Simple Summary
Cancer is a disease caused by mutations in DNA differentiating cancer cells from normal cells. Precision medicine could exploit these genetic differences to identify and create therapeutic opportunities that selectively kill cancer cells while sparing normal cells. In some cancers, the same mutations recur, making them desirable biomarkers for genetics-based therapeutic strategies. This is the case for the gene ARID1A which is lost in about 50% of ovarian clear cell carcinomas. We used genetic screening to identify targets for killing an ovarian clear cell carcinoma cell line lacking the ARID1A gene. We identified a cancer stress response regulator called KEAP1 as a potential target for ARID1A-deficient ovarian clear cell carcinomas. Using different approaches and cell lines, we observed a relationship where ARID1A-deficient cells required KEAP1 for fitness. Together these data suggest that KEAP1 could be pursued as a means to selectively kill certain types of ARID1A-deficient cancer.
Abstract
ARID1A is the core DNA-binding subunit of the BAF chromatin remodeling complex and is mutated in about 8% of all cancers. The frequency of ARID1A loss varies between cancer subtypes, with clear cell ovarian carcinoma (CCOC) presenting the highest incidence at > 50% of cases. Despite a growing understanding of the consequences of ARID1A loss in cancer, there remains limited targeted therapeutic options for ARID1A-deficient cancers. Using a genome-wide CRISPR screening approach, we identify KEAP1 as a genetic dependency of ARID1A in CCOC. Depletion or chemical perturbation of KEAP1 results in selective growth inhibition of ARID1A-KO cell lines and edited primary endometrial epithelial cells. While we confirm that KEAP1-NRF2 signalling is dysregulated in ARID1A-KO cells, we suggest that this synthetic lethality is not due to aberrant NRF2 signalling. Rather, we find that KEAP1 perturbation exacerbates genome instability phenotypes associated with ARID1A deficiency. Together, our findings identify a potentially novel synthetic lethal interaction of ARID1A-deficient cells.
1. Introduction
AT-rich interaction domain 1A (ARID1A) is the DNA-binding subunit of the BAF complex, the canonical SWI/SNF chromatin-remodeling complex, which regulates a variety of processes within the cell, including chromatin accessibility, gene expression, and the maintenance of genome integrity. ARID1A is mutated in ~8% of all cancers, with clear cell ovarian carcinoma (CCOC) presenting the highest incidence of ARID1A loss at > 50% of cases [1,2]. ARID1A mutations typically occur throughout the length of the gene and result in loss of protein expression. While ARID1A loss has shown prognostic value for several neoplastic malignancies (e.g., gastric [3,4,5], lung [6], hepatocellular [7,8,9], and breast cancer [10]), it remains ambiguous how ARID1A status influences the prognosis of gynecologic malignancies. Previous studies have reported adverse clinical outcomes for patients harboring ARID1A mutations [10,11], though conflicting reports have emerged suggesting no clinical association (e.g., stage, survival, and histopathological features) between ARID1A-positive and negative cohorts [12,13,14,15,16,17]. Nevertheless, loss of ARID1A is widely recognized as an enabling factor for cancer development and progression.
ARID1A deficiency results in a broad range of phenotypes, including genome instability [18,19,20], transcriptional dysregulation [21,22], and metabolic dysfunction [23,24,25]. Despite an increasing body of evidence documenting context-specific consequences of ARID1A loss in cancer, there remains a critical gap in knowledge as to how to selectively treat patients harboring these lesions. Genetic dependencies of ARID1A have been used to suggest novel therapeutic avenues against ARID1A-deficient cancer cells. For example, taking advantage of the antagonistic relationship between BAF and the polycomb repressive 2 (PRC2) complex on the regulation of gene expression [26,27,28], the EZH2 inhibitor tazemetostat is currently in phase II clinical trials for the treatment of ARID1A-deficient ovarian cancers [29,30,31] (EPZ-IST-001). Other promising therapies include BET inhibitors (e.g., JQ1) [32], agents targeting DNA, and replication stress responses (ATR/PARP inhibitors) [18]. The diverse targets of these inhibitors reflect the diversity of cellular activities disrupted by ARID1A loss. Nevertheless, treatment options for CCOC patients harboring ARID1A mutations remain poorly specific, with surgery and chemotherapy (platinum + taxane) being the standard of care [33,34].
In this study, we employ a genome-wide CRISPR screen to identify genes important for fitness in an isogenic CCOC cell line model of ARID1A loss. We identify the oxidative stress regulator KEAP1 as a potentially novel synthetic lethal partner of ARID1A. Using clinical and cell line samples, we document that KEAP1 perturbation with CRISPR, small molecule inhibitors, and RNAi selectively impair the growth of ARID1A-deficient cells, and results in enhanced genome instability phenotypes. Building on these findings, we show that the combination treatment of ATR inhibitors and small molecules targeting KEAP1 impairs the growth of ARID1A-KO cells, suggesting that the genome instability induced in ARID1A-KO following KEAP1 perturbation can be harnessed therapeutically. We also observe a general dysregulation of KEAP1-NRF2 signalling in ARID1A-KO cells, suggesting that perhaps other functions of KEAP1 underlie the genetic dependency between KEAP1 and ARID1A. Altogether, our findings uncover a novel vulnerability of ARID1A-deficient CCOC cells.
2. Results
2.1. Genome-Wide CRISPR Screen Identifies Genes Important for Fitness in ARID1A-KO CCOC Cells
To identify genes important for fitness when ARID1A is lost in the context of CCOC, we performed genome-wide CRISPR screens in biological duplicates in RMG-1 cells and its ARID1A knockout (ARID1A-KO) derivative using the Toronto Knockout version 3 library (TKOv3) [35] (Figure 1A). RMG-1 ARID1A-KO cells were generated using CRISPR as documented previously [19] (Supplementary Figure S1A). Cells were infected at > 500× coverage of the library and were cultured until 14 days post-puromycin selection to capture robust fitness defects induced by sgRNA knockouts. Genomic DNA was purified and sgRNA representation was assessed by deep amplicon sequencing (> 99% sgRNA representation across all samples). Reads were aligned to the TKOv3 library using Bowtie2 [36] and sgRNA enrichment was calculated using the BAGEL2 pipeline [37], which integrates the fitness scores from all four sgRNAs into a single value for each target gene. Bayes Factor (BF) scores obtained from the BAGEL2 analysis were compared between the ARID1A-WT and ARID1A-KO datasets to identify genes important for fitness, specifically within the ARID1A-KO populations. As expected, non-targeting control sgRNAs targeting eGFP, LacZ, and Luciferase presented the lowest BF scores (Supplementary Table S1) across all samples. Furthermore, our approach recovered > 94% (ARID1A-WT) and > 87.5% (ARID1A-KO) of the training dataset of core essential genes (CEGs) as important for fitness (Supplementary Table S1.1-6 highlights replicate read counts (RC), fold changes (FC) and precision recall (PR)). As further validation to our experimental approach, precision-recall (PR) curves were plotted to assess screen performance, and the high precision-to-recall ratio suggested a low false positive (precision) and high false negative rates (recall) (Figure 1B).
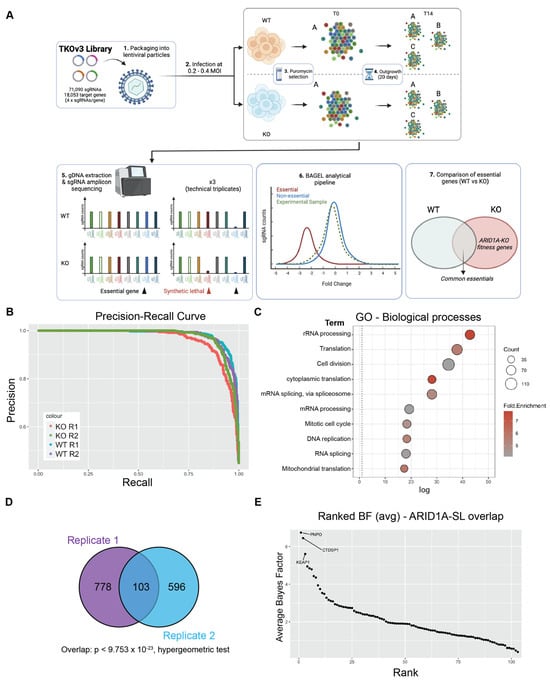
Figure 1.
Genome-wide CRISPR screen identifies KEAP1 as a synthetic lethal partner of ARID1A. (A) Schematic of CRISPR screening workflow with the TKOv3 library. (B) Precision-Recall curve assessing CRISPR screen performance of individual replicates (values obtained from BAGEL2 algorithm, Supplementary Table S1.5-6). (C) Top 10 enriched Biological Processes GO terms identified using David v 6.8 (p < 0.05, with FDR correction) for the ARID1A-KO fitness genes identified by either of our CRISPR screens (n = 1484 genes, Supplementary Table S1.7). (D) Overlap of ARID1A-SL hits from both biological replicates of the CRISPR screen. (E) Ranked ARID1A-KO specific hits (average BF from both biological replicates, Supplementary Table S1.8). The top 3 hits are labeled on the graph. Panel (A) was created using Biorender.
In total, each biological replicate of our screen identified a consensus of 1484 genes important for fitness of ARID1A-KO cells (Supplementary Table S1.7). To investigate the potential functional repertoire of all the identified hits, we performed a GO analysis on this dataset, which revealed an enrichment in pathways relating to gene expression, cell cycle, and mitochondrial function, pathways known to impact fitness in cancer cells [38,39,40,41] (Figure 1C and Supplementary Table S2.1). To focus on potential genetic dependencies unique to ARID1A-KO cells, we filtered out fitness genes that were also identified in ARID1A-WT cells. This resulted in 881 and 699 ARID1A-KO-specific hits from each replicate, with a significant overlapping consensus of 103 high-confidence synthetic lethal partners, dubbed “ARID1A-SL” (p < 9.753 × 10−23, hypergeometric test, Figure 1D and Supplementary Tables S1.8 and S2.2). Importantly, the overlap of 103 genes successfully identifies known ARID1A synthetic lethal partners in DDX19A [42], SMARCC1 [43], and FAAP24 [42] amongst others (Supplementary Table S1.8). Importantly, these genes are also identified within the top 100 dependencies in the Cancer Dependency Map project database. Additional known synthetic lethal partners were also recovered in either of the screen replicates (e.g., SMARCB1/E1 [42,43], ARID1B [44], and BRD2 [32]; Supplementary Table S1.8).
To prioritize and pursue strong novel candidate synthetic lethal partners of ARID1A for further validation, we averaged the BF score of the ARID1A-SL dataset and ranked the top hits (Figure 1E). This approach identified PNPO, CTDSP1, and KEAP1 as the top essential genes when ARID1A is lost. PNPO encodes pyridoxine 5′-phosphate oxidase, which converts dietary vitamin B6 into its biologically active form, contributing to a variety of metabolic processes. CTDSP1 encodes a small phosphatase that targets serine 5 in the heptameric repeats of the C-terminal domain of the large RNA polymerase II subunit POLR2A, controlling gene expression dynamics for most PolII genes. PNPO and CTDSP1 are interesting hits, but likely have pleiotropic effects on global cellularly phenomena. KEAP1 on the other hand is a ubiquitin E3 ligase whose major target, NRF2, a transcription factor with roles in the regulation of metabolism, inflammation, mitochondrial function, and more [45]. NRF2′s activity is tightly regulated by KEAP1 under normal conditions, whereby KEAP1 sequesters NRF2 and targets it for proteasomal degradation. Given these data, the fact that ARID1A functionally interacts with NRF2, and that the disruption of this interaction results in fitness defects [24], we chose to focus on validating KEAP1 as a synthetic lethal partner of ARID1A in CCOC. We further assessed the validity of KEAP1 as a hit by looking at the normalized read counts for all 4 sgRNAs and confirmed the dropout of these guides in the ARID1A-KO samples (Supplementary Figure S1D).
2.2. Validation of KEAP1 as a Fitness Gene in ARID1A-Deficient Cells
To validate the potential synthetic lethal relationship between ARID1A and KEAP1, we independently transfected our RMG-1 ARID1A isogenic cell line pair with two independent siRNAs targeting KEAP1 (KEAP1_5 and KEAP1_8), and performed a proliferation assay with crystal violet staining to assess the sensitivity of ARID1A-KO cells to KEAP1 depletion. ARID1A-KO cells showed decreased proliferation following KEAP1 knockdown when compared to their ARID1A-WT parental cells, as measured by the decrease in crystal violet staining (Figure 2A). As expected, knockdown of KEAP1 resulted in decreased KEAP1 protein expression that was accompanied by an increase in the levels of its major target protein, NRF2 (Supplementary Figure S2A). Since siKEAP1_5 appeared to yield more consistent knockdowns, we decided to focus on this siRNA moving forward.
