Proteomic Analysis of Spatial Heterogeneity Identifies HMGB2 as Putative Biomarker of Tumor Progression in Adult-Type Diffuse Astrocytomas

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Discovery Cohort
2.1.1. Protein Isolation and Mass Spectrometry
2.1.2. Pathway Analysis
2.2. Validation Cohorts
2.2.1. Molecular Characterization (Validation Cohort 1)
- Genomic and methylomic profiling:
- Transcriptomic profiling:
2.2.2. Histopathology and Immunohistochemistry (IHC) (Validation Cohort 2)
2.3. Biostatistical Analyses
3. Results
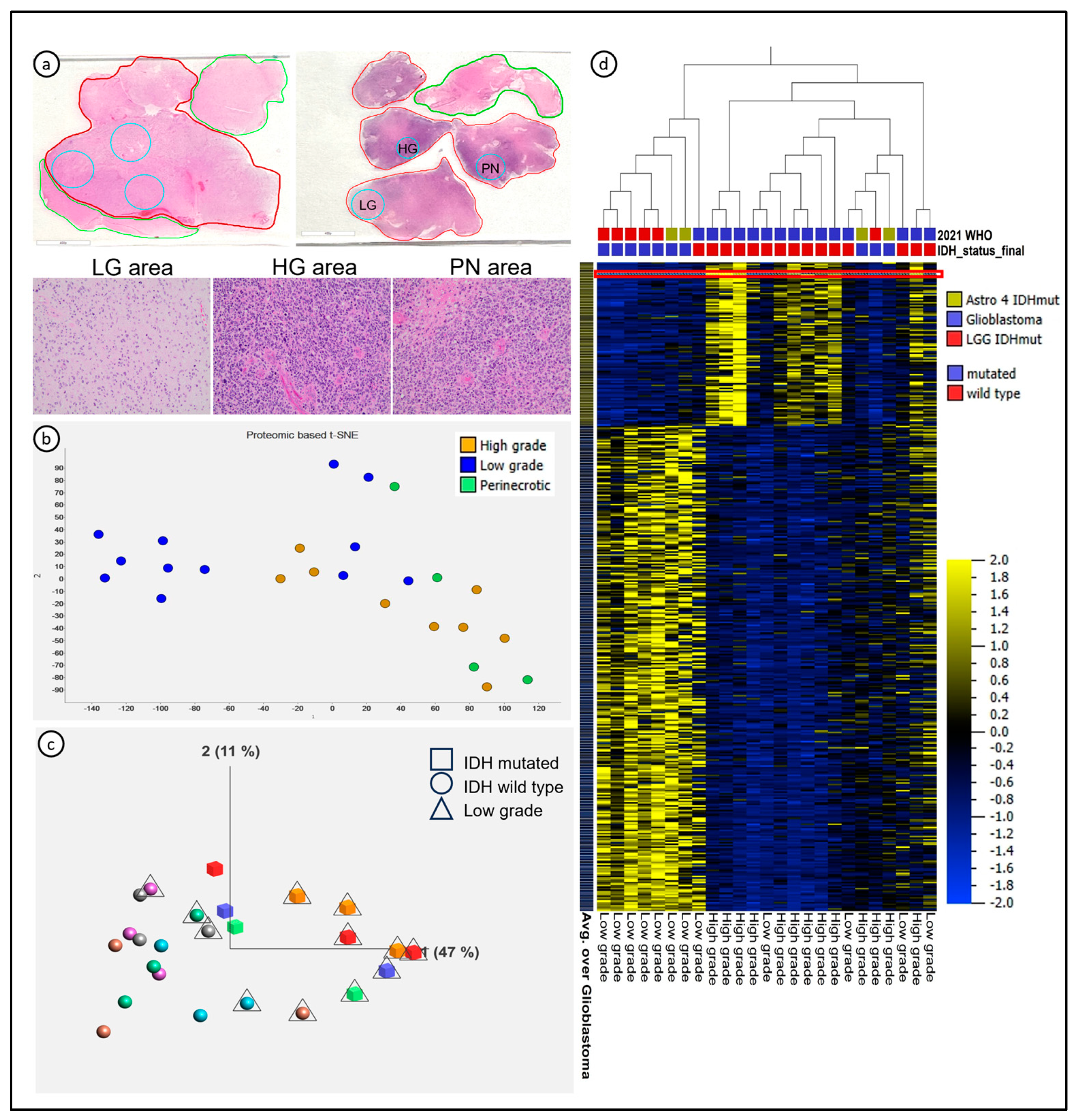
3.1. LC-MS/MS-Detected Proteomic Signatures Characterizing Glioma Evolution towards Higher Grades, Which Were Associated with, but Not Completely Dependent on, IDH Status

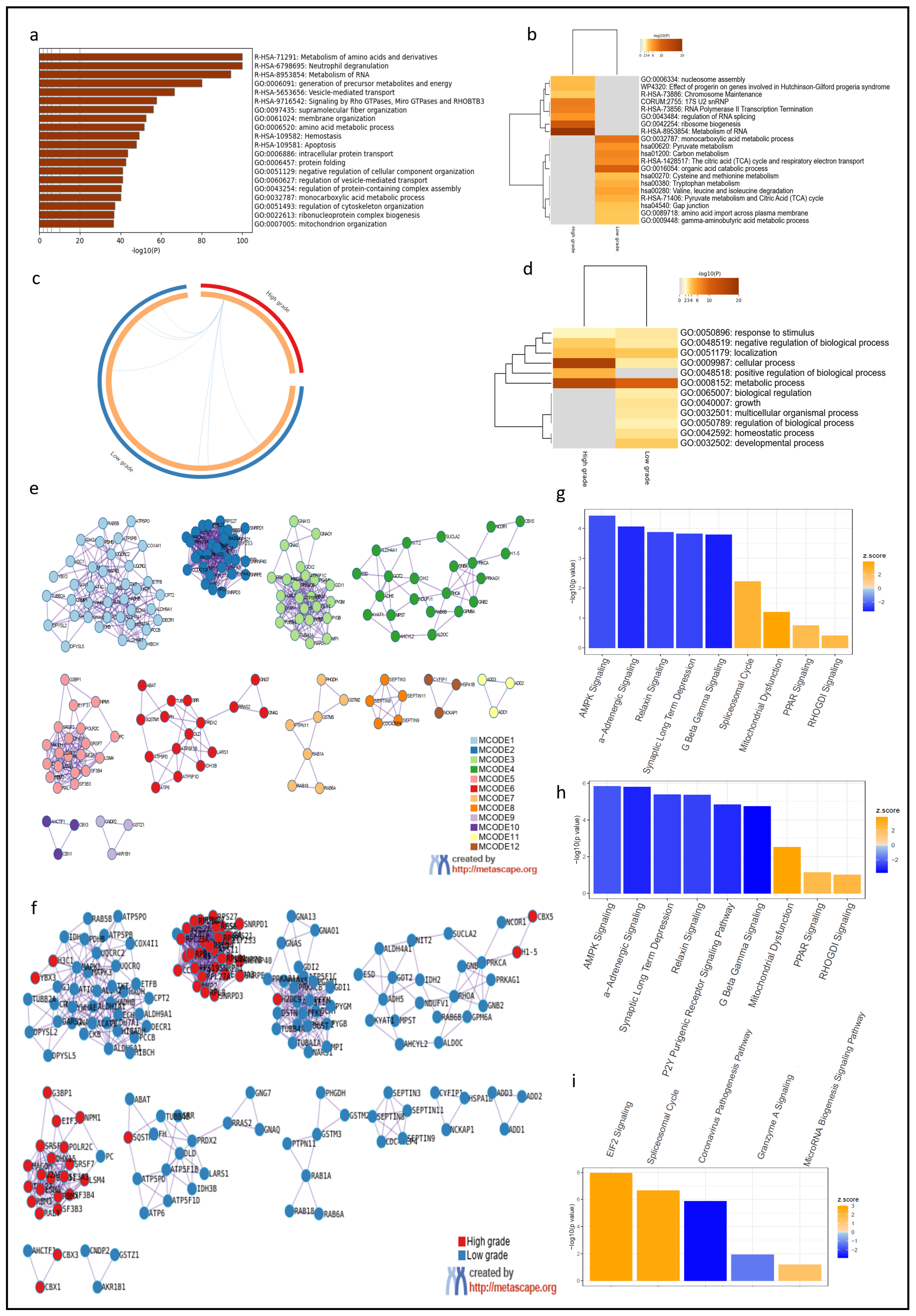
3.2. Spatial Heterogeneity of Diffuse Astrocytomas Was Associated with Dysregulation of Specific Molecular Pathways and HMGB2 Was Identified as a Putative Driver of Tumor Progression
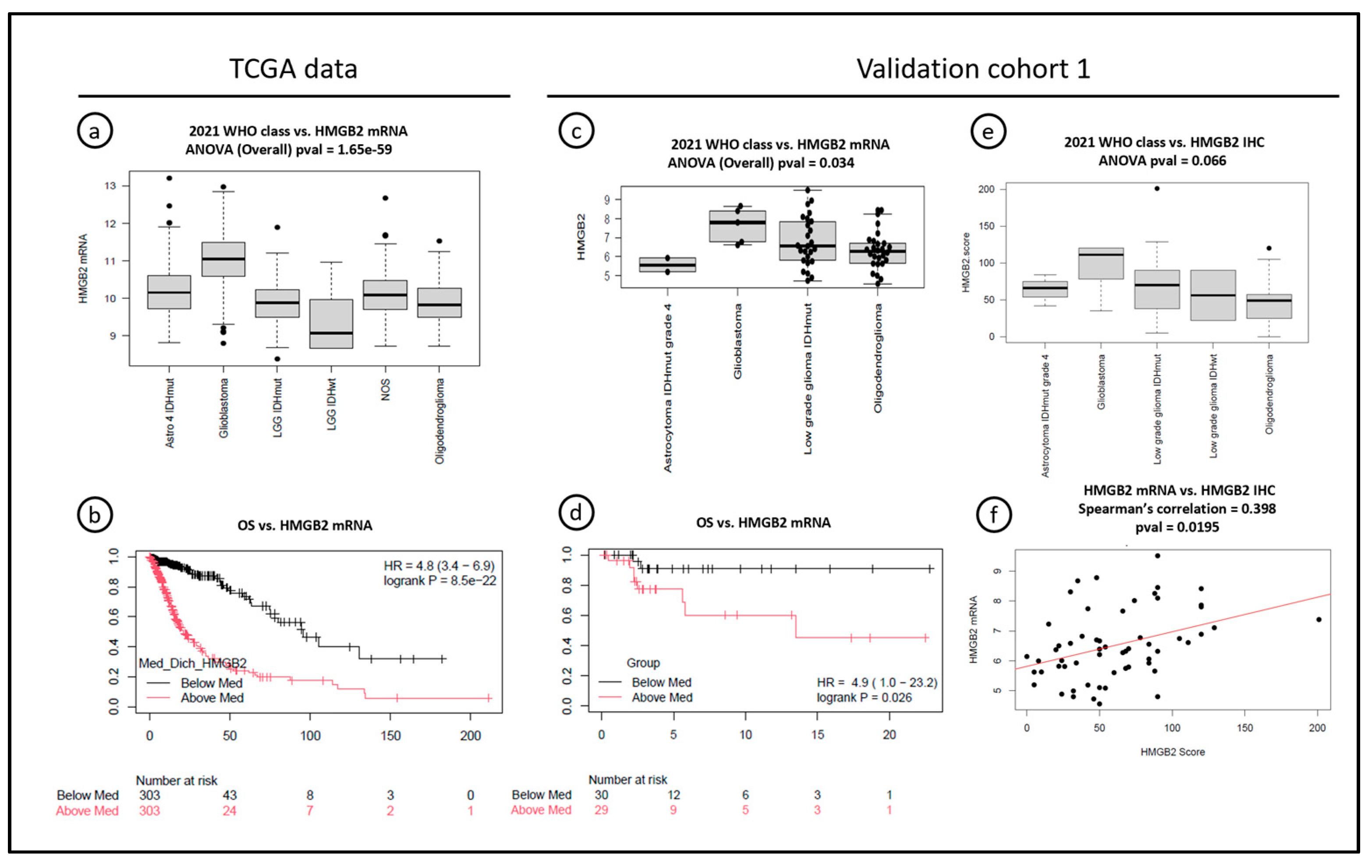
3.3. High HMGB2 Expression, Both at mRNA and Protein Levels, Is an Early Marker of Worse Overall Survival in Grades 2 and 3 Diffuse Gliomas, and It Is, at Least in Part, Regulated by DNA Methylation
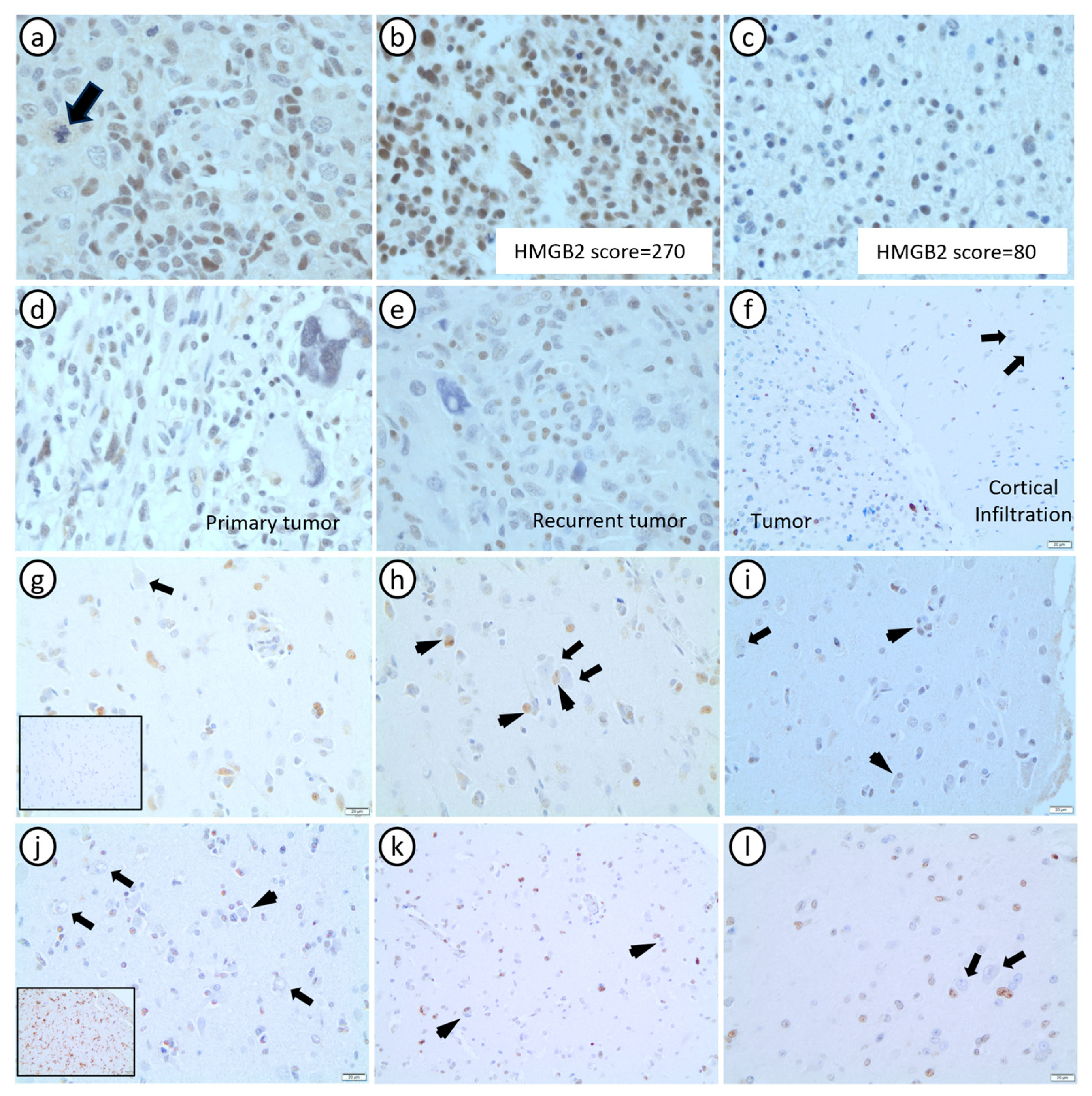
3.4. HMGB2 Is Significantly Associated with Histopathological Markers of Grade 4 Astrocytomas, and It Is Minimally Expressed in Non-Neoplastic Glial Cells
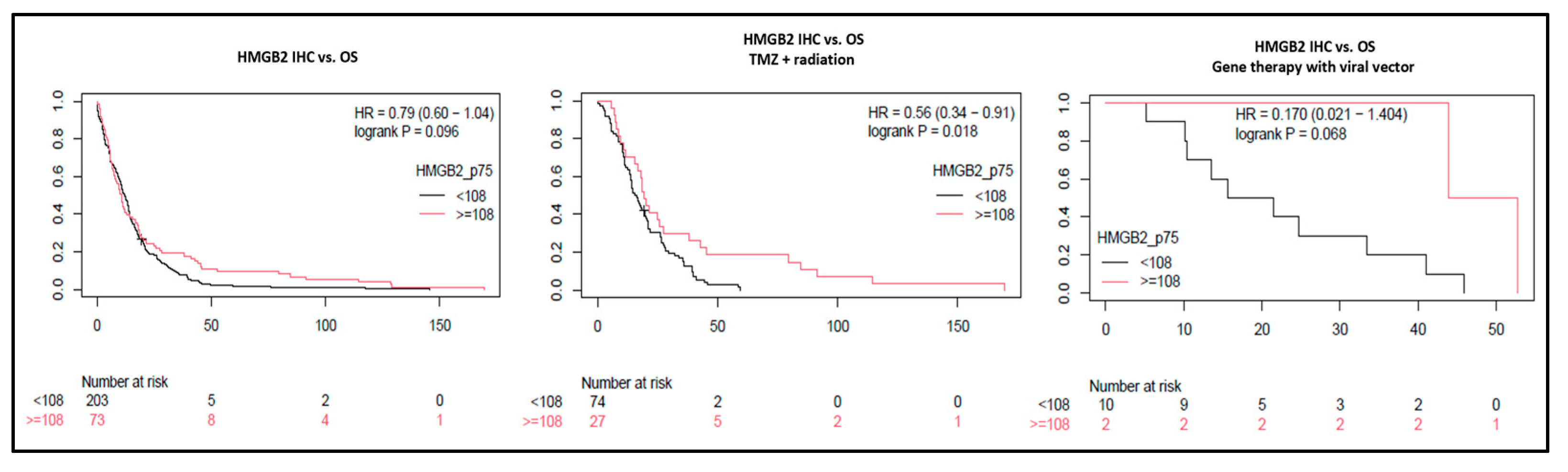
3.5. High HMGB2 Expression Identifies Glioblastomas with Better Response to Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Becker, A.P.; Sells, B.E.; Haque, S.J.; Chakravarti, A. Tumor Heterogeneity in Glioblastomas: From Light Microscopy to Molecular Pathology. Cancers 2021, 13, 761. [Google Scholar] [CrossRef] [PubMed]
- Daumas-Duport, C.; Scheithauer, B.; O’Fallon, J.; Kelly, P. Grading of astrocytomas. A simple and reproducible method. Cancer 1988, 62, 2152–2165. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.J.; Fuller, G.N.; Abi-Said, D.; Lang, F.F.; Gokaslan, Z.L.; Shi, W.M.; Wildrick, D.M.; Sawaya, R. Limitations of stereotactic biopsy in the initial management of gliomas. Neuro Oncol. 2001, 3, 193–200. [Google Scholar] [CrossRef]
