

A Plausible Framework Reveals Potential Similarities in the Regulation of Immunity against Some Cancers and Some Infectious Agents: Implications for Prevention and Treatment


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Prospect: The Impetus for Writing This Paper
2. Non-Contentious Issues That Provide the Context for Discussing Contemporary Issues That Should Be Challenged
- Classical studies gave rise to the view that there are central and peripheral mechanisms of tolerance against self-antigens [6]. Self-antigens, when sufficiently present in primary lymphoid organs where lymphocytes are generated, cause the ablation of their corresponding lymphocytes [7] by a mechanism first proposed by Lederberg [8]. This mechanism accounts for central tolerance. Some self-antigens, of which insulin is a prototype, are insufficiently present in the primary lymphoid organs to ablate all the lymphocytes specific for these antigens [9]. Such self-antigens are called peripheral self-antigens. It is recognized that there must be a mechanism by which peripheral self-antigens can inactivate their corresponding lymphocytes, which emigrate from primary lymphoid organs, in order to ensure self-tolerance [6]. This inactivation process results in peripheral tolerance.
- The two-signal model of lymphocyte activation, which proposes how antigen can activate and inactivate mature, naïve lymphocytes, was proposed in 1970 [10]. This theory posited that the activation of a target lymphocyte required its antigen-mediated interaction with a helper lymphocyte. The activation of all lymphocytes thus requires lymphocyte cooperation. Antigen inactivates single lymphocytes. The theory was proposed because it accounted for diverse observations, some paradoxical at the time, and because it accounted for how peripheral tolerance can be achieved. This latter feature is germane to the current considerations.
- The theory explained peripheral tolerance in the context of an idea first proposed by Burnet and Fenner [11] and later extended by Lederberg [9]. The extended idea was that tolerance to self-antigens requires their early presence in development, or in the history of the individual, before lymphocytes are generated, and their continuous presence thereafter. This idea is referred to as The Historical Postulate. Thus, lymphocytes specific for peripheral self-antigens are inactivated as they are generated, one or a few at a time, by virtue of the early and continuous presence of the peripheral self-antigen. Lymphocytes specific for a foreign antigen, F, accumulate in its absence; once F impinges upon the immune system, it can mediate the interactions between the accumulated lymphocytes required to generate an immune response [12].
- It is convenient to refer to the signal generated from the interaction of antigen with the antigen-specific receptors of the lymphocyte as signal 1, and the delivery of the signal to the target lymphocyte, following the recognition of antigen by the helper lymphocyte, as signal 2. The generation of signal 1 alone is envisaged to result, in time, in the inactivation of the lymphocyte. The activation of the target lymphocyte requires the generation of both signal 1 and of signal 2 [10]. This theory is often referred to as the two-signal model of lymphocyte activation. However, there are other two-signal models of lymphocyte activation, as I make clear below.Much evidence supports this two-signal model. Thus, it is generally accepted that the activation of most B cells [13,14,15,16] and CD8 T cells [17], to, respectively generate antibody-producing cells and cytotoxic T lymphocytes (CTL), requires activated CD4 T helper cells. In the absence of this help, antigen inactivates the B [18,19] and the CD8 T cells [20]. Such observations, made over the years, have led virtually all immunologists to recognize the critical importance of activating and inactivating CD4 T helper cells. The central importance of helper T cells is acknowledged by referring to them as “the conductor of the immunological orchestra”. This topic of how antigen activates and inactivates CD4 T cells is the gate to controversy.
- It became evident by the late 1970s that the antigen-specific receptors of T cells recognize peptides, derived by processing the nominal antigen, bound to class I or II MHC molecules [21,22,23]. Antigen-presenting cells (APC) had the means of taking an antigen up, processing it, and presenting it [24]. Various discoveries led to the realization that the APC, in order to activate the CD4 T cell, had to deliver a co-stimulatory signal to the CD4 T cell. This required the APC to bear costimulatory molecules and the CD4 T cell to bear counter receptors. It was envisaged that the CD4 T cell was inactivated in the absence of costimulatory signals [25,26,27]. An interaction between the B7 molecules on the APC, and their counter receptors, CD28, expressed by CD4 T cells, represents the prototypical costimulatory (CoS) signal [28].
3. The PAMP/DAMP-Centric View on How Immune Responses Are Regulated
4. The Variables of Immunization Known to Affect the Th1/Th2 Phenotype of the Ensuing Immune Response and the Physiological Significance of the Class of Immunity Generated
- (a)
- Pearson and Raffel argued, on the basis of observation, that minimally foreign antigens, due either to their small size or being larger but only varying slightly from a corresponding self-antigen, are only able to generate Th1, delayed-type hypersensitivity (DTH) responses [39].
