
Chronobiology of Cancers in the Liver and Gut


Abstract
Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
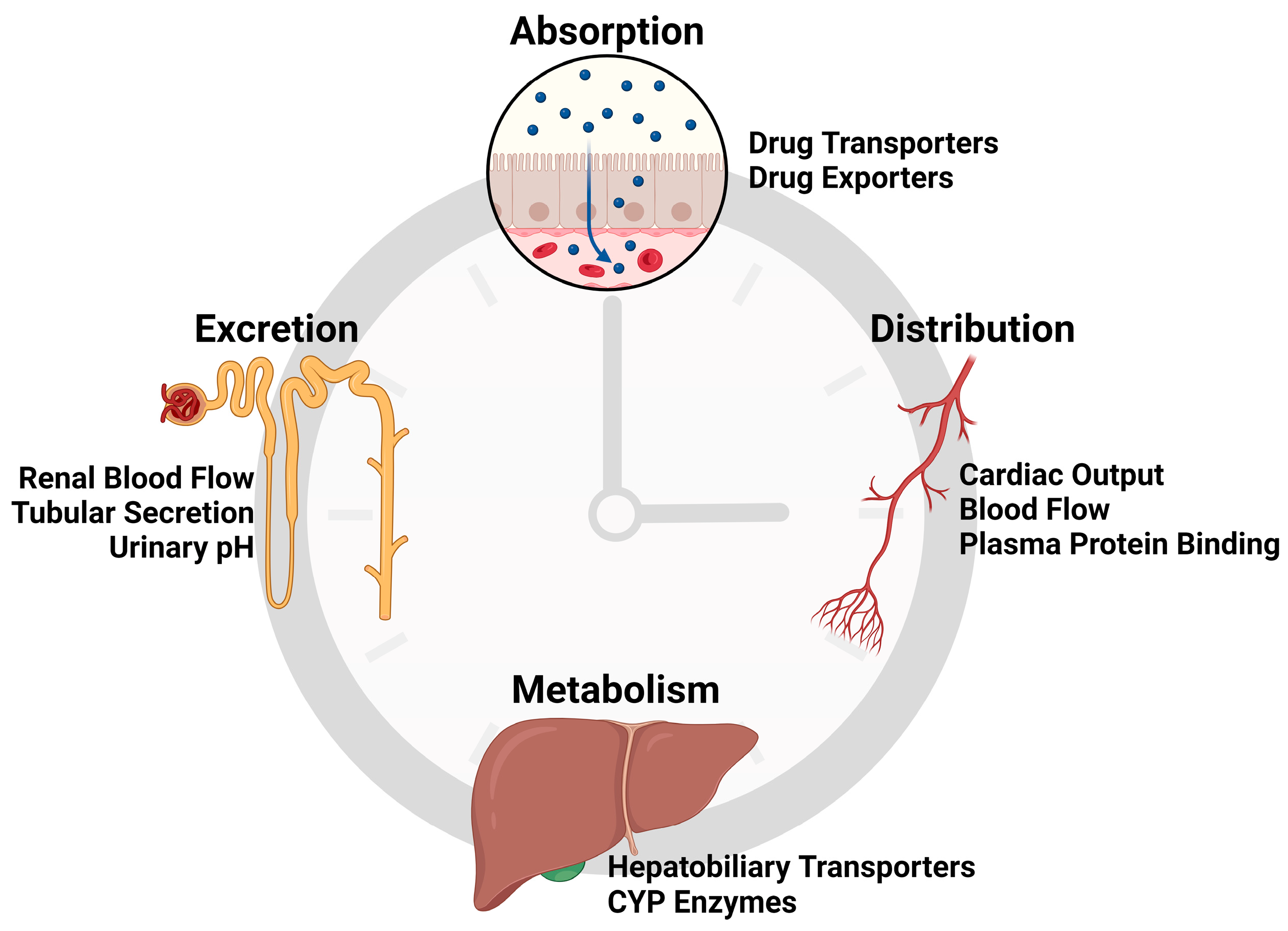
1. Introduction
2. Circadian Rhythms in the Liver and Gut
Consequences of Rhythm Disruption
3. Circadian Rhythms and Cancers of the Liver and Gut
3.1. Hepatocellular Carcinoma
3.2. Cholangiocarcinoma and Cholangitis
3.3. Colorectal Cancer
4. Chronotherapy for Cancers of the Liver and Gut
4.1. Chronotherapy in the Liver
4.2. Chronotherapy in the Gut
4.3. Clinical Applications
5. Conclusions
Funding
Conflicts of Interest
References
- Novak, C.M.; Ehlen, J.C.; Albers, H.E. Photic and Nonphotic Inputs to the Diurnal Circadian Clock. Biol. Rhythm Res. 2008, 39, 291–304. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kantermann, T.; Juda, M.; Vetter, C.; Allebrandt, K.V. Light and the Human Circadian Clock BT—Circadian Clocks; Kramer, A., Merrow, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 311–331. ISBN 978-3-642-25950-0. [Google Scholar]
- Hastings, M.H.; Brancaccio, M.; Maywood, E.S. Circadian Pacemaking in Cells and Circuits of the Suprachiasmatic Nucleus. J. Neuroendocrinol. 2014, 26, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Gut Clock: Implication of Circadian Rhythms in the Gastrointestinal Tract. J. Physiol. Pharmacol. 2011, 62, 139–150. [Google Scholar]
- Reinke, H.; Asher, G. Circadian Clock Control of Liver Metabolic Functions. Gastroenterology 2016, 150, 574–580. [Google Scholar] [CrossRef]
- Fishbein, A.B.; Knutson, K.L.; Zee, P.C. Circadian Disruption and Human Health. J. Clin. Investig. 2021, 131, e148286. [Google Scholar] [CrossRef]
- Wang, Y.; Song, L.; Liu, M.; Ge, R.; Zhou, Q.; Liu, W.; Li, R.; Qie, J.; Zhen, B.; Wang, Y.; et al. A Proteomics Landscape of Circadian Clock in Mouse Liver. Nat. Commun. 2018, 9, 1553. [Google Scholar] [CrossRef]
- Weger, B.D.; Gobet, C.; David, F.P.A.; Atger, F.; Martin, E.; Phillips, N.E.; Charpagne, A.; Weger, M.; Naef, F.; Gachon, F. Systematic Analysis of Differential Rhythmic Liver Gene Expression Mediated by the Circadian Clock and Feeding Rhythms. Proc. Natl. Acad. Sci. USA 2021, 118, e2015803118. [Google Scholar] [CrossRef] [PubMed]
- Hurni, C.; Weger, B.D.; Gobet, C.; Naef, F. Comprehensive Analysis of the Circadian Nuclear and Cytoplasmic Transcriptome in Mouse Liver. PLOS Genet. 2022, 18, e1009903. [Google Scholar] [CrossRef] [PubMed]
- Barber, L.E.; VoPham, T.; White, L.F.; Roy, H.K.; Palmer, J.R.; Bertrand, K.A. Circadian Disruption and Colorectal Cancer Incidence in Black Women. Cancer Epidemiol. Biomark. Prev. 2023, 32, 927–935. [Google Scholar] [CrossRef]
- He, C.; Anand, S.T.; Ebell, M.H.; Vena, J.E.; Robb, S.W. Circadian Disrupting Exposures and Breast Cancer Risk: A Meta-Analysis. Int. Arch. Occup. Environ. Health 2015, 88, 533–547. [Google Scholar] [CrossRef]
- Wichert, K.; Rabstein, S.; Stang, A.; Erbel, R.; Eisele, L.; Arendt, M.; Keimer, A.; Dragano, N.; Hoffmann, W.; Lerch, M.M.; et al. Associations between Shift Work and Risk of Colorectal Cancer in Two German Cohort Studies. Chronobiol. Int. 2020, 37, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Lorca, M.; Olmedo-Requena, R.; Vega-Galindo, M.-V.; Vázquez-Alonso, F.; Jiménez-Pacheco, A.; Salcedo-Bellido, I.; Sánchez, M.-J.; Jiménez-Moleón, J.-J. Night Shift Work, Chronotype, Sleep Duration, and Prostate Cancer Risk: CAPLIFE Study. Int. J. Environ. Res. Public Health 2020, 17, 6300. [Google Scholar] [CrossRef]
- Cordina-Duverger, E.; Menegaux, F.; Popa, A.; Rabstein, S.; Harth, V.; Pesch, B.; Brüning, T.; Fritschi, L.; Glass, D.C.; Heyworth, J.S.; et al. Night Shift Work and Breast Cancer: A Pooled Analysis of Population-Based Case–Control Studies with Complete Work History. Eur. J. Epidemiol. 2018, 33, 369–379. [Google Scholar] [CrossRef]