- Paulus, W.; Peiffer, J. Intratumoral histologic heterogeneity of gliomas. A quantitative study. Cancer 1989, 64, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Zolotovskaia, M.A.; Kovalenko, M.A.; Tkachev, V.S.; Simonov, A.M.; Sorokin, M.I.; Kim, E.; Kuzmin, D.V.; Karademir-Yilmaz, B.; Buzdin, A.A. Next-Generation Grade and Survival Expression Biomarkers of Human Gliomas Based on Algorithmically Reconstructed Molecular Pathways. Int. J. Mol. Sci. 2022, 23, 7330. [Google Scholar] [CrossRef] [PubMed]
- Fowler, C.B.; O’Leary, T.J.; Mason, J.T. Toward Improving the Proteomic Analysis of Formalin-fixed, Paraffin-embedded Tissue. Expert Rev. Proteom. 2013, 10, 389–400. [Google Scholar] [CrossRef]
- Shi, S.R.; Liu, C.; Balgley, B.M.; Lee, C.; Taylor, C.R. Protein extraction from formalin-fixed, paraffin-embedded tissue sections: Quality evaluation by mass spectrometry. J. Histochem. Cytochem. 2006, 54, 739–743. [Google Scholar] [CrossRef]
- Neagu, A.N.; Jayathirtha, M.; Baxter, E.; Donnelly, M.; Petre, B.A.; Darie, C.C. Applications of Tandem Mass Spectrometry (MS/MS) in Protein Analysis for Biomedical Research. Molecules 2022, 27, 2411. [Google Scholar] [CrossRef]
- Beyer, S.J.; Bell, E.H.; McElroy, J.P.; Fleming, J.L.; Cui, T.; Becker, A.; Bassett, E.; Johnson, B.; Gulati, P.; Popp, I.; et al. Oncogenic transgelin-2 is differentially regulated in isocitrate dehydrogenase wild-type vs. mutant gliomas. Oncotarget 2018, 9, 37097–37111. [Google Scholar] [CrossRef]
- Bassett, E.A.; Palanichamy, K.; Pearson, M.; McElroy, J.P.; Haque, S.J.; Bell, E.H.; Chakravarti, A. Calpastatin phosphorylation regulates radiation-induced calpain activity in glioblastoma. Oncotarget 2018, 9, 14597–14607. [Google Scholar] [CrossRef]
- Becker, A.P.; Osorio, D.S.; Bell, E.H.; Giglio, P.; Fleming, J.L.; Cottrell, C.E.; Mardis, E.R.; Miller, K.E.; Schieffer, K.M.; Kelly, B.J.; et al. Correspondence comprehensive characterization of a brainstem aggregoma (light and heavy chain deposition disease). Brain Pathol. 2023, 33, e13154. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.; Becker, K.-F.; Schott, C.; Erpenbach, A.; Porschewsk, P. Protein Expression Analysis in Formalin-Fixed, Paraffin-Embedded (FFPE) Tissue; Qiagen Inc.: Venlo, The Netherlands; Available online: https://www.qiagen.com/us/resources/download.aspx?id=95754d31-8696-438f-a797-836aa9fa3d12&lang=en (accessed on 13 November 2023).
