
Glycomic, Glycoproteomic, and Proteomic Profiling of Philippine Lung Cancer and Peritumoral Tissues: Case Series Study of Patients Stages I–III

 , , , , , ,
, , , , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement, Clinical Sample Collection, and Characterization
2.2. Sample Preparation for Mass Spectrometric Analyses
2.3. N-Glycan Release and Glycomic Analysis Using Chip-QToF LC-MS/MS
2.4. Proteomics and Glycoproteomic Analysis Using nLC-Orbitrap LC-MS/MS
3. Results and Discussion
3.1. Clinical Profile of the Filipino Lung Cancer Cohort
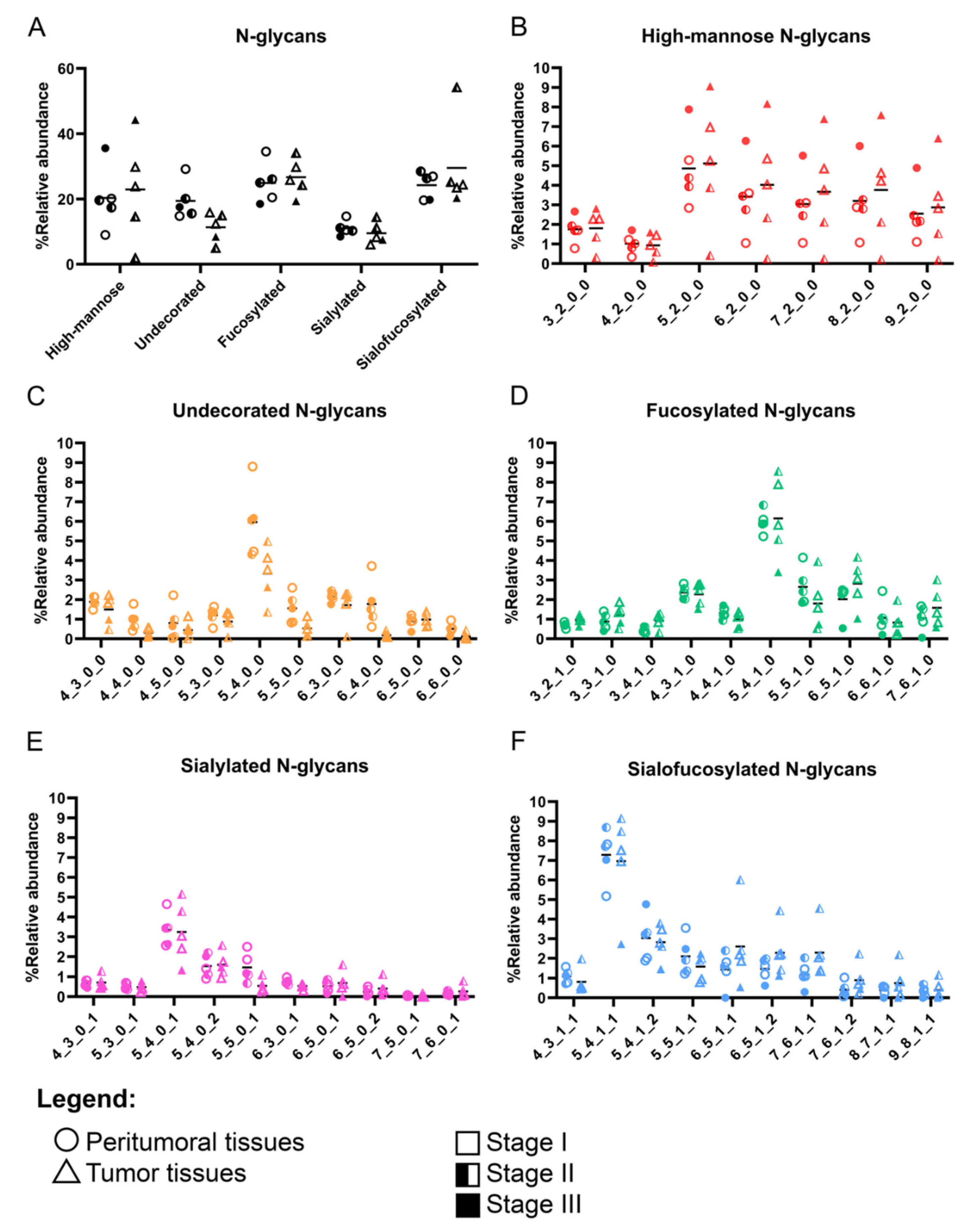
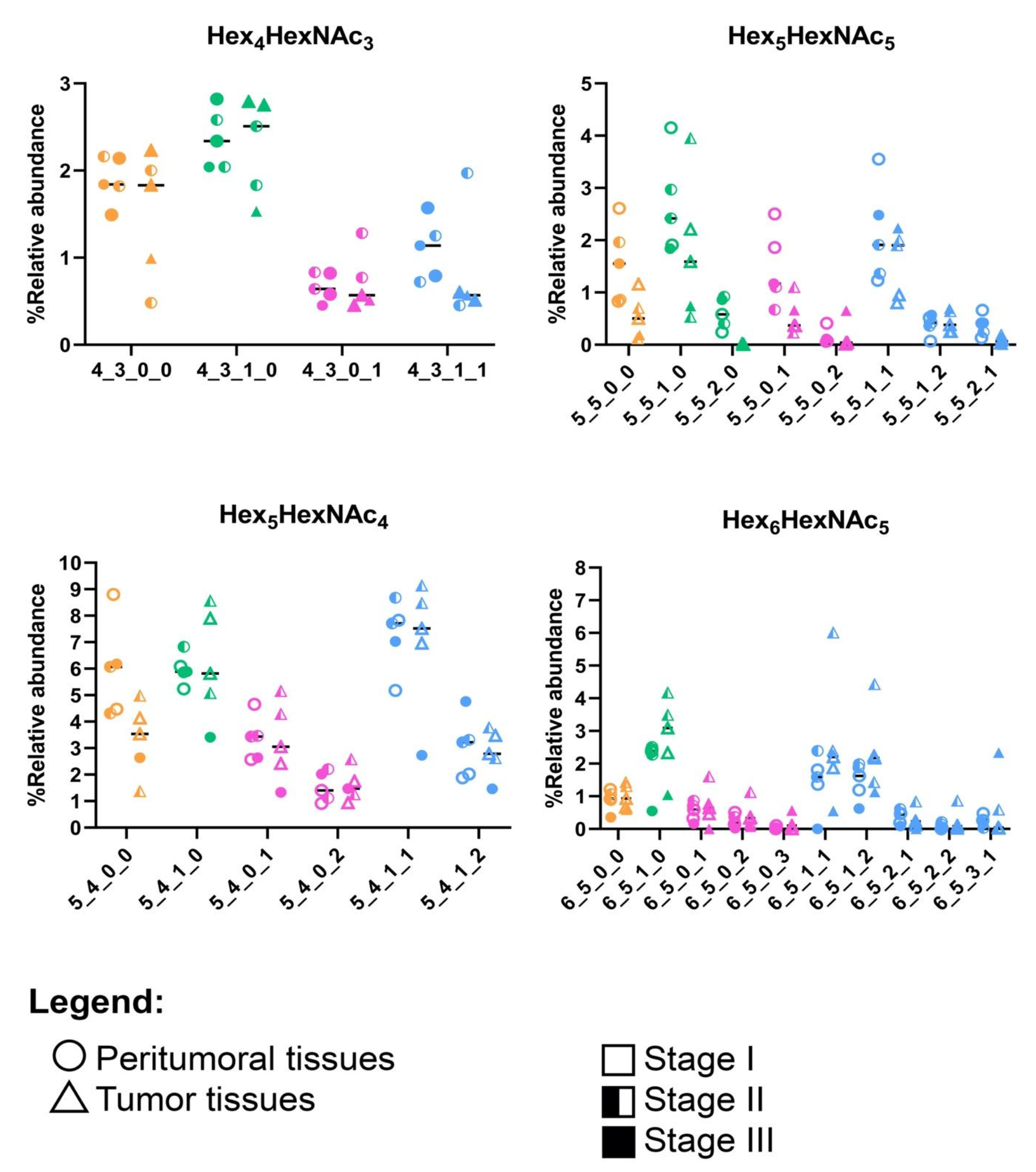
3.2. N-Glycomic Profiles of the Filipino Lung Cancer Cohort
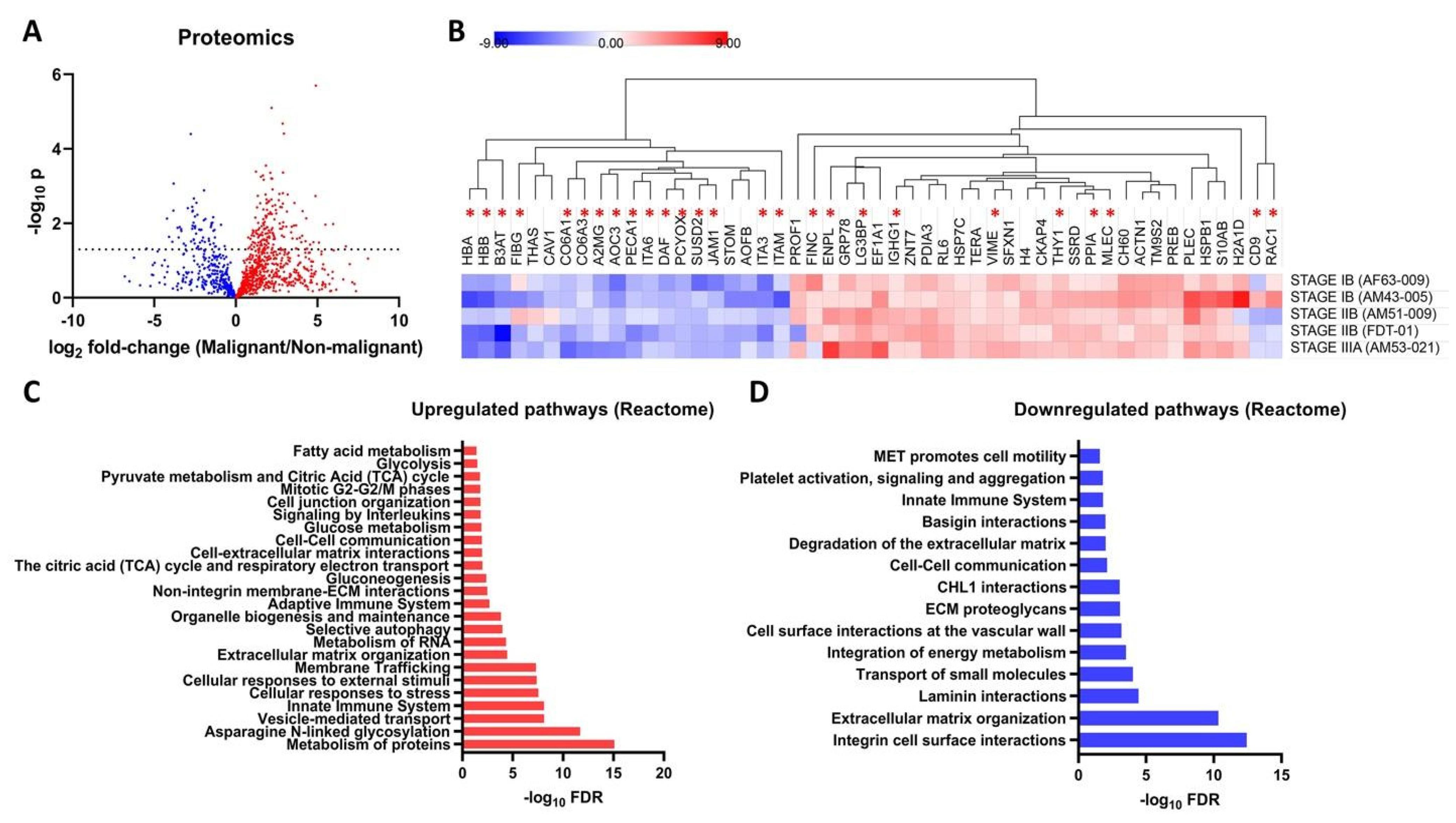
3.3. Proteomic Profiles of the Filipino Lung Cancer Cohort
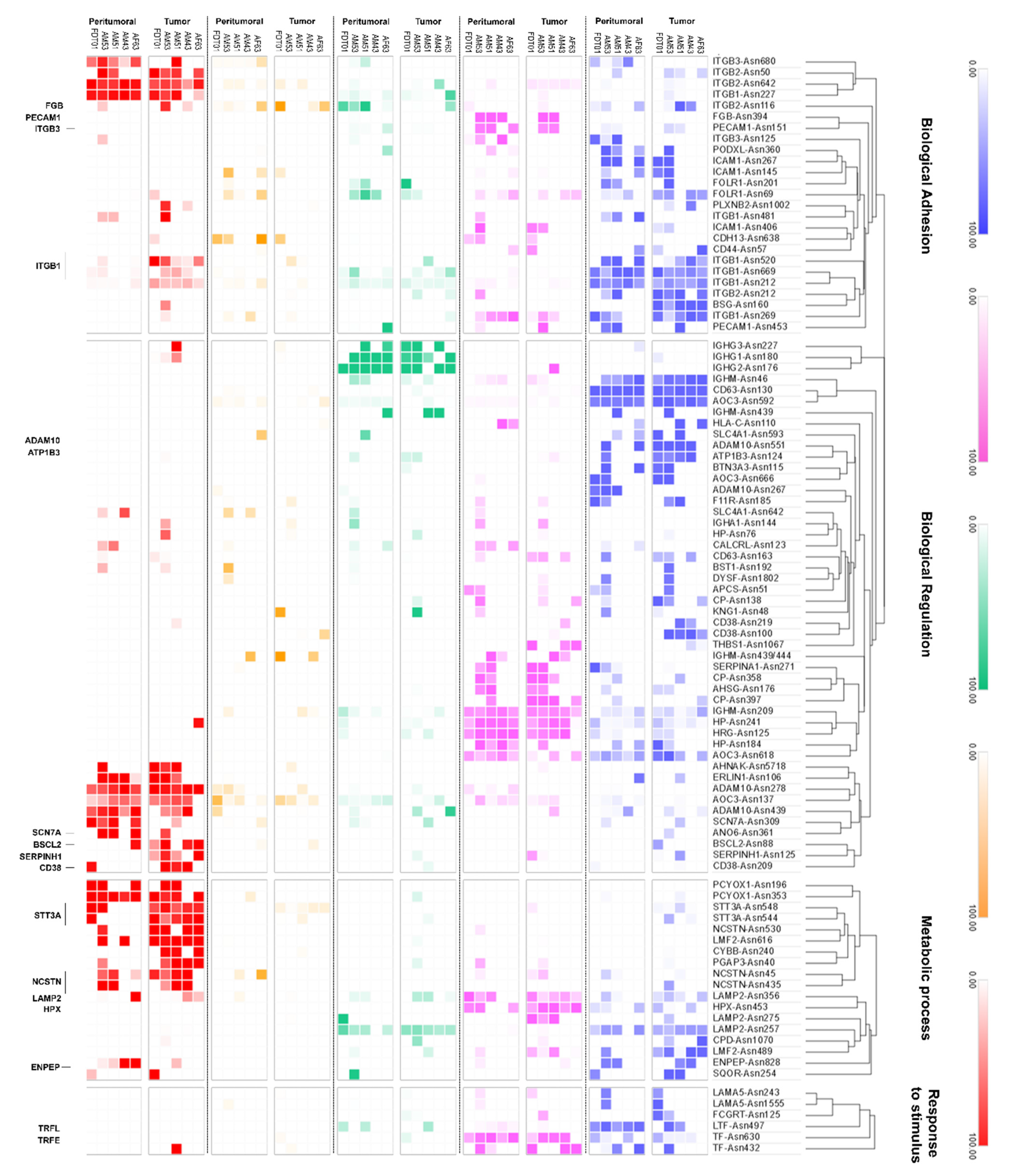
3.4. Glycoproteomic Profiles of the Filipino Lung Cancer Cohort
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Navada, S.; Lai, P.; Schwartz, A.G.; Kalemkerian, G.P. Temporal Trends in Small Cell Lung Cancer: Analysis of the National Surveillance, Epidemiology, and End-Results (SEER) Database. JCO 2006, 24, 7082. [Google Scholar] [CrossRef]
- Alberg, A.J.; Brock, M.V.; Samet, J.M. Epidemiology of Lung Cancer: Looking to the Future. JCO 2005, 23, 3175–3185. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.; Najafi, F.; Dobson, A. Meta-Analysis of Studies of Passive Smoking and Lung Cancer: Effects of Study Type and Continent. Int. J. Epidemiol. 2007, 36, 1048–1059. [Google Scholar] [CrossRef]
- Freudenheim, J.L.; Ritz, J.; Smith-Warner, S.A.; Albanes, D.; Bandera, E.V.; van den Brandt, P.A.; Colditz, G.; Feskanich, D.; Goldbohm, R.A.; Harnack, L.; et al. Alcohol Consumption and Risk of Lung Cancer: A Pooled Analysis of Cohort Studies. Am. J. Clin. Nutr. 2005, 82, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-J.; Lozano, G.; Amos, C.I.; Strong, L.C. Germline P53 Mutations in a Cohort with Childhood Sarcoma: Sex Differences in Cancer Risk. Am. J. Hum. Genet. 2003, 72, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-I.; Jeong, J.; Lee, C.W. Association between EGFR Mutation and Ageing, History of Pneumonia and Gastroesophageal Reflux Disease among Patients with Advanced Lung Cancer. Eur. J. Cancer 2019, 122, 101–108. [Google Scholar] [CrossRef]
