
Hypoxia-Targeted Immunotherapy with PD-1 Blockade in Head and Neck Cancer

 , , , , , and
, , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Patients and Immunohistochemistry (IHC)
2.3. Cell Culture and Hypoxic Conditions
2.4. Synthetic Peptide
2.5. Flow Cytometry
2.6. In Vitro Induction of MTH1-Specific CD4+ T Cells
2.7. Antigen-Specific Responses by MTH1-Specific HTLs
2.8. Cytotoxicity Assay
2.9. Cell Cycle Assay
2.10. Statistical Analysis
3. Results
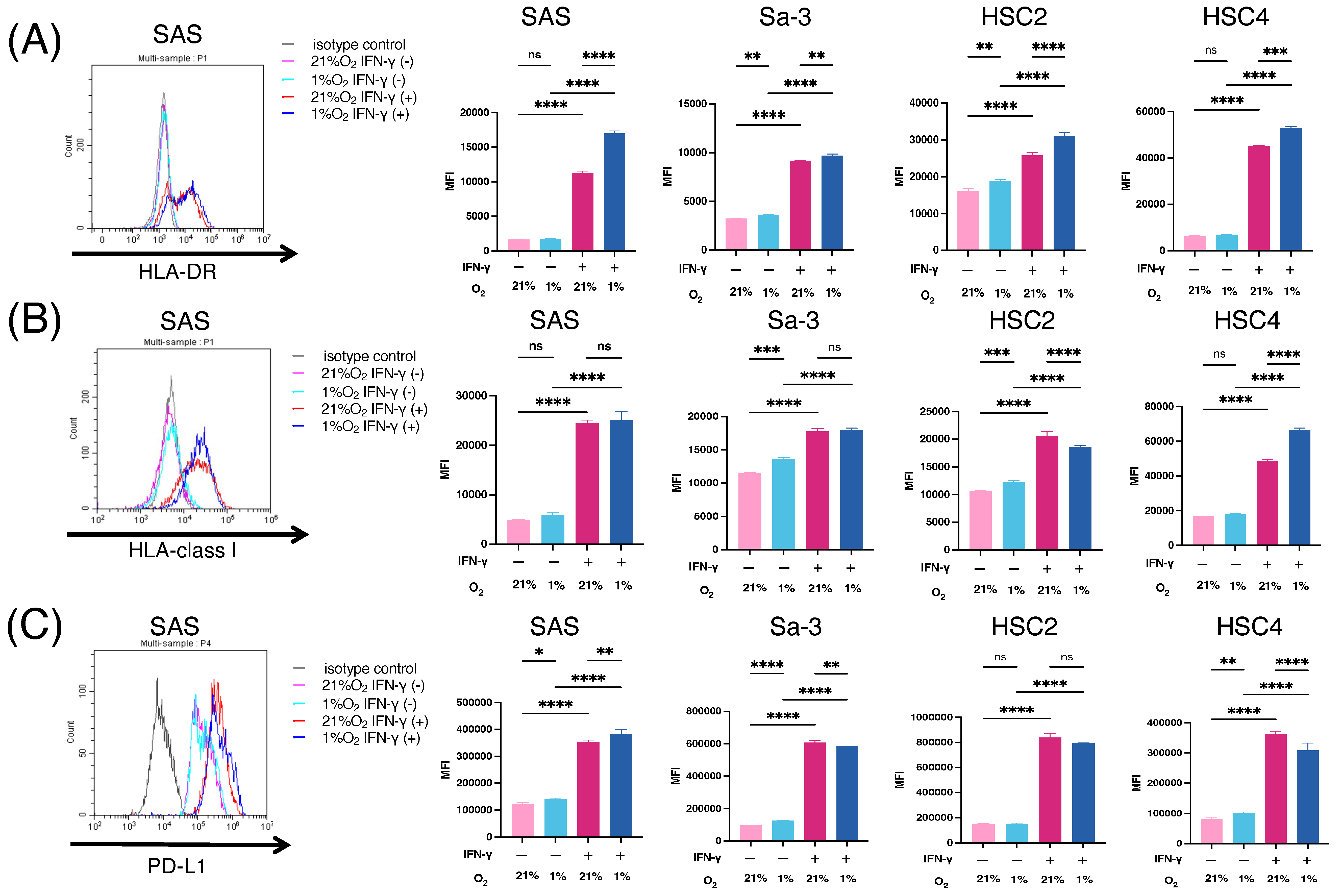
3.1. Immunogenic Changes in HNSCC Cell Lines under Hypoxic Condition
3.2. Viability and Proliferation of T Cells under Hypoxic Conditions
3.3. MTH1 Expression in Patients with HNSCC and HNSCC Cell Lines
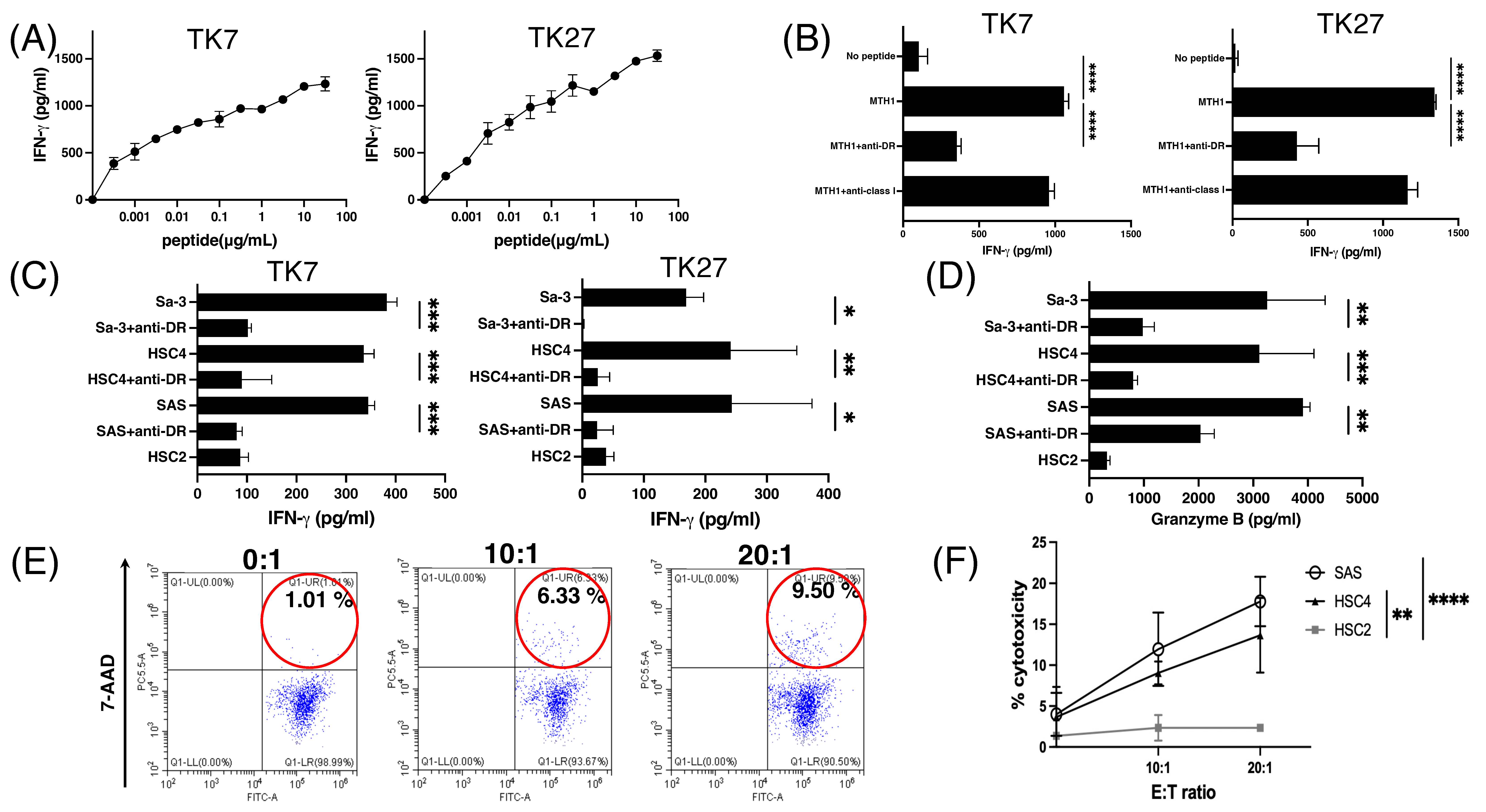
3.4. Generation of MTH145–59 Reactive CD4+ T-Cell Lines with Cytotoxic Activity
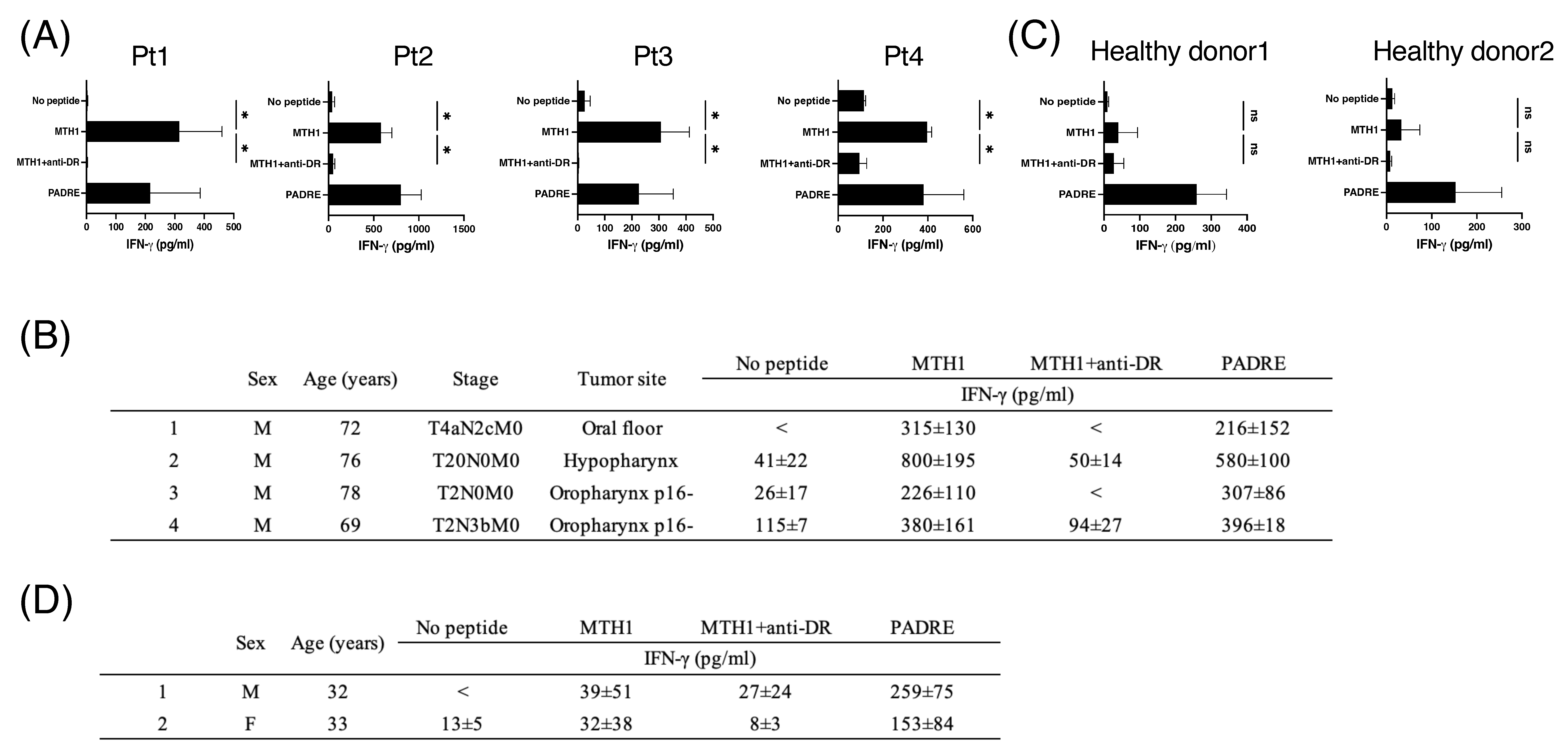
3.5. MTH145–59-Reactive T Cells in Periphery Blood from Patients with HNSCC
3.6. Antitumor Activity of MTH145–59-Reactive CD4+ T Cells with PD-1 Blockade in Hypoxia
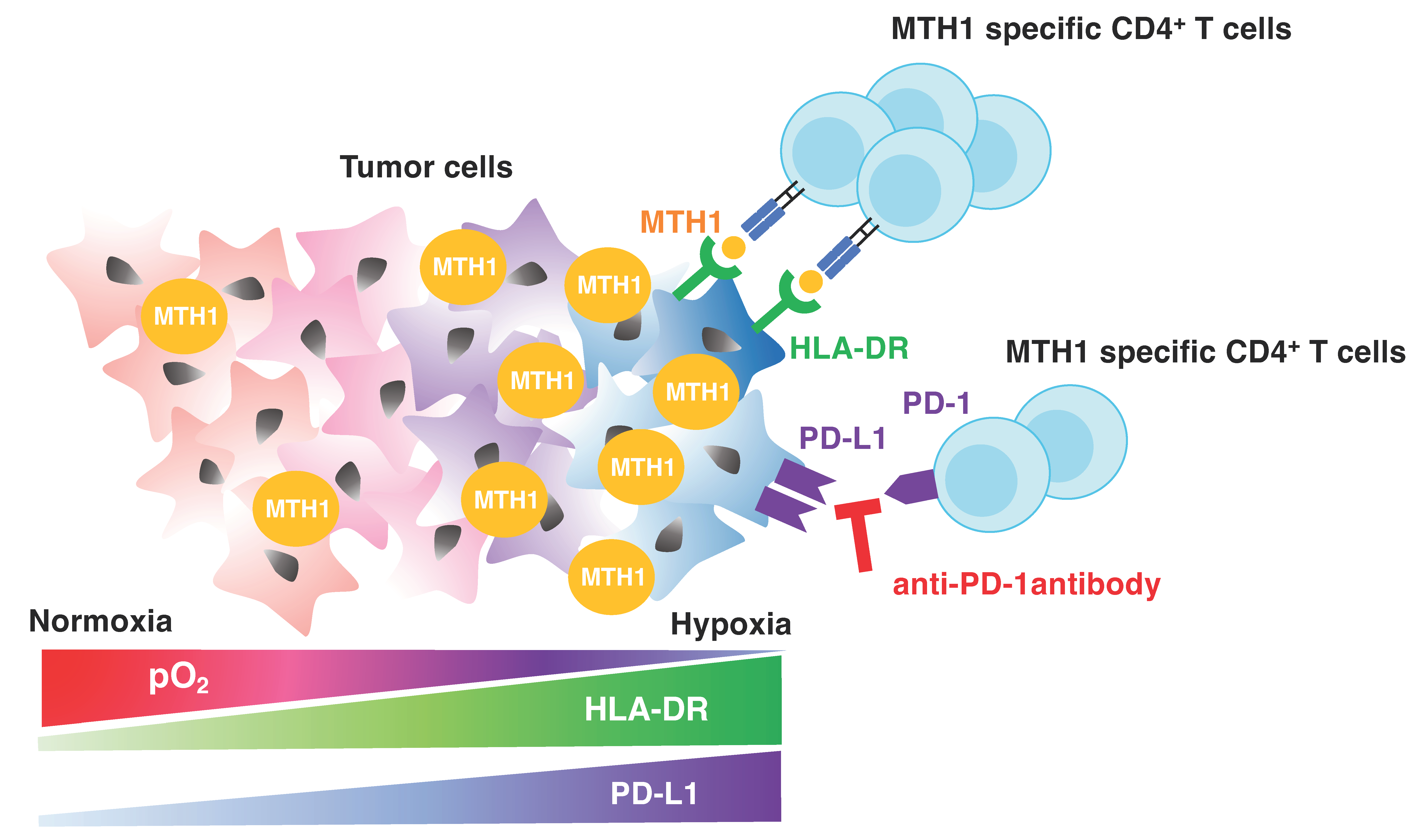
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mody, M.D.; Rocco, J.W.; Yom, S.S.; Haddad, R.I.; Saba, N.F. Head and neck cancer. Lancet 2021, 398, 2289–2299. [Google Scholar] [CrossRef] [PubMed]
- Starska-Kowarska, K. Dietary Carotenoids in Head and Neck Cancer-Molecular and Clinical Implications. Nutrients 2022, 14, 531. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef] [PubMed]
- McKeown, S.R. Defining normoxia, physoxia and hypoxia in tumours-implications for treatment response. Br. J. Radiol. 2014, 87, 20130676. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef]
