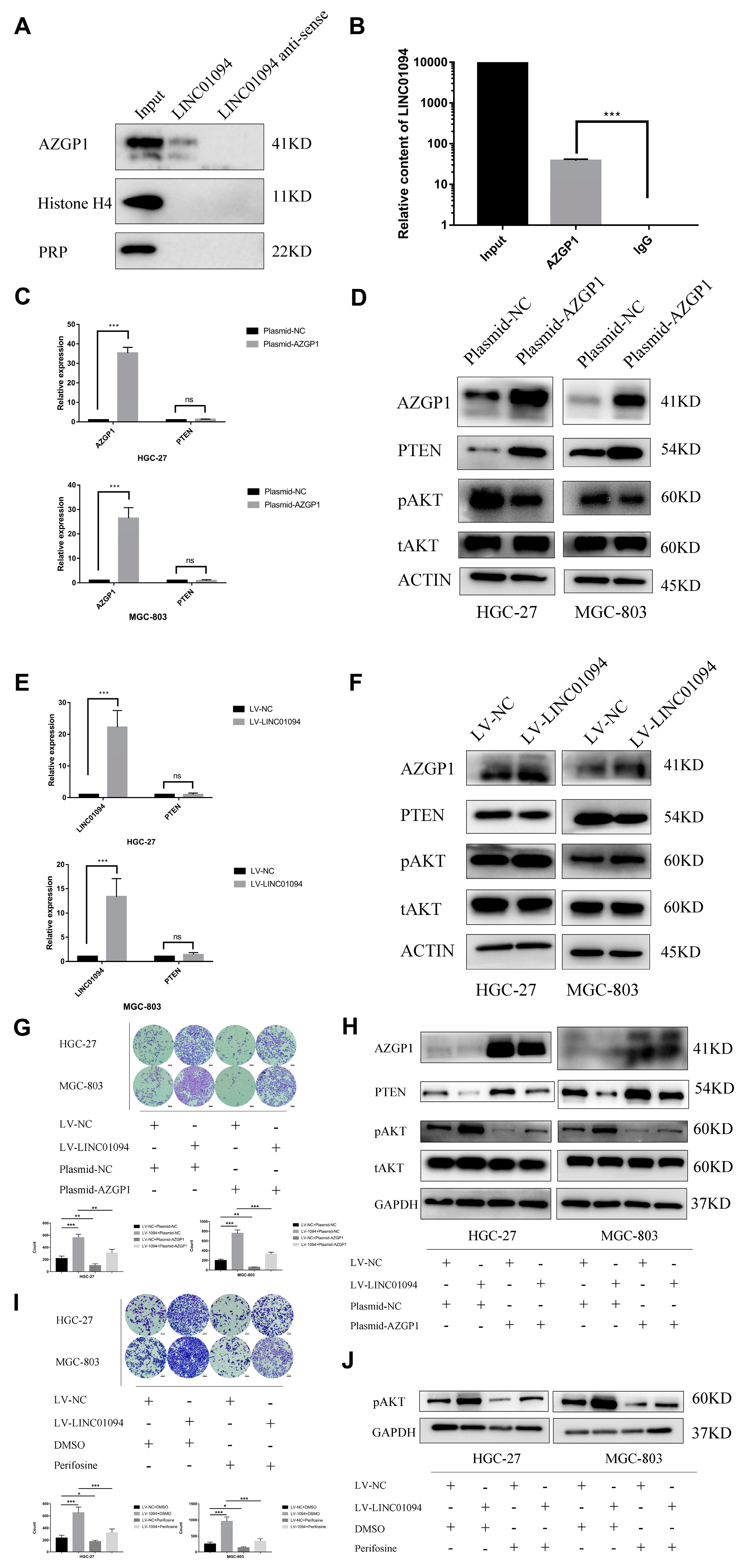
3.4. LINC01094 Directly Binds to AZGP1 in GC
3.5. LINC01094 Regulates PTEN Expression by Antagonizing the Function of AZGP1
3.6. LINC01094 Promotes Migration of Tumor Cells through the PTEN/AKT Pathway by Targeting AZGP1 in GC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, J.; Chen, Y.; Xu, R.; Zhao, Q.; Guo, W. MUC4, MUC16, and TTN genes mutation correlated with prognosis, and predicted tumor mutation burden and immunotherapy efficacy in gastric cancer and pan-cancer. Clin. Transl. Med. 2020, 10, e155. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Ding, Y.; Zong, W.; Ju, S. Non-coding RNAs in regulating gastric cancer metastasis. Clin. Chim. Acta 2019, 496, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.D.; Huang, M.Z.; Wang, Y.S.; Feng, W.J.; Chen, Z.Y.; He, Y.F.; Zhang, X.W.; Liu, X.; Wang, C.C.; Zhang, W.; et al. XELOX doublet regimen versus EOX triplet regimen as first-line treatment for advanced gastric cancer: An open-labeled, multicenter, randomized, prospective phase III trial (EX-ELOX). Cancer Commun. 2022, 42, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.; Engleitner, T.; Maresch, R.; Zukowska, M.; Lange, S.; Kaltenbacher, T.; Konukiewitz, B.; Öllinger, R.; Zwiebel, M.; Strong, A.; et al. Evolutionary routes and KRAS dosage define pancreatic cancer phenotypes. Nature 2018, 554, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, F.; Yang, Y.; Yu, N.; Weng, X.; Yang, Y.; Gong, Z.; Huang, S.; Gan, L.; Sun, S.; et al. Integrated DNA and RNA sequencing reveals early drivers involved in metastasis of gastric cancer. Cell Death Dis. 2022, 13, 392. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Xiang, Z.; Wu, H.; He, Q.; Dou, R.; Lin, Z.; Yang, C.; Huang, S.; Song, J.; Di, Z.; et al. The lncRNA BDNF-AS/WDR5/FBXW7 axis mediates ferroptosis in gastric cancer peritoneal metastasis by regulating VDAC3 ubiquitination. Int. J. Biol. Sci. 2022, 18, 1415–1433. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Gou, H.; Wang, D.; Li, C.; Li, Y.; Su, H.; Wang, X.; Zhang, X.; Yu, J. LncRNA TNFRSF10A-AS1 promotes gastric cancer by directly binding to oncogenic MPZL1 and is associated with patient outcome. Int. J. Biol. Sci. 2022, 18, 3156–3166. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Huang, T.; Cheng, A.S.; Yu, J.; Kang, W.; To, K.F. The Interplay of LncRNA-H19 and Its Binding Partners in Physiological Process and Gastric Carcinogenesis. Int. J. Mol. Sci. 2017, 18, 450. [Google Scholar] [CrossRef]
- Fattahi, S.; Kosari-Monfared, M.; Golpour, M.; Emami, Z.; Ghasemiyan, M.; Nouri, M.; Akhavan-Niaki, H. LncRNAs as potential diagnostic and prognostic biomarkers in gastric cancer: A novel approach to personalized medicine. J. Cell. Physiol. 2019, 235, 3189–3206. [Google Scholar] [CrossRef]
