
Inorganic Nanomedicine—Mediated Ferroptosis: A Synergistic Approach to Combined Cancer Therapies and Immunotherapy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
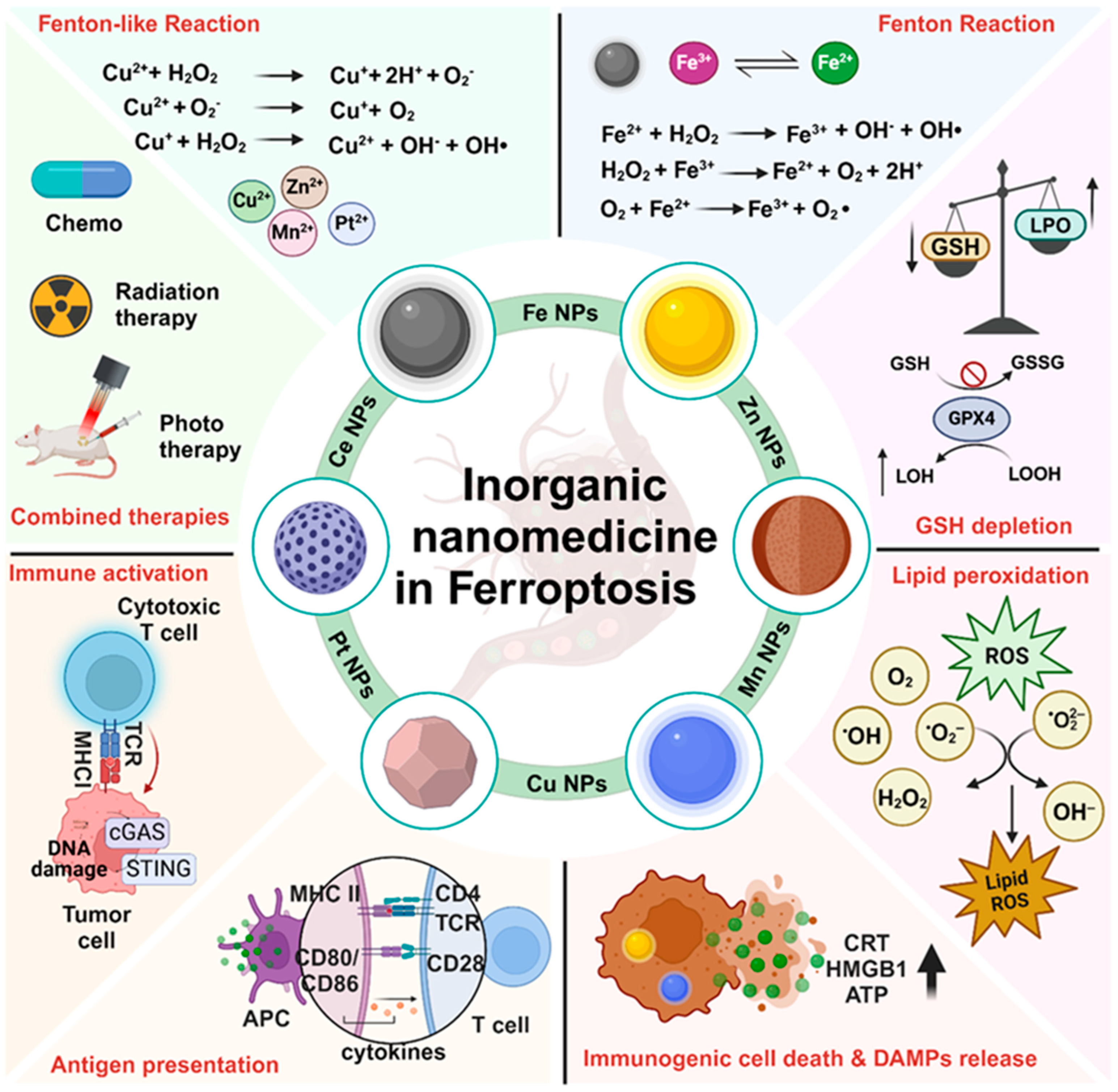
2. Inorganic Nanoparticles Induce Ferroptosis-Mediated Cell Death
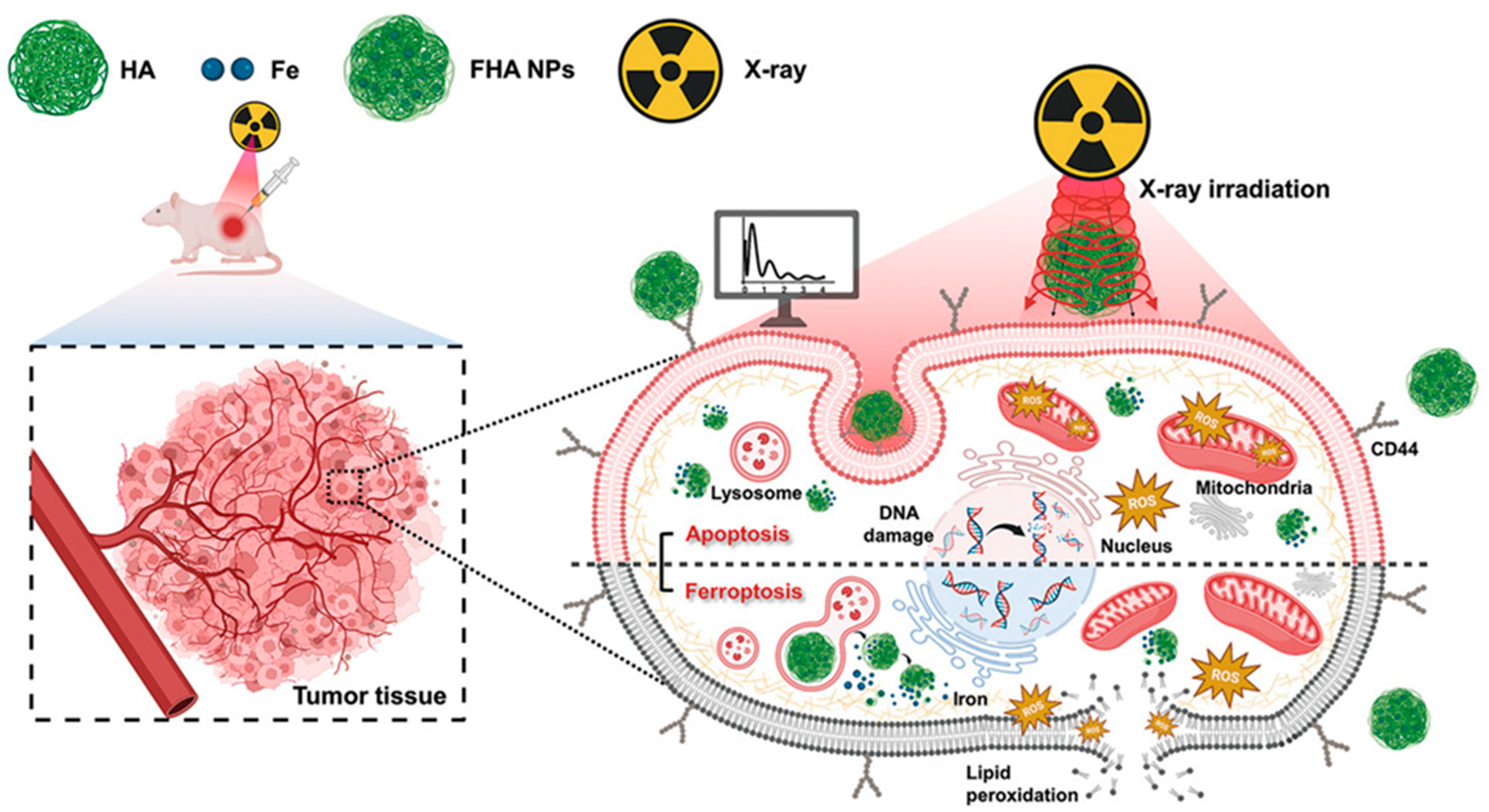
2.1. Iron Oxide NP-Based Ferroptosis
2.2. Copper NP-Based Ferroptosis
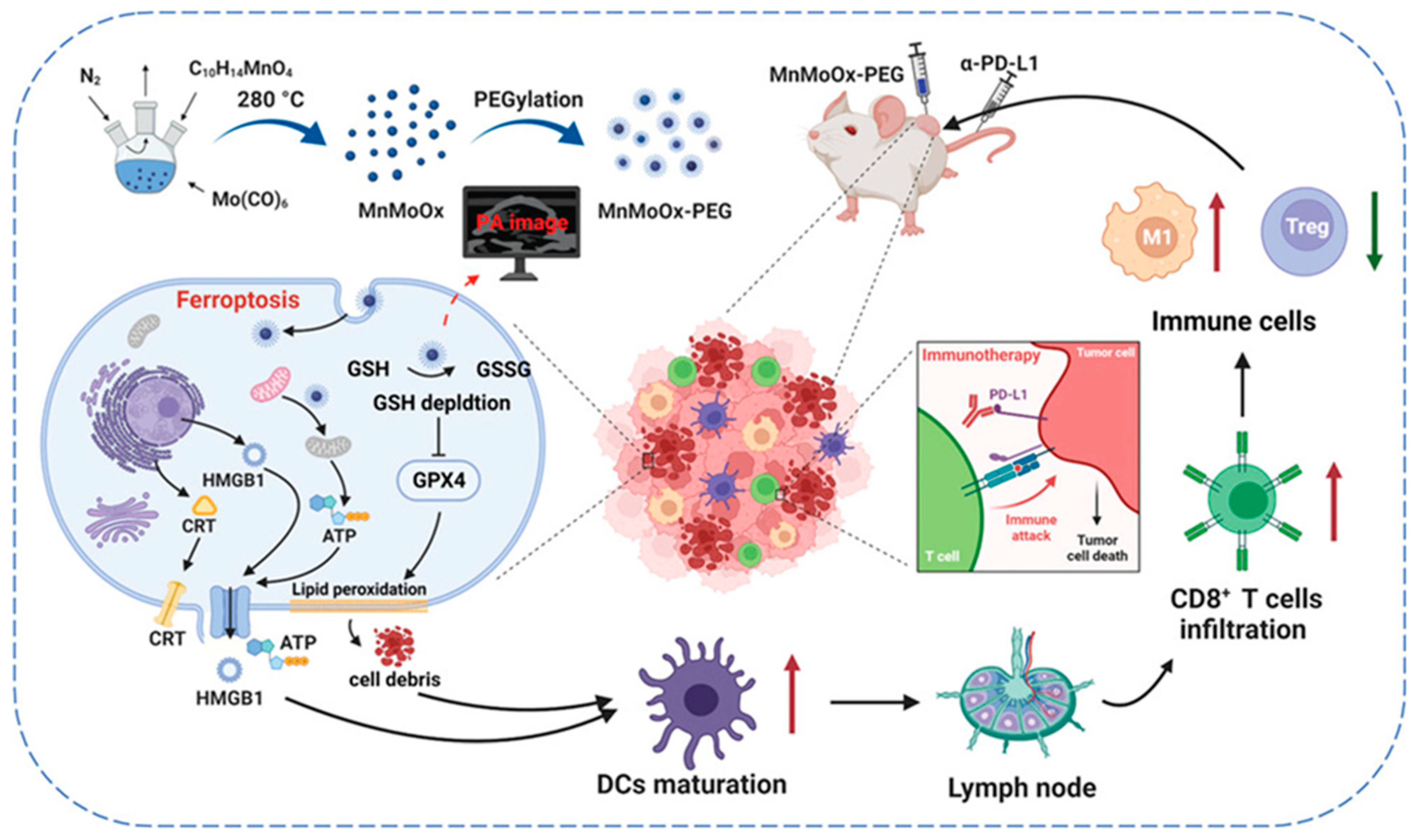
2.3. Manganese NP-Based Ferroptosis
2.4. Zinc NP-Based Ferroptosis
2.5. Platinum NP-Based Ferroptosis
3. Inorganic Nanoparticle-Based Ferroptosis in Combinational Anticancer Therapy
3.1. Ferroptosis and Cancer Immunotherapies
3.2. Ferroptosis Triggers Immunogenic Cell Death, Dendritic Cell Maturation, and T Cell Activation
3.3. Ferroptosis and Phototherapy
3.4. Ferroptosis and Radiotherapy
3.5. Ferroptosis and Chemotherapy
4. Clinical Challenges and Opportunities in Inorganic Nanoparticle-Based Ferroptosis
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROS | Reactive Oxygen Species |
| L-ROS | Lipid Reactive Oxygen Species |
| PUFAs | Polyunsaturated Fatty Acids |
| GPX4 | Glutathione Peroxidase 4 |
| GSH | Glutathione |
| HSBP1 | Heat Shock Factor-Binding Protein 1 |
| Nrf2 | Nuclear Factor Erythroid 2-Related Factor 2 |
| P53 | Tumor Protein P53 |
| ACSL4 | Acyl-CoA Synthetase Long-Chain Family Member 4 |
| MUFAs | Monounsaturated Fatty Acids |
| NPs | Nanoparticles |
| XC— | Cystine/Glutamate Antiporter System |
| RSL3 | Ras-Selective Lethal 3 |
| FIN56 | Ferroptosis Inducing Compound 56 |
| DPI2 | Diphenyleneiodonium Chloride 2 |
| Fe | Iron |
| Mn | Manganese |
| Cu | Copper |
| Pt | Platinum |
| PTT | Photothermal Therapy |
| PDT | Photodynamic Therapy |
| PEG | Poly(ethylene) Glycol |
| MRI | Magnetic Resonance Imaging |
| IFN-γ | Interferon Gamma |
| NOXs | NADPH Oxidases |
| TA | Tannic Acid |
| 1O2 | Singlet Oxygen |
| OH | Hydroxyl Radicals |
| TME | Tumor Microenvironment |
| Cu | Copper |