- (b)
- (c)
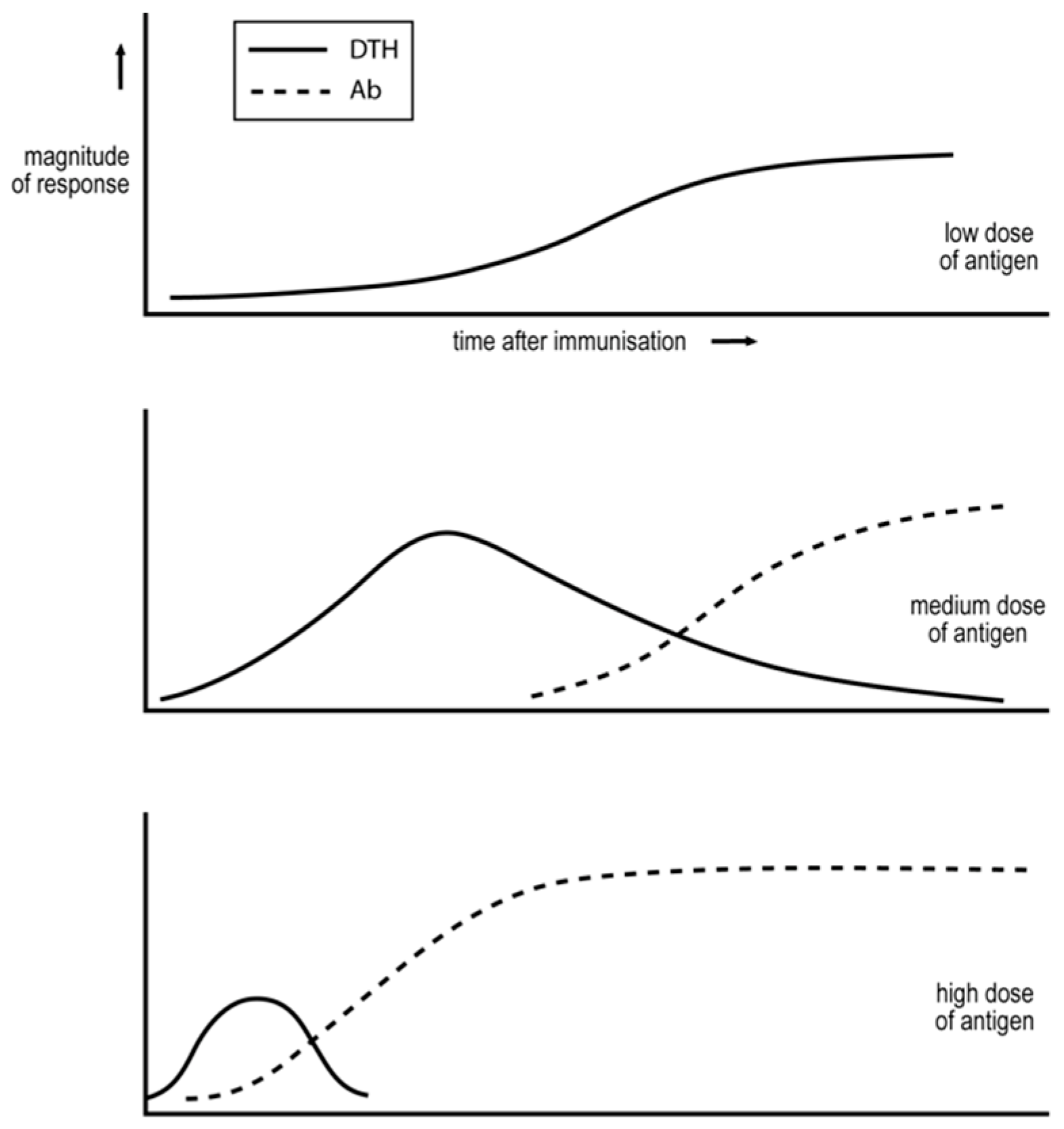
- Salvin also examined the effect of antigen dose [40]. Low doses can generate an exclusive Th1, DTH response; see Figure 1. Medium doses result in the more rapid generation of a Th1 response and, with time, Th2 cells appear and copious IgG antibody is produced. Even higher doses result in an even more rapid or transient Th1 response, and in a predominant Th2 response.
- (d)
- Immunization of an animal in a manner that results in an IgG antibody humoral response renders the animal unable to generate a DTH Th1 response to a challenge that generates such a response in a naïve animal [42]. The animal’s response to this antigen appears to have been locked into a humoral, IgG antibody mode. This phenomenon is referred to as “humoral immune deviation” [5]. Parish showed that repetitive immunization, over several weeks, of animals with a low dose of antigen, sub-immunogenic for an antibody response, resulted in a state of DTH, which is now recognized as being associated with the generation of Th1 cells. These exposed animals were partially unresponsive/unresponsive for the production of antibody [43,44]. This phenomenon is referred to as “cell-mediated immune deviation”.
- (e)
- It was first recognized, in studies on immunity of leprosy patients, that the class of immunity generated against the pathogen largely determines the course of disease. In this case, the generation of a stable and predominant Th1 cell-mediated response is associated with minimal disease, and mixed and predominant antibody responses with distinct but severe pathologies [45]. In more recent decades, the mouse model of the human disease of cutaneous leishmaniasis has become the major experimental model for a disease caused by a pathogen preferentially contained by cell-mediated attack [46]. The pertinent intracellular, protozoan pathogen, Leishmania major, inhabits macrophages. We will discuss below several studies conducted using this experimental system.