- Vaupel, P.; Mayer, A. Hypoxia in cancer: Significance and impact on clinical outcome. Cancer Metastasis Rev. 2007, 26, 225–239. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, L.; Li, X.F. Hypoxia and the Tumor Microenvironment. Technol. Cancer Res. Treat. 2021, 20, 15330338211036304. [Google Scholar] [CrossRef]
- Infantino, V.; Santarsiero, A.; Convertini, P.; Todisco, S.; Iacobazzi, V. Cancer Cell Metabolism in Hypoxia: Role of HIF-1 as Key Regulator and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 5703. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 and tumor progression: Pathophysiology and therapeutics. Trends Mol. Med. 2002, 8, S62–S67. [Google Scholar] [CrossRef]
- Alessandrini, L.; Astolfi, L.; Daloiso, A.; Sbaraglia, M.; Mondello, T.; Zanoletti, E.; Franz, L.; Marioni, G. Diagnostic, Prognostic, and Therapeutic Role for Angiogenesis Markers in Head and Neck Squamous Cell Carcinoma: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 10733. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Yokota, A.; Harada, H.; Huang, G. Hypoxia/pseudohypoxia-mediated activation of hypoxia-inducible factor-1alpha in cancer. Cancer Sci. 2019, 110, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Cowman, S.J.; Koh, M.Y. Revisiting the HIF switch in the tumor and its immune microenvironment. Trends Cancer 2022, 8, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Masoud, G.N.; Li, W. HIF-1alpha pathway: Role, regulation and intervention for cancer therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Maynard, M.A.; Ohh, M. The role of hypoxia-inducible factors in cancer. Cell Mol. Life Sci. 2007, 64, 2170–2180. [Google Scholar] [CrossRef] [PubMed]
- Zhan, D.; Zhang, X.; Li, J.; Ding, X.; Cui, Y.; Jia, J. MTH1 Inhibitor TH287 Suppresses Gastric Cancer Development Through the Regulation of PI3K/AKT Signaling. Cancer Biother. Radiopharm. 2020, 35, 223–232. [Google Scholar] [CrossRef]