| Cin | Cinnamaldehyde |
| GOx | Glucose Oxidase |
| Au | Gold |
| TAX1BP1 | Tax1 Binding Protein 1 |
| Mn | Manganese |
| Pt (IV) | Platinum (IV) |
| ZAL | Zalcitabine |
| BSA | Bovine Serum Albumin |
| DAMPs | Damage-Associated Molecular Patterns |
| ICD | Immunogenic Cell Death |
| HMGB1 | High-Mobility Group Box 1 |
| CRT | Calreticulin |
| ATP | Adenosine Triphosphate |
| TME | Tumor Microenvironment |
| DCs | Dendritic Cells |
| MDSCs | Myeloid-derived suppressor cells |
| PLGA | Poly(lactic-co-glycolic Acid) |
| TNF-α | Tumor Necrosis Factor Alpha |
| IL-6 | Interleukin 6 |
| IL-12p70 | Interleukin 12 p70 |
| RNS | Reactive Nitrogen Species |
References
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Stockwell, B.R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem. Biol. 2008, 15, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, W.; Abdul Razak, S.R.; Han, T.; Ahmad, N.H.; Li, X. Ferroptosis as a potential target for cancer therapy. Cell Death Dis. 2023, 14, 460. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Yu, J.; Pan, H.; Lu, L. Novel insights on targeting ferroptosis in cancer therapy. Biomark. Res. 2020, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef]
- Gao, W.; Wang, X.; Zhou, Y.; Wang, X.; Yu, Y. Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct. Target. Ther. 2022, 7, 196. [Google Scholar] [CrossRef]
- Jacquemyn, J.; Ralhan, I.; Ioannou, M.S. Driving factors of neuronal ferroptosis. Trends Cell Biol. 2024, 34, 535–546. [Google Scholar] [CrossRef]
- Guiney, S.J.; Adlard, P.A.; Bush, A.I.; Finkelstein, D.I.; Ayton, S. Ferroptosis and cell death mechanisms in Parkinson’s disease. Neurochem. Int. 2017, 104, 34–48. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Freitas, F.P.; Seibt, T. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis. Cell 2018, 172, 409–422.E21. [Google Scholar] [CrossRef] [PubMed]