5. The Threshold Hypothesis Explains How the Th1/Th2 Phenotype of the Response Is Determined
6. The Plausibility/Implausibility of the PAMP/DAMP View
- It is known that the dose of antigen is important for determining the Th1/Th2 phenotype of the response in the manner outlined above, with lower doses favoring Th1 responses. This is true of vertebrate, PAMP-free proteins, administered with adjuvant containing PAMPs [40]; of more complex, foreign, PAMP-free vertebrate antigens, administered without adjuvant, such as SRBC in mice [47]; of protozoa administered to mice [52,53]; and of mycobacteria administered to mice [5] and to cattle [54]. The protozoa and mycobacteria clearly bear very different PAMPs. It seems likely, given the generality of this dependence of the class of immunity on antigen dose, generated in response to diverse types of antigen, that a PAMP-independent mechanism is likely critical, as envisaged by the threshold mechanism [5,38].
- Most immune responses evolve from an exclusive Th1 mode to one with a significant Th2 component. This is true of foreign, vertebrate, PAMP-free protein antigens, administered in PAMP-containing adjuvants [40]; of foreign, vertebrate, PAMP-free, and chemically complex antigens, administered without adjuvant, such as SRBC in mice [46]; of protozoa administered to mice [52,53]; and of mycobacteria administered to mice [5] and cattle [54]. Again, the generality of this pattern in responses to such diverse antigens leads to the idea that it can only be accounted for by a PAMP/DAMP-independent mechanism. Such a mechanism is provided by The Threshold Hypothesis.
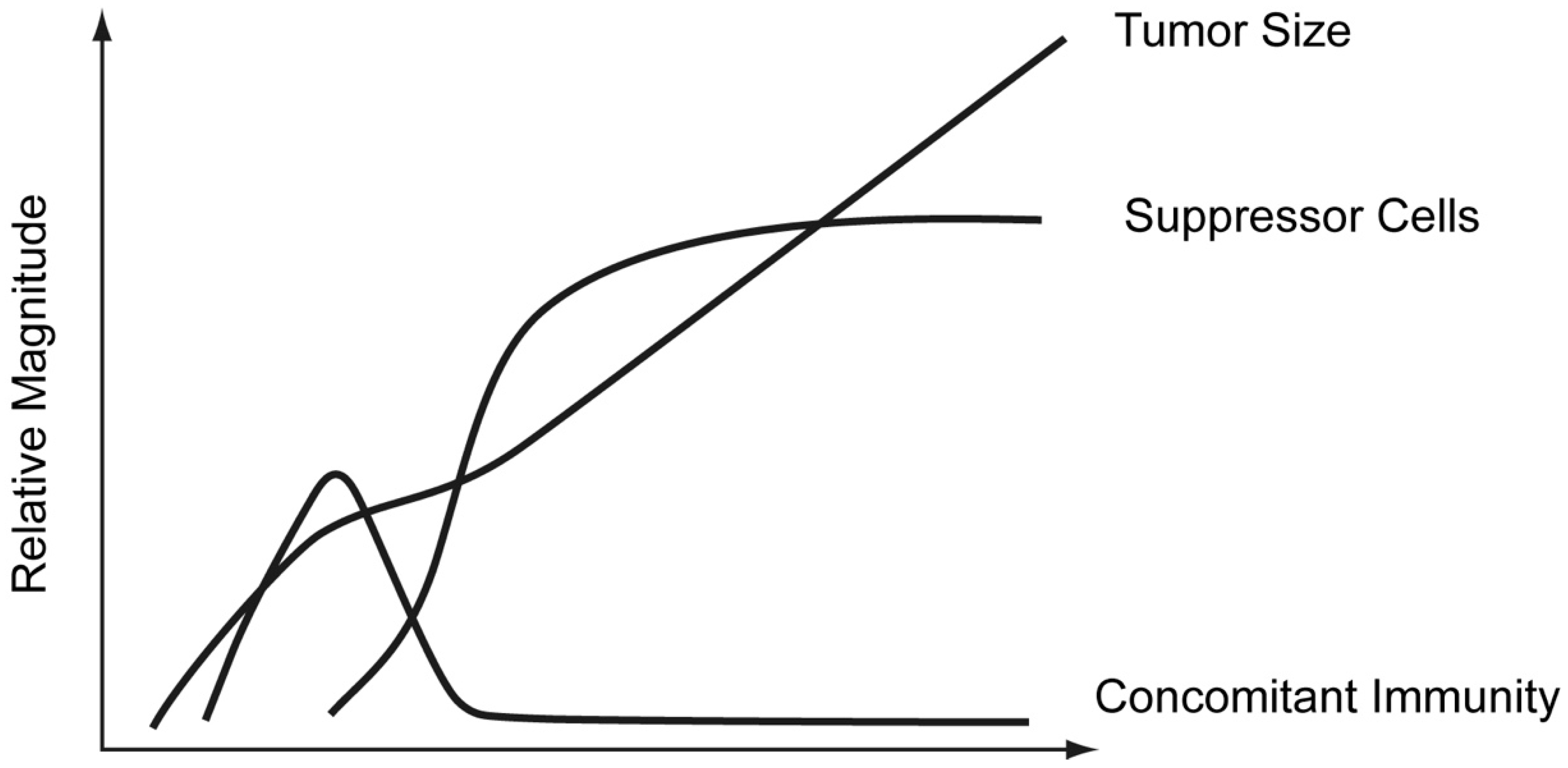
- We have briefly outlined “the CD4 T cell depletion experiment” above, showing that partial depletion modulates the response to decrease the Th2 and increase the Th1 component. Most of these observations were designed to test a critical prediction of the threshold mechanism [5,49]. However, such experiments also provide evidence against the PAMP/DAMP-centric view. No proponents of this view have explained how the number of CD4 T cells can affect the nature of the PAMP or DAMP signal. Thus, these observations present problems for the PAMP/DAMP-centric view. Again, these observations support the threshold mechanism.