- Narwal, M.; Jemth, A.S.; Gustafsson, R.; Almlof, I.; Warpman Berglund, U.; Helleday, T.; Stenmark, P. Crystal Structures and Inhibitor Interactions of Mouse and Dog MTH1 Reveal Species-Specific Differences in Affinity. Biochemistry 2018, 57, 593–603. [Google Scholar] [CrossRef]
- Bhavya, B.; Easwer, H.V.; Vilanilam, G.C.; Anand, C.R.; Sreelakshmi, K.; Urulangodi, M.; Rajalakshmi, P.; Neena, I.; Padmakrishnan, C.J.; Menon, G.R.; et al. MutT Homolog1 has multifaceted role in glioma and is under the apparent orchestration by Hypoxia Inducible factor1 alpha. Life Sci. 2021, 264, 118673. [Google Scholar] [CrossRef]
- Gad, H.; Koolmeister, T.; Jemth, A.S.; Eshtad, S.; Jacques, S.A.; Strom, C.E.; Svensson, L.M.; Schultz, N.; Lundback, T.; Einarsdottir, B.O.; et al. MTH1 inhibition eradicates cancer by preventing sanitation of the dNTP pool. Nature 2014, 508, 215–221. [Google Scholar] [CrossRef]
- Kennedy, C.H.; Pass, H.I.; Mitchell, J.B. Expression of human MutT homologue (hMTH1) protein in primary non-small-cell lung carcinomas and histologically normal surrounding tissue. Free Radic. Biol. Med. 2003, 34, 1447–1457. [Google Scholar] [CrossRef]
- Wani, G.; Milo, G.E.; D’Ambrosio, S.M. Enhanced expression of the 8-oxo-7,8-dihydrodeoxyguanosine triphosphatase gene in human breast tumor cells. Cancer Lett. 1998, 125, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zheng, H.; Sun, L.H.; Peng, K.; Xiao, W.D.; Yang, H. Hypoxia-inducible factor-1 modulates upregulation of mutT homolog-1 in colorectal cancer. World J. Gastroenterol. 2015, 21, 13447–13456. [Google Scholar] [CrossRef] [PubMed]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef]