- Rivera, A.S.; Akanbi, M.; O’Dwyer, L.C.; McHugh, M. Shift Work and Long Work Hours and Their Association with Chronic Health Conditions: A Systematic Review of Systematic Reviews with Meta-Analyses. PLoS ONE 2020, 15, e0231037. [Google Scholar] [CrossRef]
- Manouchehri, E.; Taghipour, A.; Ghavami, V.; Ebadi, A.; Homaei, F.; Latifnejad Roudsari, R. Night-Shift Work Duration and Breast Cancer Risk: An Updated Systematic Review and Meta-Analysis. BMC Womens. Health 2021, 21, 89. [Google Scholar] [CrossRef]
- Sweeney, M.R.; Sandler, D.P.; Niehoff, N.M.; White, A.J. Shift Work and Working at Night in Relation to Breast Cancer Incidence. Cancer Epidemiol. Biomark. Prev. 2020, 29, 687–689. [Google Scholar] [CrossRef]
- Gan, Y.; Li, L.; Zhang, L.; Yan, S.; Gao, C.; Hu, S.; Qiao, Y.; Tang, S.; Wang, C.; Lu, Z. Association between Shift Work and Risk of Prostate Cancer: A Systematic Review and Meta-Analysis of Observational Studies. Carcinogenesis 2018, 39, 87–97. [Google Scholar] [CrossRef]
- Papantoniou, K.; Devore, E.E.; Massa, J.; Strohmaier, S.; Vetter, C.; Yang, L.; Shi, Y.; Giovannucci, E.; Speizer, F.; Schernhammer, E.S. Rotating Night Shift Work and Colorectal Cancer Risk in the Nurses’ Health Studies. Int. J. Cancer 2018, 143, 2709–2717. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, Z.; Dong, D.; Sun, L.; Zhang, G. Sex Differences in the Association between Night Shift Work and the Risk of Cancers: A Meta-Analysis of 57 Articles. Dis. Markers 2018, 2018, 925219. [Google Scholar] [CrossRef] [PubMed]
- Dun, A.; Zhao, X.; Jin, X.; Wei, T.; Gao, X.; Wang, Y.; Hou, H. Association Between Night-Shift Work and Cancer Risk: Updated Systematic Review and Meta-Analysis. Front. Oncol. 2020, 10, 1006. [Google Scholar]
- McElvenny, D.M.; Crawford, J.O.; Davis, A.; Dixon, K.; Alexander, C.; Cowie, H.; Cherrie, J.W. A Review of the Impact of Shift-Work on Occupational Cancer: Part 1-Epidemiological Research. Policy Pract. Health Saf. 2018, 16, 71–108. [Google Scholar] [CrossRef]
- Ward, E.M.; Germolec, D.; Kogevinas, M.; McCormick, D.; Vermeulen, R.; Anisimov, V.N.; Aronson, K.J.; Bhatti, P.; Cocco, P.; Costa, G.; et al. Carcinogenicity of Night Shift Work. Lancet Oncol. 2019, 20, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Xu, S.; Devesa, S.S.; Zhang, F.; Klerman, E.B.; Graubard, B.I.; Caporaso, N.E. Longitude Position in a Time Zone and Cancer Risk in the United States. Cancer Epidemiol. Biomark. Prev. 2017, 26, 1306–1311. [Google Scholar] [CrossRef]
- VoPham, T.; Weaver, M.D.; Vetter, C.; Hart, J.E.; Tamimi, R.M.; Laden, F.; Bertrand, K.A. Circadian Misalignment and Hepatocellular Carcinoma Incidence in the United States. Cancer Epidemiol. Biomark. Prev. 2018, 27, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Brown, C.; Law, M.; Colacino, J.A.; Ritov, Y. Longitudinal Position and Cancer Risk in the United States Revisited. Cancer Res. Commun. 2024, 4, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Wong, G.; Anstee, Q.M.; Henry, L. The Global Burden of Liver Disease. Clin. Gastroenterol. Hepatol. 2023, 21, 1978–1991. [Google Scholar] [CrossRef]
- Wu, Z.; Hu, H.; Zhang, Q.; Wang, T.; Li, H.; Qin, Y.; Ai, X.; Yi, W.; Wei, X.; Gao, W.; et al. Four Circadian Rhythm-Related Genes Predict Incidence and Prognosis in Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 937403. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, C.; Wang, X.; Zhao, W.; Li, Y.; He, X.; Chen, Y.; Liu, J.; Wu, X.; Shen, A. Development and Validation of a Novel Circadian Rhythm-Related Signature to Predict the Prognosis of the Patients with Hepatocellular Carcinoma. Biomed. Res. Int. 2022, 2022, 4263261. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, S.; Huang, X.; Chai, R.; Tang, Q.; Yang, R.; Huang, X.; Wang, X.; Zheng, K. Dysregulation of Circadian Clock Genes as Significant Clinic Factor in the Tumorigenesis of Hepatocellular Carcinoma. Comput. Math. Methods Med. 2021, 2021, 8238833. [Google Scholar] [CrossRef]