- Lewerenz, J.; Ates, G.; Methner, A.; Conrad, M.; Maher, P. Oxytosis/Ferroptosis-(Re-) Emerging Roles for Oxidative Stress-Dependent Non-apoptotic Cell Death in Diseases of the Central Nervous System. Front. Neurosci. 2018, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Zhang, J. Iron metabolism, ferroptosis, and the links with Alzheimer’s disease. Front. Neurosci. 2020, 13, 1443. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, L.; Ding, J.; Chen, Y. Iron metabolism in cancer. Int. J. Mol. Sci. 2018, 20, 95. [Google Scholar] [CrossRef]
- Li, D.; Li, Y. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Dixon, S.J.; Patel, D.N.; Welsch, M.; Skouta, R.; Lee, E.D.; Hayano, M.; Thomas, A.G.; Gleason, C.E.; Tatonetti, N.P.; Slusher, B.S.; et al. Pharmacological inhibition of cystine–glutamate exchange induces endoplasmic reticulum stress and ferroptosis. eLife 2014, 3, e02523. [Google Scholar] [CrossRef]
- Zheng, H.; Jiang, J.; Xu, S.; Liu, W.; Xie, Q.; Cai, X.; Zhang, J.; Liu, S.; Li, R. Nanoparticle-induced ferroptosis: Detection methods, mechanisms and applications. Nanoscale 2021, 13, 2266–2285. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, F.; Deng, J.; Dai, P.; Qin, Y.; Li, Z.; Wang, B.; Fan, A.; Wang, Z.; Zhao, Y. Electron-Accepting Micelles Deplete Reduced Nicotinamide Adenine Dinucleotide Phosphate and Impair Two Antioxidant Cascades for Ferroptosis-Induced Tumor Eradication. ACS Nano 2020, 14, 14715–14730. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, T.; Li, X.; Sheng, H.; Ma, X.; Hao, L. Ferroptosis-Inducing Nanomedicine for Cancer Therapy. Front. Pharmacol. 2021, 12, 735965. [Google Scholar] [CrossRef]
- Sheikh, A.; Kesharwani, P.; Almalki, W.H.; Almujri, S.S.; Dai, L.; Chen, Z.-S.; Sahebkar, A.; Gao, F. Understanding the Novel Approach of Nanoferroptosis for Cancer Therapy. Nano-Micro Lett. 2024, 16, 188. [Google Scholar] [CrossRef] [PubMed]