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.H.; Zhang, P.; Shaw, E.G.; Buckner, J.C.; Barger, G.R.; Bullard, D.E.; Mehta, M.P.; Gilbert, M.R.; Brown, P.D.; Stelzer, K.J.; et al. Comprehensive Genomic Analysis in NRG Oncology/RTOG 9802: A Phase III Trial of Radiation Versus Radiation Plus Procarbazine, Lomustine (CCNU), and Vincristine in High-Risk Low-Grade Glioma. J. Clin. Oncol. 2020, 38, 3407–3417. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Bell, E.H.; McElroy, J.; Becker, A.P.; Gulati, P.M.; Geurts, M.; Mladkova, N.; Gray, A.; Liu, K.; Yang, L.; et al. miR-4516 predicts poor prognosis and functions as a novel oncogene via targeting PTPN14 in human glioblastoma. Oncogene 2019, 38, 2923–2936. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhang, X.; Huang, C.C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, K.N.; Wang, Q.W.; Li, G.Z.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.C.; Wang, Z.; Zhang, C.B.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Puchalski, R.B.; Shah, N.; Miller, J.; Dalley, R.; Nomura, S.R.; Yoon, J.G.; Smith, K.A.; Lankerovich, M.; Bertagnolli, D.; Bickley, K.; et al. An anatomic transcriptional atlas of human glioblastoma. Science 2018, 360, 660–663. [Google Scholar] [CrossRef]
- Louis, D.N.; Wesseling, P.; Aldape, K.; Brat, D.J.; Capper, D.; Cree, I.A.; Eberhart, C.; Figarella-Branger, D.; Fouladi, M.; Fuller, G.N.; et al. cIMPACT-NOW update 6: New entity and diagnostic principle recommendations of the cIMPACT-Utrecht meeting on future CNS tumor classification and grading. Brain Pathol. 2020, 30, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Brat, D.J.; Aldape, K.; Bridge, J.A.; Canoll, P.; Colman, H.; Hameed, M.R.; Harris, B.T.; Hattab, E.M.; Huse, J.T.; Jenkins, R.B.; et al. Molecular Biomarker Testing for the Diagnosis of Diffuse Gliomas. Arch. Pathol. Lab. Med. 2022, 146, 547–574. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.; Ammar, A.; Riutta, A.; Waagmeester, A.; Slenter, D.N.; Hanspers, K.; Miller, A.R.; Digles, D.; Lopes, E.N.; Ehrhart, F.; et al. WikiPathways: Connecting communities. Nucleic Acids Res. 2021, 49, D613–D621. [Google Scholar] [CrossRef] [PubMed]