- Kumai, T.; Ohkuri, T.; Nagato, T.; Matsuda, Y.; Oikawa, K.; Aoki, N.; Kimura, S.; Celis, E.; Harabuchi, Y.; Kobayashi, H. Targeting HER-3 to elicit antitumor helper T cells against head and neck squamous cell carcinoma. Sci. Rep. 2015, 5, 16280. [Google Scholar] [CrossRef] [PubMed]
- Kumai, T.; Matsuda, Y.; Oikawa, K.; Aoki, N.; Kimura, S.; Harabuchi, Y.; Celis, E.; Kobayashi, H. EGFR inhibitors augment antitumour helper T-cell responses of HER family-specific immunotherapy. Br. J. Cancer 2013, 109, 2155–2166. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, F. Targeting human MutT homolog 1 (MTH1) for cancer eradication: Current progress and perspectives. Acta Pharm. Sin. B 2020, 10, 2259–2271. [Google Scholar] [CrossRef]
- Vaupel, P.; Mayer, A. Hypoxia in tumors: Pathogenesis-related classification, characterization of hypoxia subtypes, and associated biological and clinical implications. Adv. Exp. Med. Biol. 2014, 812, 19–24. [Google Scholar] [CrossRef]
- Abu-El-Rub, E.; Sareen, N.; Lester Sequiera, G.; Ammar, H.I.; Yan, W.; ShamsEldeen, A.M.; Rubinchik, I.; Moudgil, M.; Shokry, H.S.; Rashed, L.A.; et al. Hypoxia-induced increase in Sug1 leads to poor post-transplantation survival of allogeneic mesenchymal stem cells. FASEB J. 2020, 34, 12860–12876. [Google Scholar] [CrossRef]
- Kono, M.; Komatsuda, H.; Yamaki, H.; Kumai, T.; Hayashi, R.; Wakisaka, R.; Nagato, T.; Ohkuri, T.; Kosaka, A.; Ohara, K.; et al. Immunomodulation via FGFR inhibition augments FGFR1 targeting T-cell based antitumor immunotherapy for head and neck squamous cell carcinoma. Oncoimmunology 2022, 11, 2021619. [Google Scholar] [CrossRef]
- Siemens, D.R.; Hu, N.; Sheikhi, A.K.; Chung, E.; Frederiksen, L.J.; Pross, H.; Graham, C.H. Hypoxia increases tumor cell shedding of MHC class I chain-related molecule: Role of nitric oxide. Cancer Res. 2008, 68, 4746–4753. [Google Scholar] [CrossRef]
