
Immune Microenvironment in Childhood Cancers: Characteristics and Therapeutic Challenges

Department of Biochemistry and Molecular Biology, The Fred and Pamela Buffett Cancer Center, University of Nebraska Medical Center, Omaha, NE 68198, USA
Cancers 2024, 16(12), 2201; https://doi.org/10.3390/cancers16122201
Submission received: 9 February 2024
/
Revised: 23 May 2024
/
Accepted: 5 June 2024
/
Published: 12 June 2024
(This article belongs to the Collection Oncology: State-of-the-Art Research in the USA)
Abstract
:Simple Summary
The immune microenvironment surrounding a growing tumor is complex and diverse. Understanding this complexity is crucial for predicting prognosis and response to cancer therapy. Various immune cells, signaling molecules, and extracellular matrix components within this microenvironment interact intricately, influencing tumor progression and treatment effectiveness. A deeper understanding of these interactions can lead to better prognostic markers and more effective, personalized therapeutic strategies. Compared to adults, the pediatric tumor immune microenvironment is less studied. Research on different pediatric tumors has shown that regulatory signaling networks between tumors and surrounding immune cells are critical in shifting the balance between pro- and anti-tumor immune responses. Therefore, gaining insights into the tumor immune composition is key to improving the efficacy of various pediatric cancer therapies, particularly immunotherapies that enhance host anti-tumor immunity.
Abstract
The tumor immune microenvironment is pivotal in cancer initiation, advancement, and regulation. Its molecular and cellular composition is critical throughout the disease, as it can influence the balance between suppressive and cytotoxic immune responses within the tumor’s vicinity. Studies on the tumor immune microenvironment have enriched our understanding of the intricate interplay between tumors and their immunological surroundings in various human cancers. These studies illuminate the role of significant components of the immune microenvironment, which have not been extensively explored in pediatric tumors before and may influence the responsiveness or resistance to therapeutic agents. Our deepening understanding of the pediatric tumor immune microenvironment is helping to overcome challenges related to the effectiveness of existing therapeutic strategies, including immunotherapies. Although in the early stages, targeted therapies that modulate the tumor immune microenvironment of pediatric solid tumors hold promise for improved outcomes. Focusing on various aspects of tumor immune biology in pediatric patients presents a therapeutic opportunity that could improve treatment outcomes. This review offers a comprehensive examination of recent literature concerning profiling the immune microenvironment in various pediatric tumors. It seeks to condense research findings on characterizing the immune microenvironment in pediatric tumors and its impact on tumor development, metastasis, and response to therapeutic modalities. It covers the immune microenvironment’s role in tumor development, interactions with tumor cells, and its impact on the tumor’s response to immunotherapy. The review also discusses challenges targeting the immune microenvironment for pediatric cancer therapies.
1. Introduction
Despite cancer being considered a rare disease in children due to its low prevalence, it is the second leading cause of death in children aged 1 to 14. According to the World Health Organization (WHO), an estimated 400,000 children and adolescents aged 0 to 19 years develop cancer each year. Among this demographic, blood cancers reign as the most prevalent, with leukemias comprising 36.1% of cases in children aged 0–4 years and 15.4% in those aged 15–19 years, while lymphomas account for 5.3% and 22.5%, respectively, in the same age groups [1,2]. Following leukemia, central nervous system (CNS) tumors make up roughly 17.2% of cases in children aged 0–4 years, 26.3% in those aged 5–9 years, and 20.0% in those aged 10–14 years.
Subsequently, sympathetic nervous system tumors like neuroblastoma are most common in the 0–4 years age group (12.5%), followed by renal tumors (8.9% in 0–4 years but less prevalent in 15–19 years), and bone tumors (4.7% in children aged 0–14 years, and 7.8% in those aged 15–19 years). Adolescents aged 15–19 years exhibit a heightened incidence of epithelial tumors, melanoma, and lymphoma [1]. Data from United States (US) Cancer Statistics spanning from 2003 to 2019 show that the overall cancer incidence rate among children and adolescents under 20 years old was 178.3 per 1 million, with the highest rates observed in leukemia (46.6), CNS neoplasms (30.8), and lymphoma (27.3) [3].
Despite an overall increase in pediatric cancer incidence, the death rate has declined by more than half from 1970 to 2021 and remains significantly lower than that of adults with cancer [4,5]. According to WHO’s 2020 report, the most frequently diagnosed adult cancers globally are breast (2.26 million cases), lung (2.21 million cases), colon and rectum (1.93 million cases), and prostate (1.41 million cases) [6]. The majority of cancer-related deaths are attributed to lung (1.80 million), colon and rectum (916,000), liver (830,000), stomach (769,000), and breast (685,000) [6]. In the US, breast, lung, prostate, and colorectal cancers comprise almost 50% of new cases, while lung, colorectal, pancreatic, and breast cancers make up nearly 50% of cancer-related deaths [5].
Pediatric cancers differ significantly from adult cancers, possessing distinct characteristics rather than simply being miniature versions of adult tumors. A comprehensive pan-cancer DNA study on approximately 1700 pediatric cancer patients, including those with acute lymphoblastic leukemia (ALL), acute myeloid leukemia (AML), neuroblastoma, Wilms (kidney) tumor, and osteosarcoma, revealed significant differences in somatic DNA alterations between childhood and adult tumors [7]. Compared to adult tumors, childhood tumors exhibit a lower mutational burden and commonly show somatic DNA alterations, such as deletions, duplications, single-nucleotide variants (SNVs), or rearrangements, which are more specific to the particular histotype. Additionally, the report highlights the identification of 142 driver genes in pediatric cancers, of which only 45% overlap with those found in adult pan-cancer studies [7].
Contrary to adult tumors, which exhibit multiple-driver genetic alterations, pediatric cancers are enriched with single-driver mutations [8]. These mutations are more specific to individual cancer types and show minimal overlap across different pediatric cancers [7,8]. However, the presence of inherited germline mutations is less than 10% in both children and adolescents with cancer [8,9]. Like adult tumors, many pediatric ALL, neuroblastoma, and osteosarcoma, show increased mutations in their relapsed state, making them resistant to anticancer drugs [10,11,12]. This suggests that selective mutational pressure plays a significant role in their survival and conferring resistance to the drugs.
Advancements in DNA sequencing technologies in recent years have greatly contributed to our understanding of both solid and hematological malignancies in children. Comprehensive studies employing whole-genome and exome sequencing have uncovered notable differences in driver genes affecting essential cell proliferation pathways like RAS, Janus kinase/signal transducer and activator of transcription (JAK-STAT), and Phosphoinositide 3-Kinases (PI3K) signaling among pediatric cancer populations [7]. For example, somatic alterations in genes like anaplastic lymphoma kinase (ALK), neurofibromatosis type 1(NF1), and phosphatase and tensin homolog (PTEN) primarily contribute to pathway deregulation in solid tumors, whereas in leukemias, mutations in fms-like tyrosine kinase 3 (FLT3), phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha (PIK3CA) or p110α, phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1), and RAS genes are more common.
Additionally, specific mutations exhibit varying prevalence between solid and blood cancers, highlighting their distinct genetic profiles [7]. For instance, two KRAS isoforms are common in leukemias, accounting for approximately 70% of cases, whereas they are infrequent in solid tumors. Additionally, somatic alterations in cyclin-dependent kinase inhibitor 2A (CDKN2A), particularly deletions, are highly prevalent in T-cell ALL (T-ALL) and B-cell ALL (B-ALL) cases, occurring in 78% and 42%, respectively, in contrast to solid tumors where they are present in 11% or less of cases [7].
Further distinctions between pediatric solid and hematological malignancies are evident in the composition of immune cells within the tumor microenvironment (TME). Various immune cell populations, including tumor-infiltrating lymphocytes (TILs), regulatory T cells (Tregs), natural killer (NK) cells, B cells, macrophages, dendritic cells (DCs), and myeloid-derived suppressor cells (MDSCs), play crucial roles in the development and progression of these cancers. The proportions of these immune cells within the TME influence anti-tumor immunity and impact various tumor characteristics. Additionally, the TME encompasses tumor stromal cells, such as fibroblasts, endothelial cells, pericytes, and extracellular matrix (ECM) components like collagen, laminin, and fibronectin, along with non-coding RNAs, growth factors, cytokines, and chemokines, all of which further influence tumor behavior.
In comparison to solid tumors, hematological cancers develop and progress within lymphoid organs where immune cells reside and often initiate anti-tumor immune responses. This close association with immune cells primarily arises from deficiencies in antigen priming [13]. Conversely, in many solid tumors, T-cell priming typically occurs but is impeded at the tumor site by diverse immune suppressive mechanisms. However, recent studies on pediatric AML and B-ALL reveal that these cancers contain infiltrating activated cytotoxic T cells capable of generating neoantigen-specific CD8+ T-cell responses, yet they suffer from chronic exhaustion and lack phenotypes associated with stem-like memory cells [14]. This differs from solid brain and CNS tumors, which are considered immune cold, characterized by high myeloid signatures and low T-cell infiltration [15,16,17]. This distinction is further underscored in a recent study, where an analysis of the immunogenomic and transcriptomic profiles of 202 pediatric tumor cases, focusing on relapsed or refractory solid tumors, uncovered a significant lack of immune infiltration across different tumor types [18].
Similar to solid tumors, pediatric AML blasts increase the number of MDSCs, drive macrophages toward a pro-tumoral phenotype, and exhibit signs of exhausted NK cells [14,19]. The correlation of NK cell infiltration and activation with enhanced outcomes in pediatric leukemia is well-documented in studies, thus sparking considerable interest in NK cell-based immunotherapies for this ailment [20]. With the advent of immunotherapy, analyzing the immune microenvironment to comprehend the interplay between tumors and adjacent immune cells has garnered substantial attention in recent decades. Various types of immunotherapies including monoclonal antibodies (mAbs), transfer of immune cells from donor to patient, checkpoint inhibitors, and vaccines have been tested in different solid tumors. Unlike traditional cancer treatments such as surgery, chemotherapy, or radiation therapy that directly target cancer cells, immunotherapy aims to stimulate the immune system’s natural ability to detect and attack abnormal cancer cells in the body. Immunotherapy has shown remarkable success in shrinking tumors and improving survival rates in some adult cancer patients with various tumor types [21].
Given its promising outcomes, numerous studies have aimed to characterize the immune microenvironment of pediatric tumors. While much research has focused on adult tumors, there is a growing emphasis on profiling the immune landscape surrounding pediatric cancers. The success of Chimeric Antigen Receptor (CAR) T-cell therapy, particularly in targeting the CD19 antigen in B-ALL, holds promise for enhancing immune responses against childhood tumors [22].
However, the immune microenvironment of pediatric tumors differs from that of adult tumors in several ways. The immune system in children is less developed because it is still maturing and exhibits distinct histological and molecular features compared to adults. Children rely more on innate immunity, which is generally non-specific, rather than specific adaptive immunity. In the context of the TME, pediatric tumors have a lower mutational burden and different genetic alterations compared to adult tumors, which can result in unique antigen profiles and immune interactions within the pediatric TME. Additionally, the absence of mutation-derived antigen expression makes these tumors less immunogenic, allowing them to evade or subvert anti-tumor immunity and develop acquired resistance to immunotherapy. Consequently, pediatric tumor patients show different outcomes when treated with drugs studied in adult tumors. To achieve better and improved outcomes, a proper understanding of tumors and their microenvironment in pediatric populations of different ages is crucial for effective cancer therapy.
This review aims to provide a comprehensive overview of recent literature on profiling the immune microenvironment of different pediatric tumors. It summarizes findings on the characterization of the immune microenvironment in pediatric tumors and its role in influencing the TME, interactions between the immune system and tumor cells, tumor growth and development, metastasis, and the response of tumors to immunotherapy. Additionally, the review discusses the substantial challenges associated with targeting the immune microenvironment for future therapies in pediatric cancers.
2. Profiling the Immune Microenvironment of Pediatric Tumors
Many studies have shown that densities of tumor-infiltrating immune cells, including cytotoxic lymphocytes, MDSC, tumor-associated macrophages (TAM), and mast cells in the pediatric TME are strongly associated with immune suppression and anti-tumor immune escape. The TME can be classified into different categories based on the presence and distribution of T lymphocytes. TME with infiltrated immune cells is referred to as immune-inflamed, whereas if T cells are confined to the periphery of the tumor and unable to effectively penetrate the tumor mass, it is called immune-excluded. An “immune-desert” TME indicates the complete absence of T cells.
Molecular profiling of the immune microenvironment of high-risk pediatric tumors based on Immune Pediatric Signature Score (IPASS), which combines immunohistochemistry staining data of T lymphocytes with RNA and whole-genome sequencing, estimated that 31% of high-risk cancers in pediatric patients have infiltrating T cells [23]. In CNS tumors, pediatric gliomas have the most T-cell receptor (TCR) clones, while osteosarcoma and neuroblastoma show more diversity in these clones. Having a diverse range of TCR clones suggests a strong immune response [24]. This diversity in TCR repertoires is linked to prognosis and response to immune therapy in these tumors. Interestingly, the study found that tumor mutational burden (TMB) and neoantigen load, which reflect genetic changes in cancer cells, did not predict T-cell infiltration in pediatric cancers [23]. This implies that the presence of a high TMB or a large number of neoantigens may not necessarily correlate with increased T-cell infiltration and immune response in these cancers.
Similar findings were observed in a murine-based tumor progression glioma model that mimics the progression from low-grade gliomas (LGG) to high-grade gliomas (HGG) [25]. LGG tumors demonstrated a high level of T-cell homing, infiltration, and secretion of interferon-gamma (IFNγ), which is an activation marker for T cells, in comparison to HGG tumors. Furthermore, the study found a significant difference in the proportions of cytotoxic CD8+ and CD4+ T cells between the LGG and HGG tumor cores. LGG tumors exhibited a higher number of CD8+ T cells compared to HGG tumors. However, both LGG and HGG tumors showed relatively small proportions of CD4+ T cells. Moreover, the TME of LGG and HGG tumors harbors distinct macrophage clusters exhibiting different phenotypes [25]. These macrophages transition from an immune-activated phenotype in LGG to an immunosuppressive state in HGG, suggesting their role in immune evasion during different phases of tumor development.
The ability of TAMs to adopt various functional phenotypes is a critical factor in pediatric tumors, influencing tumor growth, the anti-tumor immune response, and therapy outcomes. For example, chemotherapy-treated MYCN-amplified neuroblastoma patients showed increased tumor infiltration of HLA-DR+, CD11c+, CD68+, and CD14+ pro-tumorigenic macrophages that expressed immunosuppressive molecules such as Tim-3, B7-H3, and CD163 [26]. Treatment of these patient’s tumors with anti-colony stimulating factor 1 receptor (CSF1R), which impairs macrophage infiltration, prevents the regrowth of chemotherapy-resistant relapsed tumors [26]. This implies that tumor-infiltrated macrophages play a key role in therapeutic resistance to chemotherapy in neuroblastoma patients.
Similarly, in retinoblastoma, the microenvironment is enriched with pro-tumorigenic macrophages expressing the M2-type microglial marker Iba1+ [27]. The population of these TAMs increases after therapy [27]. Tumor cells reprogram TAMs to promote immune suppression and resistance to anti-tumor drugs. This is supported by studies demonstrating that macrophages cultured in a conditioned medium obtained from retinoblastoma cells exhibit a more pro-tumor M2 phenotype [27,28]. M2-type TAMs are associated with various other types of pediatric tumors, including osteosarcoma, rhabdomyosarcoma, leukemia, lymphoma, and brain tumors. These TAMs express M2-phenotype markers such as CD163, CD204, and CD206, which have been shown to prevent cytotoxic T lymphocytes (CTL) infiltration into the tumor core [29,30,31,32,33].
In pediatric TME, TAMs can interact with MDSCs and other immune cells, thereby influencing the overall immune response within the tumor. MDSCs are a heterogeneous population of immature myeloid cells that commonly arise in the bone marrow and are recruited to the tumor site. MDSCs can suppress immune responses and potentiate TAM-mediated immune suppression, promoting tumor development and growth [34]. Comparative transcriptomic studies suggest that the TME of osteosarcoma pulmonary metastatic sites is significantly enriched with populations of pro-tumor TAMs and polymorphonuclear (PMN) MDSCs compared to primary bone tumors [35]. These TAMs express programmed death ligand-1 (PD-L1) and colocalize with CD8+ lymphocytes at the interface of pulmonary metastatic sites. The expression of PD-L1 on TAMs leads to inhibitory signaling in activated T cells, which downregulates T-cell activity during immune responses and confines activated T cells to peripheral regions [35].
Furthermore, PMN MDSCs are a crucial microenvironment component in hematological malignancies like leukemia and lymphoma. The population of PMN MDSCs decreases in patients who respond well to therapy, indicating their potential as a biomarker for treatment response [34]. In comparison to their normal counterparts, children with ALL exhibit a higher number of both CD33+CD11b+ monocytic MDSCs (M-MDSCs) and PMN-MDSCs. This increase in MDSC populations is significantly upregulated after chemotherapy [36]. Similarly, an elevated population of PMN-MDSCs, identified as CD45+CD19-HLA-DR-CD11b+CD33+CD15+ granulocytic MDSCs (G-MDSCs), is observed in the peripheral blood and bone marrow of children with B-ALL, in comparison to age-matched healthy controls [37].