- Lin, Y.-M.; Chang, J.H.; Yeh, K.-T.; Yang, M.-Y.; Liu, T.-C.; Lin, S.-F.; Su, W.-W.; Chang, J.-G. Disturbance of Circadian Gene Expression in Hepatocellular Carcinoma. Mol. Carcinog. 2008, 47, 925–933. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, F.; Zhou, F.; Chen, Y.; Wang, X.; Zhang, H.; Zhu, Y.; Bi, J.; Zhang, Y. Functional Polymorphisms of Circadian Negative Feedback Regulation Genes Are Associated with Clinical Outcome in Hepatocellular Carcinoma Patients Receiving Radical Resection. Med. Oncol. 2014, 31, 179. [Google Scholar] [CrossRef] [PubMed]
- Bodard, S.; Liu, Y.; Guinebert, S.; Yousra, K.; Asselah, T. Prognostic Value of Genotyping in Hepatocellular Carcinoma: A Systematic Review. J. Viral Hepat. 2023, 30, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Kettner, N.M.; Voicu, H.; Finegold, M.J.; Coarfa, C.; Sreekumar, A.; Putluri, N.; Katchy, C.A.; Lee, C.; Moore, D.D.; Fu, L. Circadian Homeostasis of Liver Metabolism Suppresses Hepatocarcinogenesis. Cancer Cell 2016, 30, 909–924. [Google Scholar] [CrossRef]
- Qu, M.; Zhang, G.; Qu, H.; Vu, A.; Wu, R.; Tsukamoto, H.; Jia, Z.; Huang, W.; Lenz, H.-J.; Rich, J.N.; et al. Circadian Regulator BMAL1::CLOCK Promotes Cell Proliferation in Hepatocellular Carcinoma by Controlling Apoptosis and Cell Cycle. Proc. Natl. Acad. Sci. USA 2023, 120, e2214829120. [Google Scholar] [CrossRef]
- Jiang, H.; Garcia, V.; Yanum, J.A.; Lee, J.; Dai, G. Circadian Clock Core Component Bmal1 Dictates Cell Cycle Rhythm of Proliferating Hepatocytes during Liver Regeneration. Am. J. Physiol. Liver Physiol. 2021, 321, G389–G399. [Google Scholar] [CrossRef]
- Hassan, S.A.; Schmithals, C.; von Harten, M.; Piiper, A.; Korf, H.-W.; von Gall, C. Time-Dependent Changes in Proliferation, DNA Damage and Clock Gene Expression in Hepatocellular Carcinoma and Healthy Liver of a Transgenic Mouse Model. Int. J. Cancer 2021, 148, 226–237. [Google Scholar] [CrossRef]
- Yang, X.; Wang, J.; Wang, H. Association between Sleep Traits and Primary Liver Cancer: A Mendelian Randomization Analysis. Eur. J. Clin. Investig. 2023, 53, e14002. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Meng, F.; Venter, J.; Wu, N.; Wan, Y.; Standeford, H.; Francis, H.; Meininger, C.; Greene, J.; Trzeciakowski, J.P.; et al. MiR-34a-Dependent Overexpression of Per1 Decreases Cholangiocarcinoma Growth. J. Hepatol. 2016, 64, 1295–1304. [Google Scholar] [CrossRef]
- Mteyrek, A.; Filipski, E.; Guettier, C.; Oklejewicz, M.; van der Horst, G.T.J.; Okyar, A.; Lévi, F. Critical Cholangiocarcinogenesis Control by Cryptochrome Clock Genes. Int. J. Cancer 2017, 140, 2473–2483. [Google Scholar] [CrossRef]
- Hrad, V.; Abebe, Y.; Ali, S.H.; Velgersdyk, J.; Al Hallak, M.; Imam, M. Risk and Surveillance of Cancers in Primary Biliary Tract Disease. Gastroenterol. Res. Pract. 2016, 2016, 3432640. [Google Scholar] [CrossRef]
- Lavery, M.J.; Stull, C.; Kinney, M.O.; Yosipovitch, G. Nocturnal Pruritus: The Battle for a Peaceful Night’s Sleep. Int. J. Mol. Sci. 2016, 17, 425. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.; Cazzagon, N.; Franceschet, I.; Formentin, C.; Frighetto, G.; Giordani, F.; Cellini, N.; Mazzotta, G.; Costa, R.; Middleton, B.; et al. Morning Bright Light Treatment for Sleep-Wake Disturbances in Primary Biliary Cholangitis: A Pilot Study. Front. Physiol. 2018, 9, 1530. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, T.; Wu, N.; O’Brien, A.; Venter, J.; Ceci, L.; Kyritsi, K.; Onori, P.; Gaudio, E.; Sybenga, A.; et al. Pinealectomy or Light Exposure Exacerbates Biliary Damage and Liver Fibrosis in Cholestatic Rats through Decreased Melatonin Synthesis. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1525–1539. [Google Scholar] [CrossRef]