- Bailey, C.M.; Liu, Y.; Liu, M.; Du, X.; Devenport, M.; Zheng, P.; Liu, Y.; Wang, Y. Targeting HIF-1alpha abrogates PD-L1-mediated immune evasion in tumor microenvironment but promotes tolerance in normal tissues. J. Clin. Investig. 2022, 132, e150846. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.Z.; Desantis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. PD-L1 is a novel direct target of HIF-1alpha, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Minami, T.; Matsumura, N.; Sugimoto, K.; Shimizu, N.; De Velasco, M.; Nozawa, M.; Yoshimura, K.; Harashima, N.; Harada, M.; Uemura, H. Hypoxia-inducing factor (HIF)-1alpha-derived peptide capable of inducing cancer-reactive cytotoxic T lymphocytes from HLA-A24(+) patients with renal cell carcinoma. Int. Immunopharmacol. 2017, 44, 197–202. [Google Scholar] [CrossRef]
- Olin, M.R.; Andersen, B.M.; Zellmer, D.M.; Grogan, P.T.; Popescu, F.E.; Xiong, Z.; Forster, C.L.; Seiler, C.; SantaCruz, K.S.; Chen, W.; et al. Superior efficacy of tumor cell vaccines grown in physiologic oxygen. Clin. Cancer Res. 2010, 16, 4800–4808. [Google Scholar] [CrossRef] [PubMed]
- Alspach, E.; Lussier, D.M.; Miceli, A.P.; Kizhvatov, I.; DuPage, M.; Luoma, A.M.; Meng, W.; Lichti, C.F.; Esaulova, E.; Vomund, A.N.; et al. MHC-II neoantigens shape tumour immunity and response to immunotherapy. Nature 2019, 574, 696–701. [Google Scholar] [CrossRef] [PubMed]
- Kumai, T.; Matsuda, Y.; Ohkuri, T.; Oikawa, K.; Ishibashi, K.; Aoki, N.; Kimura, S.; Harabuchi, Y.; Celis, E.; Kobayashi, H. c-Met is a novel tumor associated antigen for T-cell based immunotherapy against NK/T cell lymphoma. Oncoimmunology 2015, 4, e976077. [Google Scholar] [CrossRef]
- Marty Pyke, R.; Thompson, W.K.; Salem, R.M.; Font-Burgada, J.; Zanetti, M.; Carter, H. Evolutionary Pressure against MHC Class II Binding Cancer Mutations. Cell 2018, 175, 416–428.e413. [Google Scholar] [CrossRef]