The studies in the murine model of T-ALL have revealed important insights into the dynamic and functional interactions between MDSCs and other immune cells within the TME [38]. One significant finding is that MDSCs play a role in maintaining the proliferation of immune suppressive CD4+CD8+ T cells in T-ALL-bearing mice. These CD4+CD8+ T cells, in turn, promote the expansion of CD11b+Gr-1+ MDSCs. This reciprocal interaction between MDSCs and CD4+CD8+ T cells contributes to the immune suppressive environment within the TME. Interestingly, treatment with an anti-interleukin 6 (IL6) antibody impedes the accumulation of intratumoral MDSCs, correlating with reduced percentages of CD4+CD8+ T cells [38].
Chemokines and cytokines are critical immune components in the pediatric TME that play an important role in regulating the local balance of pro-tumor or anti-tumor phenotypes. Selective targeting of cytokine/chemokine-cytokine/chemokine receptor signaling networks can promote anti-tumor immunity by altering tumor biological and immunological phenotypes [39]. Moreover, many pediatric neuroblastoma, Ewing sarcoma (ES), osteosarcoma, and ALL, show altered expression of circulating cytokines/chemokines such as CCL2, CXCL4, CXCL6, CXCL10, and CXCL12 [40,41,42]. These circulating chemokines might be clinically utilized as prognostic markers in these tumors.
3. Influence of the Immune Microenvironment on Pediatric Tumor Biology
Genetic and epigenetic alterations in pediatric tumors play an important role in fostering the pro-tumorigenic features of the tumor immune microenvironment. Altered genetic expression in tumor cells drives the expression of cytokines that play a critical role in the recruitment and phenotype of immune cells, especially cells of the myeloid lineage. Understanding these complex and dynamic interactions is essential for comprehending the mechanisms behind immune evasion strategies in pediatric cancers.
3.1. Immune Cell–Tumor Cell Interactions and Their Role in Tumor Progression
Single-cell RNA sequencing analysis of neuroblastoma tumors reveals distinct subtypes of immune cell populations, including myeloid, NK, B, and T cells [43]. These cells form a complex, dynamic, and heterogeneous ecosystem with tumor and stroma cell populations in the TME. Continuous interactions occur between membrane receptors expressed on immune cells and extracellular ligands present on tumor or stroma cells. These interactions play a significant role in shaping the TME ecosystem of pediatric tumors, including neuroblastoma, and can influence tumor growth as well as the response to therapy.
For example, neuroblastoma tumors exhibit elevated levels of the immune checkpoint molecule CD200, which interacts with its corresponding receptor CD200R [44]. CD200R is present on the surface of HLA-DR+CD14+ myeloid cells, CD11c+ dendritic cells, and CD4+ and CD8+ T cells. CD200 expression in neuroblastoma tumors plays a role in regulating the composition of these immune cells and impacting the anti-tumor immune response within TME. Neuroblastoma tumors with increased CD200 expression display reduced numbers of CD4+ and CD8+ T cells, which exhibit more differentiated phenotypes and produce lower amounts of effector and proliferation marker proteins such as IFN-γ and tumor necrosis factor-alpha (TNF-α) [44].
In pediatric AML and B-ALL, overexpression of CD200, along with other receptors such as CD123 and CD56, on leukemic cells and leukemia stem cells has been associated with adverse prognostic features and poorer clinical outcomes [45,46]. This suggests that CD200 expression in these cancers may contribute to treatment resistance and disease aggressiveness. The specific mechanisms by which CD200 expression impacts these cancers are linked to immune modulation and evasion, as seen in neuroblastoma. Upregulation of CD200 in AML cells is associated with an increased population of tumor-suppressive FoxP3+ Tregs, low T-cell cytokine production due to reduced MAPK and STAT3 signaling, impaired T-cell metabolism, decreased production of CD107a+CD8+ memory T cells, reduced TNFα, IL-2, and IFNγ production in CD4+ memory cells, and upregulation of the exhaustion marker TIM-3 in tumor-infiltrating CD4+ bone marrow T cells [47,48,49,50].
Overexpression of immune checkpoint molecules in the TME is an important mechanism in evading immune surveillance and destruction in pediatric tumors. The interaction of immune checkpoint molecules such as cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), programmed cell death protein 1 (PD-1), or B7-H3 receptor with corresponding ligands on tumor cells inhibits their anti-tumor activities, leading to immune suppression. For instance, CTLA-4 overexpression by leukemic cells decreases the expression of CD80, a co-stimulatory molecule, on T cells required for proper T-cell stimulation and activation. CD80 downregulation impairs the ability of T cells to mount an effective immune response against the tumor cells [51]. Pediatric patients with B-ALL exhibit significantly elevated levels of serum CTLA-4 that correspond to the B-ALL cell population, suggesting its role as a marker of disease severity [52].
Furthermore, CTLA-4 upregulation in different intratumoral T-cell populations, such as CD4+CD25highFoxp3+ Tregs, CD4+, and CD8+ T cells, has been associated with abnormal cytokine signaling and poor prognosis in B-ALL and aggressive pediatric sarcoma patients [53,54]. CTLA-4, expressed in T cells, competes with another receptor, CD28, for binding to the proteins CD80 and CD86, which are expressed on the surface of antigen-presenting cells (APCs). CD28 provides a co-stimulatory signal necessary for T-cell activation. When CTLA-4 binds with CD80 and CD86, it decreases CD28 interactions with these proteins, leading to the inhibition of T-cell activation and the maintenance of immune homeostasis. The overexpression of CTLA-4 on Tregs helps to dampen anti-tumor immunity [55]. Moreover, the interaction of CTLA-4 on Tregs with CD80 and CD86 on DCs leads to the downregulation of these co-stimulatory molecules [56]. This results in the instability of contacts between Treg cells and DCs, which limits local Treg cell proliferation. Disruption of this feedback loop through CTLA-4 blockade can induce Treg cell hyper-proliferation [56]. This suggests that, following anti-CTLA4 treatment, Treg cells can continue to proliferate and limit the anti-tumor immune response. Therefore, the functions of CTLA-4 need to be carefully considered in the TME when targeting CTLA-4 in cancer therapy.
Certain pediatric solid tumors, including neuroblastoma, osteosarcoma, retinoblastoma, and rhabdomyosarcoma, display increased PD-L1 expression on their surfaces following treatment. This overexpression is strongly associated with tumor recurrence and a poor prognosis in patients [57,58,59,60]. PD-L1 interacts with PD-1 receptors on immune cells, helping the tumor evade the immune system [57]. Prolonged immunotherapy can elevate PD-1 expression in CD4+ and CD8+ T cells, leading to reduced infiltration of these cells into the tumor and decreased levels of activation markers such as IFNγ, TNFα, and granzyme B [61]. Similarly, chemotherapy can enhance PD-1 and PD-L1 expressions on both tumor-infiltrating inflammatory immune cells and tumor cells, as observed in retinoblastoma. Notably, before treatment, most retinoblastoma tumors exhibit low or negative PD-L1 expression [62].
The loss of function of the RB1 gene, a tumor suppressor gene implicated in retinoblastoma, is associated with increased PD-L1 expression. The RB protein, regulated by phosphorylation at specific sites, can suppress PD-L1 expression by modulating the NF-κB signaling pathway [63]. Phosphorylation of the RB protein at serine-249/threonine-252 (S249/T252) by CDK4/6 promotes its interaction with the NF-κB protein p65, resulting in the inhibition of the NF-κB pathway. Consequently, the loss of RB function, either through knocking down RB or using a CDK4/6 inhibitor, selectively upregulates NF-κB pathway genes, including PD-L1 [63]. These findings suggest that RB plays a crucial role in regulating the immune response within the TME by modulating the NF-κB-PD-L1 pathway. Furthermore, when compared to neuroblastoma and retinoblastoma, osteosarcoma’s TME, particularly in metastatic sites, exhibits significant expression of PD-L1 on tumor cells, macrophages, and T cells, which is upregulated after chemotherapy [60,64]. By combining chemotherapy with PD-L1 inhibition, the immune response can be reactivated, thereby enhancing the anti-tumor activity of T cells in osteosarcoma tumors [64]. These findings demonstrate that the PD-1/PD-L1 signaling pathway promotes the establishment of an immunosuppressive TME that facilitates malignant development.
Moreover, B7-H3 exhibits sustained surface expression on tumor cells, primarily detected in the tumor microenvironment of certain pediatric malignancies. Similar to PD-L1, B7-H3 can interact with T cells, and suppress their anti-tumor activity [65]. Immune profiling of pediatric brain tumor patient-derived orthotopic xenografts (PDOX), representing both low and high grades, consistently demonstrated the presence of B7-H3 and GD2 on tumor cell surfaces [66]. Similarly, B7-H3 has been found to be associated with poor anti-tumor immunity, therapy response, and prognosis in other pediatric tumor types such as AML, medulloblastoma, Wilms tumor, and rhabdomyosarcoma [67,68,69,70]. This suggests that B7-H3 may be involved in regulating the immune response within the pediatric brain tumor microenvironment. Figure 1 depicts the dynamic interplay between tumor and immune cells and their roles within the pediatric TME.
3.2. Metastasis and Immune Microenvironment
Metastasis is a crucial event in tumor progression as it enables cancer cells to escape from the primary tumor and establish secondary tumors or metastases in distant sites of the body. Metastasis is responsible for the majority of cancer-related deaths [71]. In pediatric TME, non-cancer cells, particularly immune cells, secrete factors that can induce ECM remodeling, creating a more permissive environment for cancer cell movement and invasion. For instance, M2-TAMs associated with Wilms tumors activate AKT/PI3K signaling in cancer cells, initiating cellular changes linked to epithelial–mesenchymal transition (EMT) in the tumor cells [72]. In osteosarcoma tumors, TAMs upregulate COX-2, MMP9, and STAT3 phosphorylation in cancer cells, which promotes their migration, invasion, and metastasis to the lungs [73].
In human osteosarcoma lung metastasis specimens, a subpopulation of CD163+ macrophages was found to express erythropoietin receptor (EPOR), CD206, CD163, and PD1, which are known to play significant roles in TAMs immune suppressive and tumor-promoting functions [74]. Interestingly, another study observed more TAM infiltration into patient tumors that showed reduced metastasis and improved survival in high-grade osteosarcoma. Furthermore, it has been observed that macrophage numbers tend to increase following chemotherapy treatment, which is associated with a good therapeutic response [75]. These findings contradict the general understanding of TAMs, which are believed to be tumor-promoting, and suggest that TAMs may have different roles and functions depending on the specific tumor context. Moreover, the induction of M2-type polarization of TAMs by neuroblastoma tumor cells upregulates CXCL2 secretion in TAMs, which enhances tumor cell invasion [76].
TME data from neuroblastoma patients suggest that there is a higher infiltration of TAMs at tumor metastatic sites compared to localized tumors. Furthermore, the expression of specific markers on TAMs, including CD33, CD16, IL6R, IL10, and FCGR3, has been found to have a strong correlation with the survival prediction of patients with metastatic MYCN-nonamplified neuroblastoma [77]. This suggests that macrophage polarization significantly impacts molecular features and therapy-associated outcomes of metastatic neuroblastomas. MDSCs are another subset of the immune cell population that contributes to remodeling the ECM in the pediatric TME. A study conducted on M3-9-M mice, which is a syngeneic orthotopic tumor model of rhabdomyosarcoma that metastasizes to the lungs and shares similarities with human metastatic rhabdomyosarcoma, suggests that MDSCs are the main recruited cell population to secondary organ sites during metastasis. These MDSCs express higher levels of immunosuppressive factors and play a crucial role in the formation of pre-metastatic niches in the lungs [78].
MDSCs can interact with TAMs, dendritic cells, and Tregs to facilitate tumor metastasis. Tregs have been reported to impair local immune cell activity to support metastasis at secondary sites [79]. In patients with primary metastatic ES, Tregs were found at significantly higher frequencies compared to patients with localized disease, suggesting that Tregs may play a role in the development of metastatic disease in ES [80]. Similarly, lower intratumoral cytotoxic CD8+ T cell and higher FOXP3+ Treg cell populations were observed in osteosarcoma patients diagnosed with primary metastasis [81]. These findings indicate the significance of the interplay between various immune cells, including MDSCs, TAMs, dendritic cells, CD8+ T cells, and Tregs, in influencing tumor metastasis. Understanding these complex interactions may aid in developing targeted therapies to counteract the immunosuppressive microenvironment and improve outcomes for pediatric patients with metastatic tumors.
3.3. Immune Cell-Derived Cytokines
Secretion of immune-derived cytokines and chemokines, stimulated by cancer cells, is another critical point of interaction between tumor cells and infiltrated immune cells. These signaling molecules create a complex communication network within the TME that influences various aspects of tumor progression in most pediatric tumors. Specific changes in cytokine levels in the serum, including interleukins and TNFα, have been linked to disease severity, tumor progression, and therapeutic response in children with ALL and AML [82,83]. Tregs and MDSCs are the key immune cells that progressively accumulate within the TME and contribute to immune-suppressive interleukins, leading to the inhibition of anti-tumor activity driven by T and NK cells. Data suggest that Tregs-derived IL-10 and IL-35, work together to promote CD8+ T-cell exhaustion in the TME [84]. Both IL-10 and IL-35 induce the transcriptional repressor BLIMP1, which is known to promote terminal differentiation in T and B cell lineages and the expression of inhibitory receptors like PD-1, LAG3, TIM3, TIGIT, and 2B4 in CD4+ and CD8+ TILs [84,85]. More specifically, IL-35 promotes inhibitory receptor induction on TILs, inhibiting the differentiation of T cells into TCM (central memory T cells), whereas IL-10 regulates cytokine production and effector functions of TILs [84]. IL-10 has been found elevated and associated with the pathogenesis of pediatric neuroblastoma [86], medulloblastoma [87], leukemia, and lymphomas [88,89,90].
Cytokine profiling of CD4+ and CD8+ T cells from children with ALL revealed a significant reduction in IL-2 and IFNγ-producing CD4+ and CD8+ T cells below the normal range, while IL-4 production in CD4+ and CD8+ T-cell subsets increased [91]. A recent similar study in children with ALL has identified a correlation between elevated blood cytokine levels and ALL, suggesting a link between early-life immune events, cytokine imbalances, and the risk of ALL. The study found that patients with ALL exhibit higher levels of a group of correlated cytokines, including IL-1β, IL-8, TNF-α, and VEGF, at birth compared to their healthy counterparts [92]. Similar cytokine profiling has been conducted in children with B-ALL, anaplastic lymphoma kinase-positive anaplastic large cell lymphoma, pediatric pilocytic astrocytomas, as well as children with cancer and febrile neutropenia [93,94,95,96]. These findings collectively suggest an intriguing association between cytokine levels and tumor development.
Furthermore, the dysregulation of serum inflammatory cytokines, such as IL-1β, IL-6, and TNF-α, has been observed in pediatric patients with major depressive disorder (MDD), and it is linked to depression in children with cancer [97]. Cytokine deregulation can arise in response to cancer and its treatments, resulting in the upregulation of inflammatory reactions. Excessive cytokine production induces chronic inflammation, fostering an environment that promotes the development and progression of cancer. Therefore, dysregulated cytokine signaling signatures can be informative biomarkers in pediatric cancer research and clinical management. These signatures could improve cancer detection, patient stratification, and personalized treatment approaches by providing insights into the immune response and its interaction with cancer cells. Table 1 discusses some key cytokines in the pediatric TME and their roles and potential as predictive biomarkers.
3.4. Immunotherapy Response and Predictors in Pediatric Tumors
Before immunotherapy became widely recognized and used in cancer treatment, surgery, radiation, and chemotherapy were standard treatment options for childhood cancers. Immunotherapy has shown remarkable success in some advanced-stage cancers, leading to complete remission and potential cures. Immunotherapy as a multimodal therapy with surgery, radiation, or chemotherapy has shown better survival outcomes in pediatric patients [155,156]. A range of cancer immunotherapy approaches has been tested in pediatric cancer patients, including the use of cytokines like IL-2 [157], adoptive therapies like CAR-T and NK cell therapies [158,159], the use of targeted mAbs [160], immune checkpoint inhibitors [161], or oncolytic viruses that selectively infect and destroy cancer cells [162]. Despite these advances, the effectiveness of immunotherapy is limited in high-risk pediatric cancers due to the absence of recognizable markers. As a result, targeted mAbs and adoptive T-cell therapies have shown particular promise in treating specific pediatric cancer types that express tumor-specific antigens, such as leukemia, neuroblastoma, and melanoma.
Results from two clinical trials by Locatelli et al. and Brown et al. of antibody blinatumomab (Blincyto) in children and young adults with relapsed B-ALL showed that blinatumomab is more effective than chemotherapy [160,163]. Blinatumomab is a CD3/CD19-directed bispecific T-cell engager molecule (BiTE) that brings CD19-expressing leukemia cells and CD3-positive cytotoxic T cells together to facilitate T-cell-mediated leukemia cell killing. In both trials, blinatumomab-treated patients had better outcomes and experienced fewer side effects than patients who received additional chemotherapy as consolidation therapy [160,163]. In the Locatelli et al. study (n = 108), patients treated with blinatumomab exhibited a 2-year event-free survival of 66.2% compared to 27.1% for those receiving chemotherapy. Moreover, blinatumomab showcased higher rates of minimal residual disease (MRD)-negative remission (90% vs. 54%) and a greater proportion of patients proceeded to hematopoietic stem cell transplantation (HSCT) (88.9% vs. 70.4%) [160].