- Aldrighetti, C.M.; Niemierko, A.; Van Allen, E.; Willers, H.; Kamran, S.C. Racial and Ethnic Disparities among Participants in Precision Oncology Clinical Studies. JAMA Netw. Open 2021, 4, e2133205. [Google Scholar] [CrossRef]
- Bruno, D.S.; Hess, L.M.; Li, X.; Su, E.W.; Zhu, Y.E.; Patel, M. Racial Disparities in Biomarker Testing and Clinical Trial Enrollment in Non-Small Cell Lung Cancer (NSCLC). JCO 2021, 39, 9005. [Google Scholar] [CrossRef]
- Cheng, T.-Y.D.; Cramb, S.M.; Baade, P.D.; Youlden, D.R.; Nwogu, C.; Reid, M.E. The International Epidemiology of Lung Cancer: Latest Trends, Disparities, and Tumor Characteristics. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2016, 11, 1653–1671. [Google Scholar] [CrossRef]
- Guerrero, S.; López-Cortés, A.; Indacochea, A.; García-Cárdenas, J.M.; Zambrano, A.K.; Cabrera-Andrade, A.; Guevara-Ramírez, P.; González, D.A.; Leone, P.E.; Paz-y-Miño, C. Analysis of Racial/Ethnic Representation in Select Basic and Applied Cancer Research Studies. Sci. Rep. 2018, 8, 13978. [Google Scholar] [CrossRef] [PubMed]
- Manne, U.; Jadhav, T.; Putcha, B.-D.K.; Samuel, T.; Soni, S.; Shanmugam, C.; Suswam, E.A. Molecular Biomarkers of Colorectal Cancer and Cancer Disparities: Current Status and Perspective. Curr. Color. Cancer Rep. 2016, 12, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Meany, D.L.; Chan, D.W. Aberrant Glycosylation Associated with Enzymes as Cancer Biomarkers. Clin. Proteom. 2011, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, F.; Malagolini, N.; Trinchera, M.; Chiricolo, M. Mechanisms of Cancer-Associated Glycosylation Changes. Front. Biosci. 2012, 17, 670–699. [Google Scholar] [CrossRef]
- Dall’Olio, F.; Chiricolo, M. Sialyltransferases in Cancer. Glycoconj. J. 2001, 18, 841–850. [Google Scholar] [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in Cancer: Mechanisms and Clinical Implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Yen, H.-Y.; Chen, C.-Y.; Chen, C.-H.; Cheng, P.-F.; Juan, Y.-H.; Chen, C.-H.; Khoo, K.-H.; Yu, C.-J.; Yang, P.-C.; et al. Sialylation and Fucosylation of Epidermal Growth Factor Receptor Suppress Its Dimerization and Activation in Lung Cancer Cells. Proc. Natl. Acad. Sci. USA 2011, 108, 11332–11337. [Google Scholar] [CrossRef]