7. Consequences of the Popularity of the PAMP/DAMP-Centric View
8. The View of Tumor Immunology Prevalent in the Late 1960s and Early 1970s
9. Robert North’s Studies
10. Our Studies in North’s Systems Leading to the Th2-Skewing Hypothesis of Tumor Escape
11. The Experimental Definition of Nt, the Transition Number, and the Prospect of Universally Efficacious Vaccination against Entities Preferentially Susceptible to Cell Mediated Attack
12. The Implications of the Threshold Mechanism for Health
- (a)
- The basis of the treatment of human visceral leishmaniasis
- (b)
- Treatment of cutaneous leishmaniasis in the mouse model of the human disease
- (c)
- Personalized immunotherapy of HIV-1 infections
- (d)
- Immune correlates of protection and disease upon infection by Mycobacterium tuberculosis: implications for the basis of failure to control the pathogen
- (e)
- Treatment of TB
13. Prevention and Treatment of Human Cancer
- (a)
- Prologue
- (b)
- Current treatment of human cancer
- (c)
- Modern developments pertinent to cancer immunotherapy
- (d)
- Centrality of monitoring the nature of the anti-cancer immune response
- (e)
- Improving current treatment
- (f)
- Epilogue: The international panel on immunotherapy of HIV-1 infections
14. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, D.; Barabasi, A. The Science of Science; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Chu, J.S.G.; Evans, J.A. Slowed canonical progress in large fields of science. Proc. Natl. Acad. Sci. USA 2021, 118, e2021636118. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Leahey, E.; Funk, R.J. Papers and patents are becoming less disruptive with time. Nature 2023, 613, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Bloom, N.; Jones, C.I.; Van Reenen, J.; Webb, N. Are ideas getting harder to find? Am. Econ. Rev. 2020, 110, 1104–1144. [Google Scholar] [CrossRef]
- Bretscher, P.A. Analyzing some concepts of immune regulation of the last three decades: Fostering greater research resilience despite the information overload. A personal view. Front. Immunol. 2022, 13, 960742. [Google Scholar] [CrossRef] [PubMed]
- Dresser, D.W.; Mitchison, N.A. The mechanism of immunological paralysis. Adv. Immunol. 1968, 8, 129–181. [Google Scholar] [PubMed]
- Kappler, J.W.; Roehm, N.; Marrack, P. T cell tolerance by clonal elimination in the thymus. Cell 1987, 49, 273–280. [Google Scholar] [CrossRef]
- Lederberg, J. Genes and Antibodies: Do antigens bear instructions for antibody specificity ordo they select cell lines that arise by mutation? Science 1959, 129, 1649–1653. [Google Scholar] [CrossRef]
- Chentoufi, A.A.; Polychronakos, C. Insulin expression levels in the thymus modulate insulin-specific autoreactive T-cell tolerance: The mechanism by which the IDDM2 locus may predispose to diabetes. Diabetes 2002, 51, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, P.; Cohn, M. A Theory of Self-Nonself Discrimination. Science 1970, 169, 1042–1049. [Google Scholar] [CrossRef]
- Burnet, F.M.; Fenner, F. The Production of Antibodies, 2nd ed.; Macmillan: Melbourne, Australia; London, UK, 1949. [Google Scholar]
- Bretscher, P.A. The historical postulate: Is it the basis, at the level of the system, for self-nonself discrimination? Scand. J. Immunol. 2021, 94, e13033. [Google Scholar] [CrossRef]
- Mitchell, G.F.; Miller, J.F.A.P. Cell to cell interaction in the immune response. II. The source of hemolysin-forming cells in irradiated mice given bone marrow and thymus or thoracic duct lymphocytes. J. Exp. Med. 1968, 128, 821–837. [Google Scholar] [CrossRef]
- Claman, H.N.; Chaperon, E.A. Immunologic Complementation between Thymus and Marrow Cells—A Model for the Two-Cell Theory of Immunocompetence. Immunol. Rev. 1969, 1, 92–113. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, N.A. The carrier effect in the secondary response to hapten-protein conjugates. II. Cellular cooperation. Eur. J. Immunol. 1971, 1, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Raff, M. Theta isoantigen as a marker of thymus-derived lymphocytes in mice. Nature 1969, 224, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Keene, J.A.; Forman, J. Helper activity is required for the in vivo generation of cytotoxic T lymphocytes. J. Exp. Med. 1982, 155, 768–782. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, E.S.; Klinman, N.R. In vitro tolerance induction of neonatal murine B cells. J. Exp. Med. 1976, 143, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Goodnow, C.C.; Crosbie, J.; Jorgensen, H.; Brink, R.A.; Basten, A. Induction of self-tolerance in mature peripheral B lymphocytes. Nature 1989, 342, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Guerder, S.; Matzinger, P. A fail-safe mechanism for maintaining self-tolerance. J. Exp. Med. 1992, 176, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Doherty, P.C.; Zinkernagel, R.M. H-2 compatibility is required for T-cell-mediated lysis of target cells infected with lymphocytic choriomeningitis virus. J. Exp. Med. 1975, 141, 502–507. [Google Scholar] [CrossRef]