In the Brown et al. study (n = 214), the 2-year disease-free and overall survival rates were 54.4% and 71.3%, respectively, compared to 39.0% and 58.4% with chemotherapy. Blinatumomab also exhibited superior rates of MRD negativity (75% vs. 32%) and a higher percentage of patients proceeding to HSCT (70% vs. 43%) compared to the chemotherapy group [163]. However, there are some differences in patient selection in both studies that may explain some of the discrepancies in their treatment outcomes. The Locatelli et al. trial involved patients under 18, randomized after completing three cycles of chemotherapy, with approximately half already achieving MRD negativity [160]. In contrast, Brown and co-authors enrolled participants up to age 30, randomizing them after one cycle of chemotherapy, with only around 25% showing MRD negativity [163]. In addition, the Locatelli et al. study suggests that patients with very early relapse benefit most from blinatumomab, while the Brown et al. study indicates limited activity in patients responding unfavorably to reinduction therapy, emphasizing the significant impact of patient selection on treatment outcomes [163].
The other most notable mAbs approved for pediatric cancer patients include Dinutuximab, an anti-GD2 mAb for the treatment of neuroblastoma [164]. Anti-GD2 mAb is specific to neuroblastoma cells, as they express the tumor-specific antigen GD2, a glycolipid antigen with restricted expression in normal nerve tissues [165]. Anti-GD2 monoclonal antibodies have been used in combination with various administration regimens, including chemotherapy, cytokines like IL-2 and granulocyte–macrophage colony-stimulating factor (GM-CSF), or allogeneic NK cells in neuroblastoma treatment. Several types of anti-GD2 antibodies have been explored for therapeutic purposes in neuroblastoma patients, including the human anti-mouse mAb antibody 3F8 [166], human/mouse chimeric mAbs like Ch14.18/SP2.0 (dinutuximab, Unituxin®, United Therapeutics, Durham, NC, USA) [167,168], Ch14.18/CHO (dinutuximab beta, Qarziba®, EUSA Pharma, Hemel Hempstea, UK) [169], and Ch14.18K332A (Provenance Biopharmaceuticals, Waltham, MA, USA; St. Jude Children’s Research Hospital, Memphis, TN, USA) [170], alongside humanized antibodies like hu14.18-IL2 developed by the National Cancer Institute (NCI) [171] and hu3F8 (Naxitamab, DANYELZA®, naxitamab-gqgk) [172] developed by Memorial Sloan Kettering Cancer Center, New York, NY, USA.
In vitro findings indicate disparities in their binding to GD2, duration of effectiveness, and cytotoxic strength. For instance, hu3F8 exhibits stronger binding to GD2 compared to Ch14.18 but has a shorter half-life [173]. Ch14.18K332A demonstrates heightened antibody-dependent cellular cytotoxicity (ADCC) by NK cells and neutrophils, while Ch14.18 displays lower binding and cytotoxic efficacy but persists longer [174,175]. The impact of these characteristics on clinical outcomes awaits future testing. Approved anti-GD2 antibodies for high-risk neuroblastoma treatment in the US include naxitamab (Danyelza®) and dinutuximab (Unituxin®), while in Europe, dinutuximab beta (Qarziba®) is available.
Furthermore, multicenter trials have been either completed or are currently underway to compare regimens containing anti-GD2 with conventional treatments. For instance, one trial investigated the efficacy of anti-GD2 (murine 3F8) treatment alongside GM-CSF in children in remission from stage 4 neuroblastoma [176]. Patients were assigned to three regimens: regimen A received 3F8 alone, regimen B received 3F8 + intravenous GM-CSF + 13-cis-retinoic acid (CRA) post-stem-cell transplantation (SCT), and regimen C received 3F8 + subcutaneous GM-CSF + CRA. Additionally, some ultra-high-risk patients were treated with regimen C. Over a 5-year period, survival rates demonstrated improvement, with regimen C exhibiting the highest progression-free survival (62%) and overall survival (81%). Moreover, ultra-high-risk patients experienced enhanced survival with this treatment approach [176]. Similarly, a randomized phase III trial conducted by the Children’s Oncology Group (COG) showed that the combination of anti-GD2 ch14.18 (dinutuximab) with GM-CSF and IL-2 could improve survival rates among children with high-risk neuroblastoma [177]. However, in another phase 3 trial utilized ch14.18/CHO (dinutuximab beta) and isotretinoin as maintenance therapy with or without subcutaneous IL-2, adding IL-2 did not enhance outcomes and led to increased toxicity compared to dinutuximab beta alone [121]. The observed differences in these two IL-2 studies could originate from variations in treatment protocols, including the incorporation of GM-CSF alongside IL-2 and the subcutaneous delivery of IL-2 in the first study. The rationale for integrating GM-CSF and IL-2 with dinutuximab is to enhance antibody-dependent cellular cytotoxicity (ADCC), primarily by amplifying granulocyte and NK cell counts and activation, which constitute the primary mechanism of dinutuximab-induced anti-neuroblastoma immune reactions. Moreover, in both trials, immunotherapy showed greater efficacy in patients with minimal residual disease or those who responded well to induction therapy but did not achieve a complete response. This highlights the potential advantage of immunotherapy for individuals displaying favorable responses to initial treatments. However, it also underscores the pressing need for continued refinement of treatment regimens to enhance outcomes and minimize toxicity in patients with recurrent or progressive disease, even following various treatments.
Other strategies tested to augment the effect of anti-GD2 mAbs with various combination therapies in neuroblastoma include Ch14.18 + irinotecan + temozolomide + GM-CSF [178], Hu14.18K322A + induction chemotherapy [179], Hu14.18-IL2 + expanded NK cells (NCT02573896), Ch14.18 + temozolomide + expanded γδ T cells (NCT05400603), Anti-GD2 CARs [180] and many others. These diverse strategies underscore the multifaceted efforts aimed at improving outcomes for patients with neuroblastoma, particularly those at high risk or in remission. Despite the use of anti-GD2, many patients still relapsed. Continued research and clinical trials in this area are vital for advancing treatment options and ultimately improving survival rates and quality of life for children battling this challenging disease.
Other antibodies approved for human use against different protein targets include rituximab, an anti-CD20 mAb for ALL [181]; ipilimumab, an anti-CTLA-4 antibody for pediatric patients with advanced melanoma [182]; and pembrolizumab, an anti-PD1 antibody approved for specific subsets of pediatric patients [183]. A study investigated the effectiveness of rituximab in newly diagnosed pediatric ALL patients who tested positive for CD20. Rituximab, combined with chemotherapy, has shown success in treating adult ALL and has been well received. In this pediatric trial, twenty patients were randomly assigned to either receive rituximab with standard chemotherapy (Intervention group) or undergo standard chemotherapy alone (Control group). The addition of rituximab resulted in lower absolute blast count and peripheral blood MRD levels on day 8, suggesting potential benefits in pediatric ALL treatment [181]. In another study, rituximab was used to reduce allergic reactions associated with Polyethylene glycol (PEG)-asparaginase (pegaspargase), a commonly used chemotherapy agent. Despite a decrease in CD20-positive B cells, newly diagnosed pediatric ALL patients who received rituximab experienced a significant incidence of infusion reactions. Additionally, there were no significant differences observed in pegaspargase allergies, anti-pegaspargase antibodies, or MRD levels between the rituximab and control groups [184]. These findings contrast with the previous study; however, both studies had a small number of participants, indicating the need for further research. Additionally, in the later study, rituximab was administered alongside multimodal induction therapy, which could have already reduced the leukemia burden and decreased the MRD, potentially masking any rituximab effects.
Similarly, ipilimumab has been assessed in 45 pediatric patients across two clinical trials. The first trial, a dose-finding study involving 33 individuals aged 2 to 21 years with relapsed or refractory solid tumors [185], and the second, an open-label, single-arm trial with 12 adolescents aged 12 to under 18 years with unresectable stage III or IV melanoma [186]. In both trials, ipilimumab demonstrated efficacy in some melanoma patients and exhibited a safety profile comparable to that observed in adults. However, the second trial was prematurely halted due to slow accrual caused by enrollment challenges, primarily because this disease is exceedingly rare in children [186]. Despite its rarity, there is a pressing need for parallel studies comparing children and adult cohorts for surgical management and innovative treatment options. This approach will facilitate the implementation of standardized guidelines, ensuring that medical practices are tailored to the specific needs of pediatric patients while benefiting from insights gleaned from adult studies, thus enhancing overall care and outcomes across different age groups.
There are numerous ongoing trials investigating these antibodies, as well as antibodies targeting other molecules, in combination with various immunotherapies or chemotherapy, for various pediatric tumors (Table 2). Moreover, adoptive cell therapy including a CD19-targeting CAR T-cell immunotherapy has been approved for subsets of pediatric patients with advanced leukemia [187]. Despite immunotherapy becoming a viable treatment option for many pediatric cancer patients, it does not work for everyone [188]. The primary reason for this is that the success of immunotherapy depends on the patient’s endogenous immunity. Several factors can limit the effectiveness of immunotherapy, including a limited number of neoantigens for immunotherapies to target due to a lower mutational load, an immunosuppressive tumor microenvironment that lacks anti-tumor cytotoxic lymphocytes, low surface expression of MHC-I, allowing evasion from T-cell-mediated anti-tumor immunity, and tumor heterogeneity arising from the presence of several malignant subpopulations within a single tumor mass [8,189].
To address these limitations, novel innovative approaches and procedures, combined with a deep understanding of intra-tumor complexity within the patients, are needed. Molecular profiling of a patient’s tumors is a key strategy for overcoming these challenges and improving the decision-making process for targeted therapies, ultimately leading to higher cure rates in cancer patients. Identifying specific molecular markers reflecting germline and somatic mutations, epigenetic changes in DNA, chromosomal aberrations, and protein expression alterations can help clinicians accurately stratify tumors on a molecular level and predict treatment responses. It also involves utilizing computational tools to predict neoantigens in patients with low mutational loads and develop personalized cancer vaccines targeting these neoantigens to stimulate the immune system.
Furthermore, combining immunotherapy with other treatment modalities like targeted therapy, immune-checkpoint inhibitors, or radiation therapy has been proven effective in enhancing the immune system’s response by increasing the number of neoantigens and reducing tumor heterogeneity [196]. Therefore, the use of rationally designed combinatory therapy strategies is likely to have a significant impact on pediatric tumors, provided the tumors are molecularly well defined. However, a molecular profile may evolve over time due to the accrual of molecular abnormalities that arise during cancer progression and are influenced by the selective pressures of previous treatments. As a result, there is a requirement for innovative technologies to continuously monitor cancer genotypes in real time, enabling necessary adaptations in cancer therapy regimens. Moreover, as advancements continue in cell-manufacturing approaches, adoptive immune-cell therapies, such as autologous or allogeneic transplantation of T or NK cells, or genetically modified CAR-T cells expressing novel T-cell receptors or chimeric antigen receptors, have achieved success in providing enduring responses in certain cases. These strategies are particularly effective in tumors with low antigen presentation or those characterized by an immune-suppressive tumor microenvironment.
However, despite promising developments in novel approaches and procedures and the widespread implementation of immunotherapy in pediatric tumors, not all patients receive similar benefits from the treatment. In addition, higher off-target cytotoxicity in combination therapies, tumor cells’ ability to evade anti-tumor immunity, and challenges associated with adoptive cell therapies like inflammation, cytokine release syndrome (CRS), neurological toxicity, manufacturing barriers, and limited accessibility pose significant challenges for their use in the field of pediatric oncology [197].
4. Conclusions
Like adult tumors, pediatric tumors also rely on a complex TME that surrounds them and contributes significantly to their growth, development, and metastasis. However, unlike adult tumors, the pediatric immune microenvironment remains largely unexplored. One significant area that is poorly understood is the complex signaling network triggered by specific oncogenic drivers within tumor cells that educate and shape immune cells toward a pro-tumor phenotype. Given the extensive diversity in the immune microenvironment and the critical signaling pathways triggered by tumor cells across various tumor types and tissues, it is imperative to investigate how to navigate and comprehend this diversity. As our comprehension of immune–tumor interaction networks advances, we can classify the immune microenvironment in diverse pediatric tumors. This classification facilitates the recognition of vital immune cells and their related signaling mechanisms involved in suppressing anti-tumor immunity. Such insights have the potential to accelerate responses to established standard-of-care therapies and constitute a noteworthy area for future research. While currently approved U.S. Food and Drug Administration (FDA) immunotherapy treatments exhibit restricted efficacy, strategically targeting the immune microenvironment emerges as an appealing approach for cancer treatment. Encouraging preclinical investigations across diverse pediatric tumors has demonstrated the potential to leverage the immune microenvironment of tumors for therapeutic advantages [140,198,199,200].
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| WHO | World Health Organization |
| CNS | Central nervous system |
| ALL | Acute lymphoblastic leukemia |
| AML | Acute myeloid leukemia |
| JAK | Janus kinases |
| STAT | Signal transducer and activator of transcription |
| PI3K | Phosphoinositide 3-kinases |
| ALK | Anaplastic lymphoma kinase |
| NF1 | Neurofibromatosis type 1 |
| PTEN | Phosphatase and tensin homolog |
| FLT3 | FMS-like tyrosine kinase 3 |
| PIK3CA | Phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha or p110α |
| PIK3R1 | Phosphoinositide-3-kinase regulatory subunit 1 |
| CDKN2A | Cyclin-dependent kinase Inhibitor 2A |
| TME | Tumor microenvironment |
| TILs | Tumor-infiltrating lymphocytes |
| Tregs | Regulatory T cells |
| NK cells | Natural killer cells |
| DCs | Dendritic cells |
| MDSCs | Myeloid-derived suppressor cells |
| ECM | Extracellular matrix |
| mAbs | Monoclonal antibodies |
| CAR | Chimeric antigen receptor |
| TAMs | Tumor-associated macrophages |
| TMB | Tumor mutational burden |
| LGG | Low-grade gliomas |
| HGG | High-grade gliomas |
| IFNγ | Interferon-gamma |
| CSF1R | Colony stimulating factor 1 receptor |
| CTLs | Cytotoxic T lymphocytes |
| PMN | Polymorphonuclear |
| PD-L1 | Programmed death ligand-1 |
| IL6 | Interleukin 6 |
| ES | Ewing sarcoma |
| TNF-α | Tumor necrosis factor-alpha |
| CTLA-4 | Cytotoxic T-lymphocyte-associated protein 4 |
| PD-1 | Programmed cell death protein 1 |
| MRD | Minimal residual disease |
| HSCT | Hematopoietic stem cell transplantation |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| SCT | Stem-cell transplantation |
| COG | Children’s oncology group |
| ADCC | Antibody-dependent cellular cytotoxicity |
References
- Steliarova-Foucher, E.; Colombet, M.; Ries, L.A.G.; Moreno, F.; Dolya, A.; Bray, F.; Hesseling, P.; Shin, H.Y.; Stiller, C.A.; IICC-3 Contributors. International incidence of childhood cancer, 2001–2010: A population-based registry study. Lancet Oncol. 2017, 18, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Bonaventure, A.; Harewood, R.; Stiller, C.A.; Gatta, G.; Clavel, J.; Stefan, D.C.; Carreira, H.; Spika, D.; Marcos-Gragera, R.; Peris-Bonet, R.; et al. Worldwide comparison of survival from childhood leukaemia for 1995–2009, by subtype, age, and sex (CONCORD-2): A population-based study of individual data for 89 828 children from 198 registries in 53 countries. Lancet Haematol. 2017, 4, e202–e217. [Google Scholar] [CrossRef] [PubMed]
- Siegel, D.A.; King, J.B.; Lupo, P.J.; Durbin, E.B.; Tai, E.; Mills, K.; Van Dyne, E.; Buchanan Lunsford, N.; Henley, S.J.; Wilson, R.J. Counts, incidence rates, and trends of pediatric cancer in the United States, 2003–2019. J. Natl. Cancer Inst. 2023, 115, 1337–1354. [Google Scholar] [CrossRef] [PubMed]
- Curtin, S.C.; Anderson, R.N. Declines in Cancer Death Rates Among Youth: United States, 2001–2021. NCHS Data Brief 2023, 484, 1–8. [Google Scholar]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 30 April 2024).