- Han, Y.; Onori, P.; Meng, F.; DeMorrow, S.; Venter, J.; Francis, H.; Franchitto, A.; Ray, D.; Kennedy, L.; Greene, J.; et al. Prolonged Exposure of Cholestatic Rats to Complete Dark Inhibits Biliary Hyperplasia and Liver Fibrosis. Am. J. Physiol. Liver Physiol. 2014, 307, G894–G904. [Google Scholar] [CrossRef] [PubMed]
- Alsakarneh, S.; Jaber, F.; Beran, A.; Aldiabat, M.; Abboud, Y.; Hassan, N.; Abdallah, M.; Abdelfattah, T.; Numan, L.; Clarkston, W.; et al. The National Burden of Colorectal Cancer in the United States from 1990 to 2019. Cancers 2024, 16, 205. [Google Scholar] [CrossRef]
- Ilic, M.; Ilic, I. Cancer of Colon, Rectum and Anus: The Rising Burden of Disease Worldwide from 1990 to 2019. J. Public Health 2024, 46, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Martchenko, A.; Martchenko, S.E.; Biancolin, A.D.; Brubaker, P.L. Circadian Rhythms and the Gastrointestinal Tract: Relationship to Metabolism and Gut Hormones. Endocrinology 2020, 161, bqaa167. [Google Scholar] [CrossRef]
- Wang, X.; Ji, A.; Zhu, Y.; Liang, Z.; Wu, J.; Li, S.; Meng, S.; Zheng, X.; Xie, L. A Meta-Analysis Including Dose-Response Relationship between Night Shift Work and the Risk of Colorectal Cancer. Oncotarget 2015, 6, 25046. [Google Scholar] [CrossRef]
- Tran, L.; Jochum, S.B.; Shaikh, M.; Wilber, S.; Zhang, L.; Hayden, D.M.; Forsyth, C.B.; Voigt, R.M.; Bishehsari, F.; Keshavarzian, A.; et al. Circadian Misalignment by Environmental Light/Dark Shifting Causes Circadian Disruption in Colon. PLoS ONE 2021, 16, e0251604. [Google Scholar] [CrossRef]
- Lee, D.-B.; An, S.-Y.; Pyo, S.-S.; Kim, J.; Kim, S.-W.; Yoon, D.-W. Sleep Fragmentation Accelerates Carcinogenesis in a Chemical-Induced Colon Cancer Model. Int. J. Mol. Sci. 2023, 24, 4547. [Google Scholar] [CrossRef]
- Stokes, K.; Nunes, M.; Trombley, C.; Flôres, D.E.F.L.; Wu, G.; Taleb, Z.; Alkhateeb, A.; Banskota, S.; Harris, C.; Love, O.P.; et al. The Circadian Clock Gene, Bmal1, Regulates Intestinal Stem Cell Signaling and Represses Tumor Initiation. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1847–1872.e0. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Devocelle, A.; Souza, L.; Foudi, A.; Bento, S.T.; Desterke, C.; Sherrard, R.; Ballesta, A.; Adam, R.; Giron-Michel, J.; et al. BMAL1 Knockdown Triggers Different Colon Carcinoma Cell Fates by Altering the Delicate Equilibrium between AKT/MTOR and P53/P21 Pathways. Aging 2020, 12, 8067–8083. [Google Scholar] [CrossRef]
- Dong, P.; Wang, Y.; Liu, Y.; Zhu, C.; Lin, J.; Qian, R.; Hua, L.; Lu, C. BMAL1 Induces Colorectal Cancer Metastasis by Stimulating Exosome Secretion. Mol. Biol. Rep. 2022, 49, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Sahar, N.; Qadir, J.; Riaz, S.K.; Sultan, A.; Arif, A.; Malik, M.F.A. Dysregulation of Core Circadian Genes, BMAL1 and CLOCK, in Colorectal Cancer. Biol. Rhythm Res. 2022, 53, 1400–1413. [Google Scholar] [CrossRef]
- Zhang, Y.; Devocelle, A.; Desterke, C.; de Souza, L.E.; Hadadi, É.; Acloque, H.; Foudi, A.; Xiang, Y.; Ballesta, A.; Chang, Y.; et al. BMAL1 Knockdown Leans Epithelial–Mesenchymal Balance toward Epithelial Properties and Decreases the Chemoresistance of Colon Carcinoma Cells. Int. J. Mol. Sci. 2021, 22, 5247. [Google Scholar] [CrossRef]
- He, Y.; Chen, Y.; Dai, X.; Huang, S. Dysregulation of Circadian Clock Genes Associated with Tumor Immunity and Prognosis in Patients with Colon Cancer. Comput. Math. Methods Med. 2022, 2022, 4957996. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-L.; Xu, X.; Rixiati, Y.; Wang, C.-Y.; Ni, H.-L.; Chen, W.-S.; Gong, H.-M.; Zhang, Z.-L.; Li, S.; Shen, T.; et al. Dysfunctional Circadian Clock Accelerates Cancer Metastasis by Intestinal Microbiota Triggering Accumulation of Myeloid-Derived Suppressor Cells. Cell Metab. 2024, 36, 1320–1334.e9. [Google Scholar] [CrossRef]
- Marjot, T.; Tomlinson, J.W.; Hodson, L.; Ray, D.W. Timing of Energy Intake and the Therapeutic Potential of Intermittent Fasting and Time-Restricted Eating in NAFLD. Gut 2023, 72, 1607–1619. [Google Scholar] [CrossRef]