- Buus, A.; Sette, S.; Colon, M.; Miles, C.; Grey, H.M. The relation between major histocompatibility complex (MHC) restriction and the capacity of Ia to bind immunogenic peptides. Science 1987, 235, 1353–1358. [Google Scholar] [CrossRef]
- Adorini, L.; Sette, A.; Buus, S.; Grey, H.M.; Darsley, M.; Lehmann, P.V.; Doria, G.; Nagy, Z.A.; Appella, E. Interaction of an immunodominant epitope with Ia molecules in T-cell activation. Proc. Natl. Acad. Sci. USA 1988, 85, 5181–5185. [Google Scholar] [CrossRef]
- Shimonkevitz, R.; Kappler, J.; Marrack, P.; Grey, H. Antigen recognition by H-2-restricted T cells. I. Cell-free antigen processing. J. Exp. Med. 1983, 158, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.J.; Warren, H.S.; Woolnough, J.A.; Talmage, D.W. Immunological induction of T lymphocytes: Role of antigen and the lymphocyte costimulator. Blood Cells 1978, 4, 395–406. [Google Scholar] [PubMed]
- Lafferty, K.J.; Cunningham, A.J. A new analysis of allogeneic interactions. Aust. J. Exp. Biol. Med. Sci. 1975, 53, 27–42. [Google Scholar] [CrossRef]
- Mueller, D.L.; Jenkins, M.J.; Schwartz, R.H. Clonal Expansion Versus Functional Clonal Inactivation: A Costimulatory Signalling Pathway Determines the Outcome of T Cell Antigen Receptor Occupancy. Ann. Rev. Immunol. 1989, 7, 445–480. [Google Scholar] [CrossRef]
- Fife, B.T.; Bluestone, J.A. Control of peripheral T-cell tolerance and autoimmunity via the CTLA-4 and PD-1 pathways. Immunol. Rev. 2008, 224, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54 Pt 1, 1–13. [Google Scholar] [CrossRef]
- Janeway, C.A. The immune system evolved to discriminate infectious nonself from noninfectious self. Immunol. Today 1992, 13, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Medzhitov, R. Innate immune recognition. Ann. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Matzinger, P. Tolerance, danger, and the extended family. Ann. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Matzinger, P.; Kamala, T. Tissue-based class control: The other side of tolerance. Nat. Rev. Immunol. 2011, 11, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Matzinger, P. Friendly and dangerous signals: Is the tissue in control? Nat. Immunol. 2007, 8, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.M.; Hall, J.A.; Blank, R.B.; Bouladoux, N.; Oukka, M.; Mora, J.R.; Belkaid, Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 2007, 204, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef]
- Fearon, D.T.; Locksley, R.M. The instructive role of innate immunity in the acquired immune response. Science 1996, 272, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, P.A. On the control between cell-mediated, IgM and IgG immunity. Cell. Immunol. 1974, 13, 171–195. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Raffel, S. Macrophage-digested antigen as inducer of delayed hypersensitivity. J. Exp. Med. 1971, 133, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Salvin, S.B. Occurrence of delayed hypersensitivity during the development of Arthus type hypersensitivity. J. Exp. Med. 1958, 107, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, P. On analyzing how the Th1/Th2 phenotype of an immune response is determined: Classical observations must not be ignored. Front. Immunol. 2019, 10, 1234. [Google Scholar] [CrossRef]
- Asherson, G.L.; Stone, S.H. Selective and specific inhibition of 24 hour skin reactions in the guinea-pig: I. Immune deviation: Description of the phenomenon and the effect of splenectomy. Immunology 1965, 9, 205–217. [Google Scholar] [PubMed]
- Parish, C.R. The relationship between humoral and cell-mediated immunity. Transplant. Rev. 1972, 13, 35–66. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.R. Immune response to chemically modified flagellin. II. Evidence for a fundamental relationship between humoral and cell-mediated immunity. J. Exp. Med. 1971, 134, 21–47. [Google Scholar] [CrossRef] [PubMed]