- Bae, C.; Kim, H.; Kook, Y.-M.; Lee, C.; Kim, C.; Yang, C.; Park, M.H.; Piao, Y.; Koh, W.-G.; Lee, K. Induction of ferroptosis using functionalized iron-based nanoparticles for anti-cancer therapy. Mater. Today Bio 2022, 17, 100457. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Lu, J.; Xi, J.; Xu, Z.; Fan, L.; Dai, H.; Gao, L. Metal ions/nucleotide coordinated nanoparticles comprehensively suppress tumor by synergizing ferroptosis with energy metabolism interference. J. Nanobiotechnology 2022, 20, 199. [Google Scholar] [CrossRef] [PubMed]
- Vallabani, N.S.; Singh, S.; Karakoti, A.S. Investigating the role of ATP towards amplified peroxidase activity of Iron oxide nanoparticles in different biologically relevant buffers. Appl. Surf. Sci. 2019, 492, 337–348. [Google Scholar] [CrossRef]
- Hu, X.; Li, R.; Wu, W.; Fang, K.; Zhu, Z.; Wang, Y.; Zhou, L.; Chen, M.; Dong, C.; Shi, S. A Fe (III)-porphyrin-oxaliplatin (IV) nanoplatform for enhanced ferroptosis and combined therapy. J. Control. Release 2022, 348, 660–671. [Google Scholar] [CrossRef]
- Yu, M.; Yu, J.; Yi, Y.; Chen, T.; Yu, L.; Zeng, W.; Ouyang, X.-k.; Huang, C.; Sun, S.; Wang, Y.; et al. Oxidative stress-amplified nanomedicine for intensified ferroptosis-apoptosis combined tumor therapy. J. Control. Release 2022, 347, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Song, W.F.; Zeng, J.Y.; Ji, P.; Han, Z.Y.; Sun, Y.X.; Zhang, X.Z. Self-assembled copper-based nanoparticles for glutathione activated and enzymatic cascade-enhanced ferroptosis and immunotherapy in cancer treatment. Small 2023, 19, 2301148. [Google Scholar] [CrossRef]
- Lin, X.; Chen, H.; Deng, T.; Cai, B.; Xia, Y.; Xie, L.; Wang, H.; Huang, C. Improved immune response for colorectal cancer therapy triggered by multifunctional nanocomposites with self-amplifying antitumor ferroptosis. ACS Appl. Mater. Interfaces 2024, 16, 13481–13495. [Google Scholar] [CrossRef]