- Munkley, J.; Elliott, D.J. Hallmarks of Glycosylation in Cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [CrossRef]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein Glycosylation in Cancer. Annu. Rev. Pathol. Mech. Dis. 2015, 10, 473–510. [Google Scholar] [CrossRef]
- Hoja-Łukowicz, D.; Przybyło, M.; Duda, M.; Pocheć, E.; Bubka, M. On the Trail of the Glycan Codes Stored in Cancer-Related Cell Adhesion Proteins. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 3237–3257. [Google Scholar] [CrossRef]
- Oliveira-Ferrer, L.; Legler, K.; Milde-Langosch, K. Role of Protein Glycosylation in Cancer Metastasis. Semin. Cancer Biol. 2017, 44, 141–152. [Google Scholar] [CrossRef] [PubMed]
- De Coura, M.M.A.; Barbosa, E.A.; Brand, G.D.; Bloch, C.; de Sousa, J.B. Identification of Differential N-Glycan Compositions in the Serum and Tissue of Colon Cancer Patients by Mass Spectrometry. Biology 2021, 10, 343. [Google Scholar] [CrossRef] [PubMed]
- Kailemia, M.J.; Park, D.; Lebrilla, C.B. Glycans and Glycoproteins as Specific Biomarkers for Cancer. Anal. Bioanal. Chem. 2017, 409, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Stroble, C.D.; Taylor, S.; Hong, Q.; Lebrilla, C.B.; Leiserowitz, G.S.; Kim, K.; Ruhaak, L.R. Multiple Reaction Monitoring for the Quantitation of Serum Protein Glycosylation Profiles: Application to Ovarian Cancer. J. Proteome Res. 2018, 17, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Ruhaak, L.R.; Kim, K.; Stroble, C.; Taylor, S.L.; Hong, Q.; Miyamoto, S.; Lebrilla, C.B.; Leiserowitz, G. Protein-Specific Differential Glycosylation of Immunoglobulins in Serum of Ovarian Cancer Patients. J. Proteome Res. 2016, 15, 1002–1010. [Google Scholar] [CrossRef]
- Park, D.D.; Phoomak, C.; Xu, G.; Olney, L.P.; Tran, K.A.; Park, S.S.; Haigh, N.E.; Luxardi, G.; Lert-itthiporn, W.; Shimoda, M.; et al. Metastasis of Cholangiocarcinoma Is Promoted by Extended High-Mannose Glycans. Proc. Natl. Acad. Sci. USA 2020, 117, 7633–7644. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Stroble, C.; Dai, J.; Barnett, M.; Taguchi, A.; Goodman, G.E.; Miyamoto, S.; Gandara, D.; Feng, Z.; Lebrilla, C.B.; et al. Serum Glycans as Risk Markers for Non-Small Cell Lung Cancer. Cancer Prev. Res. 2016, 9, 317–323. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Taylor, S.L.; Stroble, C.; Nguyen, U.T.; Parker, E.A.; Song, T.; Lebrilla, C.B.; Rom, W.N.; Pass, H.; Kim, K.; et al. Differential N-Glycosylation Patterns in Lung Adenocarcinoma Tissue. J. Proteome Res. 2015, 14, 4538–4549. [Google Scholar] [CrossRef]