- Bishehsari, F.; Engen, P.A.; Voigt, R.M.; Swanson, G.; Shaikh, M.; Wilber, S.; Naqib, A.; Green, S.J.; Shetuni, B.; Forsyth, C.B.; et al. Abnormal Eating Patterns Cause Circadian Disruption and Promote Alcohol-Associated Colon Carcinogenesis. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 219–237. [Google Scholar] [CrossRef]
- González-Mercado, V.J.; Sarkar, A.; Penedo, F.J.; Pérez-Santiago, J.; McMillan, S.; Marrero, S.J.; Marrero-Falcón, M.A.; Munro, C.L. Gut Microbiota Perturbation Is Associated with Acute Sleep Disturbance among Rectal Cancer Patients. J. Sleep Res. 2020, 29, e12915. [Google Scholar] [CrossRef]
- Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Circadian Rhythms: A Regulator of Gastrointestinal Health and Dysfunction. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 411–424. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, T.; Zhou, C.; Xu, H.; Guo, L.; Chen, M.; Wu, B. The Circadian Clock Gene Bmal1 Controls Intestinal Exporter MRP2 and Drug Disposition. Theranostics 2019, 9, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, M.; Koyanagi, S.; Suzuki, N.; Katamune, C.; Matsunaga, N.; Watanabe, N.; Takahashi, M.; Izumi, T.; Ohdo, S. Circadian Modulation in the Intestinal Absorption of P-Glycoprotein Substrates in Monkeys. Mol. Pharmacol. 2015, 88, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Eckerbom, P.; Hansell, P.; Cox, E.; Buchanan, C.; Weis, J.; Palm, F.; Francis, S.; Liss, P. Circadian Variation in Renal Blood Flow and Kidney Function in Healthy Volunteers Monitored with Noninvasive Magnetic Resonance Imaging. Am. J. Physiol. Physiol. 2020, 319, F966–F978. [Google Scholar] [CrossRef]
- Zhao, M.; Zhao, H.; Deng, J.; Guo, L.; Wu, B. Role of the CLOCK Protein in Liver Detoxification. Br. J. Pharmacol. 2019, 176, 4639–4652. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, S.; Zhou, Z.; Guo, L.; Yu, F.; Wu, B. Bmal1 Regulates Circadian Expression of Cytochrome P450 3a11 and Drug Metabolism in Mice. Commun. Biol. 2019, 2, 378. [Google Scholar] [CrossRef]
- Ge, W.; Wang, T.; Zhao, Y.; Yang, Y.; Sun, Q.; Yang, X.; Gao, Y.; Xu, X.; Zhang, J. Period1 Mediates Rhythmic Metabolism of Toxins by Interacting with CYP2E1. Cell Death Dis. 2021, 12, 76. [Google Scholar] [CrossRef]
- Chen, M.; Chen, M.; Lu, D.; Wang, Y.; Zhang, L.; Wang, Z.; Wu, B. Period 2 Regulates CYP2B10 Expression and Activity in Mouse Liver. Front. Pharmacol. 2021, 12, 764124. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-H.; Lee, J.H.; Han, D.-H.; Cho, S.; Lee, Y.-J. Circadian Clock Is Involved in Regulation of Hepatobiliary Transport Mediated by Multidrug Resistance-Associated Protein 2. J. Pharm. Sci. 2017, 106, 2491–2498. [Google Scholar] [CrossRef]
- Geng, Y.-J.; Smolensky, M.H.; Sum-Ping, O.; Hermida, R.; Castriotta, R.J. Circadian Rhythms of Risk Factors and Management in Atherosclerotic and Hypertensive Vascular Disease: Modern Chronobiological Perspectives of an Ancient Disease. Chronobiol. Int. 2023, 40, 33–62. [Google Scholar] [CrossRef]
- Kosanovic Rajacic, B.; Sagud, M.; Pivac, N.; Begic, D. Illuminating the Way: The Role of Bright Light Therapy in the Treatment of Depression. Expert Rev. Neurother. 2023, 23, 1157–1171. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-H.; Wu, F.; Yu, S.; Guo, Y.-N.; Zhao, R.-R.; Zhang, R.-L. Therapeutic Sleep Deprivation for Major Depressive Disorder: A Randomized Controlled Trial. J. Affect. Disord. 2024, 361, 10–16. [Google Scholar] [CrossRef]
- Civelek, E.; Ozturk Civelek, D.; Akyel, Y.K.; Kaleli Durman, D.; Okyar, A. Circadian Dysfunction in Adipose Tissue: Chronotherapy in Metabolic Diseases. Biology 2023, 12, 1077. [Google Scholar] [CrossRef] [PubMed]
