Simple Summary
Pancreatic cancer has one of the worst prognoses among all the malignancies. To date, there are no effective strategies for preventing, diagnosing, or treating this aggressive disease. Increasing evidence shows that the human microbiome influences pancreatic cancer development and the response to treatment, which is the main aim of this review. Future studies will be key to determining whether or not this knowledge might be used to develop new strategies that harness the microbiome for diagnostic and therapeutic interventions in pancreatic cancer.
Abstract
Pancreatic cancer mortality is expected to rise in the next decades. This aggressive malignancy has a dismal prognosis due to late diagnosis and resistance to treatment. Increasing evidence indicates that host–microbiome interactions play an integral role in pancreatic cancer development, suggesting that harnessing the microbiome might offer promising opportunities for diagnostic and therapeutic interventions. Herein, we review the associations between pancreatic cancer and the intratumoral, gut and oral microbiomes. We also explore the mechanisms with which microbes influence cancer development and the response to treatment. We further discuss the potentials and limitations of using the microbiome as a target for therapeutic interventions, in order to improve pancreatic cancer patient outcomes.
1. Introduction
Pancreatic cancer ranks as the 12th most incident cancer and the seventh cause of cancer-related death worldwide, accounting for around 496,000 new cases and 466,000 deaths annually [1], with incidence and mortality being predicted to nearly duplicate in 2040 due to demographic changes [2].
Pancreatic cancer has a poor prognosis. The 5-year overall survival rate has increased by little with modern therapeutics, remaining as low as 5–10%, and remaining the worst among all types of cancers [3]. Pancreatic ductal adenocarcinoma (PDAC) accounts for more than 90% of pancreatic cancer diagnoses, arising from the ductal cells that constitute the exocrine portion of the pancreas. Many factors contribute to the overall bad prognosis of PDAC, including the absence of well-established risk factors and prevention strategies, the lack of a reliable screening tool, the aggressive biology of the tumor, the late diagnosis (generally made at the locally advanced or metastatic stage), and resistance to therapy.
Emerging evidence suggests that the microbiome influences pancreatic carcinogenesis, so identifying changes in the microbiome of patients with pancreatic cancer will likely set the stage for the development of novel screening, diagnostic, and therapeutic approaches for this deadly disease. In this review, we overview the current literature related to the microbiome and pancreatic cancer. A literature search was performed in the PubMed database, using a combination of terms, including “microbiome”, “microbiota”, “pancreatic cancer”, and “pancreatic ductal adenocarcinoma”, without restriction of the study design, publication date, or status. Studies not written in English were excluded. Additional studies were retrieved from the lists of references of selected articles. We aim to explore and summarize the latest findings on the mechanisms with which the microbiome modulates pancreatic cancer development and resistance to therapy. We further discuss potential therapeutic strategies to modulate the microbiome and briefly look into the challenges of future research on pancreatic cancer and the microbiome.
2. Pancreatic Cancer Risk Factors
PDAC is a paradigmatic example of an inflammation-driven cancer. Inflammation is both a risk factor for pancreatic cancer and also a consequence of carcinogenesis [4]. Oncogenic KRAS is the most common driver of mutation in human PDAC, being present in more than 90% of the cases [4]. There is a synergic relationship between KRAS activation and inflammation, as KRAS expression is itself highly proinflammatory, and the duration of pancreatitis is associated with the predisposition to KRAS mutations [5,6].
In addition to chronic pancreatitis, including hereditary and other forms, non-modifiable risk factors for PDAC include increasing age, familial cancer syndromes, being of the Afro-American race, and being of a non-O blood group [7]. Tobacco exposure and obesity are, so far, the only modifiable risk factors with convincing evidence to be considered well-established causes of pancreatic cancer [8]. A high-fat diet, diabetes [7], poor oral health status [9], and blood antibodies against selected oral pathogens [10,11] are also associated with an increased risk of pancreatic cancer. These risk factors are related with a state of chronic systemic inflammation that might be pro-tumorigenic, and are also associated with changes in the human microbiome that might be a common intermediate mechanistic step leading to pancreatic carcinogenesis [12].
3. The Human Microbiome
The human body is inhabited by a large collection of microorganisms that globally constitute the microbiota, and include bacteria, viruses, fungi, and protozoa [13]. The term microbiome generally refers to microorganisms, their genetic information, and the specific host environment which they inhabit, although currently the terms microbiota and microbiome are often used interchangeably [14]. The majority of the human microbiota reside in the gastrointestinal tract, skin and oral cavity, but organs previously considered to be sterile, such as the lung, the liver, and the pancreas, are now recognized as harboring low-biomass microbial populations. Whether microbiomes are natively established in these niches or represent the transient migration of microbes from adjacent sites is still unclear [13].
The composition of the normal microbiome varies significantly between individuals and between body sites, being influenced by local conditions, host genetics, diet, antibiotic consumption, and lifestyle [15]. Under healthy conditions, the commensal microorganisms symbiotically interact with the host across multiple body sites, playing an essential role in a variety of physiological processes. Disruption of the balance that exists between the microbiome and the host, called dysbiosis, may contribute to the pathogenesis of many diseases, including cancer [15].
4. The Microbiome in Pancreatic Cancer
4.1. The Intratumoral Microbiome
Cancer is classically considered a genetic disease, but tumors are much more than masses of abnormal proliferating cells [16]. Instead, the tumor microenvironment (TME) contains multiple distinct cell types that interact with one another in a complex way, contributing to the acquisition of hallmark traits that enable tumor growth and metastatic dissemination [16]. Since the microbiome is an intrinsic component of the tumor microenvironment, one may expect that the crosstalk between microbes and tumor cells is able to modulate oncogenesis.
Intratumoral bacteria were detected for the first time more than 100 years ago [17]. Currently there is strong evidence of an intratumoral bacterial microbiota across, at least, 33 major cancer types [18]. By directly analyzing the bacterial communities of tumors and their adjacent normal tissues in seven types of cancer, including the pancreas, Nejman et al. demonstrated that each tumor type has a distinct microbiota composition [19]. Consistently with these findings, predictive associations between different cancer types and specific microbiota profiles were recently reported by examining genome and transcriptome sequencing data of The Cancer Genome Atlas [18].
Several studies characterized the tumor microbiota in pancreatic cancer (Table 1).

Table 1.
Summary of studies evaluating the tumor microbiota in pancreatic cancer.
Geller et al. compared pancreatic tumor tissues and normal pancreatic tissues and identified bacterial DNA in 76% of the tumors and in 15% of the normal tissues [24]. They identified Gammaproteobacteria as the most common taxa, including members of the Enterobacteriaceae and Pseudomonadaceae, with Pseudomonas, Citrobacter, Klebsiella, Streptococcus, and Acinetobacter as genera with the highest mean relative abundances. These data are consistent with those reported by Nejman et al. showing that more than 60% of pancreatic tumors were positive for bacterial DNA, predominantly belonging to the phyla Proteobacteria and Firmicutes, with a high prevalence of Enterobacter asburiae, Klebsiella pneumoniae, Citrobacter freundii, and Fusobacterium nucleatum [19]. These findings suggest a bacterial reflux from the duodenum to the pancreas via the major/minor papilla and the pancreatic duct, as Proteobacteria also dominate the normal duodenal microbiome [19,24]. Accordingly, it was also observed that bacterial DNA profiles were similar between the pancreas and duodenum of the same subjects, which further supports that bacteria migrate from the gut into the pancreas [25]. Nejman and colleagues also showed that the microbial populations present in the tumor tissues of different cancer types are similar to those of their normal adjacent tissues [19], leading to the hypothesis that the tumor microbiome evolves from microbes existing in normal tissues [26]. In contrast, Riquelme et al. found that the human gut microbiome represents approximately 25% of the PDAC microbiome, while it is absent in the normal adjacent tissue, suggesting that gut microbes specifically colonize pancreatic tumors [23]. Additional studies evaluating the intratumoral microbiota reported tumor enrichment in Lactobacillus spp., Akkermansia muciniphila, and Bacteroides [20], and Pseudomonas, Herbaspirillum, and Sphingomonas [21] in comparison with adjacent normal tissue.
It remains unknown how bacteria may colonize the pancreas, but studies with fluorescently labeled bacteria administered via oral gavage to mice revealed bacteria migration to the pancreas [27]. Inflammatory conditions associated with pancreatic cancer, such as pancreatitis and obesity, may induce gut dysbiosis and alter gut permeability, thus increasing microbial translocation to the pancreas [28].