- Ma, X.; Liu, Y.; Liu, Y.; Alexandrov, L.B.; Edmonson, M.N.; Gawad, C.; Zhou, X.; Li, Y.; Rusch, M.C.; Easton, J.; et al. Pan-cancer genome and transcriptome analyses of 1699 paediatric leukaemias and solid tumours. Nature 2018, 555, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Grobner, S.N.; Worst, B.C.; Weischenfeldt, J.; Buchhalter, I.; Kleinheinz, K.; Rudneva, V.A.; Johann, P.D.; Balasubramanian, G.P.; Segura-Wang, M.; Brabetz, S.; et al. The landscape of genomic alterations across childhood cancers. Nature 2018, 555, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Walsh, M.F.; Wu, G.; Edmonson, M.N.; Gruber, T.A.; Easton, J.; Hedges, D.; Ma, X.; Zhou, X.; Yergeau, D.A.; et al. Germline Mutations in Predisposition Genes in Pediatric Cancer. N. Engl. J. Med. 2015, 373, 2336–2346. [Google Scholar] [CrossRef] [PubMed]
- Jedraszek, K.; Malczewska, M.; Parysek-Wojcik, K.; Lejman, M. Resistance Mechanisms in Pediatric B-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2022, 23, 3067. [Google Scholar] [CrossRef]
- Fletcher, J.I.; Ziegler, D.S.; Trahair, T.N.; Marshall, G.M.; Haber, M.; Norris, M.D. Too many targets, not enough patients: Rethinking neuroblastoma clinical trials. Nat. Rev. Cancer 2018, 18, 389–400. [Google Scholar] [CrossRef]
- Marchandet, L.; Lallier, M.; Charrier, C.; Baud’huin, M.; Ory, B.; Lamoureux, F. Mechanisms of Resistance to Conventional Therapies for Osteosarcoma. Cancers 2021, 13, 683. [Google Scholar] [CrossRef]
- Curran, E.K.; Godfrey, J.; Kline, J. Mechanisms of Immune Tolerance in Leukemia and Lymphoma. Trends Immunol. 2017, 38, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Bailur, J.K.; McCachren, S.S.; Pendleton, K.; Vasquez, J.C.; Lim, H.S.; Duffy, A.; Doxie, D.B.; Kaushal, A.; Foster, C.; DeRyckere, D.; et al. Risk-associated alterations in marrow T cells in pediatric leukemia. JCI Insight 2020, 5, e140179. [Google Scholar] [CrossRef] [PubMed]
- Mackay, A.; Burford, A.; Molinari, V.; Jones, D.T.W.; Izquierdo, E.; Brouwer-Visser, J.; Giangaspero, F.; Haberler, C.; Pietsch, T.; Jacques, T.S.; et al. Molecular, Pathological, Radiological, and Immune Profiling of Non-brainstem Pediatric High-Grade Glioma from the HERBY Phase II Randomized Trial. Cancer Cell 2018, 33, 829–842 e825. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, N.A.P.; DeGolier, K.; Kovar, H.M.; Davis, A.; Hoglund, V.; Stevens, J.; Winter, C.; Deutsch, G.; Furlan, S.N.; Vitanza, N.A.; et al. Characterization of the immune microenvironment of diffuse intrinsic pontine glioma: Implications for development of immunotherapy. Neuro Oncol. 2019, 21, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.D.; Franz, C.J.; Brennan, L.; Brouwer-Visser, J.; Tam, R.; Korski, K.; Koeppen, H.; Ziai, J.; Babitzki, G.; Ranchere-Vince, D.; et al. Immune contexture of paediatric cancers. Eur. J. Cancer 2022, 170, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Byron, S.A.; Hendricks, W.P.D.; Nagulapally, A.B.; Kraveka, J.M.; Ferguson, W.S.; Brown, V.I.; Eslin, D.E.; Mitchell, D.; Cornelius, A.; Roberts, W.; et al. Genomic and Transcriptomic Analysis of Relapsed and Refractory Childhood Solid Tumors Reveals a Diverse Molecular Landscape and Mechanisms of Immune Evasion. Cancer Res. 2021, 81, 5818–5832. [Google Scholar] [CrossRef] [PubMed]
- Perzolli, A.; Koedijk, J.B.; Zwaan, C.M.; Heidenreich, O. Targeting the innate immune system in pediatric and adult AML. Leukemia 2024, 38, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Mizia-Malarz, A.; Sobol-Milejska, G. NK Cells as Possible Prognostic Factor in Childhood Acute Lymphoblastic Leukemia. Dis. Markers 2019, 2019, 3596983. [Google Scholar] [CrossRef]
- Okoye, C.; Tran, M.; Soladoye, E.; Akahara, D.E.; Emeasoba, C.M.; Ojinna, B.T.; Anasonye, E.; Obadare, O.O.; Diala, C.S.; Salaudeen, B.H.; et al. A Review of 10-Year Survivability of Immunotherapy in the Management of Colon Cancer. Cureus 2023, 15, e43189. [Google Scholar] [CrossRef]
- Myers, R.M.; Li, Y.; Barz Leahy, A.; Barrett, D.M.; Teachey, D.T.; Callahan, C.; Fasano, C.C.; Rheingold, S.R.; DiNofia, A.; Wray, L.; et al. Humanized CD19-Targeted Chimeric Antigen Receptor (CAR) T Cells in CAR-Naive and CAR-Exposed Children and Young Adults With Relapsed or Refractory Acute Lymphoblastic Leukemia. J. Clin. Oncol. 2021, 39, 3044–3055. [Google Scholar] [CrossRef] [PubMed]
- Mayoh, C.; Gifford, A.J.; Terry, R.; Lau, L.M.S.; Wong, M.; Rao, P.; Shai-Hee, T.; Saletta, F.; Khuong-Quang, D.A.; Qin, V.; et al. A novel transcriptional signature identifies T-cell infiltration in high-risk paediatric cancer. Genome Med. 2023, 15, 20. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, A.; De Luca, C.D.; Locatelli, F.; Velardi, E. Thymic Function and T-Cell Receptor Repertoire Diversity: Implications for Patient Response to Checkpoint Blockade Immunotherapy. Front. Immunol. 2021, 12, 752042. [Google Scholar] [CrossRef]
- Rajendran, S.; Hu, Y.; Canella, A.; Peterson, C.; Gross, A.; Cam, M.; Nazzaro, M.; Haffey, A.; Serin-Harmanci, A.; Distefano, R.; et al. Single-cell RNA sequencing reveals immunosuppressive myeloid cell diversity during malignant progression in a murine model of glioma. Cell Rep. 2023, 42, 112197. [Google Scholar] [CrossRef]
- Valind, A.; Verhoeven, B.M.; Enoksson, J.; Karlsson, J.; Christensson, G.; Manas, A.; Aaltonen, K.; Jansson, C.; Bexell, D.; Baryawno, N.; et al. Macrophage infiltration promotes regrowth in MYCN-amplified neuroblastoma after chemotherapy. Oncoimmunology 2023, 12, 2184130. [Google Scholar] [CrossRef]
- Cuadrado-Vilanova, M.; Liu, J.; Paco, S.; Aschero, R.; Burgueno, V.; Sirab, N.; Pascual-Pasto, G.; Correa, G.; Balaguer-Lluna, L.; Castillo-Ecija, H.; et al. Identification of immunosuppressive factors in retinoblastoma cell secretomes and aqueous humor from patients. J. Pathol. 2022, 257, 327–339. [Google Scholar] [CrossRef]
- Chen, S.; Chen, X.; Qiu, J.; Chen, P.; Han, X.; Wu, Y.; Zhuang, J.; Yang, M.; Wu, C.; Wu, N.; et al. Exosomes derived from retinoblastoma cells enhance tumour deterioration by infiltrating the microenvironment. Oncol. Rep. 2021, 45, 278–290. [Google Scholar] [CrossRef]
- Gomez-Brouchet, A.; Illac, C.; Gilhodes, J.; Bouvier, C.; Aubert, S.; Guinebretiere, J.M.; Marie, B.; Larousserie, F.; Entz-Werle, N.; de Pinieux, G.; et al. CD163-positive tumor-associated macrophages and CD8-positive cytotoxic lymphocytes are powerful diagnostic markers for the therapeutic stratification of osteosarcoma patients: An immunohistochemical analysis of the biopsies fromthe French OS2006 phase 3 trial. Oncoimmunology 2017, 6, e1331193. [Google Scholar] [CrossRef]
- Poli, E.; Cattelan, M.; Zanetti, I.; Scagnellato, A.; Giordano, G.; Zin, A.; Bisogno, G.; Bonvini, P. Autoantibody profiling of alveolar rhabdomyosarcoma patients unveils tumor-associated antigens with diagnostic and prognostic significance. Oncoimmunology 2021, 10, 1954765. [Google Scholar] [CrossRef] [PubMed]
- Miari, K.E.; Guzman, M.L.; Wheadon, H.; Williams, M.T.S. Macrophages in Acute Myeloid Leukaemia: Significant Players in Therapy Resistance and Patient Outcomes. Front. Cell Dev. Biol. 2021, 9, 692800. [Google Scholar] [CrossRef]
- Barros, M.H.; Hassan, R.; Niedobitek, G. Tumor-associated macrophages in pediatric classical Hodgkin lymphoma: Association with Epstein-Barr virus, lymphocyte subsets, and prognostic impact. Clin. Cancer Res. 2012, 18, 3762–3771. [Google Scholar] [CrossRef]
- Gutmann, D.H.; Kettenmann, H. Microglia/Brain Macrophages as Central Drivers of Brain Tumor Pathobiology. Neuron 2019, 104, 442–449. [Google Scholar] [CrossRef]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Ligon, J.A.; Choi, W.; Cojocaru, G.; Fu, W.; Hsiue, E.H.; Oke, T.F.; Siegel, N.; Fong, M.H.; Ladle, B.; Pratilas, C.A.; et al. Pathways of immune exclusion in metastatic osteosarcoma are associated with inferior patient outcomes. J. Immunother. Cancer 2021, 9, e001772. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.L.; El-Shanshory, M.R.; Abdou, S.H.; Attia, M.S.; Sobhy, S.M.; Zidan, M.F.; Zidan, A.A. Chemotherapy alters the increased numbers of myeloid-derived suppressor and regulatory T cells in children with acute lymphoblastic leukemia. Immunopharmacol. Immunotoxicol. 2018, 40, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Chen, Y.Y.; He, Y.Y.; Wang, J.Y.; Yang, J.P.; Zhong, S.L.; Jiang, N.; Zhou, P.; Jiang, H.; Zhou, J. Expansion and activation of granulocytic, myeloid-derived suppressor cells in childhood precursor B cell acute lymphoblastic leukemia. J. Leukoc. Biol. 2017, 102, 449–458. [Google Scholar] [CrossRef]
- Grazioli, P.; Orlando, A.; Giordano, N.; Noce, C.; Peruzzi, G.; Abdollahzadeh, B.; Screpanti, I.; Campese, A.F. Notch-Signaling Deregulation Induces Myeloid-Derived Suppressor Cells in T-Cell Acute Lymphoblastic Leukemia. Front. Immunol. 2022, 13, 809261. [Google Scholar] [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef]
- Kacher, J.; Manches, O.; Aspord, C.; Sartelet, H.; Chaperot, L. Impaired Antitumor Immune Response in MYCN-amplified Neuroblastoma Is Associated with Lack of CCL2 Secretion and Poor Dendritic Cell Recruitment. Cancer Res. Commun. 2022, 2, 577–589. [Google Scholar] [CrossRef]
- Cillo, A.R.; Mukherjee, E.; Bailey, N.G.; Onkar, S.; Daley, J.; Salgado, C.; Li, X.; Liu, D.; Ranganathan, S.; Burgess, M.; et al. Ewing Sarcoma and Osteosarcoma Have Distinct Immune Signatures and Intercellular Communication Networks. Clin. Cancer Res. 2022, 28, 4968–4982. [Google Scholar] [CrossRef]
- de Vasconcellos, J.F.; Laranjeira, A.B.; Zanchin, N.I.; Otubo, R.; Vaz, T.H.; Cardoso, A.A.; Brandalise, S.R.; Yunes, J.A. Increased CCL2 and IL-8 in the bone marrow microenvironment in acute lymphoblastic leukemia. Pediatr. Blood Cancer 2011, 56, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, B.M.; Mei, S.; Olsen, T.K.; Gustafsson, K.; Valind, A.; Lindstrom, A.; Gisselsson, D.; Fard, S.S.; Hagerling, C.; Kharchenko, P.V.; et al. The immune cell atlas of human neuroblastoma. Cell Rep. Med. 2022, 3, 100657. [Google Scholar] [CrossRef] [PubMed]
- Xin, C.; Zhu, J.; Gu, S.; Yin, M.; Ma, J.; Pan, C.; Tang, J.; Zhang, P.; Liu, Y.; Bai, X.F.; et al. CD200 is overexpressed in neuroblastoma and regulates tumor immune microenvironment. Cancer Immunol. Immunother. 2020, 69, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, E.Z.; Madney, Y.; Eldin, D.N.; Shafik, N.F. Overexpression of CD200 and CD123 is a major influential factor in the clinical course of pediatric acute myeloid leukemia. Exp. Mol. Pathol. 2021, 118, 104597. [Google Scholar] [CrossRef] [PubMed]
- Aref, S.; Azmy, E.; El-Bakry, K.; Ibrahim, L.; Abdel Aziz, S. Prognostic impact of CD200 and CD56 expression in pediatric B-cell acute lymphoblastic leukemia patients. Pediatr. Hematol. Oncol. 2017, 34, 275–285. [Google Scholar] [CrossRef]
- Coles, S.J.; Hills, R.K.; Wang, E.C.; Burnett, A.K.; Man, S.; Darley, R.L.; Tonks, A. Increased CD200 expression in acute myeloid leukemia is linked with an increased frequency of FoxP3+ regulatory T cells. Leukemia 2012, 26, 2146–2148. [Google Scholar] [CrossRef] [PubMed]
- Herbrich, S.; Baran, N.; Cai, T.; Weng, C.; Aitken, M.J.L.; Post, S.M.; Henderson, J.; Shi, C.; Richard-Carpentier, G.; Sauvageau, G.; et al. Overexpression of CD200 is a Stem Cell-Specific Mechanism of Immune Evasion in AML. J. Immunother. Cancer 2021, 9, e002968. [Google Scholar] [CrossRef] [PubMed]
- Coles, S.J.; Hills, R.K.; Wang, E.C.; Burnett, A.K.; Man, S.; Darley, R.L.; Tonks, A. Expression of CD200 on AML blasts directly suppresses memory T-cell function. Leukemia 2012, 26, 2148–2151. [Google Scholar] [CrossRef] [PubMed]
- Blaeschke, F.; Willier, S.; Stenger, D.; Lepenies, M.; Horstmann, M.A.; Escherich, G.; Zimmermann, M.; Rojas Ringeling, F.; Canzar, S.; Kaeuferle, T.; et al. Leukemia-induced dysfunctional TIM-3(+)CD4(+) bone marrow T cells increase risk of relapse in pediatric B-precursor ALL patients. Leukemia 2020, 34, 2607–2620. [Google Scholar] [CrossRef]
- Do, P.; Beckwith, K.A.; Cheney, C.; Tran, M.; Beaver, L.; Griffin, B.G.; Mo, X.; Liu, Y.; Lapalombella, R.; Hertlein, E.; et al. Leukemic B Cell CTLA-4 Suppresses Costimulation of T Cells. J. Immunol. 2019, 202, 2806–2816. [Google Scholar] [CrossRef]
- Simone, R.; Tenca, C.; Fais, F.; Luciani, M.; De Rossi, G.; Pesce, G.; Bagnasco, M.; Saverino, D. A soluble form of CTLA-4 is present in paediatric patients with acute lymphoblastic leukaemia and correlates with CD1d+ expression. PLoS ONE 2012, 7, e44654. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.X.; Xiao, H.R.; Wang, G.B.; Chen, X.W.; Li, C.G.; Mai, H.R.; Yuan, X.L.; Liu, G.S.; Wen, F.Q. Preliminary investigation on the abnormal mechanism of CD4(+)FOXP3(+)CD25(high) regulatory T cells in pediatric B-cell acute lymphoblastic leukemia. Exp. Ther. Med. 2018, 16, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Hingorani, P.; Maas, M.L.; Gustafson, M.P.; Dickman, P.; Adams, R.H.; Watanabe, M.; Eshun, F.; Williams, J.; Seidel, M.J.; Dietz, A.B. Increased CTLA-4(+) T cells and an increased ratio of monocytes with loss of class II (CD14(+) HLA-DR(lo/neg)) found in aggressive pediatric sarcoma patients. J. Immunother. Cancer 2015, 3, 35. [Google Scholar] [CrossRef] [PubMed]
- Peggs, K.S.; Quezada, S.A.; Chambers, C.A.; Korman, A.J.; Allison, J.P. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti-CTLA-4 antibodies. J. Exp. Med. 2009, 206, 1717–1725. [Google Scholar] [CrossRef]
- Marangoni, F.; Zhakyp, A.; Corsini, M.; Geels, S.N.; Carrizosa, E.; Thelen, M.; Mani, V.; Prussmann, J.N.; Warner, R.D.; Ozga, A.J.; et al. Expansion of tumor-associated Treg cells upon disruption of a CTLA-4-dependent feedback loop. Cell 2021, 184, 3998–4015.e19. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Sho, M.; Sawai, T.; Kanehiro, H.; Maeda, K.; Yoshida, M.; Tsukada, R.; Nomura, M.; Okuyama, H. Potential role of the PD-L1 expression and tumor-infiltrating lymphocytes on neuroblastoma. Pediatr. Surg. Int. 2020, 36, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Siebert, N.; Zumpe, M.; Juttner, M.; Troschke-Meurer, S.; Lode, H.N. PD-1 blockade augments anti-neuroblastoma immune response induced by anti-GD(2) antibody ch14.18/CHO. Oncoimmunology 2017, 6, e1343775. [Google Scholar] [CrossRef] [PubMed]