- Pico, A.R.; Kelder, T.; van Iersel, M.P.; Hanspers, K.; Conklin, B.R.; Evelo, C. WikiPathways: Pathway editing for the people. PLoS Biol. 2008, 6, e184. [Google Scholar] [CrossRef] [PubMed]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.J.; Kim, M.S.; Kim, M.S.; Lee, J.; Kang, M.; Jeong, J.H. High-mobility group box 2 (HMGB2) modulates radioresponse and is downregulated by p53 in colorectal cancer cell. Cancer Biol. Ther. 2013, 14, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.B.; Cai, L.; Lin, S.J.; Xiong, Z.K.; Lu, J.L.; Mao, Y.; Yao, Y.; Zhou, L.F. High-mobility group box 2 is associated with prognosis of glioblastoma by promoting cell viability, invasion, and chemotherapeutic resistance. Neuro Oncol. 2013, 15, 1264–1275. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Barthel, F.P.; Malta, T.M.; Sabedot, T.S.; Salama, S.R.; Murray, B.A.; Morozova, O.; Newton, Y.; Radenbaugh, A.; Pagnotta, S.M.; et al. Molecular Profiling Reveals Biologically Discrete Subsets and Pathways of Progression in Diffuse Glioma. Cell 2016, 164, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Sondka, Z.; Dhir, N.B.; Carvalho-Silva, D.; Jupe, S.; Madhumita; McLaren, K.; Starkey, M.; Ward, S.; Wilding, J.; Ahmed, M.; et al. COSMIC: A curated database of somatic variants and clinical data for cancer. Nucleic Acids Res. 2023, 52, D1210–D1217. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Zhang, Y.; Lin, Z.; Peng, L. Roles of HMGBs in Prognosis and Immunotherapy: A Pan-Cancer Analysis. Front. Genet. 2021, 12, 764245. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, L.A.; Manzanera, A.G.; Bell, S.D.; Cavaliere, R.; McGregor, J.M.; Grecula, J.C.; Newton, H.B.; Lo, S.S.; Badie, B.; Portnow, J.; et al. Phase II multicenter study of gene-mediated cytotoxic immunotherapy as adjuvant to surgical resection for newly diagnosed malignant glioma. Neuro Oncol. 