- Britton, W.J.; Lockwood, D.N. Leprosy. Lancet 2004, 363, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Sher, A.; Gazzinelli, R.T.; Oswald, I.P.; Clerici, M.; Kullberg, M.; Pearce, E.J.; Berzovsky, J.A.; Mosmann, T.R.; James, S.L.; Morse, H.C. T cell derived cytokines in the downregulation of immune responses in parasitic and retroviral infection. Immunol. Rev. 1992, 127, 183–204. [Google Scholar] [CrossRef]
- Lagrange, P.H.; Mackaness, G.B.; Miller, T.E. Influence of dose and route of antigen injection on the immunological induction of T cells. J. Exp. Med. 1974, 139, 528–542. [Google Scholar] [CrossRef]
- Bretscher, P.A. A two-step, two-signal model for the primary activation of precursor helper T cells. Proc. Natl. Acad. Sci. USA 1992, 96, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, P.A. On the mechanism determining the TH1/TH2 phenotype of an immune response, and its pertinence to strategies for the prevention, and treatment, of certain infectious diseases. Scand. J. Immunol. 2014, 79, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Titus, R.G.; Ceredig, R.; Cerottini, J.C.; Louis, J.A. Therapeutic effect of anti-L3T4 monoclonal antibody GK1.5 on cutaneous leishmaniasis in genetically-susceptible BALB/c mice. J. Immunol. 1985, 135, 2108–2114. [Google Scholar] [CrossRef] [PubMed]
- Sadick, M.D.; Heinzel, F.P.; Shigekane, V.M.; Fisher, W.L.; Locksley, R.M. Cellular and humoral immunity to Leishmania major in genetically susceptible mice after in vivo depletion of L3T4+ T cells. J. Immunol. 1987, 139, 1303–1309. [Google Scholar] [CrossRef]
- Bretscher, P.A.; Wei, G.; Menon, J.N.; Bielefeldt-Ohmann, H. Establishment of stable, cell-mediated immunity that makes “susceptible” mice resistant to Leishmania major. Science 1992, 257, 539–542. [Google Scholar] [CrossRef]
- Menon, J.N.; Bretscher, P.A. Parasite dose determines the Th1/Th2 nature of the respons to Leishmania major independently of infection route and strain of host or parasite. Eur. J. Immunol. 1998, 28, 4020–4028. [Google Scholar] [CrossRef]
- Buddle, B.M.; de Lisle, G.W.; Pfeffer, A.; Aldwell, F.E. Immunological responses and protection against Mycobacterium bovis in calves vaccinated with a low dose of BCG. Vaccine 1995, 13, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The three Es of cancer immunoediting. Annu. Rev. Immunol. 2004, 22, 329–360. [Google Scholar] [CrossRef]
- Klein, G. Tumor-specific transplantation antigens: G. H. A. Clowes memorial lecture. Cancer Res. 1968, 28, 625–635. [Google Scholar] [PubMed]
- Foley, E.J. Antigenic properties of methylcholanthrene-induced tumors in mice of the strain of origin. Cancer Res. 1953, 13, 835–837. [Google Scholar] [PubMed]
- Gorelik, E. Concomitant tumor immunity and the resistance to a second tumor challenge. Adv. Cancer Res. 1983, 39, 71–120. [Google Scholar] [PubMed]
- Berendt, M.J.; North, R.J. T-cell-mediated suppression of anti-tumor immunity. An explanation for progressive growth of an immunogenic tumor. J. Exp. Med. 1980, 151, 69–80. [Google Scholar] [CrossRef]
- North, R.J.; Bursuker, I. Generation and decay of the immune response to a progressive fibrosarcoma. I. Ly-1+2- suppressor T cells down-regulate the generation of Ly-1-2+ effector T cells. J. Exp. Med. 1984, 159, 1295–1311. [Google Scholar] [CrossRef] [PubMed]
- North, R.J. The murine antitumor immune response and its therapeutic manipulation. Adv. Immunol. 1984, 35, 89–155. [Google Scholar] [CrossRef] [PubMed]
- Dye, E.S.; North, R.J. Specificity of the T cells that mediate and suppress adoptive immunotherapy of established tumors. J. Leukoc. Biol. 1984, 36, 27–37. [Google Scholar] [CrossRef]
- Hamilton, D.H.; Bretscher, P.A. Different immune correlates associated with tumor progression and regression: Implications for prevention and treatment of cancer. Cancer Immunol. Immunother. 2008, 57, 1125–1136. [Google Scholar] [CrossRef] [PubMed]
- Okulicz, J.F.; Marconi, V.C.; Landrum, M.L.; Weger, S.; Weintrob, A.; Ganesam, A.; Hale, B.; Crum-Cianflone, N.; Delmar, J.; Barthel, V.; et al. Clinical outcomes of elite controllers, viremic controllers, and long-term non-progressors in the US Department of Defense HIV natural history study. J. Infect. Dis. 2009, 200, 1714–1723. [Google Scholar] [CrossRef] [PubMed]