- Türk, D.; Scherer, N.; Selzer, D.; Dings, C.; Hanke, N.; Dallmann, R.; Schwab, M.; Timmins, P.; Nock, V.; Lehr, T. Significant Impact of Time-of-Day Variation on Metformin Pharmacokinetics. Diabetologia 2023, 66, 1024–1034. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.; Tagliamento, M.; Auclin, E.; Aldea, M.; Frelaut, M.; Levy, A.; Benitez, J.C.; Naltet, C.; Lavaud, P.; Botticella, A.; et al. Clinical Outcomes by Infusion Timing of Immune Checkpoint Inhibitors in Patients with Advanced Non-Small Cell Lung Cancer. Eur. J. Cancer 2023, 182, 107–114. [Google Scholar] [CrossRef]
- Catozzi, S.; Assaad, S.; Delrieu, L.; Favier, B.; Dumas, E.; Hamy, A.-S.; Latouche, A.; Crochet, H.; Blay, J.-Y.; Mullaert, J.; et al. Early Morning Immune Checkpoint Blockade and Overall Survival of Patients with Metastatic Cancer: An In-Depth Chronotherapeutic Study. Eur. J. Cancer 2024, 199, 113571. [Google Scholar] [CrossRef]
- Saito, Y.; Yoshida, S.; Nakaya, N.; Hata, Y.; Goto, Y. Comparison between Morning and Evening Doses of Simvastatin in Hyperlipidemic Subjects. A Double-Blind Comparative Study. Arterioscler. Thromb. A J. Vasc. Biol. 1991, 11, 816–826. [Google Scholar] [CrossRef]
- Wallace, A.; Chinn, D.; Rubin, G. Taking Simvastatin in the Morning Compared with in the Evening: Randomised Controlled Trial. BMJ 2003, 327, 788. [Google Scholar] [CrossRef]
- Acimovic, J.; Fink, M.; Pompon, D.; Bjorkhem, I.; Hirayama, J.; Sassone-Corsi, P.; Golicnik, M.; Rozman, D. CREM Modulates the Circadian Expression of CYP51, HMGCR and Cholesterogenesis in the Liver. Biochem. Biophys. Res. Commun. 2008, 376, 206–210. [Google Scholar] [CrossRef]
- Lévi, F.; Zidani, R.; Misset, J.-L. Randomised Multicentre Trial of Chronotherapy with Oxaliplatin, Fluorouracil, and Folinic Acid in Metastatic Colorectal Cancer. Lancet 1997, 350, 681–686. [Google Scholar] [CrossRef]
- Singal, A.G.; Llovet, J.M.; Yarchoan, M.; Mehta, N.; Heimbach, J.K.; Dawson, L.A.; Jou, J.H.; Kulik, L.M.; Agopian, V.G.; Marrero, J.A.; et al. AASLD Practice Guidance on Prevention, Diagnosis, and Treatment of Hepatocellular Carcinoma. Hepatology 2023, 78, 1922–1965. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Ye, Y.; Li, E.; Fan, J.; Gong, J.; Ying, J.; Cao, Y.; Li, R.; Wang, P. A Circadian Clock Gene-Related Signature for Predicting Prognosis and Its Association with Sorafenib Response in Hepatocellular Carcinoma. Transl. Cancer Res. 2023, 12, 2493. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-Q.; Cui, W.; Cai, J.; Sun, Y. Integrative Analysis Indicates the Prognostic Value of Circadian Rhythm Disruption in Liver Cancer: Potential for Therapeutic Targeting. Front. Immunol. 2022, 13, 1011264. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Xiang, Y.; Ozguc, F.M.; Kim, Y.; Liu, C.-J.; Park, P.K.; Hu, Q.; Diao, L.; Lou, Y.; Lin, C.; et al. The Genomic Landscape and Pharmacogenomic Interactions of Clock Genes in Cancer Chronotherapy. Cell Syst. 2018, 6, 314–328.e2. [Google Scholar] [CrossRef]
- Sartorelli, L.S.; Neto, R.J.B.; Moscheta-Pinheiro, M.G.; Castro, T.B.d.; Fernandes, F.L.C.; Silva, R.; Novais, A.A.; Chuffa, L.G.d.A.; Reiter, R.J.; Zuccari, D.A.P.d.C. Blood Melatonin Level Can Serve as a Potential Biomarker for Prostate and Hepatocellular Carcinomas. Melatonin Res. 2021, 4, 253–269. [Google Scholar] [CrossRef]
- Lorente, L.; Rodriguez, S.T.; Sanz, P.; Abreu-González, P.; González-Rivero, A.F.; Pérez-Cejas, A.; Padilla, J.; Díaz, D.; González, A.; Martín, M.M.; et al. Low Serum Melatonin Levels Prior to Liver Transplantation in Patients with Hepatocellular Carcinoma Are Associated with Lower Survival after Liver Transplantation. Int. J. Mol. Sci. 2019, 20, 1696. [Google Scholar] [CrossRef]
- Laothong, U.; Hiraku, Y.; Oikawa, S.; Intuyod, K.; Murata, M.; Pinlaor, S. Melatonin Induces Apoptosis in Cholangiocarcinoma Cell Lines by Activating the Reactive Oxygen Species-Mediated Mitochondrial Pathway. Oncol. Rep. 2015, 33, 1443–1449. [Google Scholar] [CrossRef]