- Landi, M.T.; Dracheva, T.; Rotunno, M.; Figueroa, J.D.; Liu, H.; Dasgupta, A.; Mann, F.E.; Fukuoka, J.; Hames, M.; Bergen, A.W.; et al. Gene Expression Signature of Cigarette Smoking and Its Role in Lung Adenocarcinoma Development and Survival. PLoS ONE 2008, 3, e1651. [Google Scholar] [CrossRef]
- Alvarez, M.R.S.; Zhou, Q.; Tena, J.; Lebrilla, C.B.; Completo, G.C.; Heralde, F.M.; Cabanatan, M.; Barzaga, M.T.; Tan-Liu, N.; Ladrera, G.I.; et al. N-Glycan and Glycopeptide Serum Biomarkers in Philippine Lung Cancer Patients Identified Using Liquid Chromatography–Tandem Mass Spectrometry. ACS Omega 2022, 7, 40230–40240. [Google Scholar] [CrossRef]
- Cocadiz, J.A.M.; Lim, C.C.Y.; Dalet, J.T.; Barzaga, M.T.; Heralde III, F.M. CD133, MUC1, and KRT19 Chromosomal Rearrangements and Gene Expressions as Potential Biomarkers for Liver Cancer. Int. J. Med. Res. Health Sci. 2017, 6, 21–31. [Google Scholar]
- Li, Q.; Xie, Y.; Wong, M.; Barboza, M.; Lebrilla, C.B. Comprehensive Structural Glycomic Characterization of the Glycocalyxes of Cells and Tissues. Nat. Protoc. 2020, 15, 2668–2704. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.R.S.; Zhou, Q.; Grijaldo, S.J.B.; Lebrilla, C.B.; Nacario, R.C.; Heralde, F.M.; Rabajante, J.F.; Completo, G.C. An Integrated Mass Spectrometry-Based Glycomics-Driven Glycoproteomics Analytical Platform to Functionally Characterize Glycosylation Inhibitors. Molecules 2022, 27, 3834. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xie, Y.; Wong, M.; Lebrilla, C. Characterization of Cell Glycocalyx with Mass Spectrometry Methods. Cells 2019, 8, 882. [Google Scholar] [CrossRef]
- Zhou, Q.; Xie, Y.; Lam, M.; Lebrilla, C.B. N-Glycomic Analysis of the Cell Shows Specific Effects of Glycosyl Transferase Inhibitors. Cells 2021, 10, 2318. [Google Scholar] [CrossRef] [PubMed]
- Kronewitter, S.R.; An, H.J.; de Leoz, M.L.; Lebrilla, C.B.; Miyamoto, S.; Leiserowitz, G.S. The Development of Retrosynthetic Glycan Libraries to Profile and Classify the Human Serum N-Linked Glycome. Proteomics 2009, 9, 2986–2994. [Google Scholar] [CrossRef]