2016, 18, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Korber, V.; Yang, J.; Barah, P.; Wu, Y.; Stichel, D.; Gu, Z.; Fletcher, M.N.C.; Jones, D.; Hentschel, B.; Lamszus, K.; et al. Evolutionary Trajectories of IDH(WT) Glioblastomas Reveal a Common Path of Early Tumorigenesis Instigated Years ahead of Initial Diagnosis. Cancer Cell 2019, 35, 692–704.e12. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Liu, Y.; Cai, S.J.; Qian, M.; Ding, J.; Larion, M.; Gilbert, M.R.; Yang, C. IDH mutation in glioma: Molecular mechanisms and potential therapeutic targets. Br. J. Cancer 2020, 122, 1580–1589. [Google Scholar] [CrossRef]
- Starkova, T.; Polyanichko, A.; Tomilin, A.N.; Chikhirzhina, E. Structure and Functions of HMGB2 Protein. Int. J. Mol. Sci. 2023, 24, 8334. [Google Scholar] [CrossRef]
- Reeves, R. High mobility group (HMG) proteins: Modulators of chromatin structure and DNA repair in mammalian cells. DNA Repair 2015, 36, 122–136. [Google Scholar] [CrossRef]
- Niu, L.; Yang, W.; Duan, L.; Wang, X.; Li, Y.; Xu, C.; Liu, C.; Zhang, Y.; Zhou, W.; Liu, J.; et al. Biological functions and theranostic potential of HMGB family members in human cancers. Ther. Adv. Med. Oncol. 2020, 12, 1758835920970850. [Google Scholar] [CrossRef] [PubMed]
- Stros, M.; Ozaki, T.; Bacikova, A.; Kageyama, H.; Nakagawara, A. HMGB1 and HMGB2 cell-specifically down-regulate the p53- and p73-dependent sequence-specific transactivation from the human Bax gene promoter. J. Biol. Chem. 2002, 277, 7157–7164. [Google Scholar] [CrossRef] [PubMed]
- Balani, P.; Boulaire, J.; Zhao, Y.; Zeng, J.; Lin, J.; Wang, S. High mobility group box2 promoter-controlled suicide gene expression enables targeted glioblastoma treatment. Mol. Ther. 2009, 17, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Kros, J.M. Grading of gliomas: The road from eminence to evidence. J. Neuropathol. Exp. Neurol. 2011, 70, 101–109. [Google Scholar] [CrossRef]
- Lu, C.; Ward, P.S.; Kapoor, G.S.; Rohle, D.; Turcan, S.; Abdel-Wahab, O.; Edwards, C.R.; Khanin, R.; Figueroa, M.E.; Melnick, A.; et al. IDH mutation impairs histone demethylation and results in a block to cell differentiation. Nature 2012, 483, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Yang, Z.; Chen, D.; Xie, Q.; Peng, T.; Wu, J.; Qi, S. Downregulation of miR-130a promotes cell growth and