- Lhuillier, C.; Rudqvist, N.P.; Yamazaki, T.; Zhang, T.; Charpentier, M.; Galluzzi, L.; Dephoure, N.; Clement, C.C.; Santambrogio, L.; Zhou, X.K.; et al. Radiotherapy-exposed CD8+ and CD4+ neoantigens enhance tumor control. J. Clin. Investig. 2021, 131, e138740. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Negishi, Y.; Shimizu, M.; Takahashi, M.; Ichikawa, M.; Takahashi, H. Effects of extracellular pH and hypoxia on the function and development of antigen-specific cytotoxic T lymphocytes. Immunol. Lett. 2015, 167, 72–86. [Google Scholar] [CrossRef]
- Gropper, Y.; Feferman, T.; Shalit, T.; Salame, T.M.; Porat, Z.; Shakhar, G. Culturing CTLs under Hypoxic Conditions Enhances Their Cytolysis and Improves Their Anti-tumor Function. Cell Rep. 2017, 20, 2547–2555. [Google Scholar] [CrossRef]
- Kono, M.; Wakisaka, R.; Komatsuda, H.; Hayashi, R.; Kumai, T.; Yamaki, H.; Sato, R.; Nagato, T.; Ohkuri, T.; Kosaka, A.; et al. Immunotherapy targeting tumor-associated antigen in a mouse model of head and neck cancer. Head Neck 2024, 46, 2056–2067. [Google Scholar] [CrossRef] [PubMed]
- McPherson, L.A.; Troccoli, C.I.; Ji, D.; Bowles, A.E.; Gardiner, M.L.; Mohsen, M.G.; Nagathihalli, N.S.; Nguyen, D.M.; Robbins, D.J.; Merchant, N.B.; et al. Increased MTH1-specific 8-oxodGTPase activity is a hallmark of cancer in colon, lung and pancreatic tissue. DNA Repair 2019, 83, 102644. [Google Scholar] [CrossRef] [PubMed]
- Li, D.N.; Yang, C.C.; Li, J.; Ou Yang, Q.G.; Zeng, L.T.; Fan, G.Q.; Liu, T.H.; Tian, X.Y.; Wang, J.J.; Zhang, H.; et al. The high expression of MTH1 and NUDT5 promotes tumor metastasis and indicates a poor prognosis in patients with non-small-cell lung cancer. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118895. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, L.; Piao, S.; Li, L.; Li, J.; Xia, Y.; Li, J.; Saiyin, W. NUDT1: A potential independent predictor for the prognosis of patients with oral squamous cell carcinoma. J. Oral Pathol. Med. 2020, 49, 210–218. [Google Scholar] [CrossRef]
- Sun, J.; Zhao, Z.; Lu, J.; An, W.; Zhang, Y.; Li, W.; Yang, L. The Tumor Microenvironment Mediates the HIF-1alpha/PD-L1 Pathway to Promote Immune Escape in Colorectal Cancer. Int. J. Mol. Sci. 2024, 25, 3735. [Google Scholar] [CrossRef]
- Barsoum, I.B.; Smallwood, C.A.; Siemens, D.R.; Graham, C.H. A mechanism of hypoxia-mediated escape from adaptive immunity in cancer cells. Cancer Res. 2014, 74, 665–674. [Google Scholar] [CrossRef]
- Umbrello, M.; Dyson, A.; Feelisch, M.; Singer, M. The key role of nitric oxide in hypoxia: Hypoxic vasodilation and energy supply-demand matching. Antioxid. Redox Signal 2013, 19, 1690–1710. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, Y.; Zhang, L.; Song, W.; Shi, L. Hypoxia enhances indoleamine 2,3-dioxygenase production in dendritic cells. Oncotarget 2018, 9, 11572–11580. [Google Scholar] [CrossRef]
- Kim, M.; Tomek, P. Tryptophan: A Rheostat of Cancer Immune Escape Mediated by Immunosuppressive Enzymes IDO1 and TDO. Front. Immunol. 2021, 12, 636081. [Google Scholar] [CrossRef]
- Biswas, P.; Stuehr, D.J. Indoleamine dioxygenase and tryptophan dioxygenase activities are regulated through control of cell heme allocation by nitric oxide. J. Biol. Chem. 2023, 299, 104753. [Google Scholar] [CrossRef]
- Sultan, H.; Wu, J.; Fesenkova, V.I.; Fan, A.E.; Addis, D.; Salazar, A.M.; Celis, E. Poly-IC enhances the effectiveness of cancer immunotherapy by promoting T cell tumor infiltration. J. Immunother. Cancer 2020, 8, e001224. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Feature | MTH1 IHC Score | p Value | |