- Keele, B.F.; Giorgi, K.E.; Salazar-Gonzalez, J.F.; Dercker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, P.A.; Al-Yassin, G. Can interruption/withdrawl of anti-retroviral therapy provide personalized immunotherapy against HIV-1? Scand. J. Immunol. 2020, 92, e12934. [Google Scholar] [CrossRef] [PubMed]
- Hailu, A.; Menon, J.N.; Behre, N.; Gedamu, L.; Hassard, T.H.; Kager, P.A.; Oloba, J.; Bretscher, P.A. Distinct immunity in patients with visceral leishmaniasis from that in subclinically infected and drug-cured people: Implications for the mechanism underlying drug cure. J. Infect. Dis. 2001, 184, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Altfeld, M.; Walker, B.D. Less is more? STI in acute and chronic HIV-1 infection. Nat. Med. 2001, 7, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Giang-Huong, N.; Candotti, D.; Goubar, A.; Autran, B.; Maynart, M.; Sicard, D.; Clauvel, J.P.; Agut, H.; Costagliola, D.; Rouzioux, C.; et al. HIV type 1-specific IgG2 antibodies: Markers of helper T cell type 1 response and prognostic marker of long-term non-progression. AIDS Res. Hum. Retrovir. 2001, 17, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Costagliola, D.; Bonduelle, O.; N’go, N.; Schnuriger, A.; Theodorou, I.; Clauvel, J.-P.; Sicard, D.; Agut, H.; Debre, P.; et al. Combination of HIV-1-specific CD4 Th1 cell responses and IgG2 antibodies is the best predictor for persistence of long-term non progression. J. Infect. Dis. 2005, 191, 2053–2063. [Google Scholar] [CrossRef]
- Capa, L.; Ayala-Suárez, R.; De La Torre Tarazona, H.E.; González-García, J.; Del Romero, J.; Alcamí, J.; Díez-Fuertes, F. Capa, Elite controllers long-term non progressors present improved survival and slower disease progression. Sci. Rep. 2022, 12, 16356. [Google Scholar] [CrossRef]
- Saez-Cirion, A.; Bacchus, C.; Hocqueloux, L.; Avettand-Fenoel, V.; Girault, I.; Lecuroux, C.; Potard, V.; Vermisse, P.; Melard, A.; Prazuck, T.; et al. Post-treatment HIV-1 controllers with a long-term virological remission after the interruption of early initiated antiretroviral therapy ANRS VISCONTI Study. PLoS Pathog. 2013, 9, e1003211. [Google Scholar] [CrossRef]
- Wong, J.K.; Hezareh, M.; Gunthard, H.F.; Havlir, D.V.; Ignacio, C.C.; Spina, C.A.; Richman, D.D. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 1997, 278, 1291–1295. [Google Scholar] [CrossRef]
- Menon, J.; Hoeppner, V.H.; Judd, A.; Power, C.A.; Bretscher, P.A. A hypothesis for the existence of two types of tuberculosis, reflecting two distinct types of immune failure to control the pathogen, based upon prevalence of mycobacterium-specific IgG subclasses. Scand. J. Immunol. 2018, 87, e12665. [Google Scholar] [CrossRef]
- Dannenberg, A.M. Pathogenesis of pulmonary Mycobacterium bovis infection: Basic principles established by the rabbit model. Tuberculosis 2001, 81, 87–96. [Google Scholar] [CrossRef]
- Nunes-Alves, C.; Booty, M.G.; Carpenter, S.M.; Jayaraman, P.; Rothchild, A.C.; Behar, S.M. In search of a new paradigm for protective immunity to TB. Nat. Rev. Microbiol. 2014, 12, 289–299. [Google Scholar] [CrossRef]
- Millington, K.A.; Gooding, S.; Hinks, T.S.; Reynolds, D.J.; Lalvani, A. Mycobacterium tuberculosis-specific cellular immune profiles suggest bacillary persistence decades after spontaneous cure in untreated tuberculosis. J. Infect. Dis. 2010, 202, 1685–1689. [Google Scholar] [CrossRef]
- Koch, R. Professor Koch’s remedy for tuberculosis. A further communication on a remedy for tuberculosis: By professor. Indian J. Med. Res. 2023, 157, 169–173. [Google Scholar] [CrossRef]
- Koch, R. A Further Communication on a Remedy for Tuberculosis. Br. Med. J. 1891, 1, 125–127. [Google Scholar] [CrossRef]
- Coghill, J.G.S. Sequel of a case treated by Koch’s tuberculin, with results of the necroscopy. Lancet 1895, 146, 1219–1221. [Google Scholar] [CrossRef]
- Bretscher, P.A. Facing the Increased Prevalence of Antibiotic-Resistant M. tuberculosis: Exploring the Feasibility of Realizing Koch’s Aspiration of Immunotherapy of Tuberculosis. Antibiotics 2022, 11, 371. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Boon, T.; Coulie, P.G.; Van den Eynde, B.J.; van der Bruggen, P. Human T cell responses against melanoma. Ann. Rev. Immunol. 2006, 24, 175–208. [Google Scholar] [CrossRef]