epithelial to mesenchymal transition by activating HMGB2 in glioma. Int. J. Biochem. Cell Biol. 2017, 93, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Camara-Quilez, M.; Barreiro-Alonso, A.; Rodriguez-Bemonte, E.; Quindos-Varela, M.; Cerdan, M.E.; Lamas-Maceiras, M. Differential Characteristics of HMGB2 Versus HMGB1 and their Perspectives in Ovary and Prostate Cancer. Curr. Med. Chem. 2020, 27, 3271–3289. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Kang, R.; Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef]
- Tesniere, A.; Schlemmer, F.; Boige, V.; Kepp, O.; Martins, I.; Ghiringhelli, F.; Aymeric, L.; Michaud, M.; Apetoh, L.; Barault, L.; et al. Immunogenic death of colon cancer cells treated with oxaliplatin. Oncogene 2010, 29, 482–491. [Google Scholar] [CrossRef]
- Lu, V.M.; O’Connor, K.P.; Shah, A.H.; Eichberg, D.G.; Luther, E.M.; Komotar, R.J.; Ivan, M.E. The prognostic significance of CDKN2A homozygous deletion in IDH-mutant lower-grade glioma and glioblastoma: A systematic review of the contemporary literature. J. Neurooncol. 2020, 148, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y. High mobility group proteins and their post-translational modifications. Biochim. Biophys. Acta 2008, 1784, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.J.; Giaccia, A.J. Radiosensitivity and Cell Age in the Mitotic Cycle. In Radiobiology for the Radiologist; Wolters Kluwer Health: Philadelphia, PA, USA, 2012; pp. 54–66. [Google Scholar]
- Birmpilis, A.I.; Paschalis, A.; Mourkakis, A.; Christodoulou, P.; Kostopoulos, I.V.; Antimissari, E.; Terzoudi, G.; Georgakilas, A.G.; Armpilia, C.; Papageorgis, P.; et al. Immunogenic Cell Death, DAMPs and Prothymosin α as a Putative Anticancer Immune Response Biomarker. Cells 2022, 11, 1415. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, G.; Chen, Y.; Wang, H.; Hua, Y.; Cai, Z. Immunogenic cell death in cancer therapy: Present and emerging inducers. J. Cell Mol. Med. 2019, 23, 4854–4865. [Google Scholar] [CrossRef] [PubMed]
- Neubert, E.N.; DeRogatis, J.M.; Lewis, S.A.; Viramontes, K.M.; Ortega, P.; Henriquez, M.L.; Buisson, R.; Messaoudi, I.; Tinoco, R. HMGB2 regulates the differentiation and stemness of exhausted CD8T cells during chronic viral infection and cancer. Nat. Commun. 2023, 14, 5631. [Google Scholar] [CrossRef] [PubMed]