- Ramli, S.; Sim, M.S.; Guad, R.M.; Gopinath, S.C.; Subramaniyan, V.; Fuloria, S.; Fuloria, N.K.; Choy, K.W.; Rana, S.; Wu, Y.S. Long Noncoding RNA UCA1 in Gastro-intestinal Cancers: Molecular Regulatory Roles and Patterns, Mechanisms, and Interactions. J. Oncol. 2021, 2021, 5519720. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Zhen-Hua, W.; Yi-Wei, G.; Li-Qin, Z.; Jie-Yun, Z.; Zhe, G.; Wei-Jian, G. Silencing of LncRNA C1RL-AS1 Suppresses the Malignant Phenotype in Gastric Cancer Cells via the AKT/beta-Catenin/c-Myc Pathway. Front. Oncol. 2020, 10, 1508. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-H.; Liu, C.-C.; Zhou, Y.-Q.; Hu, L.-N.; Guo, W.-J. OnclncRNA-626 promotes malignancy of gastric cancer via inactivated the p53 pathway through interacting with SRSF1. Am. J. Cancer Res. 2019, 9, 2249–2263. [Google Scholar] [PubMed]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. In Long Non Coding RNA Biology; Springer: Singapore, 2017; Volume 1008, pp. 1–46. [Google Scholar] [CrossRef]
- Kung, J.T.Y.; Colognori, D.; Lee, J.T. Long Noncoding RNAs: Past, Present, and Future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef]
- Wang, M.; Gu, S.; Liu, Y.; Yang, Y.; Yan, J.; Zhang, X.; An, X.; Gao, J.; Hu, X.; Su, P. miRNA-PDGFRB/HIF1A-lncRNA CTEPHA1 Network Plays Important Roles in the Mechanism of Chronic Thromboembolic Pulmonary Hypertension. Int. Heart J. 2019, 60, 924–937. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, W.; Yan, Y.; Yao, X.; Gu, W.; Zhang, H. LINC01094 triggers radio-resistance in clear cell renal cell carcinoma via miR-577/CHEK2/FOXM1 axis. Cancer Cell Int. 2020, 20, 274. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Wu, J.; Ji, H.; Chen, Z.; Guo, H.; Hou, J. Long Non-coding RNA LINC01094 Promotes the Development of Clear Cell Renal Cell Carcinoma by Upregulating SLC2A3 via MicroRNA-184. Front. Genet. 2020, 11, 562967. [Google Scholar] [CrossRef]
- Li, X.X.; Yu, Q. Linc01094 Accelerates the Growth and Metastatic-Related Traits of Glioblastoma by Sponging miR-126-5p. OncoTargets Ther. 2020, 13, 9917–9928. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, P.; Sun, H.; Liu, Y. LINC01094/miR-577 axis regulates the progression of ovarian cancer. J. Ovarian Res. 2020, 13, 122. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, W.; Liu, H.; Xu, Q.; Xu, W. LINC01094 Down-Regulates miR-330-3p and Enhances the Expression of MSI1 to Promote the Progression of Glioma. Cancer Manag. Res. 2020, 12, 6511–6521. [Google Scholar] [CrossRef]
- Wei, X.; Liu, X.; Tan, C.; Mo, L.; Wang, H.; Peng, X.; Deng, F.; Chen, L. Expression and Function of Zinc-α2-Glycoprotein. Neurosci. Bull. 2019, 35, 540–550. [Google Scholar] [CrossRef]