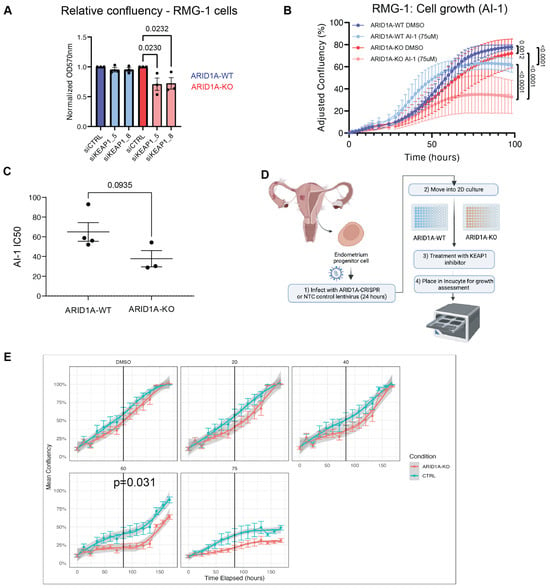
Figure 2.
ARID1A-KO cells are sensitive to KEAP1 depletion by siRNA and small molecule perturbation. (A) Quantification of crystal violet viability assays in RMG-1 cells (mean ± SEM, one-way ANOVA, n = 3, statistically significant p-values are displayed on graph). (B) ARID1A-KO RMG-1 cells grow slower than ARID1A-WT when treated with 75 µM AI-1 as measured by IncuCyte S3 imaging system. Relative confluency presented as the growth from initial time point. Error bars represent SEM of averaged triplicated wells from 3 independent experiments. P values obtained from extra sum-of-squares F test on calculated logistic growth rate are indicated on graph. (C) ARID1A-KO CCOC cells present lower IC50 values to AI-1 than ARID1-WT cells (t-test, mean ± SEM). IC50 values were obtained from dose-response curves presented in Supplementary Figure S2F–H (n = 3). (D) Patient-derived endometrium progenitor cell workflow. Normal endometrial tissue was processed and put in 2D culture to expand the progenitor cell population before CRISPR and AI-1 treatments (see Section 4). (E) Average proliferation curves of three primary endometrial epithelial cell cultures transduced with non-targeting (NTC5) or sgARID1A (ARID1A-KO) lentivirus and treated with AI-1 at the indicated concentrations. Technical quadruplicates from each patient were normalized and combined. p-value was determined at the midpoint for 60 µM AI-1 (Welch’s t-test). CRISPR knockdown western blots, and individual patient derived growth curves are shown in Supplementary Figure S2E–H. Panel (D) was created using Biorender.
In a parallel approach, we treated ARID1A-deficient or proficient RMG-1 cells with a small molecule targeting KEAP1, called AI-1. AI-1 covalently modifies cysteine residues on KEAP1′s C-terminus to release NRF2 from KEAP1 and disrupt KEAP1′s interaction with the CUL3-RBX1 complex [46,47]. RMG-1 ARID1A-KO cells presented growth defects when subjected to AI-1 treatment when compared to their ARID1A-WT counterparts (Figure 2B and Supplementary Figure S2B). As expected, treatment with AI-1 resulted in an induction of NRF2 protein levels (Supplementary Figure S3F). To further test the potential generalizability of these findings, we assessed the growth of a panel of CCOC cell lines grouped by ARID1A status when subjected to AI-1 treatment and observed a moderate trend where ARID1A-mutated cells presented lower IC50 values compared to the WT counterparts (Figure 2C and Supplementary Figure S2C,D).
Additionally, to further assess the ARID1A-KEAP1 synthetic sick/lethal relationship, we obtained primary endometrial epithelial cells and generated ARID1A-KO primary cells using lentiviral CRISPR constructs (Figure 2D). Briefly, normal endometrial tissue was processed and put in 2D culture to expand the epithelial cell population. These cells were then transduced with lentivirus containing a CRISPR sgRNA against ARID1A or a non-targeting control (NTC), and ARID1A-expression was validated by western blot (Supplementary Figure S2E). Cell growth of ARID1A-KO and NTC-transduced cells challenged with increasing concentrations of AI-1 confirmed our observations from cell lines, where ARID1A-KO primary cells from three independent donors presented growth defects compared to their NTC counterparts (Figure 2E and Supplementary Figure S2F–H). Altogether these results confirm the observations from our CRISPR screen and suggest that KEAP1 is important for cellular fitness in ARID1A-deficient ovarian/endometrial derived cells.
2.3. Gene Expression Analysis Reveals a Dysregulation of NRF2 Target Genes in ARID1A-KO Cells
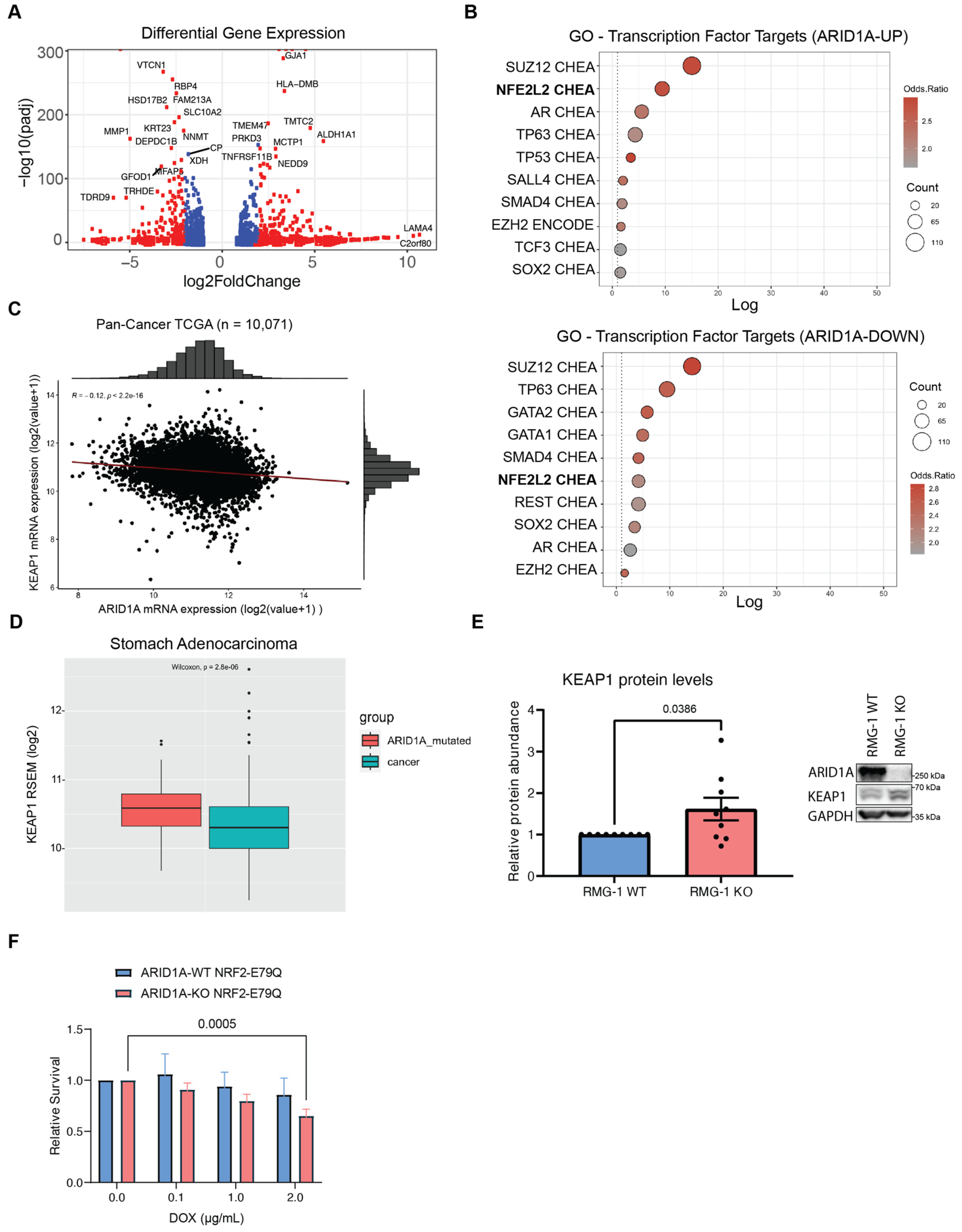
To gain further insight on the mechanisms underlying the cellular fitness relationship between KEAP1 and ARID1A, we first assessed if and how NRF2 signalling may be disrupted in ARID1A-KO cells. To do so, we revisited RNA-seq data from Wu et al. [22] in RMG-1 cells that either had intact or mutated ARID1A status. Although this isogenic line was generated using different sgRNAs, these cells possess the same genetic background as those used for our CRISPR screens. Differential gene expression (DGE) analysis using DESEQ2 [48] revealed that ARID1A loss caused a significant up-regulation of 531 genes (ARID1A-UP), and down-regulation of 500 genes (ARID1A-DOWN) (Figure 3A and Supplementary Table S3). GO analysis using the ENCODE and ChEA datasets on the differentially expressed genes revealed a significant enrichment of NRF2 (also known as NFE2L2) target genes (Figure 3B). These results highlight how ARID1A loss results in significant changes in NRF2-mediated gene expression, and perhaps support the notion that KEAP1 signalling is altered in these cells.
Since KEAP1 and ARID1A are commonly mutated in cancer [1,2,49], we compared mRNA expression of KEAP1 and ARID1A in a large cohort of solid tumors of various origins from The Cancer Genome Atlas (TCGA) datasets (Figure 3C and Supplementary Figure S3A). This analysis revealed that transcript levels from KEAP1 inversely correlated with ARID1A, albeit mildly, across a pan-cancer panel (n = 10,071, r = −0.12, p = 2.2 × 10−16). Conversely, a weak positive correlation (n = 10,071, R = 0.095, p < 2.2 × 10−16) was observed between the expression of NRF2 and ARID1A (Supplementary Figure S3B). This observation suggests that ARID1A and KEAP1 could compensate for the loss of each other in cancer development. Furthermore, stratification of patients by ARID1A mutational status confirmed this relationship, where some ARID1A-mutated cancer subtypes (e.g., stomach adenocarcinoma—STAD) appear to upregulate KEAP1 compared to wild-type (Figure 3D and Supplementary Figure S3A). To address this hypothesis, we quantified KEAP1 protein levels from western blots and found a small but significant increase in KEAP1 protein levels in RMG-1 ARID1A-KO cells compared to wild-type (Figure 3E). Overall, our data suggest that some ARID1A-deficient cells maintain elevated levels of KEAP1, perhaps as a mechanism to sustain the growth of these cancer cells.
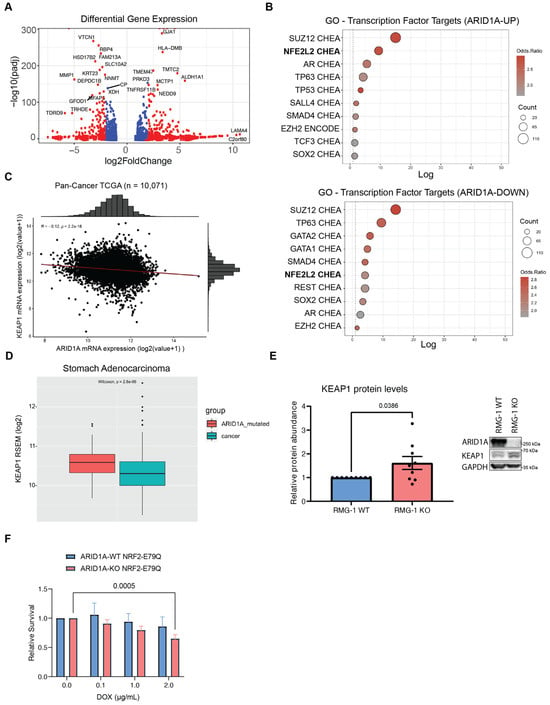
Figure 3.
ARID1A-KO sensitivity to KEAP1 depletion is only partly due to dysregulation of NRF2 (A) Volcano plot of differentially expressed genes in ARID1A-KO RMG-1 cells from RNA-seq dataset published in Wu et al. [22] (blue dots if padj < 0.01, red dots if log2FC > 1 or log2FC < −1 AND padj < 0.01). (B) Top 10 enriched transcription factor target genes identified from the ARID1A-UP (above) and ARID1A-DOWN (below) regulated genes using the ENCODE [50] and ChEA [51] datasets (EnrichR [52]; p < 0.05 with Benjamini-Hochberg correction). (C) Quantification of KEAP1 and ARID1A mRNA levels (RSEM, log2) from pan-cancer TCGA data (n = 10,071) showing a negative correlation between the two genes. (D) Quantification of KEAP1 mRNA levels (RSEM, log2) from TCGA data (stomach adenocarcinoma—STAD) showing higher KEAP1 levels in ARID1A-mutated patient samples (n = 90) compared to WT (n = 336) (box limits indicate 25% and 75% over median, Wilcoxon test, p = 2.8 × 10−6). (E) Left: Quantification of KEAP1 protein levels from western blot data showing higher levels in ARID1A-KO RMG-1 cells. KEAP1 intensity was normalized to respective GAPDH loading control, and then to ARID1A-WT (data from 9 from independent samples, mean ± SEM, t test, significant p-values displayed on graph). Right: Representative western blot image. (F) Quantification of crystal violet viability assay showing that ARID1A-KO RMG-1 cells exhibit sensitivity to NRF2E79Q expression induced by doxycycline (mean ± SEM, ANOVA, n = 3, significant p-values displayed on graph).
Figure 3.