- The Uniprot Consortium UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER Version 16: A Revised Family Classification, Tree-Based Classification Tool, Enhancer Regions and Extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Luna, H.G.; Cristal-Luna, G. 55P CTC Count & Gene Expression Profiling among Filipino NSCLC. Ann. Oncol. 2016, 27, ix15–ix16. [Google Scholar] [CrossRef]
- Situ, D.; Wang, J.; Ma, Y.; Zhu, Z.; Hu, Y.; Long, H.; Rong, T. Expression and Prognostic Relevance of MUC1 in Stage IB Non-Small Cell Lung Cancer. Med. Oncol. 2011, 28 (Suppl. S1), S596–S604. [Google Scholar] [CrossRef]
- Qu, H.; Li, R.; Liu, Z.; Zhang, J.; Luo, R. Prognostic Value of Cancer Stem Cell Marker CD133 Expression in Non-Small Cell Lung Cancer: A Systematic Review. Int. J. Clin. Exp. Pathol. 2013, 6, 2644–2650. [Google Scholar] [PubMed]
- Chang, J.S.; Chen, L.-T.; Shan, Y.-S.; Lin, S.-F.; Hsiao, S.-Y.; Tsai, C.-R.; Yu, S.-J.; Tsai, H.-J. Comprehensive Analysis of the Incidence and Survival Patterns of Lung Cancer by Histologies, Including Rare Subtypes, in the Era of Molecular Medicine and Targeted Therapy: A Nation-Wide Cancer Registry-Based Study from Taiwan. Medicine 2015, 94, e969. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Hood, B.L.; Sun, M.; Conrads, T.P.; Day, R.S.; Weissfeld, J.L.; Siegfried, J.M.; Bigbee, W.L. Lung Cancer Serum Biomarker Discovery Using Glycoprotein Capture and Liquid Chromatography Mass Spectrometry. J. Proteome Res. 2010, 9, 6440–6449. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.M.; Cho, W.; Li, B.; Prakobphol, A.; Johansen, E.; Anderson, N.L.; Regnier, F.E.; Gibson, B.W.; Fisher, S.J. Sweetening the Pot: Adding Glycosylation to the Biomarker Discovery Equation. Clin. Chem. 2010, 56, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Deng, Z.; Huang, C.; Zhu, T.; Lou, J.; Wang, L.; Li, Y. Differential N-Glycan Patterns Identified in Lung Adenocarcinoma by N-Glycan Profiling of Formalin-Fixed Paraffin-Embedded (FFPE) Tissue Sections. J. Proteom. 2018, 172, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Bao, X.; Chen, M.; Lin, R.; Zhuyan, J.; Zhen, T.; Xing, K.; Zhou, W.; Zhu, S. Mechanisms and Future of Non-Small Cell Lung Cancer Metastasis. Front. Oncol. 2020, 10, 585284. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, G.; Gagneux, P. N-Glycan Biosynthesis. Reactome 2010, 33. [Google Scholar] [CrossRef]