- Woller, N.; Gurlevik, E.; Ureche, C.I.; Schumacher, A.; Kuhnel, F. Oncolytic viruses as anticancer vaccines. Front. Oncol. 2014, 4, 188. [Google Scholar] [CrossRef] [PubMed]
- Skjulsvik, A.J.; Mork, J.N.; Torp, M.O.; Torp, S.H. Ki-67/MIB-1 immunostaining in a cohort of human gliomas. Int. J. Clin. Exp. Pathol. 2014, 7, 8905–8910. [Google Scholar]
- Dahlrot, R.H.; Bangso, J.A.; Petersen, J.K.; Rosager, A.M.; Sorensen, M.D.; Reifenberger, G.; Hansen, S.; Kristensen, B.W. Prognostic role of Ki-67 in glioblastomas excluding contribution from non-neoplastic cells. Sci. Rep. 2021, 11, 17918. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age (Years) | Tumor Order | Original Diagnosis | 2021 WHO | IDH Status | 1p19q Status | OS (Months) | Samples per Block |
|---|---|---|---|---|---|---|---|---|---|
| 1 | M | 28 | P | AA | A3IDHmut | Mut | intact | 31 | 3 |
| 2 | F | 24 | P | DA | A2IDHmut | Mut | intact | 2 | 3 |
| 3 | F | 49 | R | GBM * | A4IDHmut | Mut | intact | 15 | 2 |
| 4 | F | 55 | R | GBM * | A4IDHmut | Mut | intact | 16 | 2 |
| 5 | M | 86 | P | GBM | GBM | WT | intact | 6 | 3 |
| 6 | F | 56 | P | GBM | GBM | WT | intact | 22 | 3 |
| 7 | M | 81 | P | GBM | GBM | WT | intact | 10 | 3 |
| 8 | M | 69 | P | GBM | GBM | WT | intact | 4 | 3 |
| 9 | F | 44 | R | GBM | GBM | WT | intact | 6 | 3 |
| Sex | 184 Male | 122 Female | 5 NA | ||
| Age (categorical) | 156 < 60 years | 149 ≥ 60 years | 6 NA | ||
| Extension of surgery | 263 Resection | 42 Biopsy | 6 NA | ||
| 2021 WHO diagnosis | 287 Glioblastoma | 24 A4 IDHmut | |||
| HMGB2 overexpression 1 | 76 High | 223 Low | 12 NA | ||
| Adjuvant treatment | 110 RT + TMZ | 41 RT only | 38 None | 112 Others 2 | 10 NA |
| Age (continuous) | 25.8–91.2 years (Mean: 58.8; Median: 60.6) | ||||
| HMGB2 expression (score) 3 | 0–260 (Percentile 75 = 108) | ||||
| Overall survival | 0.07–169.4 months (Mean: 15.9; Median: 10.5) | ||||
| coef | exp(coef) | se(coef) | z | p | |
|---|---|---|---|---|---|
| IDH_IHCR132H | −0.0408 | 0.960013 | 0.545607 | −0.075 | 0.94038 |
| Geographic necrosis (yes) | −0.0727 | 0.929901 | 0.192487 | −0.378 | 0.70575 |