As with the bacteriome, fungal dysbiosis has been identified in cancer and studies have linked fungi to pancreatic carcinogenesis [29]. Aykut et al. showed that PDAC tumors in both humans and mouse models display a 3000-fold increase in fungi compared to the normal pancreas, with higher mycobiome diversity and enrichment in Malassezia [30]. The authors demonstrated that fluorescently labeled fungi translocated from the gut lumen to the pancreatic tissue. Fungal ablation was protective against tumor development, while repopulation with Malassezia—but not Candida, Aspergillus, or Saccharomyces—accelerated oncogenesis. Malassezia was shown to promote pancreatic tumorigenesis via the activation of mannose-binding lectin and the subsequent activation of the complement cascade [30], which stimulates cell growth, survival and migration [31]. Fungal dysbiosis of the digestive tract is often characterized by an overgrowth of Candida albicans, which was independently associated with a higher PDAC risk in two large population-based cohorts in Sweden [32] and in Taiwan [33]. A valuable next step will be to determine whether or not the mycobiome interacts with the bacteriome to promote PDAC progression [31].
Clinical disease heterogeneity exists in pancreatic cancer, and since limited genomic differences have been found [34], the tumor microbiota has been studied as a possible factor that may determine disease prognosis. Pancreatic tumors located in the head of the pancreas are generally easier to detect, so their overall prognosis is better than that of pancreatic body/tail cancers. Recently, prognostic and genetic differences between pancreatic head and body/tail cancers were identified [35], but no differences in intratumoral bacteria diversity or relative abundance were detected between anatomical locations [36].
Long-term and short-term PDAC survivors have different intratumoral microbiota structures (Table 1), with higher bacterial diversity being associated with longer survival [22,23]. In two independent PDAC patient cohorts from the USA, Riquelme et al. identified an intratumoral microbial signature of Pseudoxanthomonas, Streptomyces, Saccharopolyspora, and Bacillus clausii, which was predictive of long-term survivorship [23]. Huang et al. identified an enrichment in Sphingomonas, Megasphaera, Bradyrhizobium, Desulfovibrio, Flavobacterium, Enhydrobacter, and Megamonas in long-term-surviving PDAC patients of Chinese origin [22]. Patients with high relative abundances of Sphingomonas and Megasphaera had longer overall survival, whereas patients with a high abundance of Clostridium had shorter survival. Fusobacterium nucleatum, an anaerobic Gram-negative species of the oral microbiome implicated in colorectal cancer, has also been identified as a prevalent species in pancreatic cancer [19] and associated with worse prognosis [37]. Notably, Pseudomonas, a common opportunistic pathogenic bacterium, was also observed to be enriched in PDAC patients with short-term survival [23] and was identified as the most abundant genus that translocates from the gut to the pancreas in PDAC patients [27].
Taken together, these data show that the composition of a tumor’s bacterial microbiota in pancreatic cancer is highly variable, although microbial signatures might be useful to predict pancreatic cancer prognosis. Once microbial biomarker signatures are identified both in pancreatic cysts or tumors, microbial phenotyping via endoscopic ultrasound might emerge as a new approach to screen, stratify the risk of and select therapeutics for pancreatic cancer [36]. Whether intratumoral bacteria play a causal role in pancreatic carcinogenesis or simply infect tumors already established is still not clear.
4.2. The Gut and the Oral Microbiomes in Pancreatic Cancer
Multiple studies in the last decade have explored the association of the human oral and gut microbiomes with pancreatic cancer, as these may represent non-invasive approaches to early cancer diagnosis (Table 2).
Table 2.
Summary of studies evaluating the gut and oral microbiota in pancreatic cancer.
4.2.1. The Gut Microbiome and Pancreatic Cancer
An expanding body of evidence has associated gut microbiome changes with pancreatic cancer. Studies investigating the link between the gut microbiome and pancreatic cancer have shown that the structure of the gut microbial communities of pancreatic cancer patients is clearly distinct from that of healthy controls, as evaluated by beta diversity analysis [38,41,42,43]. However, reports are inconsistent regarding the gut microbial alpha diversity in pancreatic cancer patients, with studies showing higher, similar, or lower diversity compared to that of healthy controls. Overall, microbial biomarker species have high accuracy to distinguish PDAC patients from healthy controls [20,38,39,41,42,43]. In the multinational study of Nagata and colleagues, dysbiosis of the gut microbiome was reported and significant associations were identified between PDAC and 30 gut bacterial species [38]. These bacterial signatures had high levels of accuracy in predicting PDAC in the Japanese discovery and in the Spanish and German independent patient cohorts. A common PDAC gut microbial signature in all these cohorts was the enrichment in Streptococcus and Veillonella spp. and depletion of Faecalibacterium prausnitzii.
Kartal et al. showed that fecal microbiome classifiers based on 27 bacterial species were better than those of the saliva microbiome in the identification of PDAC patients, in Spanish and German cohorts [20]. Others have shown an enrichment in pro-inflammatory genera, especially those belonging to the phylum Proteobacteria in the gut microbiome of PDAC patients [27,41]. In a Chinese cohort comparing the fecal microbiome of PDAC patients with that of matched healthy controls, the gut microbial community of PDAC patients had a higher abundance of lipopolysaccharide-producing bacteria, and a reduction in beneficial microbes, such as butyrate-producing bacteria [43]. Accordingly, significant reduction in Firmicutes, especially of butyrate-producing bacteria, was also observed in the gut microbiome of PDAC patients [41]. Taxa consistently identified as enriched in the gut microbiome of pancreatic cancer patients include Veillonella [20,27,38,40,41,42,43], Streptococcus [24,38,39,40], Actinomyces [22,38,40,47], Lactobacillus [20,39,40], and Klebsiella [27,39,43]. Taxa depleted in the gut microbiome of pancreatic cancer patients include Faecalibacterium [20,38,41,42], Bifidobacterium [20,39,43], Eubacterium [38,39,41], Anaerostipes [40,42,43], Blautia [39,43], Coprococcus [39,43], and Clostridiales [38,42]. Still, there is high microbial variability between individuals, both within and across different populations. The combination of fecal microbial patterns with additional biomarkers may be necessary to increase accuracy in pancreatic cancer detection, as shown by Kartal et al. who combined fecal microbiome classifiers with serum levels of CA 19-9 to improve accuracy in the identification of PDAC patients [20].
The association between the duodenal microbiome and pancreatic cancer has been scarcely explored, but it has been hypothesized that changes in the duodenal microenvironment might contribute to a higher incidence of pancreatic head cancer compared to pancreatic body/tail cancers [52]. The microbiome of different pancreatic sites appears to be similar to the duodenum’s microbiome of the same subjects [25]. Recently, it was shown that patients with PDAC have alterations in their duodenal fluid microbial profiles, showing higher levels of bacterial and fungal DNA, reduced microbial diversity and an enrichment in Bifidobacterium [53]. Additionally, duodenal fluid samples from short-term PDAC survivors were enriched with bacteria belonging to the genera Fusobacterium, Rothia, and Neisseria. Microbial profiling of duodenal fluid acquired via endoscopy to screen and stratify the risk of pancreatic cancer may deserve further attention. Factors that influence duodenal fluid composition, such as the use of proton-pump inhibitors, must be considered. The role of Helicobacter pylori in PDAC remains controversial [54], but it is possible that infection of the gastric body mucosa with subsequent hypochlorhydria may be associated with duodenal fluid alterations.
Compared to the bacterial populations in the human gut, the enteric mycobiome has low diversity (dominated by yeasts such as Saccharomyces, Malassezia, and Candida), with high intra-individual variability over time [55]. In contrast, viruses in the gut are highly diverse with limited intra-individual variability over time [56]. While some individual viruses have causal roles in carcinogenesis [57], the relationship between the gut virome and pancreatic cancer is far from being understood. A study in Japan reported differences in the gut virome structure between pancreatic cancer patients and healthy controls [39]. The gut virome of pancreatic cancer patients was enriched with Cardiovirus, Alphabaculovirus, and Proboscivirus, whereas 16 other viral genera were more common in the gut of healthy subjects [39]. The relative abundance of Cyprinivirus was positively correlated with the cancer stage [39]. The enteric virome consists largely of bacteriophages that infect specific bacterial species and help maintain a healthy gut microbiome [58]. Nagata et al. found 58 bacteriophages that potentially infect microbial species that are consistently enriched in the gut microbiome of patients with PDAC (Streptococcus oralis, Streptococcus parasanguinis, Veillonella atypica, and Veillonella parvula) [38]. Synergistic pathogenic interactions may exist, in which virome dysbiosis leads to the emergence and persistence of opportunistic bacteria that are able to modulate the development of cancer [29]. The characterization of the gut phageome will be fundamental before considering phage therapy as an approach to treat pancreatic cancer.
Despite the potential use of gut microbial signatures for cancer diagnosis, it remains unclear whether gut dysbiosis is a result or rather causally triggers the development of pancreatic cancer.