- Jupe, S. Laminin Interactions. Reactome 2013, 46. [Google Scholar] [CrossRef]
- Matthews, L. Cell Junction Organization. Reactome 2009, 31. [Google Scholar] [CrossRef]
- Geiger, B.; Horwitz, R. Integrin Cell Surface Interactions. Reactome 2008, 25. [Google Scholar] [CrossRef]
- Jupe, S. Non-Integrin Membrane-ECM Interactions. Reactome 2013, 45. [Google Scholar] [CrossRef]
- Dapic, I.; Baljeu-Neuman, L.; Uwugiaren, N.; Kers, J.; Goodlett, D.R.; Corthals, G.L. Proteome Analysis of Tissues by Mass Spectrometry. Mass Spectrom. Rev. 2019, 38, 403–441. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fermaintt, C.S.; Cherepanova, N.A.; Gilmore, R.; Yan, N.; Lehrman, M.A. Mammalian STT3A/B Oligosaccharyltransferases Segregate N-Glycosylation at the Translocon from Lipid-Linked Oligosaccharide Hydrolysis. Proc. Natl. Acad. Sci. USA 2018, 115, 9557–9562. [Google Scholar] [CrossRef] [PubMed]
- Jouannet, S.; Saint-Pol, J.; Fernandez, L.; Nguyen, V.; Charrin, S.; Boucheix, C.; Brou, C.; Milhiet, P.-E.; Rubinstein, E. TspanC8 Tetraspanins Differentially Regulate the Cleavage of ADAM10 Substrates, Notch Activation and ADAM10 Membrane Compartmentalization. Cell. Mol. Life Sci. 2016, 73, 1895–1915. [Google Scholar] [CrossRef] [PubMed]
- Seegar, T.C.M.; Killingsworth, L.B.; Saha, N.; Meyer, P.A.; Patra, D.; Zimmerman, B.; Janes, P.W.; Rubinstein, E.; Nikolov, D.B.; Skiniotis, G.; et al. Structural Basis for Regulated Proteolysis by the α-Secretase ADAM10. Cell 2017, 171, 1638–1648.e7. [Google Scholar] [CrossRef]
- Lemjabbar, H.; Basbaum, C. Platelet-Activating Factor Receptor and ADAM10 Mediate Responses to Staphylococcus Aureus in Epithelial Cells. Nat. Med. 2002, 8, 41–46. [Google Scholar] [CrossRef]
- Gutwein, P.; Mechtersheimer, S.; Riedle, S.; Stoeck, A.; Gast, D.; Joumaa, S.; Zentgraf, H.; Fogel, M.; Altevogt, P. ADAM10-mediated Cleavage of L1 Adhesion Molecule at the Cell Surface and in Released Membrane Vesicles. FASEB J. 2003, 17, 292–294. [Google Scholar] [CrossRef]
- Bendas, G.; Borsig, L. Cancer Cell Adhesion and Metastasis: Selectins, Integrins, and the Inhibitory Potential of Heparins. Int. J. Cell Biol. 2012, 2012, 676731. [Google Scholar] [CrossRef]