ARID1A-KO sensitivity to KEAP1 depletion is only partly due to dysregulation of NRF2 (A) Volcano plot of differentially expressed genes in ARID1A-KO RMG-1 cells from RNA-seq dataset published in Wu et al. [22] (blue dots if padj < 0.01, red dots if log2FC > 1 or log2FC < −1 AND padj < 0.01). (B) Top 10 enriched transcription factor target genes identified from the ARID1A-UP (above) and ARID1A-DOWN (below) regulated genes using the ENCODE [50] and ChEA [51] datasets (EnrichR [52]; p < 0.05 with Benjamini-Hochberg correction). (C) Quantification of KEAP1 and ARID1A mRNA levels (RSEM, log2) from pan-cancer TCGA data (n = 10,071) showing a negative correlation between the two genes. (D) Quantification of KEAP1 mRNA levels (RSEM, log2) from TCGA data (stomach adenocarcinoma—STAD) showing higher KEAP1 levels in ARID1A-mutated patient samples (n = 90) compared to WT (n = 336) (box limits indicate 25% and 75% over median, Wilcoxon test, p = 2.8 × 10−6). (E) Left: Quantification of KEAP1 protein levels from western blot data showing higher levels in ARID1A-KO RMG-1 cells. KEAP1 intensity was normalized to respective GAPDH loading control, and then to ARID1A-WT (data from 9 from independent samples, mean ± SEM, t test, significant p-values displayed on graph). Right: Representative western blot image. (F) Quantification of crystal violet viability assay showing that ARID1A-KO RMG-1 cells exhibit sensitivity to NRF2E79Q expression induced by doxycycline (mean ± SEM, ANOVA, n = 3, significant p-values displayed on graph).
2.4. Aberrant NRF2 Signalling Does Not Significantly Impair the Growth of ARID1A-KO Cells
To explore the possibility that aberrant NRF2 signalling may underlie the growth defects of ARID1A-KO cells subjected to KEAP1 perturbation, we generated mutant NRF2-inducible cell lines by transducing our isogenic RMG-1 model (+/− ARID1A) with lentiviral particles containing a pIND20-NRF2E79Q-HA construct. The E79Q mutation in NRF2 confers reduced binding affinity for KEAP1, resulting in NRF2 pathway activation [53]. Upon induction of NRF2E79Q protein expression in our ARID1A-WT and KO cells using doxycycline, we observed a minor reduction in the growth of ARID1A-KO cells but not WT cells (Figure 3F and Supplementary Figure S3C,D). This phenotype was restricted to the highest dosage of doxycycline (2 µg/mL), and we observed no sensitivity to doxycycline at the doses tested (Supplementary Figure S3E). Our data therefore suggest that perhaps the dysregulation of the KEAP1 function beyond NRF2 sequestration, and not NRF2 activation only, may be responsible for the sensitivity displayed by ARID1A-KO cells.
2.5. KEAP1 Perturbation Exacerbates Genome Instability in ARID1A-Deficient Cells
Since the canonical KEAP1 substrate NRF2 does not appear to be responsible for the growth defects observed in ARID1A-KO cells, we sought other explanations. Our group has previously reported that ARID1A-deficient cells experience increased rates of DNA replication stress and DNA damage [19]. To assess whether the sensitivity of ARID1A-KO cells to KEAP1 perturbation could be associated with genome instability phenotypes, we performed immunofluorescence assays to measure p-RPA(ser33) foci formation. p-RPA(ser33) foci formation is a bona fide marker of DNA replication stress, and indicates RPA filaments modified by ATR. ARID1A-KO cells presented a higher accumulation of p-RPA(ser33) foci compared to WT cells (Figure 4A). Importantly, we observed a significant increase in p-RPA(ser33) foci in ARID1A-KO cells treated with sublethal doses of AI-1 (50 µM), but not in ARID1A-WT cells (Figure 4A). These results suggest that KEAP1 perturbation may enhance the genome instability phenotypes associated with ARID1A-loss. To assess if the replication stress associated with KEAP1 perturbation also resulted in DNA damage, we performed an immunofluorescence assay probing for γH2AX foci formation. In agreement with our previous results, we observed a significant increase in γH2AX foci formation in our RMG-1 ARID1A-KO cells, and 50 µM AI-1 treatment caused a significant increase in γH2AX foci formation specifically in the ARID1A-KO cells, but not the WT (Figure 4B).
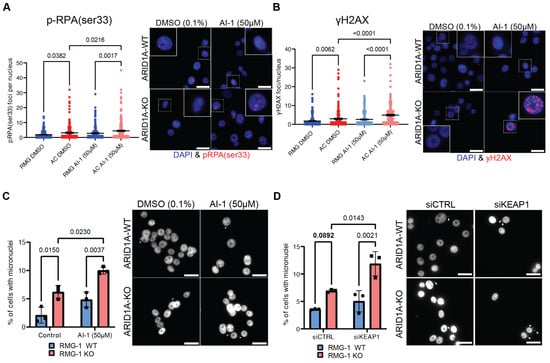
Figure 4.
KEAP1 perturbation exacerbates genome instability phenotypes of ARID1A-KO cells (A,B). Quantification (left) and representative images (right) of immunofluorescence experiment showing AI-1 (50 µM) induces higher rates of p-RPAser33 (A) or γH2AX (B) foci formation in ARID1A-KO RMG-1 cells (mean ± SEM, ANOVA, n = 3, significant p-values displayed on graph). (C,D) Quantification (left) and representative images (right) showing that ARID1A-KO RMG-1 cells accumulate higher levels of micronuclei compared to WT when treated with 50 µM AI-1 (C) or siKEAP1 (D) (mean ± SEM, ANOVA, n = 3, significant p-values displayed on graph). White arrows highlighting micronuclei. In A through D, a 20 µm scale bar is show in the lower right.
If DNA damage and replication stress impacts the fitness of ARID1A-KO cells, evidence of induced genome instability should be evident upon KEAP1 depletion. To test this, we assessed micronuclei formation by DAPI staining following KEAP1 perturbation. Micronuclei are typically formed from lagging chromosomes during mitosis or appear in senescent cells due to a defective nuclear envelope [54]. We found that under normal conditions, ARID1A-KO cells present a higher frequency of micronuclei [20] (Figure 4C). AI-1 treatment induced an increase in micronuclei formation specifically in ARID1A-KO cells, while no change could be observed in the ARID1A-WT cells in our isogenic cell line (Figure 4C). We observed the same trend in ARID1A-KO RMG-1 cells subjected to siKEAP1, (Figure 4D). In addition, western blot analysis probing for γH2AX in a representative primary endometrial progenitor sample treated with NTC- or sgARID1A- and challenged with AI-1 revealed an increase in γH2AX in ARID1A-KO (Supplementary Figure S4A). Thus, we conclude that ARID1A-deficient cells exist in a sensitized state where KEAP1 perturbation with AI-1 or siRNA enhances genome instability phenotypes.
2.6. Dual Inhibition of KEAP1 and ATR Potentiates Killing of ARID1A-KO Cells
Since ARID1A-loss sensitizes cells to ATR inhibition [55], and we observed enhanced marks of replication stress when treating cells with KEAP1 inhibitors, we set out to test whether AI-1 treatment could potentiate the selective killing of these cells. To do so, we treated our isogenic RMG-1 cells with sub-lethal concentrations of ceralasertib, a potent ATR inhibitor, alone or in combination with AI-1 (Supplementary Figure S4B). At the 60 h time point, neither AI-1 nor ceralasertib treatment resulted in significant killing of RMG-1 ARID1A-KO cells, while the combination treatment of AI-1 + ceralasertib resulted in the potentiation of the killing of ARID1A mutants (Supplementary Figure S4B). These data agree with the genome instability phenotypes induced by KEAP1 perturbation in ARID1A-KO cells and suggests that combination treatment with ATRi could be harnessed to improve the selective growth inhibition of ARID1A-KO cells by AI-1.
3. Discussion
Despite a high incidence of mutations across cancers, precision medicine options for patients harboring ARID1A mutations in cancer remain limited and unspecific. To identify genetic dependencies of ARID1A-deficient CCOC cells and to ultimately advise on new synthetic lethal target development, we performed two genome-wide CRISPR screens in an isogenic cell line model of ARID1A loss in RMG-1 cells. Our screen identified known ARID1A synthetic lethal partners in SMARCC1 and DDX19A [42,43]. Known ARID1A synthetic lethal paralogue ARID1B was only identified as significant in one of the two biological replicates, highlighting the noise of this approach and the need for replicates. The overlap between our two screens identified a consensus dataset of 103 genes that are important for fitness when ARID1A is lost (Figure 1D). While this overlap was statistically significant, our data also highlight the variability between biological replicates of CRISPR screening experiments.
The top hit from our screen, PNPO is a metabolic enzyme involved in the breakdown of vitamin B6. Despite no reports of ARID1A loss impacting vitamin B6 metabolism, there are many reports highlighting the metabolic consequences of ARID1A mutations [23,24,25]. It is therefore possible that loss of ARID1A, through metabolic reprogramming, would induce sensitivity to PNPO depletion. Further research will be needed to confirm this hypothesis. The 2nd strongest hit from our screen was assigned to CTDSP1, which functions in the regulation of transcriptional elongation by RNAPII. While we did not test effects of CTDSP1 depletion beyond the screen, there is strong evidence that loss of ARID1A results in defects in transcriptional elongation [21]. In fact, ARID1A deficiency impairs promoter proximal pausing dynamics, a crucial step for productive transcriptional elongation [21]. We hypothesize that inhibition of CTDSP1 would exacerbate this phenotype and perhaps sensitize ARID1A-KO cells to CTDSP1 depletion. While these genes are interesting and warrant further investigation, we decided to focus on the 3rd strongest fitness genes identified by our screen, KEAP1, given that ARID1A physically interacts with its downstream target NRF2, and that the disruption of this interaction was linked to fitness defects [24].
To validate the findings from our CRISPR screen, we assessed the sensitivity of KEAP1 perturbation by siRNA or by small molecule in an isogenic cell line model of ARID1A loss. AI-1 was initially characterized as an NRF2 activator, acting by covalently modifying cysteine residues of KEAP1, to promote the stabilization and transcriptional activation of NRF2 [46]. AI-1 also disrupts the ability of KEAP1 to serve as an adaptor for CUL3-KEAP1 ubiquitin ligase complex, potentially impacting other functions of KEAP1 like the regulation of protein homeostasis [46]. KEAP1 perturbation by AI-1 resulted in growth defects in a cell line model when ARID1A is lost, and also in primary endometrial progenitors depleted for ARID1A. This was further corroborated in a panel of CCOC cell lines, where we saw a moderate correlation between lower AI-1 IC50 values and ARID1A-deficiency. Overall, our findings suggest that KEAP1 perturbation impairs the growth of ARID1A-deficient cells.
To gain further insight on the potential mechanism underlying the sensitivity of ARID1A-KO cells to KEAP1 perturbation, we investigated whether NRF2 signalling, downstream KEAP1, could explain the growth defects we observed. Building on work by Wu et al. [22], we performed a differential gene expression analysis that identified large subsets of genes that are disrupted when ARID1A is lost. Among these, polycomb repressive complex 2 (PRC2) target genes were highly enriched within the DGE dataset, as seen by the enrichment of the PRC2 subunits SUZ12 and EZH2 (Figure 3B). This is unsurprising considering that PRC2 and the BAF complex have antagonistic roles in the regulation of gene expression [26,56]. Accordingly, EZH2 inhibition in ARID1A-deficient cells has been reported to behave in a synthetic lethal manner [57,58]. Interestingly, our DGE analysis also revealed that NRF2 (NFE2L2) target genes are highly enriched among both the up-regulated, and to a lesser extent, down-regulated genes (Figure 3B). Furthermore, the analysis of pan-cancer TCGA data revealed a mild but significant correlation where some ARID1A-mutated tumor subtypes present elevated KEAP1 mRNA levels (e.g., stomach adenocarcinoma—STAD, and colon adenocarcinoma—COADREAD) (Figure 3D and Supplementary Figure S3A). Unfortunately, limitations in sample size and data availability prevented us from assessing this trend in ovarian carcinoma subtypes specifically. These results suggest that KEAP1 is perhaps up-regulated when ARID1A is mutated, and this may support cancer cell growth.
Although we cannot rule out that transcriptional rewiring by NRF2 activation may be partially responsible for the growth defects of ARID1A-KO cells following KEAP1 perturbation, our findings suggest that functions of KEAP1 beyond NRF2 sequestration may underlie the genetic dependency we report here. To this extent, we demonstrated that KEAP1 perturbation enhances genome instability phenotypes in ARID1A-KO cells, though the nature of the damage sustained by these cells remains unknown. Marzio et al. [59] have shown that loss of KEAP1 can produce a BRCA-ness phenotype, which may explain our observations of elevated genome instability phenotypes in ARID1A-KO cells subjected to KEAP1 perturbation. Moreover, KEAP1 has been shown to interact with proteins like BRCA1 and PALB2, which may influence DNA repair pathway choice and cell cycle regulation. It is possible that defects in DNA repair pathway choice may also result in inadequate DNA repair, though more research is necessary to confirm this hypothesis. Furthermore, two groups have identified MCM3, a component of the replicative DNA helicase complex, as a substrate of KEAP1 [60,61]. While the exact function of this interaction remains unknown, Mulvaney et al. showed that KEAP1 associates with chromatin during S-phase [61]. The authors suggest that KEAP1 plays a role in monitoring replication fork progression and the coordination of replisome activity. Overall, these results point towards an NRF2-independent role for KEAP1 in the maintenance of genome integrity, which may be further exacerbated when ARID1A is lost. Finally, since KEAP1 is an essential component of the KEAP1-CUL3-RBX1 ubiquitin ligase complex that targets various proteins for proteasomal degradation, and also interacts with p62 to regulate autophagic flux [62,63,64,65,66], it also remains possible that KEAP1 perturbation could disrupt normal protein homeostasis, indirectly impairing the growth of ARID1A-KO cells.