4.2.2. The Oral Microbiome and Pancreatic Cancer
As for the gut microbiome, several reports have analyzed the link between the oral microbiome and pancreatic cancer (Table 2). The majority of studies, including those with a larger sample size, observed no differences in alpha diversity between cancer patients and controls [44,46,49,50], despite some reporting increased alpha diversity in the oral microbiota of pancreatic cancer patients [38,45]. Furthermore, while some studies show that the overall structure of the oral microbiome in pancreatic cancer patients can be distinguished from that of heathy control subjects [38,45,46], others do not identify such differences [48,49,50]. Still, there is supportive evidence that the oral microbiome plays a role in pancreatic cancer etiology, some of which stemmed from the hypothesis that the relationship between periodontal disease and pancreatic cancer is related to changes in the oral microbiota. A large study by Fan and colleagues, which analyzed oral wash samples from patients in the USA, identified two pathogens involved in chronic periodontitis, Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans, as being associated with increased risk of pancreatic cancer [48]. These results are in agreement with those of previous studies showing that patients with high levels of plasma antibodies against P. gingivalis were at higher risk of PDAC [10,11]. However, in a study of 273 pancreatic cancer cases and 285 controls from Iran, Porphyromonas was detected in similar proportions in cases and controls, and was not associated with pancreatic cancer [46]. Interestingly, in this study, the authors reported an increased abundance of Haemophilus being associated with decreased odds of pancreatic cancer, which corroborated previous findings in a population from the USA [49].
Farrell et al. identified various bacterial species that were significantly different between saliva samples of pancreatic cancer patients and healthy controls, including Neisseria elongata and Streptococcus mitis [51]. These two species, which were depleted in pancreatic cancer, were then validated in an independent cohort of patients and showed high sensitivity and specificity in distinguishing the two groups [51]. Other studies consistently report a decrease in the abundance of Neisseria in the oral microbiota of pancreatic cancer patients [45,47,49]. Depletion of various species of Streptococcus within a bacterial signature of 18 species was also identified in the oral microbiota of PDAC patients, which had high accuracy in predicting disease in Japanese, Spanish, and German patient cohorts [38].
In contrast to the above, other studies point to similar overall profiles in the oral microbiota of healthy controls and patients with pancreatic cancer, with no major differences in the abundance of taxa [20,44,49]. Whether oral microbiome signatures in pancreatic cancer patients represent risk factors for cancer development or are a result of the presence of cancer remains to be determined.
5. Mechanisms Linking the Microbiome to Pancreatic Cancer
The microbiome, both local and remote, can impact cancer development through a variety of mechanisms. These include the stimulation of oncogenic signaling pathways, the dampening of tumor suppressor mechanisms, the induction of DNA damage, the promotion of chronic inflammation, and the modulation of the tumor immune environment [59,60,61,62,63]. Microorganisms may influence carcinogenesis by binding to, invading, or injecting virulence factors into the host target cell, by secreting toxins and microbial metabolites, or by metabolizing host and dietary metabolites [13].
To evaluate the microbial metabolites affecting pancreatic carcinogenesis, Mendez et al. performed gut microbiome and metabolome analyses in a genetically engineered PDAC mouse model [64]. They showed that PDAC progression was accompanied by changes in the gut microbial population and, by characterizing the metabolome of the involved bacterial species, identified metabolites involved in polyamine metabolism [64]. The findings of elevated serum polyamine levels as tumors progressed in mice were also confirmed in human PDAC patients [64]. Other microbe-derived metabolites may have protective effects in cancer development, such as butyrate and pyridoxine [65]. Butyrate, a short-chain fatty acid (SCFA) produced by certain bacteria through the fermentation of non-digestible carbohydrates, was described to possess anti-inflammatory and anti-neoplastic properties [66]. Specifically, in the context of pancreatic cancer, butyrate was found to have “pro-differentiating, anti-proliferative, anti-invasive, pro-apoptotic” and chemo-sensitizing effects in PDAC cell lines in vitro [66]. Sodium butyrate also prevents the activation of pancreatic stellate cells, reducing the desmoplastic reaction in PDAC [67].
As mentioned above, a plausible explanation for the procarcinogenic effect of the microbiome involves chronic and low-grade activation of the immune system, which can perpetuate tumor-associated inflammation and immunosuppression [4,68]. It is likely that a persistent inflammatory state induced by microbial dysbiosis will result in excessive exposure to oxidative stress, leading to genetic instability and increasing the risk of tumorigenesis [69].
Since KRAS mutant signaling per se is not sufficient to initiate invasive PDAC, microbe-induced inflammation may potentiate pancreatic oncogenic signaling [70]. The local microbial activation of pattern recognition receptors, including toll-like receptor 4 (TLR4) [71], TLR7 [72], NOD-like receptor (NLRP3) inflammasome [73], and Dectin [74], may also potentiate mutant KRAS signaling and accelerate pancreatic oncogenesis. The activation of TLR2 and TLR5 was shown to suppress both innate and adaptive immunity in early and advanced PDAC [27]. Thus, the peri-pancreatic inflammatory response induced by the intratumoral microbiome may contribute not only to genetic oncogenic events, but also to helping shape an immunosuppressive microenvironment that promotes pancreatic cancer development [70]. Microorganisms remote from the pancreas, such as the gut and the oral microbiomes, may promote pancreatic carcinogenesis by triggering a systemic inflammatory response, or by reaching the pancreas (either through the bloodstream or the gastrointestinal tract) and locally modulate inflammation and the immune response [75].
Pancreatic cancer thrives in a profoundly immunosuppressive tumor microenvironment, which partially accounts for its aggressive biology. Tumors are characterized by a highly fibrotic stroma, often containing infiltration by pro-tumoral immune suppressor cells, including tumor-associated macrophages (TAMs), regulatory T cells (Tregs), and myeloid-derived suppressor cells (MDSCs) [29]. Cancer-associated fibroblasts (CAFs) and myofibroblast-like pancreatic stellate cells (PSCs) produce the collagen that composes the rich desmoplastic stroma of pancreatic tumors. TLR9 bacteria-induced stimulation may contribute to PSC activation [76]. Balachandran et al. demonstrated that tumors from long-term survivors of pancreatic cancer have a high quantity and diversity of neoantigens, which are associated with more abundant CD8+ T cell infiltrates [77]. Interestingly, the neoantigens exhibited homology to peptides derived from infectious agents, which is consistent with the existence of molecular mimicry between tumor neoantigens and microbial epitopes [77]. These data suggest that microbes might determine tumor immunogenicity and patient outcomes. Although PDAC tumors express neoantigens that can be recognized by T cells [78], there are adaptive immune suppression mechanisms that result in the inhibition of T cells’ cytotoxic activity, despite their presence in the tumor milieu [79].
Intratumoral bacteria are mainly found in the cytoplasm of tumor and immune cells [19]. They play a significant role in modulating relevant functions related to cancer hallmarks and immune activity, including cell motility, extracellular matrix interaction, complement cascade, and PD-1 signaling [19,80]. Interestingly, the tumor microbiome appears to have a dual effect, as it can promote either immune suppression or activation. Using genetically modified mouse models, Pushalkar et al. reported that bacteria induce intratumoral immune suppression mediated by TLR ligation, contributing to pancreatic cancer progression [27]. Bacterial ablation protected against PDAC progression and was associated with immunogenic reprogramming of the PDAC microenvironment, inducing an increase in intratumoral T cells and antitumoral M1 macrophages, along with a decrease in pro-tumoral MDSCs and M2 macrophages [27]. However, several studies suggested that the tumor microbiome might influence pancreatic cancer outcomes by activating the immune response. According to a recent study by Ghaddar et al., a high tumoral bacterial load was associated with decreased patient survival rates, high levels of tumor infiltration, T cell activation, and upregulation of PD-1-signaling pathways in both activated T cells and malignant cells [80]. Riquelme et al. also found that the tumor microbiome favors the recruitment and activation of CD8+ T cells into the tumor milleu, thus promoting the immune response against the tumor [23].
Overall, and in addition to identifying intratumoral populations of bacteria, fungi, and viruses, it is also important to understand the mechanisms through which these microorganisms interact with each other and with host cells in PDAC. Such knowledge could provide unique opportunities to improve clinical outcomes for patients with PDAC. For instance, understanding the role of viruses in the tumor microenvironment may enable the identification of phages that can selectively target and deplete tumor-promoting bacteria, instead of using non-selective antibiotics [68].
6. The Microbiome and Treatment Responses in Pancreatic Cancer
It is becoming increasingly evident that the microbiome has an impact on the efficacy of cancer treatment. However, the way the microbiome interacts with therapeutics is bidirectional and dynamic [14]. On the one hand, the microbiome can affect drug metabolism and absorption, modulating the efficacy and side effects of PDAC therapy [70], and on the other hand, drugs can shape the microbiome of the host.