- Boon, T.; Cerottini, J.-C.; van den Eynde, B.; van der Bruggen, P.; Van Pel, A. Tumor antigens recognized by T lymphocytes. Ann. Rev. Immunol. 1994, 12, 337–365. [Google Scholar] [CrossRef]
- Sahin, U.; Tureci, O.; Schmitt, H.; Cochlovius, B.; Johannes, T.; Schmits, R.; Stenner, F.; Luo, G.; Schobert, I.; Pfreundschuh, M. Human neoplasms elicit multiple specific immune responses in the autologous host. Proc. Natl. Acad. Sci. USA 1995, 92, 11810–11813. [Google Scholar] [CrossRef]
- Sterner, R.C.; Sterner, R.M. CAR-T cell therapy: Current limitations and potential strategies. Blood Cancer J. 2021, 11, 69. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Postow, M.A.; Sidlow, R.; Mellmann, M.D. Immune-Related Adverse Events Associated with Immune Checkpoint Blockade. N. Eng. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef]
- Bretscher, P. The role of cytokines in determining the Th1/Th2 phenotype of an immune response: Coherence of the T cell response and the Cytokine Implementation Hypothesis. Scand. J. Immunol. 2022, 95, e13110. [Google Scholar] [CrossRef]
- Nishimura, T.; Iwakabe, K.; Sekimoto, M.; Ohmi, Y.; Yahata, T.; Nakui, M.; Sato, T.; Habu, S.; Tashiro, H.; Sato, M.; et al. Distinct role of antigen-specific T helper type 1 (Th1) and Th2 cells in tumor eradication in vivo. J. Exp. Med. 1999, 190, 617–627. [Google Scholar] [CrossRef]
- Tatsumi, T.; Kierstead, L.S.; Ranieri, E.; Gesualdo, L.; Schena, F.P.; Finke, J.H.; Bukowski, R.M.; Mueller-Berghaus, J.; Kirkwood, J.M.; Kwok, W.W.; et al. Disease-Associated Bias in T Helper Type 1 (Th1)/Th2 CD4+ T Cell Responses Against MAGE-6 in HLA-DRB10401+ Patients with Renal Cell Carcinoma or Melanoma. J. Exp. Med. 2002, 196, 619–628. [Google Scholar] [CrossRef]
- Tassi, E.; Gavazzi, F.; Albarello, L.; Senyukov, V.; Longhi, R.; Dellabona, P.; Doglioni, C.; Braga, M.; Di Carlo, V.; Protti, M.P. Carcinoembryonic Antigen-Specific But Not Antiviral CD4+ T Cell Immunity Is Impaired in Pancreatic Carcinoma Patients. J. Immunol. 2008, 181, 6595–6603. [Google Scholar] [CrossRef]
- Guenova, E.; Watanabe, R.; Teague, J.E.; Desimone, J.A.; Jiang, Y.; Dowlatshahi, M.; Schlapbach, C.; Schaekel, K.; Rook, A.H.; Tawa, M.; et al. TH2 Cytokines From Malignant Cells Suppress TH1 Responses and Enforce a Global TH2 Bias in Leukemic Cutaneous T-Cell Lymphoma. Clin. Cancer Res. 2013, 19, 3755–3763. [Google Scholar] [CrossRef] [PubMed]
- Frafjord, A.; Buer, L.; Hammarström, c.; Aamodt, H.; Woldbæk, P.R.; Brustugun, O.T.; Helland, A.; Øynebråten, I.; Corthay, A. The immune landscape, of human primary lung tumors is Th2 skewed. Front. Immunol. 2021, 12, 764596. [Google Scholar] [CrossRef] [PubMed]
- Lagrange, P.H.; Mackaness, G.B.; Miller, T.E. Potentiation of T-cell-mediated immunity by selective suppression of antibody formation by cyclophosphamide. J. Exp. Med. 1974, 139, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Awwad, M.; North, R.J. Cyclophosphamide-induced Immunologically Mediated Regression of a Cyclophosphamide-resistant Murine Tumor: A Consequence of Eliminating Precursor L3T4+ Suppressor T-Cells. Cancer Res. 1989, 4, 1649–1654. [Google Scholar] [PubMed]
- North, R.J.; Awwad, M. Elimination of cycling CD4+ suppressor T cells with an anti-mitotic drug releases non-cycling CD8+ T cells to cause regression of an advanced lymphoma. Immunology 1990, 71, 90–95. [Google Scholar] [PubMed]
- Deeks, S.G.; Lewin, S.R.; Ross, A.L.; Ananworanich, J.; Benkirane, M.; Cannon, P.; Chomont, N.; Douek, D.; Lifson, J.D.; Lo, Y.-R.; et al. International AIDS Society global scientific strategy: Towards an HIV cure. Nat. Med. 2016, 22, 839–850. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bretscher, P.A. A Plausible Framework Reveals Potential Similarities in the Regulation of Immunity against Some Cancers and Some Infectious Agents: Implications for Prevention and Treatment. Cancers 2024, 16, 1431. https://doi.org/10.3390/cancers16071431
Bretscher PA. A Plausible Framework Reveals Potential Similarities in the Regulation of Immunity against Some Cancers and Some Infectious Agents: Implications for Prevention and Treatment. Cancers. 2024; 16(7):1431. https://doi.org/10.3390/cancers16071431
Chicago/Turabian StyleBretscher, Peter A. 2024. "A Plausible Framework Reveals Potential Similarities in the Regulation of Immunity against Some Cancers and Some Infectious Agents: Implications for Prevention and Treatment" Cancers 16, no. 7: 1431. https://doi.org/10.3390/cancers16071431