- Geoerger, B.; Kang, H.J.; Yalon-Oren, M.; Marshall, L.V.; Vezina, C.; Pappo, A.; Laetsch, T.W.; Petrilli, A.S.; Ebinger, M.; Toporski, J.; et al. Pembrolizumab in paediatric patients with advanced melanoma or a PD-L1-positive, advanced, relapsed, or refractory solid tumour or lymphoma (KEYNOTE-051): Interim analysis of an open-label, single-arm, phase 1–2 trial. Lancet Oncol. 2020, 21, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Sundara, Y.T.; Kostine, M.; Cleven, A.H.; Bovee, J.V.; Schilham, M.W.; Cleton-Jansen, A.M. Increased PD-L1 and T-cell infiltration in the presence of HLA class I expression in metastatic high-grade osteosarcoma: A rationale for T-cell-based immunotherapy. Cancer Immunol. Immunother. 2017, 66, 119–128. [Google Scholar] [CrossRef]
- Su, Y.; Luo, B.; Lu, Y.; Wang, D.; Yan, J.; Zheng, J.; Xiao, J.; Wang, Y.; Xue, Z.; Yin, J.; et al. Anlotinib Induces a T Cell-Inflamed Tumor Microenvironment by Facilitating Vessel Normalization and Enhances the Efficacy of PD-1 Checkpoint Blockade in Neuroblastoma. Clin. Cancer Res. 2022, 28, 793–809. [Google Scholar] [CrossRef]
- Miracco, C.; Toti, P.; Gelmi, M.C.; Aversa, S.; Baldino, G.; Galluzzi, P.; De Francesco, S.; Petrelli, F.; Sorrentino, E.; Belmonte, G.; et al. Retinoblastoma Is Characterized by a Cold, CD8+ Cell Poor, PD-L1- Microenvironment, Which Turns Into Hot, CD8+ Cell Rich, PD-L1+ After Chemotherapy. Investig. Ophthalmol. Vis. Sci. 2021, 62, 6. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Ding, D.; Yan, Y.; Li, H.; Wang, B.; Ma, L.; Ye, Z.; Ma, T.; Wu, Q.; Rodrigues, D.N.; et al. Phosphorylated RB Promotes Cancer Immunity by Inhibiting NF-kappaB Activation and PD-L1 Expression. Mol. Cell 2019, 73, 22–35.e6. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, F.; Yu, P.; Wang, J.; Liu, Z.; Bao, Q.; Zhang, W.; Wen, J. Sorafenib inhibits doxorubicin-induced PD-L1 upregulation to improve immunosuppressive microenvironment in Osteosarcoma. J. Cancer Res. Clin. Oncol. 2022, 149, 5127–5138. [Google Scholar] [CrossRef] [PubMed]
- Hofmeyer, K.A.; Ray, A.; Zang, X. The contrasting role of B7-H3. Proc. Natl. Acad. Sci. USA 2008, 105, 10277–10278. [Google Scholar] [CrossRef] [PubMed]
- Haydar, D.; Houke, H.; Chiang, J.; Yi, Z.; Ode, Z.; Caldwell, K.; Zhu, X.; Mercer, K.S.; Stripay, J.L.; Shaw, T.I.; et al. Cell-surface antigen profiling of pediatric brain tumors: B7-H3 is consistently expressed and can be targeted via local or systemic CAR T-cell delivery. Neuro Oncol. 2021, 23, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, E.I.; Du, H.; Shou, P.; Song, F.; Suzuki, K.; Ahn, S.; Li, G.; Ferrone, S.; Su, L.; Savoldo, B.; et al. Preclinical Evaluation of B7-H3-specific Chimeric Antigen Receptor T Cells for the Treatment of Acute Myeloid Leukemia. Clin. Cancer Res. 2021, 27, 3141–3153. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Poolen, G.C.; van Vliet, L.C.; Schipper, J.G.; Broekhuizen, R.; Monnikhof, M.; Van Hecke, W.; Vermeulen, J.F.; Bovenschen, N. Pediatric medulloblastoma express immune checkpoint B7-H3. Clin. Transl. Oncol. 2022, 24, 1204–1208. [Google Scholar] [CrossRef] [PubMed]
- Hont, A.B.; Dumont, B.; Sutton, K.S.; Anderson, J.; Kentsis, A.; Drost, J.; Hong, A.L.; Verschuur, A. The tumor microenvironment and immune targeting therapy in pediatric renal tumors. Pediatr. Blood Cancer 2023, 70 (Suppl. 2), e30110. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, T.; Miyachi, M.; Sugimoto, Y.; Yagyu, S.; Kikuchi, K.; Tsuchiya, K.; Iehara, T.; Hosoi, H. Reduced B7-H3 expression by PAX3-FOXO1 knockdown inhibits cellular motility and promotes myogenic differentiation in alveolar rhabdomyosarcoma. Sci. Rep. 2021, 11, 18802. [Google Scholar] [CrossRef]
- Dillekas, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef]
- Tian, K.; Du, G.; Wang, X.; Wu, X.; Li, L.; Liu, W.; Wu, R. MMP-9 secreted by M2-type macrophages promotes Wilms’ tumour metastasis through the PI3K/AKT pathway. Mol. Biol. Rep. 2022, 49, 3469–3480. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Guo, W.; Ren, T.; Huang, Y.; Wang, S.; Liu, K.; Zheng, B.; Yang, K.; Zhang, H.; Liang, X. Tumor-associated macrophages promote lung metastasis and induce epithelial-mesenchymal transition in osteosarcoma by activating the COX-2/STAT3 axis. Cancer Lett. 2019, 440–441, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, M.; Wei, R.; Wu, J. Identification and Functional Analysis of EPOR(+) Tumor-Associated Macrophages in Human Osteosarcoma Lung Metastasis. J. Immunol. Res. 2020, 2020, 9374240. [Google Scholar] [CrossRef] [PubMed]
- Buddingh, E.P.; Kuijjer, M.L.; Duim, R.A.; Burger, H.; Agelopoulos, K.; Myklebost, O.; Serra, M.; Mertens, F.; Hogendoorn, P.C.; Lankester, A.C.; et al. Tumor-infiltrating macrophages are associated with metastasis suppression in high-grade osteosarcoma: A rationale for treatment with macrophage activating agents. Clin. Cancer Res. 2011, 17, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, O.; Yoshida, M.; Koma, Y.; Yanai, T.; Hasegawa, D.; Kosaka, Y.; Nishimura, N.; Yokozaki, H. Collaboration of cancer-associated fibroblasts and tumour-associated macrophages for neuroblastoma development. J. Pathol. 2016, 240, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Asgharzadeh, S.; Salo, J.A.; Ji, L.; Oberthuer, A.; Fischer, M.; Berthold, F.; Hadjidaniel, M.; Liu, C.W.; Metelitsa, L.S.; Pique-Regi, R.; et al. Clinical significance of tumor-associated inflammatory cells in metastatic neuroblastoma. J. Clin. Oncol. 2012, 30, 3525–3532. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowska, S.; Beury, D.W.; Gopalan, V.; Tycko, A.K.; Qin, H.; Clements, M.E.; Drake, J.; Nwanze, C.; Murgai, M.; Rae, Z.; et al. Genetically engineered myeloid cells rebalance the core immune suppression program in metastasis. Cell 2021, 184, 2033–2052.e21. [Google Scholar] [CrossRef] [PubMed]
- Kos, K.; Aslam, M.A.; van de Ven, R.; Wellenstein, M.D.; Pieters, W.; van Weverwijk, A.; Duits, D.E.M.; van Pul, K.; Hau, C.S.; Vrijland, K.; et al. Tumor-educated T(regs) drive organ-specific metastasis in breast cancer by impairing NK cells in the lymph node niche. Cell Rep. 2022, 38, 110447. [Google Scholar] [CrossRef] [PubMed]
- Brinkrolf, P.; Landmeier, S.; Altvater, B.; Chen, C.; Pscherer, S.; Rosemann, A.; Ranft, A.; Dirksen, U.; Juergens, H.; Rossig, C. A high proportion of bone marrow T cells with regulatory phenotype (CD4+CD25hiFoxP3+) in Ewing sarcoma patients is associated with metastatic disease. Int. J. Cancer 2009, 125, 879–886. [Google Scholar] [CrossRef]
- Fritzsching, B.; Fellenberg, J.; Moskovszky, L.; Sapi, Z.; Krenacs, T.; Machado, I.; Poeschl, J.; Lehner, B.; Szendroi, M.; Bosch, A.L.; et al. CD8(+)/FOXP3(+)-ratio in osteosarcoma microenvironment separates survivors from non-survivors: A multicenter validated retrospective study. Oncoimmunology 2015, 4, e990800. [Google Scholar] [CrossRef]
- Almeida, R.D.S.; Ramos, A.M.L.; Luna, C.F.; Pedrosa, F.; Donadi, E.A.; Lucena-Silva, N. Cytokines and soluble HLA-G levels in bone marrow stroma and their association with the survival rate of patients exhibiting childhood T-cell acute lymphoblastic leukemia. Cytokine 2018, 102, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Z.; Yuce, D.; Kirkiz, S.; Kocak, U.; Ozmen, F. Prognostic role of serum cytokines and soluble HLA-G levels in children with leukemia who undergo allogeneic stem cell transplantation. Cytokine 2022, 153, 155869. [Google Scholar] [CrossRef] [PubMed]
- Sawant, D.V.; Yano, H.; Chikina, M.; Zhang, Q.; Liao, M.; Liu, C.; Callahan, D.J.; Sun, Z.; Sun, T.; Tabib, T.; et al. Adaptive plasticity of IL-10(+) and IL-35(+) T(reg) cells cooperatively promotes tumor T cell exhaustion. Nat. Immunol. 2019, 20, 724–735. [Google Scholar] [CrossRef]
- Rutishauser, R.L.; Martins, G.A.; Kalachikov, S.; Chandele, A.; Parish, I.A.; Meffre, E.; Jacob, J.; Calame, K.; Kaech, S.M. Transcriptional repressor Blimp-1 promotes CD8(+) T cell terminal differentiation and represses the acquisition of central memory T cell properties. Immunity 2009, 31, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Z.; Guo, X.; Liao, R.; Yang, K.; Ye, L.; You, Z. Involvement of IL-10 and TGF-beta in HLA-E-mediated neuroblastoma migration and invasion. Oncotarget 2016, 7, 44340–44349. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, S.; Fang, T.; Qiu, X.; Wang, X.; Zhou, X.; Morse, M.A.; Hobeika, A.; Wu, W.; Yang, H.; et al. Changes in Peripheral Blood Regulatory T Cells and IL-6 and IL-10 Levels Predict Response of Pediatric Medulloblastoma and Germ Cell Tumors With Residual or Disseminated Disease to Craniospinal Irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2021, 111, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, L.; Wittig, S.; Gruhn, B. Interleukin-10-592 polymorphism: Impact on relapse and survival after allogeneic hematopoietic stem cell transplantation in children with hematological malignancies. J. Cancer Res. Clin. Oncol. 2022, 148, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Yavuz, G.; Taçyildiz, N.; Babacan, E.; Ünal, E.; İkincioğullari, A.; Eğin, Y.; Göksel, D.; Ertem, U.; Dağdemir, A.; Cin, Ş.; et al. Serum Interleukin-10 in Childhood Malignant Lymphomas (Preliminary Report). Pediatr. Res. 1999, 45, 775. [Google Scholar] [CrossRef]
- Stevens, A.M.; Horton, T.M.; Glasser, C.L.; Gerbing, R.B.; Aplenc, R.; Alonzo, T.A.; Redell, M.S. IL-10 and TNFalpha are associated with decreased survival in low-risk pediatric acute myeloid leukemia; a children’s oncology group report. Pediatr. Hematol. Oncol. 2023, 40, 147–158. [Google Scholar] [CrossRef]
- Zhang, X.L.; Komada, Y.; Chipeta, J.; Li, Q.S.; Inaba, H.; Azuma, E.; Yamamoto, H.; Sakurai, M. Intracellular cytokine profile of T cells from children with acute lymphoblastic leukemia. Cancer Immunol. Immunother. 2000, 49, 165–172. [Google Scholar] [CrossRef]
- Whitehead, T.P.; Wiemels, J.L.; Zhou, M.; Kang, A.Y.; McCoy, L.S.; Wang, R.; Fitch, B.; Petrick, L.M.; Yano, Y.; Imani, P.; et al. Cytokine Levels at Birth in Children Who Developed Acute Lymphoblastic Leukemia. Cancer Epidemiol. Biomark. Prev. 2021, 30, 1526–1535. [Google Scholar] [CrossRef]
- Magalhaes-Gama, F.; Kerr, M.W.A.; de Araujo, N.D.; Ibiapina, H.N.S.; Neves, J.C.F.; Hanna, F.S.A.; Xabregas, L.A.; Carvalho, M.; Alves, E.B.; Tarrago, A.M.; et al. Imbalance of Chemokines and Cytokines in the Bone Marrow Microenvironment of Children with B-Cell Acute Lymphoblastic Leukemia. J. Oncol. 2021, 2021, 5530650. [Google Scholar] [CrossRef]
- Knorr, F.; Damm-Welk, C.; Ruf, S.; Singh, V.K.; Zimmermann, M.; Reiter, A.; Woessmann, W. Blood cytokine concentrations in pediatric patients with anaplastic lymphoma kinase-positive anaplastic large cell lymphoma. Haematologica 2018, 103, 477–485. [Google Scholar] [CrossRef]
- Bte Syed Sulaiman, N.; Kuick, C.H.; Chang, K.T.E.; Wan, K.R.; Looi, W.S.; Low, D.C.Y.; Seow, W.T.; Low, S.Y.Y. Cytokines in Pediatric Pilocytic Astrocytomas: A Clinico-Pathological Study. NeuroSci 2021, 2, 95–108. [Google Scholar] [CrossRef]
- Tapia, L.I.; Olivares, M.; Torres, J.P.; De la Maza, V.; Valenzuela, R.; Contardo, V.; Tordecilla, J.; Alvarez, A.M.; Varas, M.; Zubieta, M.; et al. Cytokine and chemokine profiles in episodes of persistent high-risk febrile neutropenia in children with cancer. Cytokine 2021, 148, 155619. [Google Scholar] [CrossRef]
- Narendran, G.; Tomfohr, L.; Schulte, F. Inflammatory cytokines and depression in children with cancer: A review of the literature. Pediatr. Hematol. Oncol. 2018, 35, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Singh, A.; Yadav, G.; Kushwaha, R.; Ali, W.; Verma, S.P.; Singh, U.S. Serum Tumor Necrosis Factor-Alpha Levels in Acute Leukemia and Its Prognostic Significance. Cureus 2022, 14, e24835. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chu, X.; Gao, L.; Ling, J.; Xiao, P.; Lu, J.; Wang, Y.; He, H.; Li, J.; Hu, Y.; et al. High Expression of Interleukin-3 Receptor Alpha Chain (CD123) Predicts Favorable Outcome in Pediatric B-Cell Acute Lymphoblastic Leukemia Lacking Prognosis-Defining Genomic Aberrations. Front. Oncol. 2021, 11, 614420. [Google Scholar] [CrossRef]
- Wittwer, N.L.; Brumatti, G.; Marchant, C.; Sandow, J.J.; Pudney, M.K.; Dottore, M.; D’Andrea, R.J.; Lopez, A.F.; Ekert, P.G.; Ramshaw, H.S. High CD123 levels enhance proliferation in response to IL-3, but reduce chemotaxis by downregulating CXCR4 expression. Blood Adv. 2017, 1, 1067–1079. [Google Scholar] [CrossRef]
- Zenatti, P.P.; Ribeiro, D.; Li, W.; Zuurbier, L.; Silva, M.C.; Paganin, M.; Tritapoe, J.; Hixon, J.A.; Silveira, A.B.; Cardoso, B.A.; et al. Oncogenic IL7R gain-of-function mutations in childhood T-cell acute lymphoblastic leukemia. Nat. Genet. 2011, 43, 932–939. [Google Scholar] [CrossRef]
- Oliveira, M.L.; Veloso, A.; Garcia, E.G.; Iyer, S.; Pereira, C.; Barreto, V.M.; Langenau, D.M.; Barata, J.T. Mutant IL7R collaborates with MYC to induce T-cell acute lymphoblastic leukemia. Leukemia 2022, 36, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kong, L.; Kim, S.; Lee, S.; Oh, S.; Jo, S.; Jang, I.; Kim, T.D. The Role of IL-7 and IL-7R in Cancer Pathophysiology and Immunotherapy. Int. J. Mol. Sci. 2022, 23, 10412. [Google Scholar] [CrossRef] [PubMed]
- Peace, D.J.; Cheever, M.A. Toxicity and therapeutic efficacy of high-dose interleukin 2. In vivo infusion of antibody to NK-1.1 attenuates toxicity without compromising efficacy against murine leukemia. J. Exp. Med. 1989, 169, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.S.; Hallner, A.; Brune, M.; Nilsson, S.; Thoren, F.B.; Martner, A.; Hellstrand, K. Immunotherapy with HDC/IL-2 may be clinically efficacious in acute myeloid leukemia of normal karyotype. Hum. Vaccin. Immunother. 2020, 16, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.; Ducassou, S.; Leblanc, T.; Pasquet, M.; Rousseau, A.; Ragu, C.; Cachanado, M.; Nelken, B.; Bertrand, Y.; Michel, G.; et al. Maintenance Therapy With Interleukin-2 for Childhood AML: Results of ELAM02 Phase III Randomized Trial. Hemasphere 2018, 2, e159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hresko, M.E.; Picton, L.K.; Su, L.; Hollander, M.J.; Nunez-Cruz, S.; Zhang, Z.; Assenmacher, C.A.; Sockolosky, J.T.; Garcia, K.C.; et al. A human orthogonal IL-2 and IL-2Rbeta system enhances CAR T cell expansion and antitumor activity in a murine model of leukemia. Sci. Transl. Med. 2021, 13, eabg6986. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.M.; Miller, J.M.; Munoz, J.O.; Gaikwad, A.S.; Redell, M.S. Interleukin-6 levels predict event-free survival in pediatric AML and suggest a mechanism of chemotherapy resistance. Blood Adv. 2017, 1, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, H.; Inoue, K.; Ogawa, H.; Yamagami, T.; Soma, T.; Miyake, S.; Hirata, M.; Kishimoto, T. The expression of IL-6 and its related genes in acute leukemia. Leuk. Lymphoma 1996, 21, 49–52. [Google Scholar] [CrossRef]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef]