- Laothong, U.; Pinlaor, P.; Boonsiri, P.; Pairojkul, C.; Priprem, A.; Johns, N.P.; Charoensuk, L.; Intuyod, K.; Pinlaor, S. Melatonin Inhibits Cholangiocarcinoma and Reduces Liver Injury in Opisthorchis Viverrini-Infected and N-Nitrosodimethylamine-Treated Hamsters. J. Pineal Res. 2013, 55, 257–266. [Google Scholar] [CrossRef]
- Hassan, S.A.; Ali, A.A.H.; Sohn, D.; Flögel, U.; Jänicke, R.U.; Korf, H.-W.; von Gall, C. Does Timing Matter in Radiotherapy of Hepatocellular Carcinoma? An Experimental Study in Mice. Cancer Med. 2021, 10, 7712–7725. [Google Scholar] [CrossRef]
- Das, M.; Kumar, D.; Sauceda, C.; Oberg, A.; Ellies, L.G.; Zeng, L.; Jih, L.J.; Newton, I.G.; Webster, N.J.G. Time-Restricted Feeding Attenuates Metabolic Dysfunction-Associated Steatohepatitis and Hepatocellular Carcinoma in Obese Male Mice. Cancers 2024, 16, 1513. [Google Scholar] [CrossRef]
- Davidson, A.J.; Straume, M.; Block, G.D.; Menaker, M. Daily Timed Meals Dissociate Circadian Rhythms in Hepatoma and Healthy Host Liver. Int. J. Cancer 2006, 118, 1623–1627. [Google Scholar] [CrossRef]
- Molina-Aguilar, C.; Guerrero-Carrillo, M.d.J.; Espinosa-Aguirre, J.J.; Olguin-Reyes, S.; Castro-Belio, T.; Vázquez-Martínez, O.; Rivera-Zavala, J.B.; Díaz-Muñoz, M. Time-Caloric Restriction Inhibits the Neoplastic Transformation of Cirrhotic Liver in Rats Treated with Diethylnitrosamine. Carcinogenesis 2017, 38, 847–858. [Google Scholar] [CrossRef]
- Pascale, A.; Allard, M.A.; Benamar, A.; Levi, F.; Adam, R.; Rosmorduc, O. Prognostic Value of the Timing of Immune Checkpoint Inhibitors Infusion in Patients with Advanced Hepatocellular Carcinoma. J. Clin. Oncol. 2024, 42, 457. [Google Scholar] [CrossRef]
- Joerg, V.; Scheiner, B.; D’Alessio, A.; Fulgenzi, C.A.M.; Schönlein, M.; Kocheise, L.; Lohse, A.W.; Huber, S.; Wege, H.; Kaseb, A.; et al. Efficacy and Safety of Atezolizumab/Bevacizumab in Patients with HCC after Prior Systemic Therapy: A Global, Observational Study. Hepatol. Commun. 2023, 7, e0302. [Google Scholar] [CrossRef]
- Burgermeister, E.; Battaglin, F.; Eladly, F.; Wu, W.; Herweck, F.; Schulte, N.; Betge, J.; Härtel, N.; Kather, J.N.; Weis, C.-A.; et al. Aryl Hydrocarbon Receptor Nuclear Translocator-like (ARNTL/BMAL1) Is Associated with Bevacizumab Resistance in Colorectal Cancer via Regulation of Vascular Endothelial Growth Factor A. eBioMedicine 2019, 45, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Innominato, P.F.; Giacchetti, S.; Moreau, T.; Smaaland, R.; Focan, C.; Bjarnason, G.A.; Garufi, C.; Iacobelli, S.; Tampellini, M.; Tumolo, S.; et al. Prediction of Survival by Neutropenia According To Delivery Schedule of Oxaliplatin–5-Fluorouracil–Leucovorin for Metastatic Colorectal Cancer in a Randomized International Trial (EORTC 05963). Chronobiol. Int. 2011, 28, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Innominato, P.F.; Ballesta, A.; Huang, Q.; Focan, C.; Chollet, P.; Karaboué, A.; Giacchetti, S.; Bouchahda, M.; Adam, R.; Garufi, C.; et al. Sex-Dependent Least Toxic Timing of Irinotecan Combined with Chronomodulated Chemotherapy for Metastatic Colorectal Cancer: Randomized Multicenter EORTC 05011 Trial. Cancer Med. 2020, 9, 4148–4159. [Google Scholar] [CrossRef]
- Hesse, J.; Müller, T.; Relógio, A. An Integrative Mathematical Model for Timing Treatment Toxicity and Zeitgeber Impact in Colorectal Cancer Cells. npj Syst. Biol. Appl. 2023, 9, 27. [Google Scholar] [CrossRef]
- Hesse, J.; Martinelli, J.; Aboumanify, O.; Ballesta, A.; Relógio, A. A Mathematical Model of the Circadian Clock and Drug Pharmacology to Optimize Irinotecan Administration Timing in Colorectal Cancer. Comput. Struct. Biotechnol. J. 2021, 19, 5170–5183. [Google Scholar] [CrossRef]
- Luo, M.; Wang, Q.; Sun, Y.; Jiang, Y.; Wang, Q.; Gu, Y.; Hu, Z.; Chen, Q.; Xu, J.; Chen, S.; et al. Fasting-Mimicking Diet Remodels Gut Microbiota and Suppresses Colorectal Cancer Progression. npj Biofilms Microbiomes 2024, 10, 53. [Google Scholar] [CrossRef]