6.1. Chemotherapy
Gemcitabine is a chemotherapeutic drug commonly used to treat PDAC. It has been shown that gemcitabine can be metabolized to its inactive form by the cytidine deaminase enzyme of Gammaproteobacteria, which are highly prevalent bacteria in human PDAC [24]. In a mouse model of colon carcinoma, gemcitabine-induced apoptosis was significantly enhanced when antibiotics were directly delivered into the tumor, suggesting a role of intratumoral bacteria in gemcitabine resistance [24]. However, gut dysbiosis induced by gemcitabine may paradoxically inhibit the drug’s anti-cancer effect and promote inflammation both locally and systemically. In a PDAC xenograft mouse model treated with gemcitabine, there was an overall increase in pro-inflammatory bacteria in the gut, with a reduction in Lachnospiraceae and Ruminococcaceae, two bacterial families known for their ability to generate butyrate [81]. Treatment with gemcitabine led to a significant reduction in serum levels of the anti-inflammatory compound inosine and its metabolites xanthine and hypoxanthine, which is likely associated with the observed dysbiosis [81].
Another chemotherapeutic option for treating advanced PDAC is the FOLFIRINOX regimen (5-FU, leucovorin, irinotecan, and oxaliplatin) [82]. Reactive oxygen species (ROS) are important for platinum-mediated cytotoxicity. The gut microbiome can increase oxaliplatin-induced ROS production, thus enhancing the efficacy of chemotherapy [83]. However, the gut microbiome may also increase the toxicity of chemotherapeutic drugs, thus limiting the effective dose that can be delivered [75]. An example is irinotecan-induced diarrhea promoted by an increase in β-glucuronidase-producing bacteria [84]. Further studies are needed to determine the impact of these drugs on the human gut microbiome and on PDAC prognosis in the clinical setting.
Neoadjuvant chemotherapy (NAC) is being employed more commonly in PDAC, not only to select and potentially increase the number of patients who can benefit from surgical resection, but also to reduce the chances of early recurrence and to improve peri-operative outcomes. Most patients receiving NAC undergo pre-operative biliary stenting, and accumulating evidence shows that these patients have a different bile microbiome compared to that of those undergoing primary surgery, with potential prognostic relevance [85]. Pre-operative biliary draining has been associated with higher risk of post-operative surgical site infections (SSIs), which is believed to be related to the introduction of intestinal bacteria into the biliary tree after stenting [85]. In a retrospective cohort, Nadeem et al. observed that NAC significantly alters the biliary microbiome of patients undergoing pancreaticoduodenectomy, as those undergoing NAC showed an increased growth of gram-negative anaerobes and an overall decrease in antibiotic resistance amongst biliary bacteria, particularly with the cephalosporin class of antibiotics [86]. These results are in contrast to those of another study in which resistance to cephalosporins was more common in patients who received NAC [85], which highlights the need to account for potential confounders in order to clarify the real impact of NAC in the biliary microbiome and thus tailor perioperative antibiotic prophylaxis. Postoperative pancreatic fistulas (POPFs) remain one of the most frequent and dangerous complications after pancreatic surgery, with common bacterial contamination resulting in higher morbidity and mortality [87]. While NAC appears to significantly reduce the incidence of POPFs [87], pre-operative biliary stenting may be an important risk factor for infectious complications after surgery [86]. Furthermore, a recent study demonstrated that the human bile reduces pancreatic cancer cell survival in vitro and peritoneal metastasis in vivo, with secondary bile acids showing more antitumor effects than primary bile acids do [88]. The anti-tumorigenic property of bile was reduced following pre-operative biliary stenting, presumably due to changes in the biliary microbiome [88].
6.2. Immunotherapy
Immunotherapy with immune checkpoint inhibitors, such as anti-PD-1, anti-PD-L1, and anti-CTLA-4, has shown remarkable success in several solid malignancies, but it has been ineffective for pancreatic cancer thus far, with the possible exception of very rare tumors with high microsatellite instability [89]. Commensal gut bacteria were found to positively influence the efficacy of PD-1-based immunotherapy in different types of epithelial tumors, with non-responders showing an imbalance in gut microbial composition that correlated with impaired immune cell activity [90]. Bifidobacterium, one of the major genera in the human gut microbiome, is believed to enhance the efficacy of the immune checkpoint blockers [91]. In several studies using mouse models of PDAC, it was observed that antibiotic-induced ablation of the gut microbiome may increase the sensitivity of the tumor to immune checkpoint inhibitors and reduce tumor burden [27,92,93]. Pushalkar et al. found that microbiome depletion promotes immunogenic reprogramming of the tumor microenvironment towards an activation phenotype, with increased PD-1 expression in intratumoral effector T cells [27]. Altogether, these studies suggest that antibiotic therapy combined with immune checkpoint blockers represents an attractive new strategy for treating patients with PDAC.
7. Microbiome Modulation in Pancreatic Cancer
Most patients with pancreatic cancer present with advanced unresectable disease, with treatment relying mainly on gemcitabine-based palliative chemotherapy. New therapeutic strategies have shown inconsistent results in PDAC, including neoantigen vaccines [94] and genomic targeted therapies [95]. Modulation of the host microbiome may represent a promising adjunct to current treatments for pancreatic cancer, as it may change the tumor microenvironment and sensitize tumors to therapeutics [96].
7.1. Diet
Changes in diet constitute an attractive approach to shape the microbial populations of the gut. A high-fat diet induces gut dysbiosis characterized by a reduction in the abundance of Bacteroidetes and an increase in the amount of Firmicutes and Proteobacteria, as well as gut inflammation and increased permeability, with the possible translocation of bacteria and its metabolites into the circulation [97]. Conversely, a high-fiber diet may protect against cancer, as it increases the microbial production of butyrate and other anti-inflammatory and pro-apoptotic SCFAs [98]. Engineered resistant starch (ERS) has emerged as a new strategy to enhance butyrate production and improve mucosal integrity [99]. Panebianco et al. found that an ERS diet retards tumor growth in a PDAC xenograft mouse model, by exerting a positive influence in the composition and metabolism of the gut microbiome [100]. Prospective clinical trials are necessary to determine how enriching a diet with nondigestible fibers may help to prevent and treat pancreatic cancer. Questions regarding the cost, acceptability, and potential adverse effects of these dietary interventions, such as gastrointestinal intolerance and enhanced cancer-associated cachexia, should be considered [13].
7.2. Probiotics
Probiotics are defined as “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [101]. The range of microorganisms with potential health benefits is increasing in parallel to the accumulating knowledge on the human microbiome [96]. Probiotics have been suggested as an approach to cancer prevention and treatment, but in multiple cases have yielded conflicting results [13]. Gut dysbiosis in PDAC human patients is often characterized by a decrease in the abundance of butyrate-producing bacteria [41,43]. Regarding the aforementioned benefits of butyrate, accompanying conventional chemotherapeutic treatments with butyrate supplementation [66] and/or SCFAs-producing bacteria (e.g., Faecalibacterium prausnitzii) [102] may help to enhance clinical outcomes in pancreatic cancer. In addition, some probiotics can ameliorate the adverse effects of cancer therapies, such as chemotherapy-induced mucositis, thereby avoiding treatment discontinuation/dose reduction and increasing the chances of cure. In PDAC xenografted mice, the administration of probiotics enhanced the efficacy of gemcitabine while reducing chemotherapy-related toxicity [103]. As gemcitabine significantly alters the gut microbiome, and microbial changes may be detrimental for its efficacy, combined strategies using probiotics to restore a favorable gut microbiome may help in enhancing its effectiveness in the treatment of PDAC [81]. The administration of microorganisms associated with long-term cancer survival might also emerge as a new therapeutic strategy in PDAC [96]. Large clinical trials are required to develop precision probiotics that could address the heterogeneous microbiome and clinical features observed in different individuals [13].
7.3. Antibiotics
The use of antibiotics for microbial eradication in cancer is controversial, being currently recommended only for H. pylori erradication in gastric carcinoma and MALT lymphoma [13]. Preclinical studies have suggested a potential positive role of antibiotics in the treatment of PDAC, showing a reduction in tumor progression [27,92,93], and an improved response to anti-PD-1 immunotherapy [27] and to gemcitabine [24]. Antibiotic usage in metastatic PDAC was linked to increased survival in patients receiving gemcitabine-based chemotherapy, possibly due to a positive modulation of the microbiome, as well as a limitation of local and systemic infections [104]. Antifungal drugs were also found to slow PDAC progression and improve response to chemotherapy [30]. Long-term antibiotic treatments can have deleterious effects, such as drug toxicity, systemic microbial dysbiosis [14], the emergence of resistant strains, unforeseeable impacts on disease course and treatment efficacy [13], and possibly an increased risk of cancer [105]. In addition, by decreasing the abundance of commensal bacteria, broad-spectrum antibiotics can further exacerbate fungal colonization and infection [106]. Thus, strategies aiming to eliminate specific bacteria, rather than relatively untargeted approaches, might emerge as promising adjuncts to conventional cancer therapies [13]. One of such strategies may be phage therapy. Bacteriophages (or phages) are ubiquitous viruses that only infect bacteria, and have been recognized as promising tools to target specific disease-associated bacteria, while bearing minimal impacts on the surrounding microbiome. Alternatively, phages can be used as cargo particles to directly deliver drugs within the tumor microenvironment. Future research is needed to overcome the challenges that currently limit the widespread applicability of phage therapy [13,107].