- Hamidi, H.; Ivaska, J. Every Step of the Way: Integrins in Cancer Progression and Metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Kdimati, S.; Mullins, C.S.; Linnebacher, M. Cancer-Cell-Derived IgG and Its Potential Role in Tumor Development. IJMS 2021, 22, 11597. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lin, D.; Peng, H.; Huang, Y.; Huang, J.; Gu, J. Cancer-Derived Immunoglobulin G Promotes Tumor Cell Growth and Proliferation through Inducing Production of Reactive Oxygen Species. Cell Death Dis. 2013, 4, e945. [Google Scholar] [CrossRef]
- Honma, R.; Kinoshita, I.; Miyoshi, E.; Tomaru, U.; Matsuno, Y.; Shimizu, Y.; Takeuchi, S.; Kobayashi, Y.; Kaga, K.; Taniguchi, N.; et al. Expression of Fucosyltransferase 8 Is Associated with an Unfavorable Clinical Outcome in Non-Small Cell Lung Cancers. Oncology 2015, 88, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Hoagland IV, L.F.M.; Campa, M.J.; Gottlin, E.B.; Herndon II, J.E.; Patz Jr, E.F. Haptoglobin and Posttranslational Glycan-Modified Derivatives as Serum Biomarkers for the Diagnosis of Nonsmall Cell Lung Cancer. Cancer 2007, 110, 2260–2268. [Google Scholar] [CrossRef] [PubMed]
- Jay, S.D.; Cherech, D.A. Integrins in Cancer: Biological Implications in Therapeutic Opportunities. Cancer Nat. Rev. 2015, 10, 9–22. [Google Scholar] [CrossRef]
- Zhao, Y.; Itoh, S.; Wang, X.; Isaji, T.; Miyoshi, E.; Kariya, Y.; Miyazaki, K.; Kawasaki, N.; Taniguchi, N.; Gu, J. Deletion of Core Fucosylation on Alpha3beta1 Integrin Down-Regulates Its Functions. J. Biol. Chem. 2006, 281, 38343–38350. [Google Scholar] [CrossRef]
- Cai, X.; Thinn, A.M.M.; Wang, Z.; Shan, H.; Zhu, J. The Importance of N-Glycosylation on Β3 Integrin Ligand Binding and Conformational Regulation. Sci. Rep. 2017, 7, 4656. [Google Scholar] [CrossRef]
- Janik, M.E.; Lityńska, A.; Vereecken, P. Cell Migration—The Role of Integrin Glycosylation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 545–555. [Google Scholar] [CrossRef]
- Marsico, G.; Russo, L.; Quondamatteo, F.; Pandit, A. Glycosylation and Integrin Regulation in Cancer. Trends Cancer 2018, 4, 537–552. [Google Scholar] [CrossRef]
- Pan, D.; Song, Y. Role of Altered Sialylation of the I-Like Domain of Β1 Integrin in the Binding of Fibronectin to Β1 Integrin: Thermodynamics and Conformational Analyses. Biophys. J. 2010, 99, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-B.; Lee, I.; Kamar, M.; Akiyama, S.K.; Pierce, M. Aberrant N-Glycosylation of β1 Integrin Causes Reduced α5β1 Integrin Clustering and Stimulates Cell Migration. Cancer Res. 2002, 62, 6837. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