While our group is the first to our knowledge to document a genetic dependency between KEAP1 and ARID1A, consequences of ARID1A loss on NRF2 signalling have been reported. Ogiwara et al. [24] have reported that ARID1A loss causes glutathione metabolism deficiencies, which sensitizes ARID1A-KO cells to SLC7A11 depletion. The authors showed that this synthetic lethal relationship was partially explained by the loss of an interaction between ARID1A and NRF2, which in turn regulates SLC7A11 expression. While the role of KEAP1 in this genetic dependency has not been explored, additional evidence suggests that KEAP1-mediated signalling is important in ARID1A-deficient cells. In fact, bromodomain and extra-terminal domain inhibitors (BETi) have been used to selectively kill ARID1A-KO cells; particularly the inhibitor JQ-1, which targets BRD4 [32]. While the authors suggest that this toxicity is due to transcriptional changes associated with an increased BRD4 recruitment to chromatin, BRD4 inhibition by JQ-1 has been shown to affect KEAP1-NRF2 signalling, supporting the idea that KEAP1 is perhaps a regulator of fitness in ARID1A-deficient cells [67,68,69]. Additionally, loss of other BAF complex subunits has been associated with the dysregulation of KEAP1-NRF2 signalling. For instance, mutations in the catalytic subunit BRG1 have been associated with changes in NRF2-mediated gene expression [70]. Overall, this suggests that there exists a convergence in functions between the BAF complex and KEAP1-NRF2 signalling, though further research is needed to explore the nature of this connection. Overall, we propose a model (Figure 5) where KEAP1 perturbation results in an upregulation of NRF2 signalling and perhaps dysregulation of other cellular processes such as protein homeostasis, which is viable when ARID1A is intact; however, when KEAP1 is perturbed in ARID1A-deficient cells, the resulting genome instability is exacerbated by KEAP1 perturbation and results in lethality.
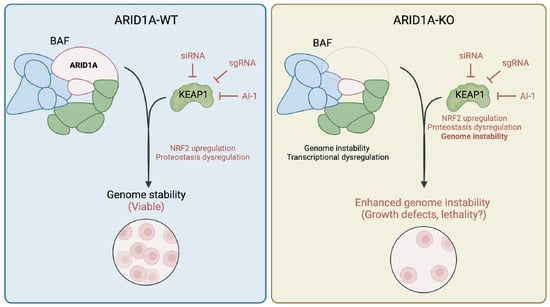
Figure 5.
Sensitivity of ARID1A-KO cells to KEAP1 perturbation is conserved in endometrium progenitor-derived organoid model. Model of ARID1A-KEAP1 synthetic lethality. Perturbation of KEAP1 dysregulates normal NRF2 signalling and proteostasis, which remains viable in normal cells. Perturbation of KEAP1 in ARID1A-KO cells specifically enhances genome instability phenotypes, resulting in growth defects. This figure was created using Biorender.
We recognize that relatively high concentrations (micromolar range) of AI-1 are required to elicit the phenotypes observed, suggesting that this inhibitor is not promising for clinical use. In fact, the detailed consequences of AI-1 inhibition remain poorly characterized. While some off-target effects have been predicted against HDAC1 [46], our group did not observe significant changes in protein expression when subjecting cells to lethal doses of this small molecule, though these observations do not exclude the possibility that HDAC1 function may be affected by this inhibitor (Supplementary Figure S3F). Nevertheless, our observations suggest that further investigation of KEAP1 function in ARID1A-deficient cells is warranted to further inform on the fitness defects described herein. In fact, our observations that sublethal concentrations of AI-1 in combination with ceralasertib allowed to selectively kill ARID1A-KO cells suggests that combination therapy may potentiate the effects of AI-1, allowing perhaps limit off-target effects. Importantly, the validation of our findings using siRNA knockdown of KEAP1 as a complementary approach supports our conclusions. Ultimately, both generating more potent KEAP1 inhibitors and understanding the mechanisms underlying the growth defects of ARID1A-KO cells is needed to confirm the clinical potential of these findings.
4. Methods
4.1. Cell Culture Conditions
RMG-1 ARID1A-WT and ARID1A-KO cells were a generous gift from the Huntsman Lab. The CCOC cell line panel (JHOC-5/7, OVISE, OVMANA, OVCA 429, and RMG-II) were also obtained from the Huntsman Lab. All CCOC cell lines were cultured in RPMI 1640 supplemented with 10% FBS and 1% penicillin/streptomycin at 37 °C, supplied with 5% CO2.
4.2. Generation of NRF2E79Q Dox-Inducible Cell Lines
The pIND20-NRF2E79Q-HA plasmid was a generous gift from Bernard Weissman. The plasmid was transformed into bacteria for amplification and validated by sequencing (Supplementary Table S4.2). To produce lentiviral particles for stable expression of the construct, 8 × 106 HEK-293T cells were transfected in a 15-cm culture dish. Cells were transfected using the TransIT-LT1 Transfection reagent (Mirus, Madison WI, MIR 2305, 3 µL/µg of DNA) with 4.8 μg psPAX2, 3.8 μg pMD2.G and 8 µg pIND20-NRF2E79Q-HA DNA 24-h after cells were seeded. After 24 h of transfection, the media were replaced with high-BSA growth media (DMEM + 1.1 g/100 mL BSA + 1x Pen/Strep) for viral harvest. Lentiviral particles were harvested 48 h post-transfection, and viral particles were concentrated by ultracentrifugation (25,000× g, 4 °C, 1.5 h), resuspended in OPTI-MEM, and frozen at −80 °C for future use. To generate the stable cell lines, 1 × 106 RMG-1 cells were infected at a MOI between 0.2–0.4 in the presence of 8 µg/mL polybrene. The infected cells were selected for 96 h in media containing 800 µg/mL geneticin (Thermo, Burnaby, BC, Canada, CAT: 10131027).
4.3. TKOv3 Library Cloning and Viral Preparation
The Toronto human knockout pooled library (TKOv3) was a gift from Jason Moffat (Addgene, Watertown, MA, #90294). The library was amplified in bacteria as described in the Moffat protocol on Addgene (https://www.addgene.org/pooled-library/moffat-crispr-knockout-tkov3/) (accessed on 1 November 2019). The amplified sgRNA library was packaged into lentiviral particles using HEK293T/17 cells by co-transfection with psPAX2 and pMD2.G viral plasmids (1:1:1 molar ratio). To produce viral particles at scale, 510 × 106 cells were transfected in 60 15 cm culture dishes. Each dish was transfected using the TransIT-LT1 Transfection reagent (Mirus, MIR 2305, 3 µL/µg of DNA) with 4.8 μg psPAX2, 3.8 μg pMD2.G, and 8 μg library DNA 24-h after cells were seeded. After 24 h of transfection, the media was replaced with high-BSA growth media (DMEM + 1.1 g/100 mL BSA + 1x Pen/Strep) for viral harvest. Lentiviral particles were harvested twice at 48 h and 72 h post-transfection. Both harvests were pooled, viral particles were concentrated by ultracentrifugation (25,000× g, 4 °C, 1.5 h) and frozen at −80 °C for future use.
4.4. TKOv3 Library Viral Transduction and Titration
Viral titre was assessed to determine the multiplicity of infection (MOI) of the concentrated viral particles. RMG-1 derived, ARID1A-WT, and ARID1A-KO cells were seeded at a density of 2.5 × 106 cells/well in 12-well plates and spin-fected (2000 rpm for 2 h at 37 °C) with increasing concentrations of virus in the presence of 8 µg/mL polybrene (Millipore, Temecula, CA, USA, TR-003-G lot#3287963). The next day, the cells were split into 6-well plates at a density of 0.5 × 106 cells/well and subjected to 2 µg/mL puromycin (Sigma, Burlington, MA, USA, P8833) for selection of infected cells. Cells not transduced with the library (no virus control) did not survive past 24 h of puromycin selection (2 µg/mL). After 48 h of selection, all cells in all wells were counted to determine the viral volume that resulted in 20–40% survival in puromycin (corresponding to an MOI of 0.2–0.4 assuming an independent infection rate).
4.5. CRISPR Knockout Pool Generation
The dropout CRISPR screen was performed in biological duplicates, in an adapted version of the Mair et al. protocol [71]. ARID1A-WT and ARID1A-KO RMG-1 cells were transduced with sgRNA libraries at a multiplicity of infection of (MOI) between 0.2 and 0.4, aiming for coverage of, on average, >500 cells per sgRNA reagent. For genome-wide screens, 120 million cells were transduced by “spin-fection” (2 h, 2000 rpm, 37 °C) per cell line in 12-well plates (2.5 × 106 cells per well) using the appropriate volume of viral particles for MOI of 0.2–0.4 and 8 µg/mL polybrene. Two additional wells per cell line were also plated to account for no puromycin and no virus controls to assess MOI at time 0 (T0). Media was aspirated and refreshed following the infection and cells were incubated overnight. The following day, the cells were split, pooled, and seeded at 8 × 106/plate in 150 mm dishes in media containing 2 µg/mL puromycin. Additional plates were seeded to assess MOI (no-puromycin, no-virus, and virus-infected). Forty-eight hours post-selection with puromycin, the cells were split, and MOI was assessed. Time 0 samples were frozen to assess initial library representation (20 × 106 cells—>250x), and the remainder of cells were maintained in culture and split as needed to ensure confluence did not exceed 90%. Cells were harvested and frozen as “dry” pellets (−80 °C) at relevant time points for subsequent sgRNA enrichment assessment (described below).
4.6. Genomic DNA Extraction and Precipitation
Genomic DNA extraction and precipitation was performed according to the method described in the TKOv3 protocol described by Mair et al. [71] using the QIAamp Blood Maxi kit (cat no. 51194) and RNase A (cat no. 19101), both obtained from Qiagen, Germantown, MD, USA.
4.7. Library Preparation and Sequencing
The sgRNA libraries were prepared using two-steps of PCR as described by Mair et al. [71]: (1) amplify the sgRNA region within genomic DNA; (2) amplify sgRNAs with Illumina (San Diego, CA, USA) TruSeq adapters with i5 and i7 indices (see Supplementary Table S4). These indices are unique sequences that are added to DNA samples during library preparation and act as sample identifiers during multiplex sequencing. All PCR steps were performed using the NEBNext Ultra II Q5 Master Mix high-fidelity polymerase, from NEB, Whitby, ON, Canada. For PCR #1, the thermocycling parameters were 98 °C for 30 s, 25 cycles of (98 °C for 10 s, 66 °C for 30 s, 72 °C for 15 s), and 72 °C for 2 min. In each PCR #1 reaction, we used 3.5 μg of gDNA. For each sample, the necessary number of PCR #1 reactions was used to capture the appropriate representation of the screen. For instance, assuming a diploid genome is ~7.2 pg and one guide-RNA per genome, 100 µg of genomic DNA yields ~200-fold coverage of the TKOv3 library. In this example, at least 29 PCR #1 reactions were necessary to capture 200x representation of the library for each replicate.
PCR #1 products were pooled for each biological sample and 5 µL was used for amplification and unique barcoding in PCR #2. The thermocycling parameters for PCR #2 were 98 °C for 30 s, followed by 10 cycles of (98 °C for 10 s, 55 °C for 30 s, and 65 °C for 15 s), and 65 °C for 5 min. The PCR #2 products were validated using gel electrophoresis and purified using the QIAquick Gel extraction kit (QIAGEN, Germantown, MD, USA). The purified libraries were then quantified by Nanodrop and Qubit prior to sequencing on an Illumina MiSeq (2 samples per lane for >100x coverage).
4.8. Amplicon Scoring, Guide-RNA Enrichment and Hit Identification
Deep amplicon sequencing data were processed for sgRNA representation using custom scripts. To summarize, the sequencing reads were first de-multiplexed using the 8 bp barcodes in the forward primer, then using the 8 bp barcodes in the reverse primers. De-multiplexed reads were trimmed to leave only the 20 bp spacer (sgRNA) sequences. The sgRNA sequences were then mapped to the reference TKOv3 library sgRNA sequences using Bowtie 2 [48]. For mapping, no mismatches were allowed, and mapped sgRNA sequences were quantified by counting the total number of reads.
Fitness scores were assigned to each gene by integrating the read counts from individual sgRNAs using BAGEL2 (each replicate analysed independently) [37]. Genes with a positive Bayes Factor and an FDR cut-off of 0.05 were determined important for fitness across datasets, and hits that were unique to the ARID1A-KO datasets (i.e., not overlapping between ARID1A-WT and KO) were identified as synthetic lethal to ARID1A. The consensus dataset of synthetic lethal partners of ARID1A corresponds to the intersection of ARID1A-KO-specific hits from both biological replicates of the screen.
4.9. Gene Ontology (GO) Analysis
Gene ontology analysis for biological processes was performed using DAVID v6.8 [72]. Gene ontology analysis for target genes of transcription factors was performed using the ENCODE and ChEA datasets from EnrichR [52].