7.4. Fecal Microbiota Transplantation
Fecal microbiota transplantation (FMT) refers to the transfer of the entire microbiota from a healthy individual into the intestine of a patient, aiming to restore the homeostasis of the gut microbiome [108]. In the last years, FMT has been investigated for the treatment of many intestinal and systemic disorders; notably, recurrent Clostridium difficile infections. Regarding cancer, FMT was shown to be a reasonable approach to mitigate the adverse effects of cancer treatment, such as colitis, and improve the efficacy of immune checkpoint inhibitors in melanoma [108]. Riquelme et al. found that the stools from PDAC patients with long-term survival harbor a specific bacterial signature that can alter the intratumoral microbiome and reduce tumor growth in fecally transplanted mice [23]. These findings suggest that FMT might be a way to modulate the microbiome in patients with PDAC, but future clinical trials are needed to validate these results. Importantly, FMT presents with several clinical, scientific, and regulatory uncertainties that need to be addressed before it can be routinely adopted in clinical practice [13]. For instance, effective donor and recipient selection, different regulatory policies across countries and institutions, and excessive costs may limit the widespread applicability of FMT. A future goal must be the identification of specific bacteria and microbial metabolites that drive the response to FMT, in the aim to develop personalized strategies to modulate the composition of the gut microbiome [13].
8. Concluding Remarks
The aggressive biology of pancreatic cancer remains poorly understood, but research on the microbiome has revealed that pancreatic cancer tissues harbor a distinct microbial community. Additionally, changes in the gut and oral microbiomes have been associated with pancreatic cancer progression and the response to treatment (Figure 1). However, consistent microbial signatures associated with pancreatic cancer features have not been disclosed. In addition to the low number of cancer patients included in some studies, factors such as the genetic background and geographic origins of the patients, as well as lifestyle habits and environmental exposure, may partially explain the differences observed between studies. Furthermore, differences in methods of sample collection, processing, analyzing, and microbiomic reporting may generate additional variation.
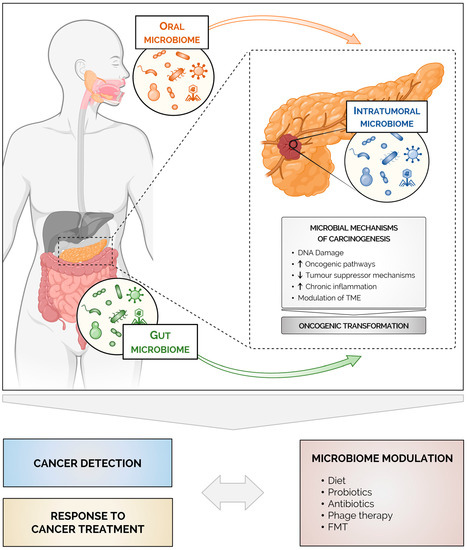
Figure 1.
Microbes as an emerging tool for pancreatic cancer diagnosis and treatment. The microbiome influences pancreatic carcinogenesis both locally (intratumoral microbiome) and at a distance (gut and oral microbiomes). Mechanistically, microbially induced inflammation, metabolism and modulation of the tumor microenvironment contribute to the development of pancreatic cancer and to the overall poor response to chemotherapy and immunotherapy. Microbial signatures might be useful as non-invasive approaches for early detection and for defining the prognosis of pancreatic cancer. Ultimately, microbiome modulation may represent a promising adjunct to current treatments for pancreatic cancer. Created with BioRender.com.
It needs to be highlighted that although bacteria are the most studied component of the microbiome in cancer, future research is needed to expand our understanding on how fungi and viruses interact with the bacterial microbiome and how they may influence pancreatic cancer development and treatment. Nevertheless, one of the major challenges in this area of research is to evolve from the identification of associations towards finding causality, since changes in the microbiome of pancreatic cancer patients may represent a risk factor for pancreatic cancer, but may also be the result of the overall cancer-induced disturbance in host homeostasis. Evidence from animal studies together with large prospective clinical studies will be fundamental to elucidating plausible mechanistic links and to definitely clarify whether microbial signatures are causative or the result of pancreatic cancer progression. Pancreatic cancer microbiome research will likely generate new knowledge, which may be translated into enhanced strategies for screening, diagnosis, and treatment of this deadly disease, eventually helping to reduce its burden.
Author Contributions
Conceptualization: C.F.; literature screening and data acquisition: A.R.B., R.M.F. and C.F.; data analysis and interpretation: A.R.B., J.P.-M., R.M.F. and C.F.; writing—original draft: A.R.B. and C.F.; writing—critical review and editing: all authors; tables and figures: J.P.-M., R.M.F. and C.F.; supervision: C.F. All authors have read and agreed to the published version of the manuscript.
Funding
R.M.F. has a Fundação para a Ciência e a Tecnologia (FCT) researcher position under the Individual Call to Scientific Employment Stimulus (CEECIND/01854/2017). The team is also funded by national funds through FCT (PTDC/BTM-TEC/0367/2021 and 2022.02141.PTDC).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Laversanne, M.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I. Global Cancer Observatory: Cancer Tomorrow. Lyon, France: International Agency for Research on Cancer. 2020. Available online: https://gco.iarc.fr/tomorrow (accessed on 16 November 2022).
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic Cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Zambirinis, C.P.; Pushalkar, S.; Saxena, D.; Miller, G. Pancreatic Cancer, Inflammation, and Microbiome. Cancer J. 2014, 20, 195–202. [Google Scholar] [CrossRef]
- Guerra, C.; Schuhmacher, A.J.; Cañamero, M.; Grippo, P.J.; Verdaguer, L.; Pérez-Gallego, L.; Dubus, P.; Sandgren, E.P.; Barbacid, M. Chronic Pancreatitis Is Essential for Induction of Pancreatic Ductal Adenocarcinoma by K-Ras Oncogenes in Adult Mice. Cancer Cell 2007, 11, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Löhr, M.; Klöppel, G.; Maisonneuve, P.; Lowenfels, A.B.; Lüttges, J. Frequency of K-Ras Mutations in Pancreatic Intraductal Neoplasias Associated with Pancreatic Ductal Adenocarcinoma and Chronic Pancreatitis: A Meta-Analysis. Neoplasia 2005, 7, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Midha, S.; Chawla, S.; Garg, P.K. Modifiable and Non-Modifiable Risk Factors for Pancreatic Cancer: A Review. Cancer Lett. 2016, 381, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Carreras-Torres, R.; Johansson, M.; Gaborieau, V.; Haycock, P.C.; Wade, K.H.; Relton, C.L.; Martin, R.M.; Davey Smith, G.; Brennan, P. The Role of Obesity, Type 2 Diabetes, and Metabolic Factors in Pancreatic Cancer: A Mendelian Randomization Study. J. Natl. Cancer Inst. 2017, 109, djx012. [Google Scholar] [CrossRef]
- Hujoel, P.P.; Drangsholt, M.; Spiekerman, C.; Weiss, N.S. An Exploration of the Periodontitis-Cancer Association. Ann. Epidemiol. 2003, 13, 312–316. [Google Scholar] [CrossRef]
- Michaud, D.S.; Izard, J.; Wilhelm-Benartzi, C.S.; You, D.-H.; Grote, V.A.; Tjønneland, A.; Dahm, C.C.; Overvad, K.; Jenab, M.; Fedirko, V.; et al. Plasma Antibodies to Oral Bacteria and Risk of Pancreatic Cancer in a Large European Prospective Cohort Study. Gut 2013, 62, 1764–1770. [Google Scholar] [CrossRef]
- Ahn, J.; Segers, S.; Hayes, R.B. Periodontal Disease, Porphyromonas Gingivalis Serum Antibody Levels and Orodigestive Cancer Mortality. Carcinogenesis 2012, 33, 1055–1058. [Google Scholar] [CrossRef]