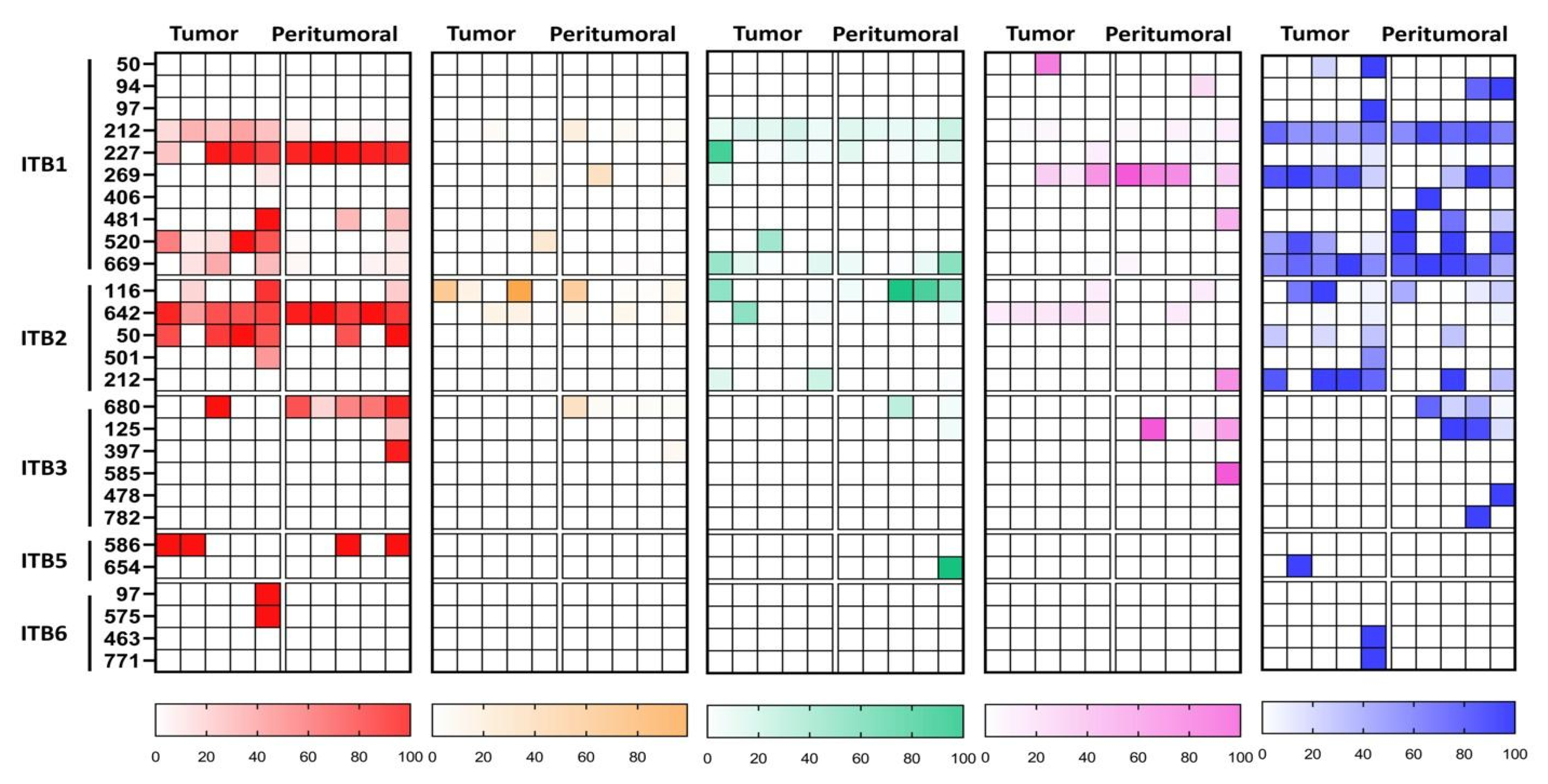{kind=link}
| Subject ID | Stage | Age/Sex | Mutation Status | 3-Gene Panel Expression | ||||
|---|---|---|---|---|---|---|---|---|
| TTF1 | ALK | EGFR | CD133 | KRT19 | MUC1 | |||
| AF63-009 | I-B | 63/F | Positive | Negative | Negative | Down | Down | Up |
| AM43-005 | I-B | 43/M | Positive | Negative | Negative | Up | Down | Down |
| AM51-009 | II-B | 51/M | Positive | Negative | Negative | Up | Down | Down |
| FDT-01 | II-B | 75/F | Positive | Negative | Positive (Exon18) | Up | Up | Down |
| AM53-021 | III-A | 53/M | Positive | Negative | Positive (Exon19) | No data | No data | No data |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, M.R.; Zhou, Q.; Tena, J.; Barboza, M.; Wong, M.; Xie, Y.; Lebrilla, C.B.; Cabanatan, M.; Barzaga, M.T.; Tan-Liu, N.; et al. Glycomic, Glycoproteomic, and Proteomic Profiling of Philippine Lung Cancer and Peritumoral Tissues: Case Series Study of Patients Stages I–III. Cancers 2023, 15, 1559. https://doi.org/10.3390/cancers15051559
Alvarez MR, Zhou Q, Tena J, Barboza M, Wong M, Xie Y, Lebrilla CB, Cabanatan M, Barzaga MT, Tan-Liu N, et al. Glycomic, Glycoproteomic, and Proteomic Profiling of Philippine Lung Cancer and Peritumoral Tissues: Case Series Study of Patients Stages I–III. Cancers. 2023; 15(5):1559. https://doi.org/10.3390/cancers15051559
Chicago/Turabian StyleAlvarez, Michael Russelle, Qingwen Zhou, Jennyfer Tena, Mariana Barboza, Maurice Wong, Yixuan Xie, Carlito B. Lebrilla, Michelle Cabanatan, Ma. Teresa Barzaga, Nelia Tan-Liu, and et al. 2023. "Glycomic, Glycoproteomic, and Proteomic Profiling of Philippine Lung Cancer and Peritumoral Tissues: Case Series Study of Patients Stages I–III" Cancers 15, no. 5: 1559. https://doi.org/10.3390/cancers15051559
APA StyleAlvarez, M. R., Zhou, Q., Tena, J., Barboza, M., Wong, M., Xie, Y., Lebrilla, C. B., Cabanatan, M., Barzaga, M. T., Tan-Liu, N., Heralde, F. M., III, Serrano, L., Nacario, R. C., & Completo, G. C. (2023). Glycomic, Glycoproteomic, and Proteomic Profiling of Philippine Lung Cancer and Peritumoral Tissues: Case Series Study of Patients Stages I–III. Cancers, 15(5), 1559. https://doi.org/10.3390/cancers15051559