4.10. Incucyte Growth Assay
Cells were seeded at 10,000 (RMG-1) cells per well in a clear bottom 96-well plate on day 0. For drug treatments, cells were treated with indicated drug concentrations on day 1 and placed in an IncuCyte S3 live-cell imaging system (Sartorius, Goettingen, Germany) contained in an incubator kept at 37 °C and 5% CO2. Images were taken at 2 h intervals in quadruplicate/well for 72–96 h.
4.11. RNA Interference
Cells of 2 × 105 were reverse-transfected in 6-well plates with Flexitube siRNA constructs (Qiagen) targeting KEAP1 (KEAP1_5 or KEAP1_8), NRF2 (NRF2_10), or a non-targeting control (siCTRL) at concentrations of 25 nM with HiPerFect Transfection Reagent (Qiagen, Germantown, MD, USA, #301705) according to the manufacturer’s protocol. Target sequences are available in Supplementary Table S4. Cells were cultured for 48 h after transfection and before subsequent analysis. Alternatively, for reverse transfection, cells were seeded at 10,000 (RMG-1) cells per well in 24-well plates on day 0. On day 1, cells were transfected with 25 nM siRNA (final concentration).
4.12. Crystal Violet Proliferation Assay
KEAP1 siRNA proliferation: For siRNA experiments, cells were seeded at 10,000 (RMG-1) cells per well in 24-well plates on day 0. On day 1, cells were transfected with 25 nM siRNA (final concentration). Cells were fixed for crystal violet staining after 7 days of outgrowth on day 8.
For experiments with drug treatments, cells were seeded at 5000 (OVCA 429) or 10,000 (RMG-1, RMG-II, OVMANA, OVISE, JHOC-5/7) cells per well in 24-well plates, and incubated overnight prior to addition of the drug. Cells were fixed for crystal violet staining after outgrowth of 7 days on day 8.
For experiments with RMG-1 p.IND20-NRF2-E79-HA expressing cells, 10,000 cells were seeded in a 6-well plate on day 0. On day 1, doxycycline was added to the media at concentrations indicated in Figure legends. Media was changed on day 4 to replenish doxycycline. Cells were fixed for crystal violet staining on day 7.
Briefly, cells were washed with PBS and fixed with 3.7% formaldehyde for 15 min at RT. Cells were washed with PBS and stained with crystal violet for 30 min at RT, after which cells were washed with distilled H2O until no residual crystal violet could be observed. The plates were dried at RT overnight. The following day, the plates were incubated for 15 min at RT (in dark) following the addition of 500 µL of 10% acetic acid in methanol. The absorbance at 570 nm was measured using a Tecan Infinite M200Pro plate reader (Tecan, Mannedorf, Switzerland).
4.13. Immunofluorescence
For all immunofluorescence experiments, cells were grown on coverslips overnight. For experiments with drug treatments, cells were treated at indicated concentrations for 24 h prior to fixation. For siRNA transfections, cells were reverse transfected at time of seeding with appropriate siRNA concentrations and grown for 48 h prior to fixation.
Cells were fixed with 4% paraformaldehyde for 10 min and permeabilized with 0.2% Triton X-100 for 10 min on ice. After permeabilization, cells were washed with PBS and blocked in 3%BSA, 0.1% Tween 20 in 4x saline sodium citrate buffer (SSC) for 1 h at room temperature. Cells were then incubated with primary antibodies (Supplementary Table S4) overnight at 4 °C. Following PBS wash, cells were then incubated with Alexa-Fluor-488 or 568-conjugated secondary antibodies for 1 h at room temperature, washed with PBS for 10 min 3 times, and stained with DAPI before mounting and imaging on LeicaDMI8 microscope (Leica, Wetzlar, Germany) at 100x magnification. ImageJ was used for image processing and quantification [73]. For assessment of micronuclei formation, DAPI staining was used to identify micronuclei-positive cells, which were quantified as a percentage of all cells analyzed.
4.14. Western Blotting
Whole-cell lysates were prepared with RIPA buffer containing protease inhibitor (Sigma) and phosphatase inhibitor (Roche Applied Science, Mississauga, ON, Canada) cocktail tablets and the protein concentration was determined by Bio-Rad Protein assay (Bio-Rad, Redmond, WA, USA). Equivalent amounts of protein were resolved by SDS-PAGE and transferred to polyvinylidene fluoride microporous membrane (Millipore), blocked with 1.5% BSA in H2O containing 0.1% Tween-20 (TBS-T), and membranes were probed with the primary antibodies listed in (Supplementary Table S4). Secondary antibodies were conjugated to horseradish peroxidase (HRP) and peroxidase activity was visualized using Chemiluminescent HRP substrate (Thermo Scientific, Burnaby, BC, Canada). For quantification of protein samples, western blot images were analyzed using Image Lab Software for PC (v6.1). KEAP1 protein levels were normalized to GAPDH loading control and compared between samples.
4.15. TCGA Dataset Analysis
mRNA expression levels were obtained from the TCGA cBioportal [50] (RSEM—Batch normalized from Illumina HiSeq_RNASeqV2). Correlations of ARID1A and KEAP1 were analyzed using all samples (n = 10,071). For stratification by ARID1A-mutational status, ARID1A-mutated samples were sub-setted using VEG scores of “High” (meaning high impact on protein function—e.g., frameshift or loss). Cancer subtypes with at least 5 data points for either of ARID1A-WT or ARID1A-KO were selected for analysis.
4.16. Differential Gene Expression Analysis
RNA-seq data for RMG-1 cells isogenic for ARID1A were obtained from Wu et al. [22]. Differentially expressed genes were generated using DESEQ2 [48].
4.17. Culture of Patient-Derived Endometrial Progenitor Cells
Primary endometrial epithelial cells and organoids were derived as previously reported by Cochrane et al. [74] from hysterectomy tissue from surgeries for non-cancer reasons performed at the Vancouver General Hospital (VGH) and University of British Columbia (UBC) hospitals. Tissues were collected as part of the OVCARE’s Gynaecologic Tissue Bank. These studies were approved by the Institutional Review Board (IRB) of UBC and British Columbia Cancer Agency (H05-60119), and use of the tissue for research purposes was approved by written informed consent by the patients.
Cells were seeded in six well plates to be transfected by ARID1A and non-targeting control CRISPR lentiviruses. ARID1A (12354, 1:1000, Cell Signaling, Danvers, MA, USA) western blot was performed to confirm the downregulation of ARID1A protein in sgARID1A-infected cells compared to NTC controls. Then, cells were seeded at a density of 1000 cells/well in 96 well plates. KEAP1 inhibitor treatment was performed after 24 h using increasing concentrations and the plates were placed into an IncuCyte S3 live-cell imaging system contained in an incubator kept at 37 °C and 5% CO2 to assess proliferation for 7 days.
4.18. Statistical Analysis, Data Availability, and Reproducibility
Statistical analysis was performed using GraphPad Prism 9. Experiments were repeated three times unless otherwise stated. The representative images were shown unless otherwise stated. Quantitative data were expressed as means ± SEM unless otherwise stated. Analysis of variance (ANOVA) was used to identify significant differences in multiple comparisons. Strains and plasmids are available upon request. The authors affirm that all data necessary for confirming the conclusions of the article are present within the article, figures, and tables.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cancers16172949/s1, Supplementary Figure S1. (A). Representative western blot images of isogenic RMG-1 ARID1A-WT and KO cells. (B, C) Overlap of ARID1A-WT (B) or ARID1A-KO (C) fitness genes with the core essential gene training dataset (CEG2) for each replicate of our TKOv3 CRISPR screen. (D) Normalized read count data obtained using BAGEL2 [75] for all four TKOv3 sgRNAs targeting KEAP1 (n = 2). Supplementary Figure S2. (A) Representative western blots showing KEAP1 siRNA knockdown efficiency in RMG-1 cells. (B) Severely impaired growth of RMG-1 ARID1A-WT and KO cells treated with 100 µM AI-1 as measured by IncuCyte S3 imaging system. Relative confluency presented as the growth from initial time point. Error bars represent SEM from triplicated wells from three independent experiments. p values obtained from extra sum-of-squares F test on calculated logistic growth rate are indicated on graph. (C) Representative western blot image of ARID1A, KEAP1, and NRF2 levels of the CCOC cell line panel. (D) Dose-response regression curves obtained from crystal violet proliferation assays for CCOC cell line panels. Confluency after 7 days of growth at indicated AI-1 concentrations was measured using crystal violet (averaged technical triplicate data from 3-4 independent experiments—or 2 independent experiments for 25 µM, mean ± SEM). IC50 values compared in Figure 2C. (E) Upper left: representative western blot images confirming ARID1A status in NTC or sgARID1A-transduced primary endometrium progenitor cells. Right and below: shows raw proliferation curves for individual patient-derived cell cultures transduced with non-targeting (NTC5) or sgARID1A (ARID1A-KO) lentivirus and treated with AI-1 at indicated concentrations as measured by IncuCyte S3 imaging system (data presented as mean of quadruplicate wells from cells derived from a single patient). Note that despite different primary cell responses, the red ARID1A-KO curves are always more sensitive to AI-1. Supplementary Figure S3. (A) Quantification of KEAP1 mRNA levels (RSEM, log2) from pan-cancer TCGA data stratified by cancer subtype and ARID1A-status (significant p-values are displayed on graph). (B) Quantification of NRF2 and ARID1A mRNA levels (RSEM, log2) from pan-cancer TCGA data (n = 10,071) showing a mild positive correlation between the two genes. (C) Representative western blot image showing dose-dependent induction of NRF2E79Q expression using doxycycline. (D) Representative images of crystal violet staining measuring proliferation of cells following 7 days of dox induction. (E) Quantification of RMG-1 ARID1A-WT and KO proliferation (normalized to DMSO control) showing no sensitivity to increasing concentrations of doxycycline in untransformed cells (mean ± SEM, n = 2 or 3—see Table S5). (F) Representative western blot image showing NRF2 induction by AI-1 treatment but no effect on HDAC1 protein levels of RMG-1 cells treated with increasing concentrations of AI-1 for 24 h. Supplementary Figure S4. (A) Representative western blot image showing increased γH2AX staining in sgARID1A-CRISPR cells after 48 h treatment with AI-1 (data from cells derived from a single patient). (B) Quantification of confluency (relative to DMSO control) of RMG-1 cells treated with AI-1 (75 µM)/ceralasertib (25 nM) alone or in combination as measured after 60 h of treatment by IncuCyte S3 imaging system (technical duplicate wells from three independent experiments; mean ± SEM, ANOVA, significant p-values displayed on graph). Supplementary Figure S5. Raw western blot images. Supplementary Table S1. CRISPR screen datasets. Sheet 1: Normalized read counts from the analysis of biological replicate 1 of our TKOv3 CRISPR screen. Sheet 2: Normalized read counts from the analysis of biological replicate 2 of our TKOv3 CRISPR screen. Sheet 3: Fold change of sgRNAs from the analysis of biological replicate 1 of our TKOv3 CRISPR screen. Sheet 4: Fold change of sgRNAs from the analysis of biological replicate 2 of our TKOv3 CRISPR screen. Sheet 5: BAGEL output of Bayes Factor, Precision, Recall and FDR values of fitness genes in ARID1A-WT and KO cells (replicate 1). Sheet 6: BAGEL output of Bayes Factor, Precision, Recall and FDR values of fitness genes in ARID1A-WT and KO cells (replicate 2). Sheet 7: Summary of all significantly enriched fitness genes identified in ARID1A-KO cells. Sheet 8: ARID1A-KO-specific hits for each biological replicate of the TKOv3 screen (i.e., synthetic lethal partners of ARID1A), along with overlap ARID1A-SL consensus from the two replicates. Supplementary Table S2. GO analysis (David v6.8). Sheet 1: GO analysis of enriched biological processes for the ARID1A fitness genes identified in both CRISPR screen replicates (overlap presented in Supplemental Table S1.7). Sheet 2: GO analysis of enriched biological processes for the consensus ARID1A-SL dataset (103 genes, Supplemental Table S1.8). Supplementary Table S3. Differential gene expression analysis of up-regulated genes (Sheet 1) and down-regulated genes (Sheet 2) in RMG-1 ARID1A-KO cells from Wu et al. [22] using DESEQ2 [47]. Supplementary Table S4. Miscellaneous. Sheet 1: Reagent List. Sheet 2: p.IND20-NRF2-E79Q-HA plasmid sequence. Supplementary Table S5. Raw Data for each Figure presented in separate sheets.
Author Contributions
Conceptualization, L.-A.F., M.H. and P.C.S.; methodology, L.-A.F., M.H., D.G.H. and P.C.S.; formal analysis, L.-A.F., F.K., J.P.W., J.S.L., G.T.-G., M.M.M., T.S., J.W., A.S., S.L.W., E.S. and Y.W.; investigation, L.-A.F., F.K. and P.C.S.; resources, P.C.S. and D.G.H.; data curation, L.-A.F., J.S.L., T.S., S.L.W. and E.S.; writing—original draft preparation, L.-A.F. and P.C.S.; visualization, L.-A.F. and S.L.W.; supervision, P.C.S., D.G.H. and M.H.; funding acquisition, P.C.S., D.G.H. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
LAF holds a CIHR Frederick Banting and Charles Best doctoral scholarship and a UBC Four Year Fellowship. This work was supported by a Terry Fox Research Institute Program Project grant to P.C.S., M.H. and D.G.H.