- Xue, Q.; Yan, D.; Chen, X.; Li, X.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Chen, X.; Tang, D.; Liu, J. Copper-dependent autophagic degradation of GPX4 drives ferroptosis. Autophagy 2023, 19, 1982–1996. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Z.; Zhang, Z.; Pandey, N.K.; Amador, E.; Nguyen, W.; Chudal, L.; Xiong, L.; Chen, W.; Wen, Y. Copper-cysteamine nanoparticle-mediated microwave dynamic therapy improves cancer treatment with induction of ferroptosis. Bioact. Mater. 2023, 24, 322–330. [Google Scholar] [CrossRef]
- Lin, L.S.; Song, J.; Song, L.; Ke, K.; Liu, Y.; Zhou, Z.; Shen, Z.; Li, J.; Yang, Z.; Tang, W. Simultaneous Fenton-like ion delivery and glutathione depletion by MnO2-based nanoagent to enhance chemodynamic therapy. Angew. Chem. 2018, 130, 4996–5000. [Google Scholar] [CrossRef]
- Lv, M.; Chen, M.; Zhang, R.; Zhang, W.; Wang, C.; Zhang, Y.; Wei, X.; Guan, Y.; Liu, J.; Feng, K. Manganese is critical for antitumor immune responses via cGAS-STING and improves the efficacy of clinical immunotherapy. Cell Res. 2020, 30, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhu, Y.; Xing, X.; Xiao, J.; Chen, H.; Zhang, H.; Wang, D.; Zhang, Y.; Zhang, G.; Wu, Z.; et al. Manganese-deposited iron oxide promotes tumor-responsive ferroptosis that synergizes the apoptosis of cisplatin. Theranostics 2021, 11, 5418. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Tong, X.; Sun, Y.; Dong, C.; Li, C.; Wang, C.; Zhang, M.; Wen, Y.; Ye, P.; Li, R.; et al. A ferroptosis amplifier based on triple-enhanced lipid peroxides accumulation strategy for effective pancreatic cancer therapy. Biomaterials 2024, 309, 122574. [Google Scholar] [CrossRef] [PubMed]
- Asif, N.; Amir, M.; Fatma, T. Recent advances in the synthesis, characterization and biomedical applications of zinc oxide nanoparticles. Bioprocess Biosyst. Eng. 2023, 46, 1377–1398. [Google Scholar] [CrossRef]
- Liu, J.; Feng, X.; Wei, L.; Chen, L.; Song, B.; Shao, L. The toxicology of ion-shedding zinc oxide nanoparticles. Crit. Rev. Toxicol. 2016, 46, 348–384. [Google Scholar] [CrossRef]