- Bruserud, O.; Tore Gjertsen, B.; Brustugun, O.T.; Bassoe, C.F.; Nesthus, I.; Espen Akselsen, P.; Buhring, H.J.; Pawelec, G. Effects of interleukin 10 on blast cells derived from patients with acute myelogenous leukemia. Leukemia 1995, 9, 1910–1920. [Google Scholar]
- Chang, J.S.; Zhou, M.; Buffler, P.A.; Chokkalingam, A.P.; Metayer, C.; Wiemels, J.L. Profound deficit of IL10 at birth in children who develop childhood acute lymphoblastic leukemia. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1736–1740. [Google Scholar] [CrossRef] [PubMed]
- Fitch, B.A.; Zhou, M.; Situ, J.; Surianarayanan, S.; Reeves, M.Q.; Hermiston, M.L.; Wiemels, J.L.; Kogan, S.C. Decreased IL-10 accelerates B-cell leukemia/lymphoma in a mouse model of pediatric lymphoid leukemia. Blood Adv. 2022, 6, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, H.; Yan, M.; Zhang, L.; Tang, J.; Li, Q.; Gu, X.; Gong, Q. Interleukin-33 Promotes Cell Survival via p38 MAPK-Mediated Interleukin-6 Gene Expression and Release in Pediatric AML. Front. Immunol. 2020, 11, 595053. [Google Scholar] [CrossRef] [PubMed]
- Naef, P.; Radpour, R.; Jaeger-Ruckstuhl, C.A.; Bodmer, N.; Baerlocher, G.M.; Doehner, H.; Doehner, K.; Riether, C.; Ochsenbein, A.F. IL-33-ST2 signaling promotes stemness in subtypes of myeloid leukemia cells through the Wnt and Notch pathways. Sci. Signal. 2023, 16, eadd7705. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.; Edwards, D.K.t.; Eide, C.A.; Newell, L.; Traer, E.; Medeiros, B.C.; Pollyea, D.A.; Deininger, M.W.; Collins, R.H.; Tyner, J.W.; et al. Identification of Interleukin-1 by Functional Screening as a Key Mediator of Cellular Expansion and Disease Progression in Acute Myeloid Leukemia. Cell Rep. 2017, 18, 3204–3218. [Google Scholar] [CrossRef] [PubMed]
- Fitter, S.; Bradey, A.L.; Kok, C.H.; Noll, J.E.; Wilczek, V.J.; Venn, N.C.; Law, T.; Paisitkriangkrai, S.; Story, C.; Saunders, L.; et al. CKLF and IL1B transcript levels at diagnosis are predictive of relapse in children with pre-B-cell acute lymphoblastic leukaemia. Br. J. Haematol. 2021, 193, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Arriero, M.D.M.; Villatoro, A. Interleukin-1beta as emerging therapeutic target in hematological malignancies and potentially in their complications. Blood Rev. 2017, 31, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Fultang, L.; Gamble, L.D.; Gneo, L.; Berry, A.M.; Egan, S.A.; De Bie, F.; Yogev, O.; Eden, G.L.; Booth, S.; Brownhill, S.; et al. Macrophage-Derived IL1beta and TNFalpha Regulate Arginine Metabolism in Neuroblastoma. Cancer Res. 2019, 79, 611–624. [Google Scholar] [CrossRef]
- Ladenstein, R.; Potschger, U.; Valteau-Couanet, D.; Luksch, R.; Castel, V.; Yaniv, I.; Laureys, G.; Brock, P.; Michon, J.M.; Owens, C.; et al. Interleukin 2 with anti-GD2 antibody ch14.18/CHO (dinutuximab beta) in patients with high-risk neuroblastoma (HR-NBL1/SIOPEN): A multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1617–1629. [Google Scholar] [CrossRef]
- Pession, A.; Prete, A.; Locatelli, F.; Pierinelli, S.; Pession, A.L.; Maccario, R.; Magrini, E.; De Bernardi, B.; Paolucci, P.; Paolucci, G. Immunotherapy with low-dose recombinant interleukin 2 after high-dose chemotherapy and autologous stem cell transplantation in neuroblastoma. Br. J. Cancer 1998, 78, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Cicek, F.; Troschke-Meurer, S.; Ceylan, K.; Jahns, L.J.; Zumpe, M.; Siebert, N.; Ehlert, K.; Lode, H.N. Impact of IL-2 on Treatment Tolerance in Patients With High-Risk Neuroblastoma Treated With Dinutuximab Beta-Based Immunotherapy. Front. Pediatr. 2020, 8, 582820. [Google Scholar] [CrossRef] [PubMed]
- Raeber, M.E.; Sahin, D.; Karakus, U.; Boyman, O. A systematic review of interleukin-2-based immunotherapies in clinical trials for cancer and autoimmune diseases. EBioMedicine 2023, 90, 104539. [Google Scholar] [CrossRef] [PubMed]
- Ara, T.; Song, L.; Shimada, H.; Keshelava, N.; Russell, H.V.; Metelitsa, L.S.; Groshen, S.G.; Seeger, R.C.; DeClerck, Y.A. Interleukin-6 in the bone marrow microenvironment promotes the growth and survival of neuroblastoma cells. Cancer Res. 2009, 69, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Louault, K.; Porras, T.; Lee, M.H.; Muthugounder, S.; Kennedy, R.J.; Blavier, L.; Sarte, E.; Fernandez, G.E.; Yang, F.; Pawel, B.R.; et al. Fibroblasts and macrophages cooperate to create a pro-tumorigenic and immune resistant environment via activation of TGF-beta/IL-6 pathway in neuroblastoma. Oncoimmunology 2022, 11, 2146860. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lathia, J.D.; Wu, Q.; Wang, J.; Li, Z.; Heddleston, J.M.; Eyler, C.E.; Elderbroom, J.; Gallagher, J.; Schuschu, J.; et al. Targeting interleukin 6 signaling suppresses glioma stem cell survival and tumor growth. Stem Cells 2009, 27, 2393–2404. [Google Scholar] [CrossRef]
- Holst, C.B.; Christensen, I.J.; Skjoth-Rasmussen, J.; Hamerlik, P.; Poulsen, H.S.; Johansen, J.S. Systemic Immune Modulation in Gliomas: Prognostic Value of Plasma IL-6, YKL-40, and Genetic Variation in YKL-40. Front. Oncol. 2020, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. Interaction between human osteosarcoma and mesenchymal stem cells via an interleukin-8 signaling loop in the tumor microenvironment. Cell Commun. Signal. 2018, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Baglio, S.R.; Schrijver, C.D.; Massaro, C.; Lagerweij, T.; Martin, C.G.; Giorgio, C.; Brandolini, L.; Gavioli, E.M.; Allegretti, M.; Pegtel, D.M. Combined IL-6 and IL-8 inhibition to overcome mesenchymal stem cell (MSC)-induced resistance to antimetastatic drugs in osteosarcoma. J. Clin. Oncol. 2022, 40, 10037. [Google Scholar] [CrossRef]
- Cheng, M.; Cai, W.; Huang, W.; Chen, Y.; Wu, Z.; Luo, P.; Yan, W. Histone deacetylase 6 regulated expression of IL-8 is involved in the doxorubicin (Dox) resistance of osteosarcoma cells via modulating ABCB1 transcription. Eur. J. Pharmacol. 2018, 840, 1–8. [Google Scholar] [CrossRef]
- Xiao, H.; Chen, L.; Luo, G.; Son, H.; Prectoni, J.H.; Zheng, W. Effect of the cytokine levels in serum on osteosarcoma. Tumour Biol. 2014, 35, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.S.; Jian, T.Y.; Sung, S.Y.; Hsieh, C.L.; Huang, M.H.; Fang, C.L.; Wong, T.T.; Lin, Y.L. Enrichment of Tumor-Infiltrating B Cells in Group 4 Medulloblastoma in Children. Int. J. Mol. Sci. 2022, 23, 5287. [Google Scholar] [CrossRef] [PubMed]
- Huettner, C.; Czub, S.; Kerkau, S.; Roggendorf, W.; Tonn, J.C. Interleukin 10 is expressed in human gliomas in vivo and increases glioma cell proliferation and motility in vitro. Anticancer Res. 1997, 17, 3217–3224. [Google Scholar] [PubMed]
- Sreenivasan, L.; Wang, H.; Yap, S.Q.; Leclair, P.; Tam, A.; Lim, C.J. Autocrine IL-6/STAT3 signaling aids development of acquired drug resistance in Group 3 medulloblastoma. Cell Death Dis. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- White, C.L.; Jayasekara, W.S.N.; Picard, D.; Chen, J.; Watkins, D.N.; Cain, J.E.; Remke, M.; Gough, D.J. A Sexually Dimorphic Role for STAT3 in Sonic Hedgehog Medulloblastoma. Cancers 2019, 11, 1702. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nong, L.; Wei, Y.; Qin, S.; Zhou, Y.; Tang, Y. Association of interleukin-12 polymorphisms and serum IL-12p40 levels with osteosarcoma risk. DNA Cell Biol. 2013, 32, 605–610. [Google Scholar] [CrossRef]
- Tugues, S.; Burkhard, S.H.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; Vom Berg, J.; Kulig, P.; Becher, B. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Zalacain, M.; Bunuales, M.; Marrodan, L.; Labiano, S.; Gonzalez-Huarriz, M.; Martinez-Velez, N.; Laspidea, V.; Puigdelloses, M.; Garcia-Moure, M.; Gonzalez-Aparicio, M.; et al. Local administration of IL-12 with an HC vector results in local and metastatic tumor control in pediatric osteosarcoma. Mol. Ther. Oncolytics 2021, 20, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.E.; Starr, R.; Aguilar, B.; Shami, A.F.; Martinez, C.; D’Apuzzo, M.; Barish, M.E.; Forman, S.J.; Jensen, M.C. Stem-like tumor-initiating cells isolated from IL13Ralpha2 expressing gliomas are targeted and killed by IL13-zetakine-redirected T Cells. Clin. Cancer Res. 2012, 18, 2199–2209. [Google Scholar] [CrossRef]
- Rechberger, J.S.; Porath, K.A.; Zhang, L.; Nesvick, C.L.; Schrecengost, R.S.; Sarkaria, J.N.; Daniels, D.J. IL-13Ralpha2 Status Predicts GB-13 (IL13.E13K-PE4E) Efficacy in High-Grade Glioma. Pharmaceutics 2022, 14, 922. [Google Scholar] [CrossRef]
- Pollack, I.F.; Jakacki, R.I.; Butterfield, L.H.; Hamilton, R.L.; Panigrahy, A.; Potter, D.M.; Connelly, A.K.; Dibridge, S.A.; Whiteside, T.L.; Okada, H. Antigen-specific immune responses and clinical outcome after vaccination with glioma-associated antigen peptides and polyinosinic-polycytidylic acid stabilized by lysine and carboxymethylcellulose in children with newly diagnosed malignant brainstem and nonbrainstem gliomas. J. Clin. Oncol. 2014, 32, 2050–2058. [Google Scholar] [CrossRef]
- Burga, R.A.; Yvon, E.; Chorvinsky, E.; Fernandes, R.; Cruz, C.R.Y.; Bollard, C.M. Engineering the TGFbeta Receptor to Enhance the Therapeutic Potential of Natural Killer Cells as an Immunotherapy for Neuroblastoma. Clin. Cancer Res. 2019, 25, 4400–4412. [Google Scholar] [CrossRef]
- Fuchs, B.; Inwards, C.Y.; Janknecht, R. Vascular endothelial growth factor expression is up-regulated by EWS-ETS oncoproteins and Sp1 and may represent an independent predictor of survival in Ewing’s sarcoma. Clin. Cancer Res. 2004, 10, 1344–1353. [Google Scholar] [CrossRef]
- DuBois, S.G.; Marina, N.; Glade-Bender, J. Angiogenesis and vascular targeting in Ewing sarcoma: A review of preclinical and clinical data. Cancer 2010, 116, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.S.; Kleinerman, E.S. Tumor Vessel Development and Expansion in Ewing’s Sarcoma: A Review of the Vasculogenesis Process and Clinical Trials with Vascular-Targeting Agents. Sarcoma 2011, 2011, 165837. [Google Scholar] [CrossRef] [PubMed]
- Capasso, L.; Florio, M.; Lillo, M.; Basilico, M.; De Santis, V.; Ziranu, A.; Grasso, A.; Minutillo, F.; Maccauro, G. Vascular endothelial growth factor expression as a biomarker of prognosis in patients with chondrosarcoma, Ewing’s sarcoma and osteosarcoma. Current concepts. J. Biol. Regul. Homeost. Agents 2019, 33, 39–43. [Google Scholar] [PubMed]
- Ghanem, M.A.; van Steenbrugge, G.J.; Sudaryo, M.K.; Mathoera, R.B.; Nijman, J.M.; van der Kwast, T.H. Expression and prognostic relevance of vascular endothelial growth factor (VEGF) and its receptor (FLT-1) in nephroblastoma. J. Clin. Pathol. 2003, 56, 107–113. [Google Scholar] [CrossRef]
- Rowe, D.H.; Huang, J.; Kayton, M.L.; Thompson, R.; Troxel, A.; O’Toole, K.M.; Yamashiro, D.; Stolar, C.J.; Kandel, J.J. Anti-VEGF antibody suppresses primary tumor growth and metastasis in an experimental model of Wilms’ tumor. J. Pediatr. Surg. 2000, 35, 30–32; discussion 32–33. [Google Scholar] [CrossRef]
- Hong, B.; Dong, R. Research advances in the targeted therapy and immunotherapy of Wilms tumor: A narrative review. Transl. Cancer Res. 2021, 10, 1559–1567. [Google Scholar] [CrossRef]
- Tang, M.J.; Ma, X.L.; He, X.L.; Pan, W.H.; Zhang, X.H.; Jiang, S.Y.; Gao, J.; Li, F.; Yao, W.; Gu, S.; et al. A multicenter prospective study on the management of hepatoblastoma in children: A report from the Chinese Children’s Cancer Group. World J. Pediatr. 2023. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Li, C.; Wang, S.; Jiang, W.; Liu, Z.; Zhou, S.; Liu, X.; McNutt, M.A.; Li, G. Alpha-fetoprotein: A new member of intracellular signal molecules in regulation of the PI3K/AKT signaling in human hepatoma cell lines. Int. J. Cancer 2011, 128, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Dai, X.; Dai, J.; Ding, C.; Zhang, Z.; Lin, Z.; Hu, J.; Lu, M.; Wang, Z.; Qi, Y.; et al. AFP promotes HCC progression by suppressing the HuR-mediated Fas/FADD apoptotic pathway. Cell Death Dis. 2020, 11, 822. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.A.; Malogolowkin, M.H.; Furman, W.L.; Piao, J.; Krailo, M.D.; Chung, N.; Brock, L.; Towbin, A.J.; McCarville, E.B.; Finegold, M.J.; et al. Vincristine/irinotecan/temsirolimus upfront window treatment of high-risk hepatoblastoma: A report from the Children’s Oncology Group AHEP0731 Study Committee. Pediatr. Blood Cancer 2023, 70, e30365. [Google Scholar] [CrossRef] [PubMed]
- Hutzen, B.; Paudel, S.N.; Naeimi Kararoudi, M.; Cassady, K.A.; Lee, D.A.; Cripe, T.P. Immunotherapies for pediatric cancer: Current landscape and future perspectives. Cancer Metastasis Rev. 2019, 38, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.; Ludwig, K.; Pacenta, H.L.; Klesse, L.J.; Watt, T.C.; Laetsch, T.W. Recent progress in the treatment of cancer in children. CA Cancer J. Clin. 2021, 71, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Reaman, G.H.; Hank, J.A.; Cairo, M.S.; Anderson, P.; Blazar, B.R.; Frierdich, S.; Sondel, P.M. A phase II trial of human recombinant interleukin-2 administered as a 4-day continuous infusion for children with refractory neuroblastoma, non-Hodgkin’s lymphoma, sarcoma, renal cell carcinoma, and malignant melanoma. A Childrens Cancer Group study. Cancer 1995, 75, 2959–2965. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Kochenderfer, J.N.; Stetler-Stevenson, M.; Cui, Y.K.; Delbrook, C.; Feldman, S.A.; Fry, T.J.; Orentas, R.; Sabatino, M.; Shah, N.N.; et al. T cells expressing CD19 chimeric antigen receptors for acute lymphoblastic leukaemia in children and young adults: A phase 1 dose-escalation trial. Lancet 2015, 385, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Rubnitz, J.E.; Inaba, H.; Kang, G.; Gan, K.; Hartford, C.; Triplett, B.M.; Dallas, M.; Shook, D.; Gruber, T.; Pui, C.H.; et al. Natural killer cell therapy in children with relapsed leukemia. Pediatr. Blood Cancer 2015, 62, 1468–1472. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, F.; Zugmaier, G.; Rizzari, C.; Morris, J.D.; Gruhn, B.; Klingebiel, T.; Parasole, R.; Linderkamp, C.; Flotho, C.; Petit, A.; et al. Effect of Blinatumomab vs Chemotherapy on Event-Free Survival Among Children With High-risk First-Relapse B-Cell Acute Lymphoblastic Leukemia: A Randomized Clinical Trial. JAMA 2021, 325, 843–854. [Google Scholar] [CrossRef]