Institutional Review Board Statement
These studies were approved by the Institutional Review Board (IRB) of UBC and British Columbia Cancer Agency (H05-60119).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All research data are available within this article and associated supplemental data.
Acknowledgments
We thank DGH and Michael Anglesio for the RMG-1 ARID1A-WT and ARID1A-KO cells, as well as the other CCOC cell lines. We also thank Rodrigo Vallejos and Dawn Cochrane for their support with data analysis and organoid culture, respectively. We also acknowledge Grace Cheng, Fraser Johnson, Dylan Farnsworth, and the Morin Lab for their assistance with the IncuCyte. The pIND20-NRF2E79Q-HA construct was a generous gift from Bernard Weissman. Some of the results shown herein are in whole or part based upon data generated by the TCGA Research Network: https://www.cancer.gov/tcga. We also thank Arun Kumar for the insightful discussions that helped shape this manuscript. LAF would also like to acknowledge René Fournier for providing continuous support through this work.
Conflicts of Interest
LAF is a member of the Office of the Canada Chief Science Advisor Youth Council (CSA-YC). PCS is CEO of Arrowsmith Genetics Inc.
References
- Wiegand, K.C.; Shah, S.P.; Al-Agha, O.M.; Zhao, Y.; Tse, K.; Zeng, T.; Senz, J.; McConechy, M.K.; Anglesio, M.S.; Kalloger, S.E.; et al. ARID1A Mutations in Endometriosis-Associated Ovarian Carcinomas. N. Engl. J. Med. 2010, 363, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Wang, T.-L.; Shih, I.-M.; Mao, T.-L.; Nakayama, K.; Roden, R.; Glas, R.; Slamon, D.; Diaz, L.A.; Vogelstein, B.; et al. Frequent Mutations of Chromatin Remodeling Gene ARID1A in Ovarian Clear Cell Carcinoma. Science 2010, 330, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Ashizawa, M.; Saito, M.; Min, A.K.T.; Ujiie, D.; Saito, K.; Sato, T.; Kikuchi, T.; Okayama, H.; Fujita, S.; Endo, H.; et al. Prognostic Role of ARID1A Negative Expression in Gastric Cancer. Sci. Rep. 2019, 9, 6769. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Y.; Pan, K.; Wang, W.; Chen, S.; Chen, J.; Zhao, J.; Lv, L.; Pan, Q.; Li, Y.; et al. Decreased Expression of the ARID1A Gene Is Associated with Poor Prognosis in Primary Gastric Cancer. PLoS ONE 2012, 7, e40364. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Sheng, L.L.; Wu, J.; Yang, M.; Cheng, X.F.; Wu, N.N.; Ye, X.B.; Cai, J.; Wang, L.; Shen, Q.; et al. Loss of ARID1A Expression Is Associated with Poor Prognosis in Patients with Gastric Cancer. Hum. Pathol. 2018, 78, 28–35. [Google Scholar] [CrossRef]
- Wang, T.; Guo, J.; Liu, W.; Guo, Q.; Cheng, L.; Zheng, R.; Hu, X. Downregulation of ARID1A Is Correlated with Poor Prognosis in Non-Small Cell Lung Cancer. Transl. Cancer Res. 2020, 9, 4896–4905. [Google Scholar] [CrossRef]
- Yang, H.; Huo, J.; Li, X. Identification and Validation of a Five-Gene Prognostic Signature for Hepatocellular Carcinoma. World J. Surg. Oncol. 2021, 19, 90. [Google Scholar] [CrossRef]
- Yim, S.Y.; Kang, S.H.; Shin, J.-H.; Jeong, Y.S.; Sohn, B.H.; Um, S.H.; Lee, J.-S. Low ARID1A Expression Is Associated with Poor Prognosis in Hepatocellular Carcinoma. Cells 2020, 9, 2002. [Google Scholar] [CrossRef]
- He, F.; Li, J.; Xu, J.; Zhang, S.; Xu, Y.; Zhao, W.; Yin, Z.; Wang, X. Decreased Expression of ARID1A Associates with Poor Prognosis and Promotes Metastases of Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 47. [Google Scholar] [CrossRef]
- Cho, H.D.; Lee, J.E.; Jung, H.Y.; Oh, M.-H.; Lee, J.-H.; Jang, S.-H.; Kim, K.-J.; Han, S.W.; Kim, S.Y.; Kim, H.J.; et al. Loss of Tumor Suppressor ARID1A Protein Expression Correlates with Poor Prognosis in Patients with Primary Breast Cancer. J. Breast Cancer 2015, 18, 339–346. [Google Scholar] [CrossRef]
- Ayhan, A.; Mao, T.-L.; Seckin, T.; Wu, C.-H.; Guan, B.; Ogawa, H.; Futagami, M.; Mizukami, H.; Yokoyama, Y.; Kurman, R.J.; et al. Loss of ARID1A Expression Is an Early Molecular Event in Tumor Progression from Ovarian Endometriotic Cyst to Clear Cell and Endometrioid Carcinoma. Int. J. Gynecol. Cancer 2012, 22, 1310–1315. [Google Scholar] [CrossRef] [PubMed]
- Lowery, W.J.; Schildkraut, J.M.; Akushevich, L.; Bentley, R.; Marks, J.R.; Huntsman, D.; Berchuck, A. Loss of ARID1A-Associated Protein Expression Is a Frequent Event in Clear Cell and Endometrioid Ovarian Cancers. Int. J. Gynecol. Cancer 2012, 22, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Maeda, D.; Mao, T.-L.; Fukayama, M.; Nakagawa, S.; Yano, T.; Taketani, Y.; Shih, I.-M. Clinicopathological Significance of Loss of ARID1A Immunoreactivity in Ovarian Clear Cell Carcinoma. Int. J. Mol. Sci. 2010, 11, 5120–5128. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Tsuda, H.; Takano, M.; Tamai, S.; Matsubara, O. PIK3CA Mutations and Loss of ARID1A Protein Expression Are Early Events in the Development of Cystic Ovarian Clear Cell Adenocarcinoma. Virchows Arch. Int. J. Pathol. 2012, 460, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Han, H.H.; Kim, Y.T.; Lee, J.H.; Kim, B.G.; Kang, S.; Cho, N.H. Ovarian Clear Cell Carcinoma Sub-Typing by ARID1A Expression. Yonsei Med. J. 2017, 58, 59–66. [Google Scholar] [CrossRef]
- Katagiri, A.; Nakayama, K.; Rahman, M.T.; Rahman, M.; Katagiri, H.; Ishikawa, M.; Ishibashi, T.; Iida, K.; Otsuki, Y.; Nakayama, S.; et al. Frequent Loss of Tumor Suppressor ARID1A Protein Expression in Adenocarcinomas/Adenosquamous Carcinomas of the Uterine Cervix. Int. J. Gynecol. Cancer 2012, 22, 208–212. [Google Scholar] [CrossRef]
- Heinze, K.; Nazeran, T.M.; Lee, S.; Krämer, P.; Cairns, E.S.; Chiu, D.S.; Leung, S.C.; Kang, E.Y.; Meagher, N.S.; Kennedy, C.J.; et al. Validated Biomarker Assays Confirm That ARID1A Loss Is Confounded with MMR Deficiency, CD8+ TIL Infiltration, and Provides No Independent Prognostic Value in Endometriosis-Associated Ovarian Carcinomas. J. Pathol. 2022, 256, 388–401. [Google Scholar] [CrossRef]
- Mandal, J.; Mandal, P.; Wang, T.-L.; Shih, I.-M. Treating ARID1A Mutated Cancers by Harnessing Synthetic Lethality and DNA Damage Response. J. Biomed. Sci. 2022, 29, 71. [Google Scholar] [CrossRef]
- Tsai, S.; Fournier, L.-A.; Chang, E.Y.-C.; Wells, J.P.; Minaker, S.W.; Zhu, Y.D.; Wang, A.Y.-H.; Wang, Y.; Huntsman, D.G.; Stirling, P.C. ARID1A Regulates R-Loop Associated DNA Replication Stress. PLoS Genet. 2021, 17, e1009238. [Google Scholar] [CrossRef]