- Anand, A.S.; Jain, K.; Chauhan, A.; Prasad, D.N.; Kohli, E. Zinc oxide nanoparticles trigger dysfunction of mitochondrial respiratory complexes and repair dynamics in human alveolar cells. Toxicol. Ind. Health 2023, 39, 127–137. [Google Scholar] [CrossRef]
- Naji, R.M.; Bashandy, M.A.; Fathy, A.H. Ameliorative effects of some natural antioxidants against blood and cardiovascular toxicity of oral subchronic exposure to silicon dioxide, aluminum oxide, or zinc oxide nanoparticles in wistar rats. Int. J. Food Sci. 2023, 2023, 8373406. [Google Scholar] [CrossRef]
- Singh, S. Zinc oxide nanoparticles impacts: Cytotoxicity, genotoxicity, developmental toxicity, and neurotoxicity. Toxicol. Mech. Methods 2019, 29, 300–311. [Google Scholar] [CrossRef]
- Ali, D.; Alarifi, S.; Kumar, S.; Ahamed, M.; Siddiqui, M.A. Oxidative stress and genotoxic effect of zinc oxide nanoparticles in freshwater snail Lymnaea luteola L. Aquat. Toxicol. 2012, 124, 83–90. [Google Scholar] [CrossRef]
- Wang, J.; Liu, M.; Wang, J.; Li, Z.; Feng, Z.; Xu, M.; Wang, H.; Li, H.; Li, Z.; Yu, J.; et al. Zinc oxide nanoparticles with catalase-like nanozyme activity and near-infrared light response: A combination of effective photodynamic therapy, autophagy, ferroptosis, and antitumor immunity. Acta Pharm. Sin. B 2024. [Google Scholar] [CrossRef]
- Wang, W.; Cai, J.; Wen, J.; Li, X.; Yu, Y.; Zhang, L.; Han, Q.; Wei, Z.; Ma, Y.; Ying, F.; et al. Boosting ferroptosis via abplatin(iv) for treatment of platinum-resistant recurrent ovarian cancer. Nano Today 2022, 44, 101459. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, S.-A.; Kerwood, D.J.; Goodisman, J.; Dabrowiak, J.C. Pt (IV) complexes as prodrugs for cisplatin. J. Inorg. Biochem. 2012, 107, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, B.; Han, Q.; Zhou, H.; Xia, Y.; Gong, C.; Dai, X.; Li, Z.; Wu, G. Ferroptosis: A novel anti-tumor action for cisplatin. Cancer Res. Treat. 2018, 50, 445–460. [Google Scholar] [CrossRef]