| Mitotic activity | 0.01406 | 1.014155 | 0.005462 | 2.573 | 0.01008 |
| Glomeruloid vasc (yes) | −0.0025 | 0.997489 | 0.185236 | −0.014 | 0.98917 |
| Sex (male) | 0.06345 | 1.065509 | 0.176581 | 0.359 | 0.71934 |
| Surgery (resection) | −0.2213 | 0.801442 | 0.312497 | −0.708 | 0.47876 |
| Adj treatment (others) | −2.6391 | 0.071428 | 0.377554 | −6.99 | 2.75 × 10−12 |
| Adj treatment (rt) | −1.4625 | 0.231664 | 0.323833 | −4.516 | 6.30 × 10−6 |
| Adj treatment (tmz + rt) | −2.5327 | 0.079442 | 0.301773 | −8.393 | <2.00 × 10−16 |
| Age cat (>=60 years) | 0.37144 | 1.449826 | 0.176933 | 2.099 | 0.03579 |
| HMGB2Categ (score >= 108) | −0.6352 | 0.529813 | 0.206351 | −3.078 | 0.00208 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, A.P.; Becker, V.; McElroy, J.; Webb, A.; Han, C.; Guo, Y.; Bell, E.H.; Fleming, J.; Popp, I.; Staszewski, O.; et al. Proteomic Analysis of Spatial Heterogeneity Identifies HMGB2 as Putative Biomarker of Tumor Progression in Adult-Type Diffuse Astrocytomas. Cancers 2024, 16, 1516. https://doi.org/10.3390/cancers16081516
Becker AP, Becker V, McElroy J, Webb A, Han C, Guo Y, Bell EH, Fleming J, Popp I, Staszewski O, et al. Proteomic Analysis of Spatial Heterogeneity Identifies HMGB2 as Putative Biomarker of Tumor Progression in Adult-Type Diffuse Astrocytomas. Cancers. 2024; 16(8):1516. https://doi.org/10.3390/cancers16081516
Chicago/Turabian StyleBecker, Aline P., Valesio Becker, Joseph McElroy, Amy Webb, Chunhua Han, Yingshi Guo, Erica H. Bell, Jessica Fleming, Ilinca Popp, Ori Staszewski, and et al. 2024. "Proteomic Analysis of Spatial Heterogeneity Identifies HMGB2 as Putative Biomarker of Tumor Progression in Adult-Type Diffuse Astrocytomas" Cancers 16, no. 8: 1516. https://doi.org/10.3390/cancers16081516
APA StyleBecker, A. P., Becker, V., McElroy, J., Webb, A., Han, C., Guo, Y., Bell, E. H., Fleming, J., Popp, I., Staszewski, O., Prinz, M., Otero, J. J., Haque, S. J., Grosu, A.-L., & Chakravarti, A. (2024). Proteomic Analysis of Spatial Heterogeneity Identifies HMGB2 as Putative Biomarker of Tumor Progression in Adult-Type Diffuse Astrocytomas. Cancers, 16(8), 1516. https://doi.org/10.3390/cancers16081516