|---|---|---|---|
| Low | High | ||
| Gender | |||
| Female | 1 | 6 | 0.2187 |
| Male | 22 | 26 | |
| Age (years) | |||
| <70 | 14 | 17 | 0.5678 |
| ≥70 | 9 | 15 | |
| HPV status | |||
| Negative | 11 | 19 | 0.3962 |
| Positive | 12 | 13 | |
| T classificaion | |||
| T1–2 | 11 | 18 | 0.5371 |
| T3–4 | 12 | 14 | |
| N classification | |||
| N0–1 | 16 | 20 | 0.5868 |
| N2–3 | 7 | 12 | |
| Stage | |||
| I, II | 13 | 13 | 0.2441 |
| III, IV | 10 | 19 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wakisaka, R.; Yamaki, H.; Kono, M.; Inoue, T.; Sato, R.; Komatsuda, H.; Ohara, K.; Kosaka, A.; Ohkuri, T.; Nagato, T.; et al. Hypoxia-Targeted Immunotherapy with PD-1 Blockade in Head and Neck Cancer. Cancers 2024, 16, 3013. https://doi.org/10.3390/cancers16173013
Wakisaka R, Yamaki H, Kono M, Inoue T, Sato R, Komatsuda H, Ohara K, Kosaka A, Ohkuri T, Nagato T, et al. Hypoxia-Targeted Immunotherapy with PD-1 Blockade in Head and Neck Cancer. Cancers. 2024; 16(17):3013. https://doi.org/10.3390/cancers16173013
Chicago/Turabian StyleWakisaka, Risa, Hidekiyo Yamaki, Michihisa Kono, Takahiro Inoue, Ryosuke Sato, Hiroki Komatsuda, Kenzo Ohara, Akemi Kosaka, Takayuki Ohkuri, Toshihiro Nagato, and et al. 2024. "Hypoxia-Targeted Immunotherapy with PD-1 Blockade in Head and Neck Cancer" Cancers 16, no. 17: 3013. https://doi.org/10.3390/cancers16173013
APA StyleWakisaka, R., Yamaki, H., Kono, M., Inoue, T., Sato, R., Komatsuda, H., Ohara, K., Kosaka, A., Ohkuri, T., Nagato, T., Kishibe, K., Nakayama, K., Kobayashi, H., Kumai, T., & Takahara, M. (2024). Hypoxia-Targeted Immunotherapy with PD-1 Blockade in Head and Neck Cancer. Cancers, 16(17), 3013. https://doi.org/10.3390/cancers16173013