- del Valle, A.C.; Yeh, C.-K.; Huang, Y.-F. Near Infrared-Activatable Platinum-Decorated Gold Nanostars for Synergistic Photothermal/Ferroptotic Therapy in Combating Cancer Drug Resistance. Adv. Healthc. Mater. 2020, 9, 2000864. [Google Scholar] [CrossRef]
- Yu, B.; Choi, B.; Li, W.; Kim, D.-H. Magnetic field boosted ferroptosis-like cell death and responsive MRI using hybrid vesicles for cancer immunotherapy. Nat. Commun. 2020, 11, 3637. [Google Scholar] [CrossRef]
- Liu, Y.; Niu, R.; Zhao, H.; Wang, Y.; Song, S.; Zhang, H.; Zhao, Y. Single-Site Nanozymes with a Highly Conjugated Coordination Structure for Antitumor Immunotherapy via Cuproptosis and Cascade-Enhanced T Lymphocyte Activity. J. Am. Chem. Soc. 2024, 146, 3675–3688. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Li, Q.; Pei, Z.; Liu, L.; Yang, N.; Cheng, L. Nonferrous Ferroptosis Inducer Manganese Molybdate Nanoparticles to Enhance Tumor Immunotherapy. Small 2023, 19, 2303438. [Google Scholar] [CrossRef]
- Yang, C.; Wang, M.; Chang, M.; Yuan, M.; Zhang, W.; Tan, J.; Ding, B.; Ma, P.; Lin, J. Heterostructural Nanoadjuvant CuSe/CoSe2 for Potentiating Ferroptosis and Photoimmunotherapy through Intratumoral Blocked Lactate Efflux. J. Am. Chem. Soc. 2023, 145, 7205–7217. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Z.; Li, L.; Wang, M.; Chang, J.; Gao, M.; Wang, D.; Li, C. Ultra-small Janus nanoparticle-induced activation of ferroptosis for synergistic tumor immunotherapy. Acta Biomater. 2024, 181, 362–374. [Google Scholar] [CrossRef]
- Bae, C.; Hernández Millares, R.; Ryu, S.; Moon, H.; Kim, D.; Lee, G.; Jiang, Z.; Park, M.H.; Kim, K.H.; Koom, W.S.; et al. Synergistic Effect of Ferroptosis-Inducing Nanoparticles and X-Ray Irradiation Combination Therapy. Small 2024, 20, 2310873. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Cheng, X.; Xu, P.; Gu, Y.; Xu, H.; Xu, J.; Wang, Y.; Zhang, L.W.; Wang, Y. Radiotherapy-Driven Nanoprobes Targeting for Visualizing Tumor Infiltration Dynamics and Inducing Ferroptosis in Myeloid-Derived Suppressor Cells. J. Am. Chem. Soc. 2024, 146, 22455–22468. [Google Scholar] [CrossRef] [PubMed]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of glutathione in cancer progression and chemoresistance. Oxid. Med. Cell Longev. 2013, 2013, 972913. [Google Scholar] [CrossRef]