- Zhao, B.; Lin, J.; Rong, L.; Wu, S.; Deng, Z.; Fatkhutdinov, N.; Zundell, J.; Fukumoto, T.; Liu, Q.; Kossenkov, A.; et al. ARID1A Promotes Genomic Stability through Protecting Telomere Cohesion. Nat. Commun. 2019, 10, 4067. [Google Scholar] [CrossRef]
- Trizzino, M.; Barbieri, E.; Petracovici, A.; Wu, S.; Welsh, S.A.; Owens, T.A.; Licciulli, S.; Zhang, R.; Gardini, A. The Tumor Suppressor ARID1A Controls Global Transcription via Pausing of RNA Polymerase II. Cell Rep. 2018, 23, 3933–3945. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Fatkhutdinov, N.; Rosin, L.; Luppino, J.M.; Iwasaki, O.; Tanizawa, H.; Tang, H.-Y.; Kossenkov, A.V.; Gardini, A.; Noma, K.-I.; et al. ARID1A Spatially Partitions Interphase Chromosomes. Sci. Adv. 2019, 5, eaaw5294. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.R.; Bitler, B.G.; Schug, Z.; Conejo-Garcia, J.R.; Zhang, R.; Speicher, D.W. The Primary Effect on the Proteome of ARID1A-Mutated Ovarian Clear Cell Carcinoma Is Downregulation of the Mevalonate Pathway at the Post-Transcriptional Level. Mol. Cell. Proteom. MCP 2016, 15, 3348–3360. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Takahashi, K.; Sasaki, M.; Kuroda, T.; Yoshida, H.; Watanabe, R.; Maruyama, A.; Makinoshima, H.; Chiwaki, F.; Sasaki, H.; et al. Targeting the Vulnerability of Glutathione Metabolism in ARID1A-Deficient Cancers. Cancer Cell 2019, 35, 177–190.e8. [Google Scholar] [CrossRef]
- Zhang, X.; Shetty, M.; Clemente, V.; Linder, S.; Bazzaro, M. Targeting Mitochondrial Metabolism in Clear Cell Carcinoma of the Ovaries. Int. J. Mol. Sci. 2021, 22, 4750. [Google Scholar] [CrossRef]
- Zhang, W.; Chronis, C.; Chen, X.; Zhang, H.; Spalinskas, R.; Pardo, M.; Chen, L.; Wu, G.; Zhu, Z.; Yu, Y.; et al. The BAF and PRC2 Complex Subunits Dpf2 and Eed Antagonistically Converge on Tbx3 to Control ESC Differentiation. Cell Stem Cell 2019, 24, 138–152.e8. [Google Scholar] [CrossRef]
- Kadoch, C.; Copeland, R.A.; Keilhack, H. PRC2 and SWI/SNF Chromatin Remodeling Complexes in Health and Disease. Biochemistry 2016, 55, 1600–1614. [Google Scholar] [CrossRef]
- Wilson, M.R.; Reske, J.J.; Koeman, J.; Adams, M.; Joshi, N.R.; Fazleabas, A.T.; Chandler, R.L. SWI/SNF Antagonism of PRC2 Mediates Estrogen-Induced Progesterone Receptor Expression. Cells 2022, 11, 1000. [Google Scholar] [CrossRef]
- Bitler, B.G.; Aird, K.M.; Garipov, A.; Li, H.; Amatangelo, M.; Kossenkov, A.V.; Schultz, D.C.; Liu, Q.; Shih, I.-M.; Conejo-Garcia, J.R.; et al. Synthetic Lethality by Targeting EZH2 Methyltransferase Activity in ARID1A-Mutated Cancers. Nat. Med. 2015, 21, 231–238. [Google Scholar] [CrossRef]
- Yamada, L.; Saito, M.; Thar Min, A.K.; Saito, K.; Ashizawa, M.; Kase, K.; Nakajima, S.; Onozawa, H.; Okayama, H.; Endo, H.; et al. Selective Sensitivity of EZH2 Inhibitors Based on Synthetic Lethality in ARID1A-Deficient Gastric Cancer. Gastric Cancer 2021, 24, 60–71. [Google Scholar] [CrossRef]
- Sen, M.; Wang, X.; Hamdan, F.H.; Rapp, J.; Eggert, J.; Kosinsky, R.L.; Wegwitz, F.; Kutschat, A.P.; Younesi, F.S.; Gaedcke, J.; et al. ARID1A Facilitates KRAS Signaling-Regulated Enhancer Activity in an AP1-Dependent Manner in Colorectal Cancer Cells. Clin. Epigenet. 2019, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Berns, K.; Caumanns, J.J.; Hijmans, E.M.; Gennissen, A.M.C.; Severson, T.M.; Evers, B.; Wisman, G.B.A.; Jan Meersma, G.; Lieftink, C.; Beijersbergen, R.L.; et al. ARID1A Mutation Sensitizes Most Ovarian Clear Cell Carcinomas to BET Inhibitors. Oncogene 2018, 37, 4611–4625. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lyu, J.; Yang, E.J.; Liu, Y.; Zhang, B.; Shim, J.S. Targeting AURKA-CDC25C Axis to Induce Synthetic Lethality in ARID1A-Deficient Colorectal Cancer Cells. Nat. Commun. 2018, 9, 3212. [Google Scholar] [CrossRef]
- Hart, T.; Tong, A.H.Y.; Chan, K.; Van Leeuwen, J.; Seetharaman, A.; Aregger, M.; Chandrashekhar, M.; Hustedt, N.; Seth, S.; Noonan, A.; et al. Evaluation and Design of Genome-Wide CRISPR/SpCas9 Knockout Screens. G3 Genes Genomes Genet. 2017, 7, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Kim, E.; Hart, T. Improved Analysis of CRISPR Fitness Screens and Reduced Off-Target Effects with the BAGEL2 Gene Essentiality Classifier. Genome Med. 2021, 13, 2. [Google Scholar] [CrossRef]
- Mittal, P.; Roberts, C.W.M. The SWI/SNF Complex in Cancer—Biology, Biomarkers and Therapy. Nat. Rev. Clin. Oncol. 2020, 17, 435–448. [Google Scholar] [CrossRef]
- Mathur, R. ARID1A Loss in Cancer: Towards a Mechanistic Understanding. Pharmacol. Ther. 2018, 190, 15–23. [Google Scholar] [CrossRef]
- Hodges, C.; Kirkland, J.G.; Crabtree, G.R. The Many Roles of BAF (mSWI/SNF) and PBAF Complexes in Cancer. Cold Spring Harb. Perspect. Med. 2016, 6, a026930. [Google Scholar] [CrossRef]
- Lebedev, T.; Kousar, R.; Patrick, B.; Usama, M.; Lee, M.-K.; Tan, M.; Li, X.-G. Targeting ARID1A-Deficient Cancers: An Immune-Metabolic Perspective. Cells 2023, 12, 952. [Google Scholar] [CrossRef]
- Feng, X.; Tang, M.; Dede, M.; Su, D.; Pei, G.; Jiang, D.; Wang, C.; Chen, Z.; Li, M.; Nie, L.; et al. Genome-Wide CRISPR Screens Using Isogenic Cells Reveal Vulnerabilities Conferred by Loss of Tumor Suppressors. Sci. Adv. 2022, 8, eabm6638. [Google Scholar] [CrossRef] [PubMed]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576.e16. [Google Scholar] [CrossRef] [PubMed]
- Helming, K.C.; Wang, X.; Wilson, B.G.; Vazquez, F.; Haswell, J.R.; Manchester, H.E.; Kim, Y.; Kryukov, G.V.; Ghandi, M.; Aguirre, A.J.; et al. ARID1B Is a Specific Vulnerability in ARID1A-Mutant Cancers. Nat. Med. 2014, 20, 251–254. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Hur, W.; Sun, Z.; Jiang, T.; Mason, D.E.; Peters, E.C.; Zhang, D.D.; Luesch, H.; Schultz, P.G.; Gray, N.S. A Small-Molecule Inducer of the Antioxidant Response Element. Chem. Biol. 2010, 17, 537–547. [Google Scholar] [CrossRef]
- Hu, C.; Eggler, A.L.; Mesecar, A.D.; van Breemen, R.B. Modification of Keap1 Cysteine Residues by Sulforaphane. Chem. Res. Toxicol. 2011, 24, 515–521. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Taguchi, K.; Yamamoto, M. The KEAP1-NRF2 System in Cancer. Front. Oncol. 2017, 7, 85. [Google Scholar] [CrossRef]