- Tsourlakis, M.C.; Kluth, M.; Kleinhaus, S.; Masser, S.; Burdelski, C.; Huland, H.; Heinzer, H.; Graefen, M.; Beyer, B.; Steurer, S.; et al. Abstract 572: Reduced AZGP1 expression is an independent predictor of early PSA recurrence and associated with ERG fusion positive, PTEN deleted prostate cancers. Int. J. Cancer 2014, 74, 572. [Google Scholar] [CrossRef]
- Li, W.; Liu, J. Overexpression of the zinc-α2-glycoprotein accelerates apoptosis and inhibits growth via the mTOR/PTEN signaling pathway in gastric carcinoma cells. Life Sci. 2020, 240, 117117. [Google Scholar] [CrossRef]
- Severo, J.S.; Morais, J.B.S.; Beserra, J.B.; dos Santos, L.R.; de Sousa Melo, S.R.; de Sousa, G.S.; de Matos Neto, E.M.; Henriques, G.S.; do Nascimento Marreiro, D. Role of Zinc in Zinc-α2-Glycoprotein Metabolism in Obesity: A Review of Literature. Biol. Trace Elem. Res. 2020, 193, 81–88. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Zhao, J.-J.; Lv, L.; Chen, Y.-B.; Li, Y.-F.; Jiang, S.-S.; Wang, W.; Pan, K.; Zheng, Y.; Zhao, B.-W.; et al. Decreased Expression of AZGP1 Is Associated with Poor Prognosis in Primary Gastric Cancer. PLoS ONE 2013, 8, e69155. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Ge, C.; Zhao, F.; Zhu, M.; Zhang, L.; Huo, Q.; Li, H.; Chen, T.; Xie, H.; Cui, Y.; et al. Downregulation of AZGP1 by Ikaros and histone deacetylase promotes tumor progression through the PTEN/Akt and CD44s pathways in hepatocellular carcinoma. Carcinogenesis 2017, 38, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Yu, F.; Yan, D.; Cui, F.; Tang, H.; Wang, X.; Chen, J.; Lu, H.; Zhao, S.; Peng, Z. Zinc-α-2-Glycoprotein: A Candidate Biomarker for Colon Cancer Diagnosis in Chinese Population. Int. J. Mol. Sci. 2014, 16, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-H.; Lin, C.; Liu, C.-C.; Jiang, W.-W.; Huang, M.-Z.; Liu, X.; Guo, W.-J. MiR-616-3p promotes angiogenesis and EMT in gastric cancer via the PTEN/AKT/mTOR pathway. Biochem. Biophys. Res. Commun. 2018, 501, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Xu, M.; Hua, R.-X.; Tan, C.; Zhang, J.; Gong, Y.; Wu, Z.; Weng, W.; Sheng, W.; Guo, W. The polycomb group protein EZH2 induces epithelial–mesenchymal transition and pluripotent phenotype of gastric cancer cells by binding to PTEN promoter. J. Hematol. Oncol. 2018, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.-H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Shariati, M.; Meric-Bernstam, F. Targeting AKT for cancer therapy. Expert Opin. Investig. Drugs 2019, 28, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.W.; Chau, I. Molecular target: Pan-AKT in gastric cancer. ESMO Open 2020, 5, e000728. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Ge, X.; Zhao, W.; Xue, L.; Dai, C.; Lin, F.; Peng, W. PIPKIgamma Regulates CCL2 Expression in Colorectal Cancer by Activating AKT-STAT3 Signaling. J. Immunol. Res. 2019, 2019, 3690561. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ji, H.; Niu, X.; Yin, L.; Wang, Y.; Gu, Y.; Wang, J.; Zhou, X.; Zhang, H.; Zhang, Q. Tumor-associated macrophages secrete CC-chemokine ligand 2 and induce tamoxifen resistance by activating PI3K/Akt/mTOR in breast cancer. Cancer Sci. 2020, 111, 47–58. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | LINC01094-High | LINC01094-Low | p-Value | |||