- Roy, N.; Paira, P. Glutathione Depletion and Stalwart Anticancer Activity of Metallotherapeutics Inducing Programmed Cell Death: Opening a New Window for Cancer Therapy. ACS Omega 2024, 9, 20670–20701. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Glandorff, C.; Sun, M. GSH and Ferroptosis: Side-by-Side Partners in the Fight against Tumors. Antioxidants 2024, 13, 697. [Google Scholar] [CrossRef]
- Li, Y.; Fan, W.; Gu, X.; Liu, S.; He, T.; Gou, S.; Meng, W.; Li, M.; Liu, X.; Ren, Y.; et al. Biodegradable Ferric Phosphate Nanocarriers with Tumor-Specific Activation and Glutathione Depletion for Tumor Self-Enhanced Ferroptosis and Chemotherapy. Adv. Funct. Mater. 2024, 34, 2313540. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohapatra, A.; Mohanty, A.; Park, I.-K. Inorganic Nanomedicine—Mediated Ferroptosis: A Synergistic Approach to Combined Cancer Therapies and Immunotherapy. Cancers 2024, 16, 3210. https://doi.org/10.3390/cancers16183210
Mohapatra A, Mohanty A, Park I-K. Inorganic Nanomedicine—Mediated Ferroptosis: A Synergistic Approach to Combined Cancer Therapies and Immunotherapy. Cancers. 2024; 16(18):3210. https://doi.org/10.3390/cancers16183210
Chicago/Turabian StyleMohapatra, Adityanarayan, Ayeskanta Mohanty, and In-Kyu Park. 2024. "Inorganic Nanomedicine—Mediated Ferroptosis: A Synergistic Approach to Combined Cancer Therapies and Immunotherapy" Cancers 16, no. 18: 3210. https://doi.org/10.3390/cancers16183210
APA StyleMohapatra, A., Mohanty, A., & Park, I.-K. (2024). Inorganic Nanomedicine—Mediated Ferroptosis: A Synergistic Approach to Combined Cancer Therapies and Immunotherapy. Cancers, 16(18), 3210. https://doi.org/10.3390/cancers16183210