- Heczey, A.; Louis, C.U.; Savoldo, B.; Dakhova, O.; Durett, A.; Grilley, B.; Liu, H.; Wu, M.F.; Mei, Z.; Gee, A.; et al. CAR T Cells Administered in Combination with Lymphodepletion and PD-1 Inhibition to Patients with Neuroblastoma. Mol. Ther. 2017, 25, 2214–2224. [Google Scholar] [CrossRef]
- Cripe, T.P.; Ngo, M.C.; Geller, J.I.; Louis, C.U.; Currier, M.A.; Racadio, J.M.; Towbin, A.J.; Rooney, C.M.; Pelusio, A.; Moon, A.; et al. Phase 1 study of intratumoral Pexa-Vec (JX-594), an oncolytic and immunotherapeutic vaccinia virus, in pediatric cancer patients. Mol. Ther. 2015, 23, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.A.; Ji, L.; Xu, X.; Devidas, M.; Hogan, L.E.; Borowitz, M.J.; Raetz, E.A.; Zugmaier, G.; Sharon, E.; Bernhardt, M.B.; et al. Effect of Postreinduction Therapy Consolidation With Blinatumomab vs Chemotherapy on Disease-Free Survival in Children, Adolescents, and Young Adults With First Relapse of B-Cell Acute Lymphoblastic Leukemia: A Randomized Clinical Trial. JAMA 2021, 325, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Dinutuximab approved for high-risk neuroblastoma. Cancer Discov. 2015, 5, OF5. [CrossRef] [PubMed]
- Navid, F.; Santana, V.M.; Barfield, R.C. Anti-GD2 antibody therapy for GD2-expressing tumors. Curr. Cancer Drug Targets 2010, 10, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Yu, R.K.; Cheung, N.K. Ganglioside GD2 specificity of monoclonal antibodies to human neuroblastoma cell. Biochem. Biophys. Res. Commun. 1985, 127, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Uttenreuther-Fischer, M.M.; Huang, C.S.; Yu, A.L. Pharmacokinetics of human-mouse chimeric anti-GD2 mAb ch14.18 in a phase I trial in neuroblastoma patients. Cancer Immunol. Immunother. 1995, 41, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.; Mueller, B.M.; Handgretinger, R.; Herter, M.; Yu, A.L.; Reisfeld, R.A. Effect of a chimeric anti-ganglioside GD2 antibody on cell-mediated lysis of human neuroblastoma cells. Cancer Res. 1991, 51, 144–149. [Google Scholar] [PubMed]
- Ladenstein, R.; Weixler, S.; Baykan, B.; Bleeke, M.; Kunert, R.; Katinger, D.; Pribill, I.; Glander, P.; Bauer, S.; Pistoia, V.; et al. Ch14.18 antibody produced in CHO cells in relapsed or refractory Stage 4 neuroblastoma patients: A SIOPEN Phase 1 study. MAbs 2013, 5, 801–809. [Google Scholar] [CrossRef]
- Navid, F.; Sondel, P.M.; Barfield, R.; Shulkin, B.L.; Kaufman, R.A.; Allay, J.A.; Gan, J.; Hutson, P.; Seo, S.; Kim, K.; et al. Phase I trial of a novel anti-GD2 monoclonal antibody, Hu14.18K322A, designed to decrease toxicity in children with refractory or recurrent neuroblastoma. J. Clin. Oncol. 2014, 32, 1445–1452. [Google Scholar] [CrossRef]
- Albertini, M.R.; Hank, J.A.; Gadbaw, B.; Kostlevy, J.; Haldeman, J.; Schalch, H.; Gan, J.; Kim, K.; Eickhoff, J.; Gillies, S.D.; et al. Phase II trial of hu14.18-IL2 for patients with metastatic melanoma. Cancer Immunol. Immunother. 2012, 61, 2261–2271. [Google Scholar] [CrossRef]
- Cheung, I.Y.; Kushner, B.H.; Modak, S.; Basu, E.M.; Roberts, S.S.; Cheung, N.V. Phase I trial of anti-GD2 monoclonal antibody hu3F8 plus GM-CSF: Impact of body weight, immunogenicity and anti-GD2 response on pharmacokinetics and survival. Oncoimmunology 2017, 6, e1358331. [Google Scholar] [CrossRef] [PubMed]
- Tassev, D.V.; Cheung, N.K. Monoclonal antibody therapies for solid tumors. Expert Opin. Biol. Ther. 2009, 9, 341–353. [Google Scholar] [CrossRef]
- Park, J.A.; Cheung, N.V. Targets and Antibody Formats for Immunotherapy of Neuroblastoma. J. Clin. Oncol. 2020, 38, 1836–1848. [Google Scholar] [CrossRef]
- Matthay, K.K.; George, R.E.; Yu, A.L. Promising therapeutic targets in neuroblastoma. Clin. Cancer Res. 2012, 18, 2740–2753. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.K.; Cheung, I.Y.; Kushner, B.H.; Ostrovnaya, I.; Chamberlain, E.; Kramer, K.; Modak, S. Murine anti-GD2 monoclonal antibody 3F8 combined with granulocyte-macrophage colony-stimulating factor and 13-cis-retinoic acid in high-risk patients with stage 4 neuroblastoma in first remission. J. Clin. Oncol. 2012, 30, 3264–3270. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; London, W.B.; Kreissman, S.G.; Chen, H.X.; Smith, M.; Anderson, B.; Villablanca, J.G.; Matthay, K.K.; et al. Anti-GD2 antibody with GM-CSF, interleukin-2, and isotretinoin for neuroblastoma. N. Engl. J. Med. 2010, 363, 1324–1334. [Google Scholar] [CrossRef]
- Mody, R.; Naranjo, A.; Van Ryn, C.; Yu, A.L.; London, W.B.; Shulkin, B.L.; Parisi, M.T.; Servaes, S.E.; Diccianni, M.B.; Sondel, P.M.; et al. Irinotecan-temozolomide with temsirolimus or dinutuximab in children with refractory or relapsed neuroblastoma (COG ANBL1221): An open-label, randomised, phase 2 trial. Lancet Oncol. 2017, 18, 946–957. [Google Scholar] [CrossRef]
- Furman, W.L.; McCarville, B.; Shulkin, B.L.; Davidoff, A.; Krasin, M.; Hsu, C.W.; Pan, H.; Wu, J.; Brennan, R.; Bishop, M.W.; et al. Improved Outcome in Children With Newly Diagnosed High-Risk Neuroblastoma Treated With Chemoimmunotherapy: Updated Results of a Phase II Study Using hu14.18K322A. J. Clin. Oncol. 2022, 40, 335–344. [Google Scholar] [CrossRef]
- Del Bufalo, F.; De Angelis, B.; Caruana, I.; Del Baldo, G.; De Ioris, M.A.; Serra, A.; Mastronuzzi, A.; Cefalo, M.G.; Pagliara, D.; Amicucci, M.; et al. GD2-CART01 for Relapsed or Refractory High-Risk Neuroblastoma. N. Engl. J. Med. 2023, 388, 1284–1295. [Google Scholar] [CrossRef]
- Gupta, A.K.; Chopra, A.; Meena, J.P.; Singh, J.; Pandey, R.M.; Bakhshi, S.; Seth, R. Rituximab added to standard chemotherapy and its effect on minimal residual disease during induction in CD20 positive pediatric acute lymphoblastic leukemia: A pilot RCT. Am. J. Blood Res. 2021, 11, 571–579. [Google Scholar]
- FDA Expands Approval of Ipilimumab to Pediatric Patients 12 Years and Older. 2017. Available online: https://ascopost.com/issues/october-10-2017/fda-expands-approval-of-ipilimumab-to-pediatric-patients-12-years-and-older/ (accessed on 10 March 2024).
- Subbiah, V.; Solit, D.B.; Chan, T.A.; Kurzrock, R. The FDA approval of pembrolizumab for adult and pediatric patients with tumor mutational burden (TMB) >/=10: A decision centered on empowering patients and their physicians. Ann. Oncol. 2020, 31, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Hoshitsuki, K.; Zhou, Y.; Miller, A.M.; Choi, J.K.; Swanson, H.D.; Bhakta, N.H.; Jeha, S.; Karol, S.E.; Ribeiro, R.C.; Rubnitz, J.E.; et al. Rituximab administration in pediatric patients with newly diagnosed acute lymphoblastic leukemia. Leukemia 2023, 37, 1782–1791. [Google Scholar] [CrossRef]
- Merchant, M.S.; Wright, M.; Baird, K.; Wexler, L.H.; Rodriguez-Galindo, C.; Bernstein, D.; Delbrook, C.; Lodish, M.; Bishop, R.; Wolchok, J.D.; et al. Phase I Clinical Trial of Ipilimumab in Pediatric Patients with Advanced Solid Tumors. Clin. Cancer Res. 2016, 22, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Geoerger, B.; Bergeron, C.; Gore, L.; Sender, L.; Dunkel, I.J.; Herzog, C.; Brochez, L.; Cruz, O.; Nysom, K.; Berghorn, E.; et al. Phase II study of ipilimumab in adolescents with unresectable stage III or IV malignant melanoma. Eur. J. Cancer 2017, 86, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Boettcher, M.; Joechner, A.; Li, Z.; Yang, S.F.; Schlegel, P. Development of CAR T Cell Therapy in Children-A Comprehensive Overview. J. Clin. Med. 2022, 11, 2158. [Google Scholar] [CrossRef] [PubMed]
- Long, A.H.; Morgenstern, D.A.; Leruste, A.; Bourdeaut, F.; Davis, K.L. Checkpoint Immunotherapy in Pediatrics: Here, Gone, and Back Again. Am. Soc. Clin. Oncol. Educ. Book 2022, 42, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sweet-Cordero, E.A.; Biegel, J.A. The genomic landscape of pediatric cancers: Implications for diagnosis and treatment. Science 2019, 363, 1170–1175. [Google Scholar] [CrossRef]
- Diefenbach, C.S.; Hong, F.; Ambinder, R.F.; Cohen, J.B.; Robertson, M.J.; David, K.A.; Advani, R.H.; Fenske, T.S.; Barta, S.K.; Palmisiano, N.D.; et al. Ipilimumab, nivolumab, and brentuximab vedotin combination therapies in patients with relapsed or refractory Hodgkin lymphoma: Phase 1 results of an open-label, multicentre, phase 1/2 trial. Lancet Haematol. 2020, 7, e660–e670. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Frey, N.V.; Engels, B.; Barrett, D.M.; Shestova, O.; Ravikumar, P.; Cummins, K.D.; Lee, Y.G.; Pajarillo, R.; Chun, I.; et al. Antigen-independent activation enhances the efficacy of 4-1BB-costimulated CD22 CAR T cells. Nat. Med. 2021, 27, 842–850. [Google Scholar] [CrossRef]
- Jonus, H.C.; Lee, J.Y.; Silva, J.A.; Spencer, H.T.; Goldsmith, K.C. Abstract 4093: Dual targeted CAR immunotherapy for neuroblastoma using γδ T cells. Cancer Res. 2023, 83, 4093. [Google Scholar] [CrossRef]
- Dunkel, I.J.; Doz, F.; Foreman, N.K.; Hargrave, D.; Lassaletta, A.; Andre, N.; Hansford, J.R.; Hassall, T.; Eyrich, M.; Gururangan, S.; et al. Nivolumab with or without ipilimumab in pediatric patients with high-grade CNS malignancies: Safety, efficacy, biomarker, and pharmacokinetics-CheckMate 908. Neuro Oncol. 2023, 25, 1530–1545. [Google Scholar] [CrossRef]
- Turpin, B.; Vatner, R. A pilot study of pembrolizumab in combination with decitabine and hypofractionated index lesion radiation in pediatric and young adult patients with relapsed and refractory solid tumors or lymphoma. J. Clin. Oncol. 2019, 37, TPS25. [Google Scholar] [CrossRef]
- Short, S.S.; Kastenberg, Z.J.; Wei, G.; Bondoc, A.; Dasgupta, R.; Tiao, G.M.; Watters, E.; Heaton, T.E.; Lotakis, D.; La Quaglia, M.P.; et al. Histologic type predicts disparate outcomes in pediatric hepatocellular neoplasms: A Pediatric Surgical Oncology Research Collaborative study. Cancer 2022, 128, 2786–2795. [Google Scholar] [CrossRef] [PubMed]
- Vanneman, M.; Dranoff, G. Combining immunotherapy and targeted therapies in cancer treatment. Nat. Rev. Cancer 2012, 12, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Morotti, M.; Albukhari, A.; Alsaadi, A.; Artibani, M.; Brenton, J.D.; Curbishley, S.M.; Dong, T.; Dustin, M.L.; Hu, Z.; McGranahan, N.; et al. Promises and challenges of adoptive T-cell therapies for solid tumours. Br. J. Cancer 2021, 124, 1759–1776. [Google Scholar] [CrossRef] [PubMed]
- Theruvath, J.; Menard, M.; Smith, B.A.H.; Linde, M.H.; Coles, G.L.; Dalton, G.N.; Wu, W.; Kiru, L.; Delaidelli, A.; Sotillo, E.; et al. Anti-GD2 synergizes with CD47 blockade to mediate tumor eradication. Nat. Med. 2022, 28, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Mount, C.W.; Majzner, R.G.; Sundaresh, S.; Arnold, E.P.; Kadapakkam, M.; Haile, S.; Labanieh, L.; Hulleman, E.; Woo, P.J.; Rietberg, S.P.; et al. Potent antitumor efficacy of anti-GD2 CAR T cells in H3-K27M(+) diffuse midline gliomas. Nat. Med. 2018, 24, 572–579. [Google Scholar] [CrossRef]
- Upadhye, A.; Meza Landeros, K.E.; Ramirez-Suastegui, C.; Schmiedel, B.J.; Woo, E.; Chee, S.J.; Malicki, D.; Coufal, N.G.; Gonda, D.; Levy, M.L.; et al. Intra-tumoral T cells in pediatric brain tumors display clonal expansion and effector properties. Nat. Cancer 2024, 5, 791–807. [Google Scholar] [CrossRef]
Figure 1.
Dynamic tumor-immune cell interactions in the pediatric TME. The TME encompasses tumor cells alongside recruited immune cells, endothelial cells, fibroblasts, adipocytes, and non-cellular elements of the extracellular matrix. Over time, myeloid-derived suppressor cells (MDSCs), mesenchymal stromal cells (MSCs), tumor-associated macrophages (TAMs), T cells, and fibroblasts undergo education by the tumor, leading to a transition toward immune-suppressive and pro-tumorigenic functions. For instance, TAMs reprogramming toward pro-tumorigenic phenotypes by tumor cells promote myeloid-derived suppressor cell (MDSC) infiltration and proliferation, resulting in the inhibition of CTL infiltration into tumors and the upregulation of CD4+CD8+ immune-suppressive Tregs. These Tregs further support MDSC expansion, promoting diverse phenotypes within the primary tumor, including growth and metastasis. Tumor-educated immune cells secrete a plethora of pro-tumorigenic cytokines and growth factors that influence signaling in the tumor and neighboring immune cells. For example, chemokine (C-X-C motif) ligand 2 (CXCL2) secreted by TAMs upregulates cyclooxygenase-2 (COX2), matrix metalloproteinase-9 (MMP9), and STAT-3 in tumor cells, promoting tumor metastasis. IL-10 and IL-35 secretions from Tregs elevate the expression of inhibitory molecules like B lymphocyte-induced maturation protein-1 (BLIMP1), programmed cell death protein 1 (PD-1), lymphocyte-activation gene 3 (LAG3), metalloproteinase inhibitor 3 (TIMP3), and TIGIT on T and B cells, inhibiting their proliferation, activation, and tumor cytotoxicity. Similarly, cancer-associated fibroblasts (CAFs) and MSCs secrete IL-6, which increases the production of TGFβ in the TME. TGF-β1 induces the expression of IL-6 in tumor cells, promoting STAT-3 and SMAD activation, fostering the survival of TAMs, and inhibiting NK cell functions. Elevated expression of immune checkpoint molecules in tumors or antigen-presenting cells (APCs) like macrophages or dendritic cells (DCs) hinders the activation of cytotoxic NK cells and CD8+ T cells. This inhibition restricts anti-tumor responses by curtailing the activation, expansion, and infiltration of T and NK cells. Crucial to these regulatory processes is the upregulation of immune checkpoint molecules like CD200, PD-L1, CD80, B7H3, and CD20 on tumor cells, which play pivotal roles in modulating immune responses. Interactions of these molecules with their receptors on T or NK cells, such as CD200-CD200R, PD-L1-PD-1, and CD80-CTLA-4, B7-H3, and its receptor, contribute to immune suppression by inhibiting T and NK cell activation. These interactions underscore the intricate mechanisms that tumors and the immune system employ to regulate immune responses and underscore their potential as therapeutic targets in pediatric tumors. Pembrolizumab (anti-PD-1) and ipilimumab (anti-CTLA-4) have gained approval for treating pediatric cancers by modulating immune responses. Additionally, rituximab, targeting CD20, is utilized for B cell depletion in conditions such as lymphomas. These advancements in immunotherapy exemplify the progress made in treating specific cancers by targeting immune checkpoints or cell surface antigens.
Figure 1.