- Kerins, M.J.; Ooi, A. A Catalogue of Somatic NRF2 Gain-of-Function Mutations in Cancer. Sci. Rep. 2018, 8, 12846. [Google Scholar] [CrossRef]
- ENCODE Project Consortium An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [CrossRef]
- Lachmann, A.; Xu, H.; Krishnan, J.; Berger, S.I.; Mazloom, A.R.; Ma’ayan, A. ChEA: Transcription Factor Regulation Inferred from Integrating Genome-Wide ChIP-X Experiments. Bioinformatics 2010, 26, 2438–2444. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Krupina, K.; Goginashvili, A.; Cleveland, D.W. Causes and Consequences of Micronuclei. Curr. Opin. Cell Biol. 2021, 70, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.T.; Miller, R.; Pemberton, H.N.; Jones, S.E.; Campbell, J.; Konde, A.; Badham, N.; Rafiq, R.; Brough, R.; Gulati, A.; et al. ATR Inhibitors as a Synthetic Lethal Therapy for Tumours Deficient in ARID1A. Nat. Commun. 2016, 7, 13837. [Google Scholar] [CrossRef]
- Shen, J.; Peng, Y.; Wei, L.; Zhang, W.; Yang, L.; Lan, L.; Kapoor, P.; Ju, Z.; Mo, Q.; Shih, I.-M.; et al. ARID1A Deficiency Impairs the DNA Damage Checkpoint and Sensitizes Cells to PARP Inhibitors. Cancer Discov. 2015, 5, 752–767. [Google Scholar] [CrossRef]
- Kadoch, C.; Williams, R.T.; Calarco, J.P.; Miller, E.L.; Weber, C.M.; Braun, S.M.G.; Pulice, J.L.; Chory, E.J.; Crabtree, G.R. Dynamics of BAF-Polycomb Complex Opposition on Heterochromatin in Normal and Oncogenic States. Nat. Genet. 2017, 49, 213–222. [Google Scholar] [CrossRef]
- Alldredge, J.K.; Eskander, R.N. EZH2 Inhibition in ARID1A Mutated Clear Cell and Endometrioid Ovarian and Endometrioid Endometrial Cancers. Gynecol. Oncol. Res. Pract. 2017, 4, 17. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, W.; Howard, T.P.; Vazquez, F.; Tsherniak, A.; Wu, J.N.; Wang, W.; Haswell, J.R.; Walensky, L.D.; Hahn, W.C.; et al. SWI/SNF-Mutant Cancers Depend on Catalytic and Non-Catalytic Activity of EZH2. Nat. Med. 2015, 21, 1491–1496. [Google Scholar] [CrossRef]
- Kopacz, A.; Kloska, D.; Forman, H.J.; Jozkowicz, A.; Grochot-Przeczek, A. Beyond Repression of Nrf2: An Update on Keap1. Free Radic. Biol. Med. 2020, 157, 63–74. [Google Scholar] [CrossRef]
- Chen, F.; Xiao, M.; Feng, J.; Wufur, R.; Liu, K.; Hu, S.; Zhang, Y. Different Inhibition of Nrf2 by Two Keap1 Isoforms α and β to Shape Malignant Behaviour of Human Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 10342. [Google Scholar] [CrossRef]
- Shah, S.Z.A.; Zhao, D.; Hussain, T.; Sabir, N.; Mangi, M.H.; Yang, L. P62-Keap1-NRF2-ARE Pathway: A Contentious Player for Selective Targeting of Autophagy, Oxidative Stress and Mitochondrial Dysfunction in Prion Diseases. Front. Mol. Neurosci. 2018, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.-S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The Selective Autophagy Substrate P62 Activates the Stress Responsive Transcription Factor Nrf2 through Inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Tamberg, N.; Tahk, S.; Koit, S.; Kristjuhan, K.; Kasvandik, S.; Kristjuhan, A.; Ilves, I. Keap1-MCM3 Interaction Is a Potential Coordinator of Molecular Machineries of Antioxidant Response and Genomic DNA Replication in Metazoa. Sci. Rep. 2018, 8, 12136. [Google Scholar] [CrossRef] [PubMed]
- Mulvaney, K.M.; Matson, J.P.; Siesser, P.F.; Tamir, T.Y.; Goldfarb, D.; Jacobs, T.M.; Cloer, E.W.; Harrison, J.S.; Vaziri, C.; Cook, J.G.; et al. Identification and Characterization of MCM3 as a Kelch-like ECH-Associated Protein 1 (KEAP1) Substrate. J. Biol. Chem. 2016, 291, 23719–23733. [Google Scholar] [CrossRef]
- Kopacz, A.; Rojo, A.I.; Patibandla, C.; Lastra-Martínez, D.; Piechota-Polanczyk, A.; Kloska, D.; Jozkowicz, A.; Sutherland, C.; Cuadrado, A.; Grochot-Przeczek, A. Overlooked and Valuable Facts to Know in the NRF2/KEAP1 Field. Free Radic. Biol. Med. 2022, 192, 37–49. [Google Scholar] [CrossRef]
- Lv, Y.; Lv, X.; Zhang, J.; Cao, G.; Xu, C.; Zhang, B.; Lin, W. BRD4 Targets the KEAP1-Nrf2-G6PD Axis and Suppresses Redox Metabolism in Small Cell Lung Cancer. Antioxidants 2022, 11, 661. [Google Scholar] [CrossRef]
- Hussong, M.; Börno, S.T.; Kerick, M.; Wunderlich, A.; Franz, A.; Sültmann, H.; Timmermann, B.; Lehrach, H.; Hirsch-Kauffmann, M.; Schweiger, M.R. The Bromodomain Protein BRD4 Regulates the KEAP1/NRF2-Dependent Oxidative Stress Response. Cell Death Dis. 2014, 5, e1195. [Google Scholar] [CrossRef]
- Huang, M.; Zhu, L.; Garcia, J.S.; Li, M.X.; Gentles, A.J.; Mitchell, B.S. Brd4 Regulates the Expression of Essential Autophagy Genes and Keap1 in AML Cells. Oncotarget 2018, 9, 11665–11676. [Google Scholar] [CrossRef]
- Song, S.; Nguyen, V.; Schrank, T.; Mulvaney, K.; Walter, V.; Wei, D.; Orvis, T.; Desai, N.; Zhang, J.; Hayes, D.N.; et al. Loss of SWI/SNF Chromatin Remodeling Alters NRF2 Signaling in Non-Small Cell Lung Carcinoma. Mol. Cancer Res. MCR 2020, 18, 1777–1788. [Google Scholar] [CrossRef]
- Hart, T.; Chandrashekhar, M.; Aregger, M.; Steinhart, Z.; Brown, K.R.; MacLeod, G.; Mis, M.; Zimmermann, M.; Fradet-Turcotte, A.; Sun, S.; et al. High-Resolution CRISPR Screens Reveal Fitness Genes and Genotype-Specific Cancer Liabilities. Cell 2015, 163, 1515–1526. [Google Scholar] [CrossRef]
- Mair, B.; Tomic, J.; Masud, S.N.; Tonge, P.; Weiss, A.; Usaj, M.; Tong, A.H.Y.; Kwan, J.J.; Brown, K.R.; Titus, E.; et al. Essential Gene Profiles for Human Pluripotent Stem Cells Identify Uncharacterized Genes and Substrate Dependencies. Cell Rep. 2019, 27, 599–615. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, D.R.; Campbell, K.R.; Greening, K.; Ho, G.C.; Hopkins, J.; Bui, M.; Douglas, J.M.; Sharlandjieva, V.; Munzur, A.D.; Lai, D.; et al. Single Cell Transcriptomes of Normal Endometrial Derived Organoids Uncover Novel Cell Type Markers and Cryptic Differentiation of Primary Tumours. J. Pathol. 2020, 252, 201–214. [Google Scholar] [CrossRef]
- Hart, T.; Moffat, J. BAGEL: A Computational Framework for Identifying Essential Genes from Pooled Library Screens. BMC Bioinform. 2016, 17, 164. [Google Scholar] [CrossRef]
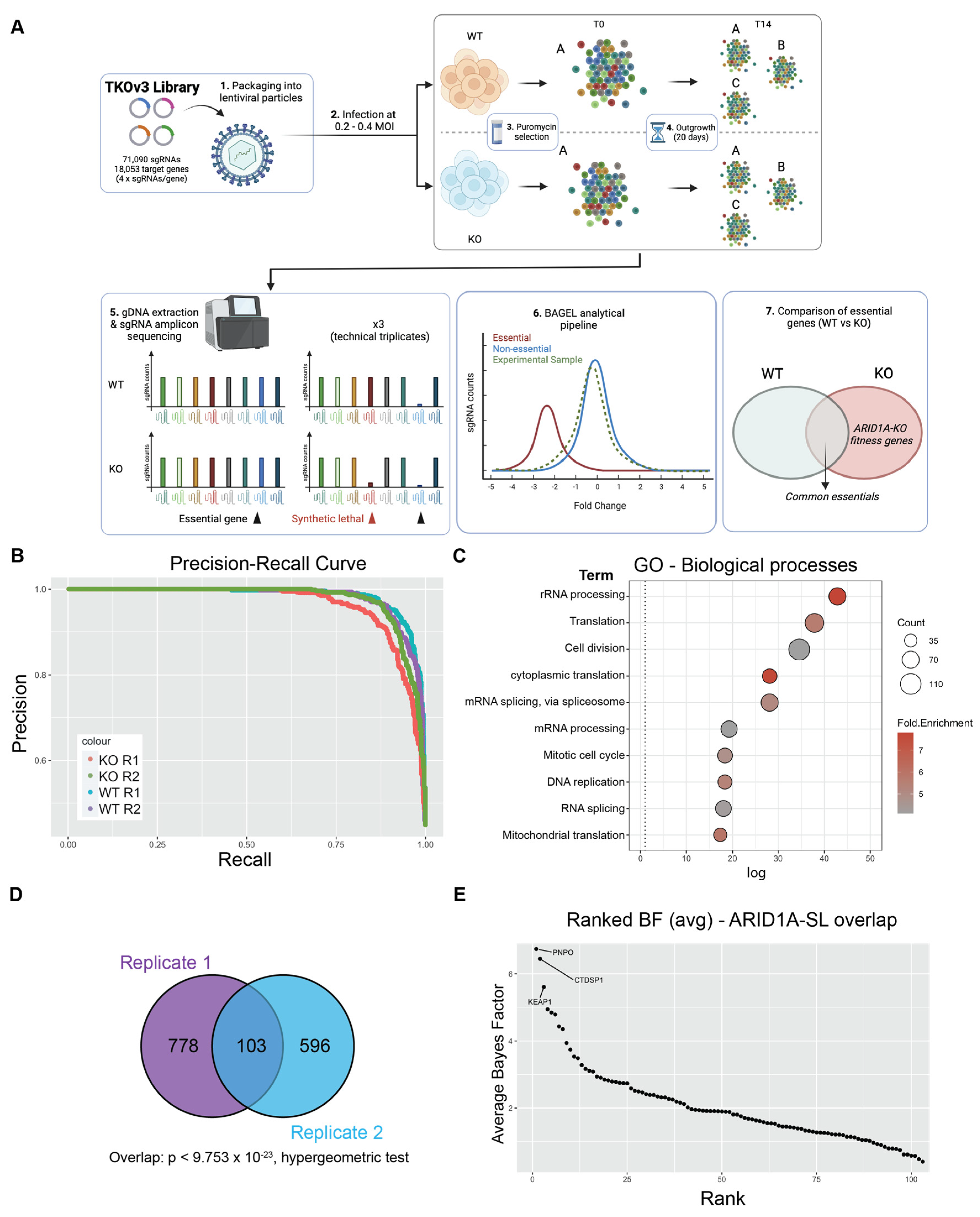
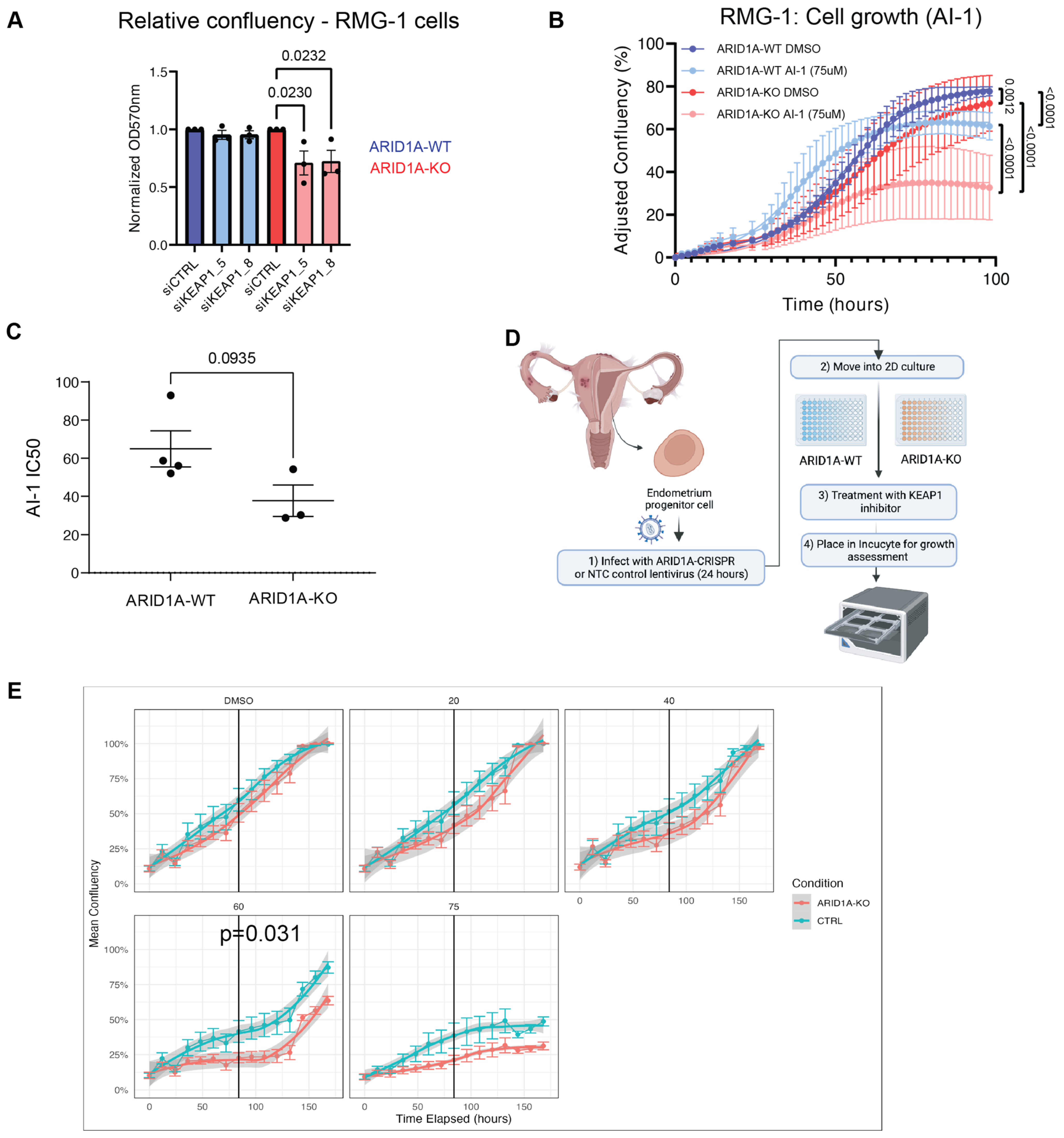
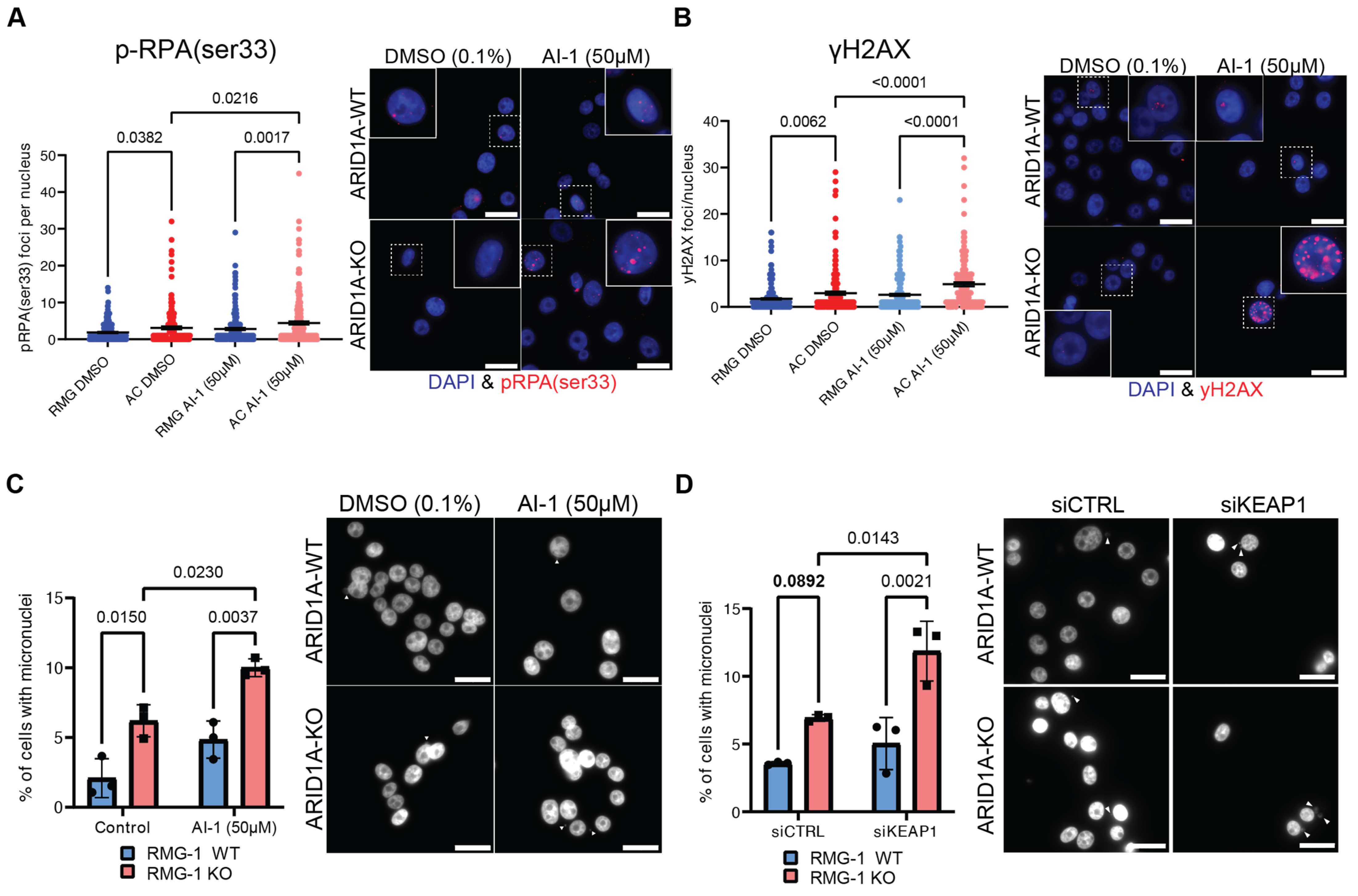
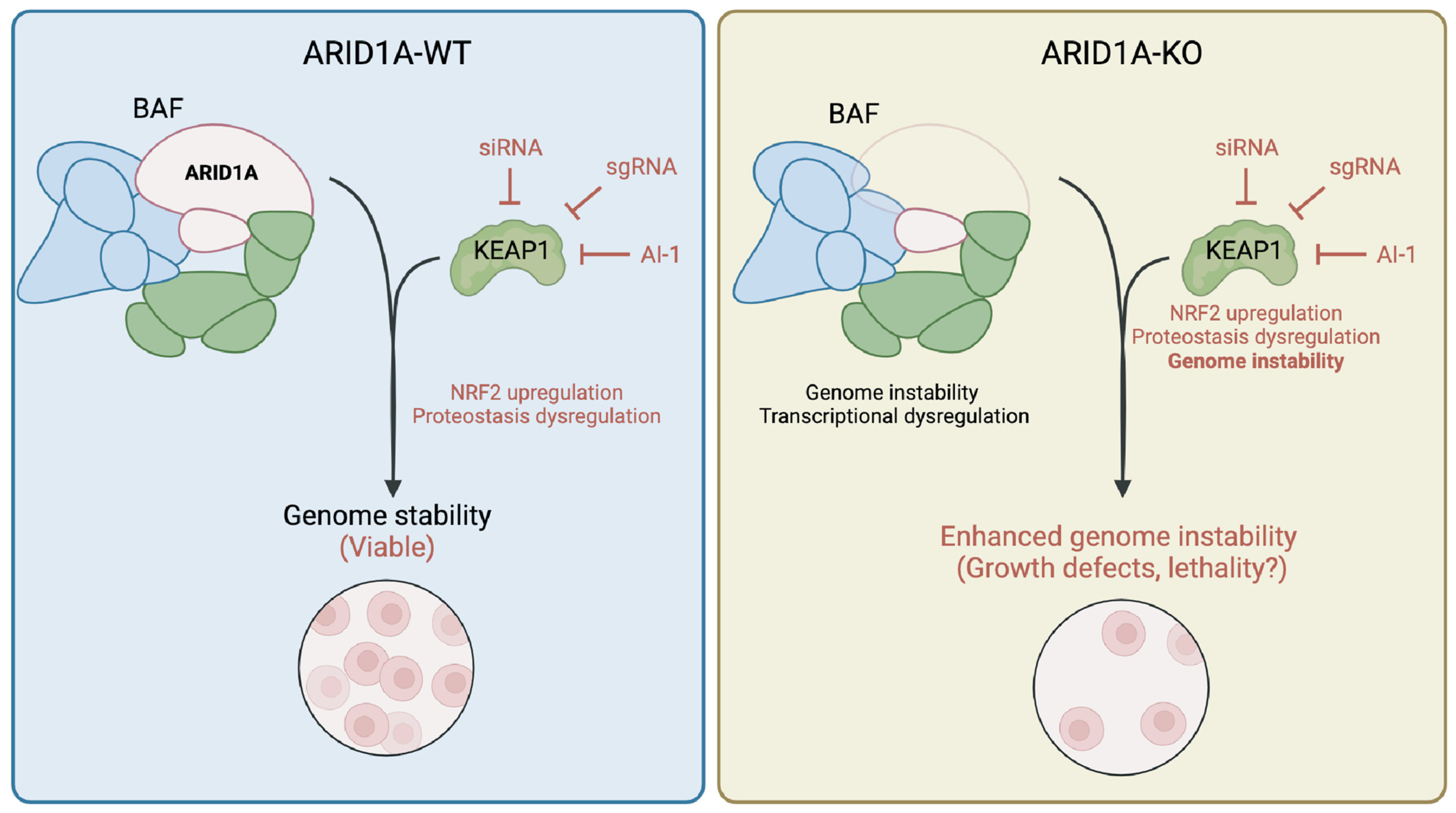
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).