- Weng, M.; Chen, W.; Chen, X.; Lu, H.; Sun, Z.; Yu, Q.; Sun, P.; Xu, Y.; Zhu, M.; Jiang, N.; et al. Fasting Inhibits Aerobic Glycolysis and Proliferation in Colorectal Cancer via the Fdft1-Mediated AKT/MTOR/HIF1α Pathway Suppression. Nat. Commun. 2020, 11, 1869. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.Y.; Eussen, S.J.P.M.; van Roekel, E.H.; Pot, G.K.; Koster, A.; Breukink, S.O.; Janssen-Heijnen, M.L.G.; Keulen, E.T.P.; Stehouwer, C.D.A.; Weijenberg, M.P.; et al. Longitudinal Associations of Circadian Eating Patterns with Sleep Quality, Fatigue and Inflammation in Colorectal Cancer Survivors up to 24 Months Post-Treatment. Br. J. Nutr. 2024, 131, 1166–1180. [Google Scholar] [CrossRef] [PubMed]
- Lima Oliveira, M.; Biggers, A.; Oddo, V.M.; Naylor, K.B.; Chen, Z.; Hamm, A.; Pezley, L.; Peñalver Bernabé, B.; Gabel, K.; Sharp, L.K.; et al. Design of a Remote Time-Restricted Eating and Mindfulness Intervention to Reduce Risk Factors Associated with Early-Onset Colorectal Cancer Development among Young Adults. Nutrients 2024, 16, 504. [Google Scholar] [CrossRef]
- Ravi, A.; Joseph, B. Circadian Mechanisms in Medicine. N. Engl. J. Med. 2021, 384, 550–561. [Google Scholar] [CrossRef]
- Tao, L.; Jiang, R.; Zhang, K.; Qian, Z.; Chen, P.; Lv, Y.; Yao, Y. Light Therapy in Non-Seasonal Depression: An Update Meta-Analysis. Psychiatry Res. 2020, 291, 113247. [Google Scholar] [CrossRef]
- Pjrek, E.; Friedrich, M.-E.; Cambioli, L.; Dold, M.; Jäger, F.; Komorowski, A.; Lanzenberger, R.; Kasper, S.; Winkler, D. The Efficacy of Light Therapy in the Treatment of Seasonal Affective Disorder: A Meta-Analysis of Randomized Controlled Trials. Psychother. Psychosom. 2019, 89, 17–24. [Google Scholar] [CrossRef]
- Cremascoli, R.; Sparasci, D.; Giusti, G.; Cattaldo, S.; Prina, E.; Roveta, F.; Bruno, F.; Ghezzi, C.; Cerri, S.; Picascia, M.; et al. Effects of Circadian Phase Tailored Light Therapy on Sleep, Mood, and Cognition in Alzheimer’s Disease: Preliminary Findings in a Pivotal Study. Front. Physiol. 2022, 12, 755322. [Google Scholar] [CrossRef]
- Yoon, J.; Heo, S.; Lee, H.; Sul, E.; Han, T.; Kwon, Y.-J. Feasibility and Efficacy of Morning Light Therapy for Adults with Insomnia: A Pilot, Randomized, Open-Label, Two-Arm Study. Medicina 2023, 59, 1066. [Google Scholar] [CrossRef]
- Lai, F.; Luo, Z.; Zhang, J.; Xia, W.; Tian, L. Bright Light Therapy Has a Positive Effect on Sleep Quality in Patients with Cancer: A Meta-Analysis. Sleep Med. Rev. 2024, 75, 101925. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, V.; Chennupati, S.; Sanchez, H.; Sun, Q.; Aly, A.; Marcus, H.; Seal, B.S. Healthcare Utilization and Costs in Hepatocellular Carcinoma (HCC) Patients Treated at a Large Referral Center in Washington (WA) State. J. Clin. Oncol. 2021, 39, e16149. [Google Scholar] [CrossRef]
- Aly, A.; Ronnebaum, S.; Patel, D.; Doleh, Y.; Benavente, F. Epidemiologic, Humanistic and Economic Burden of Hepatocellular Carcinoma in the USA: A Systematic Literature Review. Hepatic Oncol. 2020, 7, HEP27. [Google Scholar] [CrossRef] [PubMed]
- Bhimani, N.; Wong, G.Y.M.; Molloy, C.; Pavlakis, N.; Diakos, C.I.; Clarke, S.J.; Dieng, M.; Hugh, T.J. Cost of Treating Metastatic Colorectal Cancer: A Systematic Review. Public Health 2022, 211, 97–104. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrell, J.M. Chronobiology of Cancers in the Liver and Gut. Cancers 2024, 16, 2925. https://doi.org/10.3390/cancers16172925
Ferrell JM. Chronobiology of Cancers in the Liver and Gut. Cancers. 2024; 16(17):2925. https://doi.org/10.3390/cancers16172925
Chicago/Turabian StyleFerrell, Jessica M. 2024. "Chronobiology of Cancers in the Liver and Gut" Cancers 16, no. 17: 2925. https://doi.org/10.3390/cancers16172925
APA StyleFerrell, J. M. (2024). Chronobiology of Cancers in the Liver and Gut. Cancers, 16(17), 2925. https://doi.org/10.3390/cancers16172925