Dynamic tumor-immune cell interactions in the pediatric TME. The TME encompasses tumor cells alongside recruited immune cells, endothelial cells, fibroblasts, adipocytes, and non-cellular elements of the extracellular matrix. Over time, myeloid-derived suppressor cells (MDSCs), mesenchymal stromal cells (MSCs), tumor-associated macrophages (TAMs), T cells, and fibroblasts undergo education by the tumor, leading to a transition toward immune-suppressive and pro-tumorigenic functions. For instance, TAMs reprogramming toward pro-tumorigenic phenotypes by tumor cells promote myeloid-derived suppressor cell (MDSC) infiltration and proliferation, resulting in the inhibition of CTL infiltration into tumors and the upregulation of CD4+CD8+ immune-suppressive Tregs. These Tregs further support MDSC expansion, promoting diverse phenotypes within the primary tumor, including growth and metastasis. Tumor-educated immune cells secrete a plethora of pro-tumorigenic cytokines and growth factors that influence signaling in the tumor and neighboring immune cells. For example, chemokine (C-X-C motif) ligand 2 (CXCL2) secreted by TAMs upregulates cyclooxygenase-2 (COX2), matrix metalloproteinase-9 (MMP9), and STAT-3 in tumor cells, promoting tumor metastasis. IL-10 and IL-35 secretions from Tregs elevate the expression of inhibitory molecules like B lymphocyte-induced maturation protein-1 (BLIMP1), programmed cell death protein 1 (PD-1), lymphocyte-activation gene 3 (LAG3), metalloproteinase inhibitor 3 (TIMP3), and TIGIT on T and B cells, inhibiting their proliferation, activation, and tumor cytotoxicity. Similarly, cancer-associated fibroblasts (CAFs) and MSCs secrete IL-6, which increases the production of TGFβ in the TME. TGF-β1 induces the expression of IL-6 in tumor cells, promoting STAT-3 and SMAD activation, fostering the survival of TAMs, and inhibiting NK cell functions. Elevated expression of immune checkpoint molecules in tumors or antigen-presenting cells (APCs) like macrophages or dendritic cells (DCs) hinders the activation of cytotoxic NK cells and CD8+ T cells. This inhibition restricts anti-tumor responses by curtailing the activation, expansion, and infiltration of T and NK cells. Crucial to these regulatory processes is the upregulation of immune checkpoint molecules like CD200, PD-L1, CD80, B7H3, and CD20 on tumor cells, which play pivotal roles in modulating immune responses. Interactions of these molecules with their receptors on T or NK cells, such as CD200-CD200R, PD-L1-PD-1, and CD80-CTLA-4, B7-H3, and its receptor, contribute to immune suppression by inhibiting T and NK cell activation. These interactions underscore the intricate mechanisms that tumors and the immune system employ to regulate immune responses and underscore their potential as therapeutic targets in pediatric tumors. Pembrolizumab (anti-PD-1) and ipilimumab (anti-CTLA-4) have gained approval for treating pediatric cancers by modulating immune responses. Additionally, rituximab, targeting CD20, is utilized for B cell depletion in conditions such as lymphomas. These advancements in immunotherapy exemplify the progress made in treating specific cancers by targeting immune checkpoints or cell surface antigens.
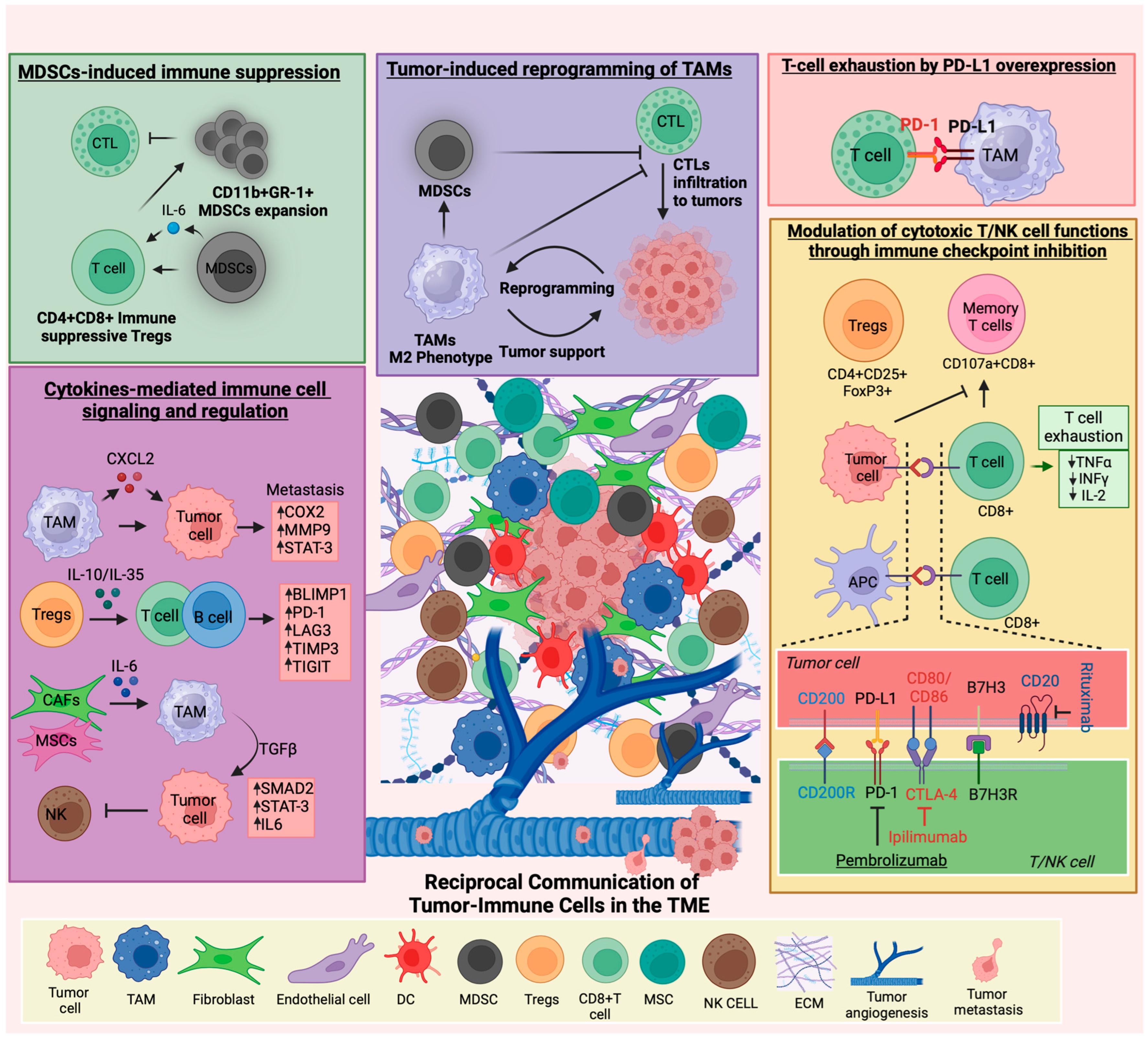

{kind=link}
Table 1.
Cytokines in the pediatric tumor microenvironment and their roles and potential as predictive biomarkers.
Table 1.
Cytokines in the pediatric tumor microenvironment and their roles and potential as predictive biomarkers.
| Cytokines and Their Receptors | Tumor Type | Cell Source | Prognosis and Correlation with Treatment | Role in Tumor Progression | Mechanisms |
|---|---|---|---|---|---|
| Hematological malignancies: | |||||
| TNFα [90,98] | AML | Blast cells, LSC, TAMs | Worse OS | Promoting | Promotes tumor inflammation by activating NF-kB, PI3K/AKT, and MAPK signaling |
| IL3Rα (CD123) [99,100] | B-ALL | Blast cells | Good OS | Promoting | Alters CXCR4/SDF-1 interaction in the BM, promotes AML cell growth |
| IL-7Rα [101,102,103] | T-ALL | T cells | Worse OS | Promoting | Promotes JAK, STAT-5, PI3K signaling |
| IL-2 [104,105,106,107] | AML | T cells | Good OS | Inhibitory | Stimulates CTLs production |
| IL-6 [108,109] | AML | Blast cells, BMSCs | Worse OS | Promoting | IL-6-induced STAT3 pathway promotes AML progression |
| IL-10 [110,111,112,113] | ALL | Myeloid, lymphoid cells | Good OS | Inhibitory | IL-10 family cytokines maintain tissue balance by regulating inflammation, enhancing immunity, and aiding tissue repair. Children with ALL have low IL-10 levels. IL-10 inhibits AML blast proliferation, while its deficiency impairs B-cell development, increases DNA damage, and raises proinflammatory cytokines IL-1α, IL-6, IL-12p40, IL-13, MIP-1β/CCL4, and G-CSF. |
| IL-33, IL1RL1 [114,115,116] | AML | AML, BM, mast, ILC2, Tregs and TAM cells | Worse OS | Promoting | Activates p38 MAPK, Wnt, and Notch pathways, promoting cell survival and stemness |
| IL1β [92,117,118,119] | AML/ALL | Leukemia cells, monocytes | Worse OS | Promoting | Activates IL-1/p38MAPK pathway and promotes leukemia progression |
| Solid tumors: | |||||
| IL-1β and TNF-α [120] | NB | TAMs | Worse OS | Promoting | Promotes ARG2 expression via p38/ERK signaling |
| IL-2 [121,122,123,124] | NB | T and NK cells | Good OS | Inhibitory | promote CD8+ T, B, and NK cell cytotoxicity activity |
| IL-6 [125,126] | NB | BMSCs, MSC, CAFs, NB cells | Worse OS | Promoting | Promotes STAT-3, ERK signaling, inhibits NK functions |
| IL-6 [87,127,128] | Glioma | Glial, stromal, or TME cells | Worse OS | Promoting | Promotes oncogenic JAK/STAT3 signaling |
| IL-8 [129,130,131,132] | Osteosarcoma | Osteosarcoma cells, MSC, TAMs | Worse OS | Promoting | Promotes FAK signaling, ABCB1/MDR1 pathway and immune suppression |
| IL-10 [87,133,134] | MB, GCT, Glioma | Bregs | Worse OS | Promoting | IL-10 suppresses T and NK cell anti-tumor immunity and promotes cell proliferation. |
| IL-6, IL-10 [87,135,136] | MB | MB cells | Worse OS | Promoting | IL-6 induces STAT3 pathway and promotes MB growth |
| IL-12 [137,138,139] | Osteosarcoma | APCs | Good OS | Inhibiting | Activates NK, CTLs, and memory T cells, inhibiting tumor growth and metastasis |
| IL-13Rα2 [140,141,142] | Glioma | Glioma cells | Worse OS | Promoting | IL-13 and its receptor IL-13Rα2 signals through JAK/STAT and AP-1 pathways |
| TGF-β [126,143] | NB | CAFs, TAMs | Worse OS | Promoting | Activates TGF-β/IL-6 pathway in NB and MSC, promotes immune suppression |
| VEGF [144,145,146,147] | ES | ES cells | Worse OS | Promoting | Attracts BM-derived EPCs to drive tumor blood vessel growth |
| VEGF [148,149,150] | WT | WT and stromal cells | Worse OS | Promoting | Regulates angiogenesis by activating VEGFR-1 and VEGFR-2 |
| AFP [151,152,153,154] | Hepato blastoma | Liver cancer cells | Worse OS | Promoting | Promotes PI3K/AKT, suppresses Fas/FADD apoptotic pathway |
AML: Acute myeloid leukemia, B-ALL: B-cell acute lymphoblastic leukemia, T-ALL: T-cell acute lymphoblastic leukemia, ALL: Acute lymphoblastic leukemia, NB: Neuroblastoma, MB: Medulloblastoma, GCT: Germ Cell Tumors, ES: Ewing Sarcoma, WT: Wilms tumor. OS: Overall survival, TNFα: Tumor necrosis factor α, LSC: Leukemia stem cells, TAM: Tumor-associated macrophages, IL: Interleukin, IL-3RA: Interleukin-3 Receptor Alpha Chain, SDF-1: Stromal-derived factor-1, CTLs: Cytotoxic T lymphocytes, MIP-1β/CCL4: Macrophage inflammatory protein-1β/CCL4, G-CSF: Granulocyte colony-stimulating factor, BM: Bone marrow, ILC2s: Group 2 innate lymphoid cells, Tregs: T regulatory cells, IL1RL1: Interleukin 1 receptor-like 1, ARG2: arginase 2, BMSCs: Bone marrow stromal cells, MSC: Mesenchymal stromal cells, CAF: Cancer-associated fibroblasts, IL-13Rα2: Interleukin-13 receptor alpha 2, Bregs: B regulatory cells (CD19+ IL-10+ or CD19+ CD24hiCD38hi), APC: Antigen-presenting cells, VEGF: Vascular endothelial growth factor, EPCs: Endothelial progenitor cells, VEGFR: Vascular endothelial growth factor receptor, AFP: Alpha-fetoprotein.
Table 2.
Clinical trials investigating the use of targeted mAbs and adoptive T-cell therapies, either in combination with standard cancer treatments or as single agents, for the treatment of pediatric cancers.
Table 2.
Clinical trials investigating the use of targeted mAbs and adoptive T-cell therapies, either in combination with standard cancer treatments or as single agents, for the treatment of pediatric cancers.
| Cancer Type | NCT; Phase | Monoclonal Antibodies or Immune Cells | Target | Combination Drugs | Status and Outcome |
|---|---|---|---|---|---|
| Hematological malignancies: | |||||
| Relapsed/Refractory Mature B-NHL | NCT05533775; I/II | Glofitamab | CD20 | Rituximab, Ifosfamide, Carboplatin, Etoposide | Recruiting |
| Ph-like ALL | NCT03571321; I | Ruxolitinib | JAK1 JAK2 | Rituximab, Cyclophosphamide, Cytarabine, Mercaptopurine, Vincristine, Pegaspargase, Methotrexate, dexamethasone, Doxorubicin, Thioguanine | Recruiting |
| Relapsed B-ALL | NCT05645718; II | Inotuzumab Ozogamicin, Blinatumomab and Rituximab | Tumor cells | Cyclophosphamide, Vincristine, and Dexamethasone | Recruiting |
| Aggressive B-cell Lymphoma | NCT03864419; I | Rituximab Hyaluronidase | CD20 | Cyclophosphamide, Vincristine, Methotrexate, Etoposide, Doxorubicin, Prednisone | Completed |
| B-ALL | NCT03150693; III | Inotuzumab Ozogamicin | CD22 | Rituximab, Allopurinol, Cytarabine, Daunorubicin, Vincristine, Dexamethasone, PLA, Methotrexate, Cyclophosphamide, Mercaptopurine, Doxorubicin, Thioguanine | Suspended due to high toxicity |
| Relapsed/Refractory HL [190] | NCT01896999; I/II | Brentuximab vedotin | CD30 | Nivolumab and Ipilimumab | CRR improved, Suspended |
| PD-L1+ve solid tumors, lymphoma [59] | NCT02332668; I/II | Pembrolizumab | PD-1 | Single agent | Recruiting |
| Relapsed/Refractory B-ALL or B-NHL | NCT04544592; I/II NCT04173988; I | CAR-T cells | CD19 | Single agent | Recruiting Not Recruiting |
| Relapsed/Refractory B-ALL [191] | NCT02650414; I/II | CAR-T cells | CD22 | Single agent | Recruiting |
| Relapsed/Refractory HL | NCT04268706; II | CAR-T cells | CD30 | Single agent | Recruiting |
| Relapsed/Refractory B-ALL, B-NHL | NCT03743246; I/II | JCAR017 (CAR-T cells) | CD19 | Fludarabine, Cyclophosphamide | Completed |
| Relapsed/ Refractory pre-B ALL | NCT03605589; I | Pembrolizumab | PD-1 | blinatumomab | Withdrawn due to low enrollment |
| Solid tumors: | |||||
| Relapsed/ Refractory NB | NCT04238819; 1b/2 | Abemaciclib | CDK4, CDK6 | Dinutuximab, GM-CSF, Irinotecan, Temozolomide | Active, Not recruiting |
| NCT03794349; II | Eflornithine | polyamines | Irinotecan, Temozolomide, Dinutuximab | Recruiting | |
| NCT02914405; I | Dinutuximab | GD2 | mlBG, Nivolumab (anti-PD1) antibody | Recruiting | |
| NCT05400603; I [192] | Allogeneic γδ T Cells | Tumor cells | Temozolomide, Irinotecan, Dinutuximab, Zoledronate | Recruiting | |
| INI1(-) or SMARCA4-def. tumors | NCT05407441; I/II | tazemetostat | EZH2 | Nivolumab and Ipilimumab | Recruiting |
| Recurrent or Progressive HGG | NCT04323046; I | nivolumab | PD-1 | Single agent. Given before and after surgery | Recruiting |
| High-grade primary CNS malignancies [193] | NCT03130959; 1b/II | Nivolumab | PD-1 | Ipilimumab | No clinical benefit in combination |
| Relapsed/Refractory solid tumors | NCT05302921; II | Nivolumab, Ipilimumab | PD-1, CTLA-4 | cryoablation therapy | Active, not recruiting |
| Relapsed, Refractory, or Progressive CNS solid tumors and lymphomas [194] | NCT03445858; I | Pembrolizumab | PD-1 | Decitabine and radiation therapy | Active, not recruiting |
| Refractory gliomas, MB | NCT02359565; I | Pembrolizumab | PD-1 | Single agent | Recruiting |
| Liver cancer [195] | NCT04134559; II | Pembrolizumab | PD-1 | Single agent | Recruiting |
| GD2+ve Brain tumors | NCT04099797; I | C7R-GD2.CAR T Cells | GD2 | Single agent | Recruiting |
NHL: Non-Hodgkin Lymphoma, ALL: Acute Lymphoblastic Leukemia, B-ALL: B-Cell Acute Lymphoblastic Leukemia, HL: Hodgkin Lymphoma, B-NHL: B-cell Non-Hodgkin Lymphoma, NB: Neuroblastoma, INI1(-): INI1 negative, SMARCA4-def: SMARCA4 deficient, HGG: High-Grade Glioma, CNS: central nervous system, MB: medulloblastoma, PLA: Pegylated L-Asparaginase, EZH2: Enhancer of zeste homolog 2, JAK: Janus kinase, mlBG: 131-l Metaiodobenzylguanidine, CRR: complete response rate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pathania, A.S. Immune Microenvironment in Childhood Cancers: Characteristics and Therapeutic Challenges. Cancers 2024, 16, 2201. https://doi.org/10.3390/cancers16122201
AMA Style
Pathania AS. Immune Microenvironment in Childhood Cancers: Characteristics and Therapeutic Challenges. Cancers. 2024; 16(12):2201. https://doi.org/10.3390/cancers16122201
Chicago/Turabian StylePathania, Anup Singh. 2024. "Immune Microenvironment in Childhood Cancers: Characteristics and Therapeutic Challenges" Cancers 16, no. 12: 2201. https://doi.org/10.3390/cancers16122201
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.