- Tijeras-Raballand, A.; Hilmi, M.; Astorgues-Xerri, L.; Nicolle, R.; Bièche, I.; Neuzillet, C. Microbiome and Pancreatic Ductal Adenocarcinoma. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101589. [Google Scholar] [CrossRef] [PubMed]
- Cullin, N.; Azevedo Antunes, C.; Straussman, R.; Stein-Thoeringer, C.K.; Elinav, E. Microbiome and Cancer. Cancer Cell 2021, 39, 1317–1341. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Gao, H.-L.; Wang, W.-Q.; Yu, X.-J.; Liu, L. Bidirectional and Dynamic Interaction between the Microbiota and Therapeutic Resistance in Pancreatic Cancer. Biochim. Biophys. acta. Rev. cancer 2021, 1875, 188484. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current Understanding of the Human Microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Glover, T. Progress in Cancer Research. Canada Lancet Pract. 1926, 67, 161–216. [Google Scholar]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome Analyses of Blood and Tissues Suggest Cancer Diagnostic Approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The Human Tumor Microbiome Is Composed of Tumor Type-Specific Intracellular Bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef]
- Kartal, E.; Schmidt, T.S.B.; Molina-Montes, E.; Rodríguez-Perales, S.; Wirbel, J.; Maistrenko, O.M.; Akanni, W.A.; Alashkar Alhamwe, B.; Alves, R.J.; Carrato, A.; et al. A Faecal Microbiota Signature with High Specificity for Pancreatic Cancer. Gut 2022, 71, 1359–1372. [Google Scholar] [CrossRef]
- Tan, Q.; Ma, X.; Yang, B.; Liu, Y.; Xie, Y.; Wang, X.; Yuan, W.; Ma, J. Periodontitis Pathogen Porphyromonas Gingivalis Promotes Pancreatic Tumorigenesis via Neutrophil Elastase from Tumor-Associated Neutrophils. Gut Microbes 2022, 14, 2073785. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, N.; Zheng, X.; Liu, Y.; Lu, H.; Yin, X.; Hao, H.; Tan, Y.; Wang, D.; Hu, H.; et al. Intratumor Microbiome Analysis Identifies Positive Association Between Megasphaera and Survival of Chinese Patients with Pancreatic Ductal Adenocarcinomas. Front. Immunol. 2022, 13, 785422. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; San Lucas, A.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef] [PubMed]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential Role of Intratumor Bacteria in Mediating Tumor Resistance to the Chemotherapeutic Drug Gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef]
- Del Castillo, E.; Meier, R.; Chung, M.; Koestler, D.C.; Chen, T.; Paster, B.J.; Charpentier, K.P.; Kelsey, K.T.; Izard, J.; Michaud, D.S. The Microbiomes of Pancreatic and Duodenum Tissue Overlap and Are Highly Subject Specific but Differ between Pancreatic Cancer and Noncancer Subjects. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Hu, Y. Microbiome Harbored within Tumors: A New Chance to Revisit Our Understanding of Cancer Pathogenesis and Treatment. Signal Transduct. Target. Ther. 2020, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Sethi, V.; Vitiello, G.A.; Saxena, D.; Miller, G.; Dudeja, V. The Role of the Microbiome in Immunologic Development and Its Implication For Pancreatic Cancer Immunotherapy. Gastroenterology 2019, 156, 2097–2115.e2. [Google Scholar] [CrossRef]
- Elaskandrany, M.; Patel, R.; Patel, M.; Miller, G.; Saxena, D.; Saxena, A. Fungi, Host Immune Response, and Tumorigenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G213–G222. [Google Scholar] [CrossRef]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The Fungal Mycobiome Promotes Pancreatic Oncogenesis via Activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Dambuza, I.M.; Brown, G.D. Fungi Accelerate Pancreatic Cancer. Nature 2019, 574, 184–185. [Google Scholar] [CrossRef]
- Huang, J.; Roosaar, A.; Axéll, T.; Ye, W. A Prospective Cohort Study on Poor Oral Hygiene and Pancreatic Cancer Risk. Int. J. Cancer 2016, 138, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.-M.; Liang, J.-A.; Lin, C.-L.; Sun, L.-M.; Kao, C.-H. Cancer Risk in Patients with Candidiasis: A Nationwide Population-Based Cohort Study. Oncotarget 2017, 8, 63562–63573. [Google Scholar] [CrossRef] [PubMed]
- Makohon-Moore, A.P.; Zhang, M.; Reiter, J.G.; Bozic, I.; Allen, B.; Kundu, D.; Chatterjee, K.; Wong, F.; Jiao, Y.; Kohutek, Z.A.; et al. Limited Heterogeneity of Known Driver Gene Mutations among the Metastases of Individual Patients with Pancreatic Cancer. Nat. Genet. 2017, 49, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Xiao, L.; Gao, Y.; Wang, G.; Gao, H.; Peng, Y.; Zhu, X.; Wei, J.; Miao, Y.; Jiang, K.; et al. Comparative Bioinformatical Analysis of Pancreatic Head Cancer and Pancreatic Body/Tail Cancer. Med. Oncol. 2020, 37, 46. [Google Scholar] [CrossRef]
- Gleeson, F.C.; Jeraldo, P.; Levy, M.J.; Murphy, S.J.; Mendes-Soares, H.; Karagouga, G.; Mccune, A.F.; Garcia Garcia Deparedes, A.; Kipp, B.R.; Song, S.D.; et al. Composition, Diversity and Potential Utility of Intervention-Naïve Pancreatic Cancer Intratumoral Microbiome Signature Profiling via Endoscopic Ultrasound. Gut 2022, 71, 441–443. [Google Scholar] [CrossRef]
- Mitsuhashi, K.; Nosho, K.; Sukawa, Y.; Matsunaga, Y.; Ito, M.; Kurihara, H.; Kanno, S.; Igarashi, H.; Naito, T.; Adachi, Y.; et al. Association of Fusobacterium Species in Pancreatic Cancer Tissues with Molecular Features and Prognosis. Oncotarget 2015, 6, 7209–7220. [Google Scholar] [CrossRef]
- Nagata, N.; Nishijima, S.; Kojima, Y.; Hisada, Y.; Imbe, K.; Miyoshi-Akiyama, T.; Suda, W.; Kimura, M.; Aoki, R.; Sekine, K.; et al. Metagenomic Identification of Microbial Signatures Predicting Pancreatic Cancer From a Multinational Study. Gastroenterology 2022, 163, 222–238. [Google Scholar] [CrossRef]
- Matsukawa, H.; Iida, N.; Kitamura, K.; Terashima, T.; Seishima, J.; Makino, I.; Kannon, T.; Hosomichi, K.; Yamashita, T.; Sakai, Y.; et al. Dysbiotic Gut Microbiota in Pancreatic Cancer Patients Form Correlation Networks with the Oral Microbiota and Prognostic Factors. Am. J. Cancer Res. 2021, 11, 3163–3175. [Google Scholar] [PubMed]
- Hashimoto, S.; Tochio, T.; Funasaka, K.; Funahashi, K.; Hartanto, T.; Togashi, Y.; Saito, M.; Nishimoto, Y.; Yoshinori, M.; Nakaoka, K.; et al. Changes in Intestinal Bacteria and Imbalances of Metabolites Induced in the Intestines of Pancreatic Ductal Adenocarcinoma Patients in a Japanese Population: A Preliminary Result. Scand. J. Gastroenterol. 2023, 58, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhang, D.; Li, Z.; Jiang, H.; Li, J.; Ren, R.; Gao, X.; Li, J.; Wang, X.; Wang, W.; et al. The Fecal Microbiota of Patients with Pancreatic Ductal Adenocarcinoma and Autoimmune Pancreatitis Characterized by Metagenomic Sequencing. J. Transl. Med. 2021, 19, 215. [Google Scholar] [CrossRef]
- Half, E.; Keren, N.; Reshef, L.; Dorfman, T.; Lachter, I.; Kluger, Y.; Reshef, N.; Knobler, H.; Maor, Y.; Stein, A.; et al. Fecal Microbiome Signatures of Pancreatic Cancer Patients. Sci. Rep. 2019, 9, 16801. [Google Scholar] [CrossRef]
- Ren, Z.; Jiang, J.; Xie, H.; Li, A.; Lu, H.; Xu, S.; Zhou, L.; Zhang, H.; Cui, G.; Chen, X.; et al. Gut Microbial Profile Analysis by MiSeq Sequencing of Pancreatic Carcinoma Patients in China. Oncotarget 2017, 8, 95176–95191. [Google Scholar] [CrossRef]