|---|---|---|---|---|---|---|
| N = 48 | N = 58 | |||||
| N | % | N | % | |||
| Gender | 0.4212 | |||||
| Male | 38 | 79.17 | 42 | 72.41 | ||
| Female | 10 | 20.83 | 16 | 27.59 | ||
| Age | 0.6143 | |||||
| ≥60 | 28 | 58.33 | 31 | 53.45 | ||
| <60 | 20 | 41.67 | 27 | 46.55 | ||
| T | 0.1503 | |||||
| 1 | 12 | 25.00 | 27 | 46.55 | ||
| 2 | 4 | 8.33 | 4 | 6.90 | ||
| 3 | 1 | 2.08 | 1 | 1.72 | ||
| 4 | 31 | 64.58 | 26 | 44.83 | ||
| N | 0.04274 | |||||
| 0 | 12 | 25.00 | 20 | 34.48 | ||
| 1 | 9 | 18.75 | 17 | 29.31 | ||
| 2 | 9 | 18.75 | 13 | 22.41 | ||
| 3 | 18 | 37.50 | 8 | 13.79 | ||
| M | 0.2694 | |||||
| 0 | 47 | 97.92 | 58 | 100.00 | ||
| 1 | 1 | 2.08 | 0 | 0.00 | ||
| Histology | 0.3803 | |||||
| Adenocarcinoma | 39 | 81.25 | 50 | 86.21 | ||
| Mucinous adenocarcinoma | 6 | 12.50 | 3 | 5.17 | ||
| Signet-ring cell carcinoma | 3 | 6.25 | 5 | 8.62 | ||
| Grade | 0.2628 | |||||
| 1–2 | 11 | 22.92 | 19 | 32.76 | ||
| 3–4 | 37 | 77.08 | 39 | 67.24 | ||
| Vessel invasion | 0.212 | |||||
| Invasion | 24 | 50.00 | 22 | 37.93 | ||
| No invasion | 24 | 50.00 | 36 | 62.07 | ||
| Neuroinvasion | 0.009942 | |||||
| Invasion | 25 | 52.08 | 16 | 27.59 | ||
| No invasion | 23 | 47.92 | 42 | 72.41 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, Z.; Zhang, Y.; Yang, Y.; Yang, Y.; Zhang, J.; Wang, Y.; Zhao, L.; Yu, N.; Wu, Z.; Guo, W. LncRNA LINC01094 Promotes Cells Proliferation and Metastasis through the PTEN/AKT Pathway by Targeting AZGP1 in Gastric Cancer. Cancers 2023, 15, 1261. https://doi.org/10.3390/cancers15041261
Gong Z, Zhang Y, Yang Y, Yang Y, Zhang J, Wang Y, Zhao L, Yu N, Wu Z, Guo W. LncRNA LINC01094 Promotes Cells Proliferation and Metastasis through the PTEN/AKT Pathway by Targeting AZGP1 in Gastric Cancer. Cancers. 2023; 15(4):1261. https://doi.org/10.3390/cancers15041261
Chicago/Turabian StyleGong, Zhe, Yanqiu Zhang, Yue Yang, Yanan Yang, Jieyun Zhang, Yixuan Wang, Liqin Zhao, Nuoya Yu, Zhenhua Wu, and Weijian Guo. 2023. "LncRNA LINC01094 Promotes Cells Proliferation and Metastasis through the PTEN/AKT Pathway by Targeting AZGP1 in Gastric Cancer" Cancers 15, no. 4: 1261. https://doi.org/10.3390/cancers15041261
APA StyleGong, Z., Zhang, Y., Yang, Y., Yang, Y., Zhang, J., Wang, Y., Zhao, L., Yu, N., Wu, Z., & Guo, W. (2023). LncRNA LINC01094 Promotes Cells Proliferation and Metastasis through the PTEN/AKT Pathway by Targeting AZGP1 in Gastric Cancer. Cancers, 15(4), 1261. https://doi.org/10.3390/cancers15041261