- Petrick, J.L.; Wilkinson, J.E.; Michaud, D.S.; Cai, Q.; Gerlovin, H.; Signorello, L.B.; Wolpin, B.M.; Ruiz-Narváez, E.A.; Long, J.; Yang, Y.; et al. The Oral Microbiome in Relation to Pancreatic Cancer Risk in African Americans. Br. J. Cancer 2022, 126, 287–296. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, X.; Zhou, Y.; Wang, J.; Ma, R.; Ren, X.; Wang, H.; Zou, L. Characterization of Oral Microbiome and Exploration of Potential Biomarkers in Patients with Pancreatic Cancer. Biomed Res. Int. 2020, 2020, 4712498. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, E.; Han, Y.; Caporaso, J.G.; Bokulich, N.; Mohamadkhani, A.; Moayyedkazemi, A.; Hua, X.; Kamangar, F.; Wan, Y.; Suman, S.; et al. Oral Microbial Community Composition Is Associated with Pancreatic Cancer: A Case-Control Study in Iran. Cancer Med. 2020, 9, 797–806. [Google Scholar] [CrossRef]
- Wei, A.-L.; Li, M.; Li, G.-Q.; Wang, X.; Hu, W.-M.; Li, Z.-L.; Yuan, J.; Liu, H.-Y.; Zhou, L.-L.; Li, K.; et al. Oral Microbiome and Pancreatic Cancer. World J. Gastroenterol. 2020, 26, 7679–7692. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Alekseyenko, A.V.; Wu, J.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Abnet, C.C.; Stolzenberg-Solomon, R.; Miller, G.; et al. Human Oral Microbiome and Prospective Risk for Pancreatic Cancer: A Population-Based Nested Case-Control Study. Gut 2018, 67, 120–127. [Google Scholar] [CrossRef]
- Olson, S.H.; Satagopan, J.; Xu, Y.; Ling, L.; Leong, S.; Orlow, I.; Saldia, A.; Li, P.; Nunes, P.; Madonia, V.; et al. The Oral Microbiota in Patients with Pancreatic Cancer, Patients with IPMNs, and Controls: A Pilot Study. Cancer Causes Control 2017, 28, 959–969. [Google Scholar] [CrossRef]
- Torres, P.J.; Fletcher, E.M.; Gibbons, S.M.; Bouvet, M.; Doran, K.S.; Kelley, S.T. Characterization of the Salivary Microbiome in Patients with Pancreatic Cancer. PeerJ 2015, 3, e1373. [Google Scholar] [CrossRef]
- Farrell, J.J.; Zhang, L.; Zhou, H.; Chia, D.; Elashoff, D.; Akin, D.; Paster, B.J.; Joshipura, K.; Wong, D.T.W. Variations of Oral Microbiota Are Associated with Pancreatic Diseases Including Pancreatic Cancer. Gut 2012, 61, 582–588. [Google Scholar] [CrossRef]
- Mei, Q.-X.; Huang, C.-L.; Luo, S.-Z.; Zhang, X.-M.; Zeng, Y.; Lu, Y.-Y. Characterization of the Duodenal Bacterial Microbiota in Patients with Pancreatic Head Cancer vs. Healthy Controls. Pancreatol. Off. J. Int. Assoc. Pancreatol. 2018, 18, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Kohi, S.; Macgregor-Das, A.; Dbouk, M.; Yoshida, T.; Chuidian, M.; Abe, T.; Borges, M.; Lennon, A.M.; Shin, E.J.; Canto, M.I.; et al. Alterations in the Duodenal Fluid Microbiome of Patients With Pancreatic Cancer. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2022, 20, e196–e227. [Google Scholar] [CrossRef] [PubMed]
- Panthangi, V.; Cyril Kurupp, A.R.; Raju, A.; Luthra, G.; Shahbaz, M.; Almatooq, H.; Foucambert, P.; Esbrand, F.D.; Zafar, S.; Khan, S. Association Between Helicobacter Pylori Infection and the Risk of Pancreatic Cancer: A Systematic Review Based on Observational Studies. Cureus 2022, 14, e28543. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The Gut Mycobiome of the Human Microbiome Project Healthy Cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Shkoporov, A.N.; Clooney, A.G.; Sutton, T.D.S.; Ryan, F.J.; Daly, K.M.; Nolan, J.A.; McDonnell, S.A.; Khokhlova, E.V.; Draper, L.A.; Forde, A.; et al. The Human Gut Virome Is Highly Diverse, Stable, and Individual Specific. Cell Host Microbe 2019, 26, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Zella, D.; Gallo, R.C. Viruses and Bacteria Associated with Cancer: An Overview. Viruses 2021, 13, 1039. [Google Scholar] [CrossRef]
- Seo, S.-U.; Kweon, M.-N. Virome-Host Interactions in Intestinal Health and Disease. Curr. Opin. Virol. 2019, 37, 63–71. [Google Scholar] [CrossRef]
- Abreu, M.T.; Peek, R.M.J. Gastrointestinal Malignancy and the Microbiome. Gastroenterology 2014, 146, 1534–1546. [Google Scholar] [CrossRef]
- Cuevas-Ramos, G.; Petit, C.R.; Marcq, I.; Boury, M.; Oswald, E.; Nougayrède, J.-P. Escherichia Coli Induces DNA Damage In Vivo and Triggers Genomic Instability in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11537–11542. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, K.; Wier, E.M.; Lei, Y.; Hodgson, A.; Xu, D.; Xia, X.; Zheng, D.; Ding, H.; Sears, C.L.; et al. Bacterial Genotoxin Accelerates Transient Infection-Driven Murine Colon Tumorigenesis. Cancer Discov. 2022, 12, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, A.; Wang, Y.; Zhang, Y. Intratumoral Microbiota: Roles in Cancer Initiation, Development and Therapeutic Efficacy. Signal Transduct. Target. Ther. 2023, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Mendez, R.; Kesh, K.; Arora, N.; Di Martino, L.; McAllister, F.; Merchant, N.; Banerjee, S.; Banerjee, S. Microbial Dysbiosis and Polyamine Metabolism as Predictive Markers for Early Detection of Pancreatic Cancer. Carcinogenesis 2020, 41, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Daillère, R.; Roberti, M.P.; Routy, B.; Kroemer, G. Anticancer Effects of the Microbiome and Its Products. Nat. Rev. Microbiol. 2017, 15, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Villani, A.; Pisati, F.; Orsenigo, F.; Ulaszewska, M.; Latiano, T.P.; Potenza, A.; Andolfo, A.; Terracciano, F.; Tripodo, C.; et al. Butyrate, a Postbiotic of Intestinal Bacteria, Affects Pancreatic Cancer and Gemcitabine Response in in Vitro and in Vivo Models. Biomed. Pharmacother. 2022, 151, 113163. [Google Scholar] [CrossRef] [PubMed]
- Kanika, G.; Khan, S.; Jena, G. Sodium Butyrate Ameliorates L-Arginine-Induced Pancreatitis and Associated Fibrosis in Wistar Rat: Role of Inflammation and Nitrosative Stress. J. Biochem. Mol. Toxicol. 2015, 29, 349–359. [Google Scholar] [CrossRef]
- Leinwand, J.; Miller, G. Regulation and Modulation of Antitumor Immunity in Pancreatic Cancer. Nat. Immunol. 2020, 21, 1152–1159. [Google Scholar] [CrossRef]
- Morgillo, F.; Dallio, M.; Della Corte, C.M.; Gravina, A.G.; Viscardi, G.; Loguercio, C.; Ciardiello, F.; Federico, A. Carcinogenesis as a Result of Multiple Inflammatory and Oxidative Hits: A Comprehensive Review from Tumor Microenvironment to Gut Microbiota. Neoplasia 2018, 20, 721–733. [Google Scholar] [CrossRef]
- Vitiello, G.A.; Cohen, D.J.; Miller, G. Harnessing the Microbiome for Pancreatic Cancer Immunotherapy. Trends in cancer 2019, 5, 670–676. [Google Scholar] [CrossRef]
- Ochi, A.; Nguyen, A.H.; Bedrosian, A.S.; Mushlin, H.M.; Zarbakhsh, S.; Barilla, R.; Zambirinis, C.P.; Fallon, N.C.; Rehman, A.; Pylayeva-Gupta, Y.; et al. MyD88 Inhibition Amplifies Dendritic Cell Capacity to Promote Pancreatic Carcinogenesis via Th2 Cells. J. Exp. Med. 2012, 209, 1671–1687. [Google Scholar] [CrossRef]
- Ochi, A.; Graffeo, C.S.; Zambirinis, C.P.; Rehman, A.; Hackman, M.; Fallon, N.; Barilla, R.M.; Henning, J.R.; Jamal, M.; Rao, R.; et al. Toll-like Receptor 7 Regulates Pancreatic Carcinogenesis in Mice and Humans. J. Clin. Investig. 2012, 122, 4118–4129. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Pandian, G.S.D.B.; Savadkar, S.; Lee, K.B.; Torres-Hernandez, A.; Aykut, B.; Diskin, B.; et al. NLRP3 Signaling Drives Macrophage-Induced Adaptive Immune Suppression in Pancreatic Carcinoma. J. Exp. Med. 2017, 214, 1711–1724. [Google Scholar] [CrossRef] [PubMed]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Ochi, A.; Heindel, D.W.; Lee, K.B.; Zambirinis, C.P.; Pandian, G.S.D.B.; Savadkar, S.; et al. Dectin 1 Activation on Macrophages by Galectin 9 Promotes Pancreatic Carcinoma and Peritumoral Immune Tolerance. Nat. Med. 2017, 23, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Jobin, C.; Thomas, R.M. Implications of the Microbiome in the Development and Treatment of Pancreatic Cancer: Thinking Outside of the Box by Looking inside the Gut. Neoplasia 2021, 23, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Zambirinis, C.P.; Levie, E.; Nguy, S.; Avanzi, A.; Barilla, R.; Xu, Y.; Seifert, L.; Daley, D.; Greco, S.H.; Deutsch, M.; et al. TLR9 Ligation in Pancreatic Stellate Cells Promotes Tumorigenesis. J. Exp. Med. 2015, 212, 2077–2094. [Google Scholar] [CrossRef]
- Balachandran, V.P.; Łuksza, M.; Zhao, J.N.; Makarov, V.; Moral, J.A.; Remark, R.; Herbst, B.; Askan, G.; Bhanot, U.; Senbabaoglu, Y.; et al. Identification of Unique Neoantigen Qualities in Long-Term Survivors of Pancreatic Cancer. Nature 2017, 551, 512–516. [Google Scholar] [CrossRef]
- Bailey, P.; Chang, D.K.; Forget, M.-A.; Lucas, F.A.S.; Alvarez, H.A.; Haymaker, C.; Chattopadhyay, C.; Kim, S.-H.; Ekmekcioglu, S.; Grimm, E.A.; et al. Exploiting the Neoantigen Landscape for Immunotherapy of Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2016, 6, 35848. [Google Scholar] [CrossRef]
- Neesse, A.; Bauer, C.A.; Öhlund, D.; Lauth, M.; Buchholz, M.; Michl, P.; Tuveson, D.A.; Gress, T.M. Stromal Biology and Therapy in Pancreatic Cancer: Ready for Clinical Translation? Gut 2019, 68, 159–171. [Google Scholar] [CrossRef]
- Ghaddar, B.; Biswas, A.; Harris, C.; Omary, M.B.; Carpizo, D.R.; Blaser, M.J.; De, S. Tumor Microbiome Links Cellular Programs and Immunity in Pancreatic Cancer. Cancer Cell 2022, 40, 1240–1253.e5. [Google Scholar] [CrossRef]
- Panebianco, C.; Adamberg, K.; Jaagura, M.; Copetti, M.; Fontana, A.; Adamberg, S.; Kolk, K.; Vilu, R.; Andriulli, A.; Pazienza, V. Influence of Gemcitabine Chemotherapy on the Microbiota of Pancreatic Cancer Xenografted Mice. Cancer Chemother. Pharmacol. 2018, 81, 773–782. [Google Scholar] [CrossRef]
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.-L.; Choné, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef] [PubMed]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Stringer, A.M.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Yeoh, A.S.J.; Keefe, D.M.K. Faecal Microflora and Beta-Glucuronidase Expression Are Altered in an Irinotecan-Induced Diarrhea Model in Rats. Cancer Biol. Ther. 2008, 7, 1919–1925.e6. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Nadler, A.; Reddy, S.; Hoffman, J.P.; Pitt, H.A. Biliary Microbiome in Pancreatic Cancer: Alterations with Neoadjuvant Therapy. HPB Off. J. Int. Hepato Pancreato Biliary Assoc. 2019, 21, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, S.O.; Jajja, M.R.; Maxwell, D.W.; Pouch, S.M.; Sarmiento, J.M. Neoadjuvant Chemotherapy for Pancreatic Cancer and Changes in the Biliary Microbiome. Am. J. Surg. 2021, 222, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hank, T.; Sandini, M.; Ferrone, C.R.; Rodrigues, C.; Weniger, M.; Qadan, M.; Warshaw, A.L.; Lillemoe, K.D.; Fernández-Del Castillo, C. Association Between Pancreatic Fistula and Long-Term Survival in the Era of Neoadjuvant Chemotherapy. JAMA Surg. 2019, 154, 943–951. [Google Scholar] [CrossRef]
- Shrader, H.R.; Miller, A.M.; Tomanek-Chalkley, A.; McCarthy, A.; Coleman, K.L.; Ear, P.H.; Mangalam, A.K.; Salem, A.K.; Chan, C.H.F. Effect of Bacterial Contamination in Bile on Pancreatic Cancer Cell Survival. Surgery 2021, 169, 617–622. [Google Scholar] [CrossRef]
- Hilmi, M.; Bartholin, L.; Neuzillet, C. Immune Therapies in Pancreatic Ductal Adenocarcinoma: Where Are We Now? World J. Gastroenterol. 2018, 24, 2137–2151. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut Microbiome Influences Efficacy of PD-1-Based Immunotherapy against Epithelial Tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti-PD-L1 Efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Sethi, V.; Kurtom, S.; Tarique, M.; Lavania, S.; Malchiodi, Z.; Hellmund, L.; Zhang, L.; Sharma, U.; Giri, B.; Garg, B.; et al. Gut Microbiota Promotes Tumor Growth in Mice by Modulating Immune Response. Gastroenterology 2018, 155, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.M.; Gharaibeh, R.Z.; Gauthier, J.; Beveridge, M.; Pope, J.L.; Guijarro, M.V.; Yu, Q.; He, Z.; Ohland, C.; Newsome, R.; et al. Intestinal Microbiota Enhances Pancreatic Carcinogenesis in Preclinical Models. Carcinogenesis 2018, 39, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, V.P.; Beatty, G.L.; Dougan, S.K. Broadening the Impact of Immunotherapy to Pancreatic Cancer: Challenges and Opportunities. Gastroenterology 2019, 156, 2056–2072. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Gong, Y.; Fan, Z.; Luo, G.; Huang, Q.; Deng, S.; Cheng, H.; Jin, K.; Ni, Q.; Yu, X.; et al. Molecular Alterations and Targeted Therapy in Pancreatic Ductal Adenocarcinoma. J. Hematol. Oncol. 2020, 13, 130. [Google Scholar] [CrossRef]
- Brandi, G.; Turroni, S.; McAllister, F.; Frega, G. The Human Microbiomes in Pancreatic Cancer: Towards Evidence-Based Manipulation Strategies? Int. J. Mol. Sci. 2021, 22, 9914. [Google Scholar] [CrossRef]
- Malesza, I.J.; Malesza, M.; Walkowiak, J.; Mussin, N.; Walkowiak, D.; Aringazina, R.; Bartkowiak-Wieczorek, J.; Mądry, E. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells 2021, 10, 3164. [Google Scholar] [CrossRef]
- Bultman, S.J. Molecular Pathways: Gene-Environment Interactions Regulating Dietary Fiber Induction of Proliferation and Apoptosis via Butyrate for Cancer Prevention. Clin. cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 799–803. [Google Scholar] [CrossRef]
- DeMartino, P.; Cockburn, D.W. Resistant Starch: Impact on the Gut Microbiome and Health. Curr. Opin. Biotechnol. 2020, 61, 66–71. [Google Scholar] [CrossRef]
- Panebianco, C.; Adamberg, K.; Adamberg, S.; Saracino, C.; Jaagura, M.; Kolk, K.; Di Chio, A.G.; Graziano, P.; Vilu, R.; Pazienza, V. Engineered Resistant-Starch (ERS) Diet Shapes Colon Microbiota Profile in Parallel with the Retardation of Tumor Growth in In Vitro and In Vivo Pancreatic Cancer Models. Nutrients 2017, 9, 331. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Sobocki, B.K.; Kaźmierczak-Siedlecka, K.; Folwarski, M.; Hawryłkowicz, V.; Makarewicz, W.; Stachowska, E. Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations. Nutrients 2021, 13, 4425. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Pisati, F.; Ulaszewska, M.; Andolfo, A.; Villani, A.; Federici, F.; Laura, M.; Rizzi, E.; Potenza, A.; Latiano, T.P.; et al. Tuning Gut Microbiota through a Probiotic Blend in Gemcitabine-Treated Pancreatic Cancer Xenografted Mice. Clin. Transl. Med. 2021, 11, e580. [Google Scholar] [CrossRef] [PubMed]
- Mohindroo, C.; Hasanov, M.; Rogers, J.E.; Dong, W.; Prakash, L.R.; Baydogan, S.; Mizrahi, J.D.; Overman, M.J.; Varadhachary, G.R.; Wolff, R.A.; et al. Antibiotic Use Influences Outcomes in Advanced Pancreatic Adenocarcinoma Patients. Cancer Med. 2021, 10, 5041–5050. [Google Scholar] [CrossRef] [PubMed]
- Boursi, B.; Mamtani, R.; Haynes, K.; Yang, Y.-X. Recurrent Antibiotic Exposure May Promote Cancer Formation--Another Step in Understanding the Role of the Human Microbiota? Eur. J. Cancer 2015, 51, 2655–2664. [Google Scholar] [CrossRef]
- Galloway-Peña, J.R.; Kontoyiannis, D.P. The Gut Mycobiome: The Overlooked Constituent of Clinical Outcomes and Treatment Complications in Patients with Cancer and Other Immunosuppressive Conditions. PLoS Pathog. 2020, 16, e1008353. [Google Scholar] [CrossRef]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; Del Fiol, F.S.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef]
- Derosa, L.; Routy, B.; Desilets, A.; Daillère, R.; Terrisse, S.; Kroemer, G.; Zitvogel, L. Microbiota-Centered Interventions: The Next Breakthrough in Immuno-Oncology? Cancer Discov. 2021, 11, 2396–2412. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).