
Identification and In Vivo Validation of Unique Anti-Oncogenic Mechanisms Involving Protein Kinase Signaling and Autophagy Mediated by the Investigational Agent PV-10


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cytotoxicity Assays
2.4. Western Blot Analysis
2.5. Phospho-Kinase Array
2.6. Wound Healing Assay
2.7. In Vivo Xenograft Models
2.8. Statistical Analysis
3. Results
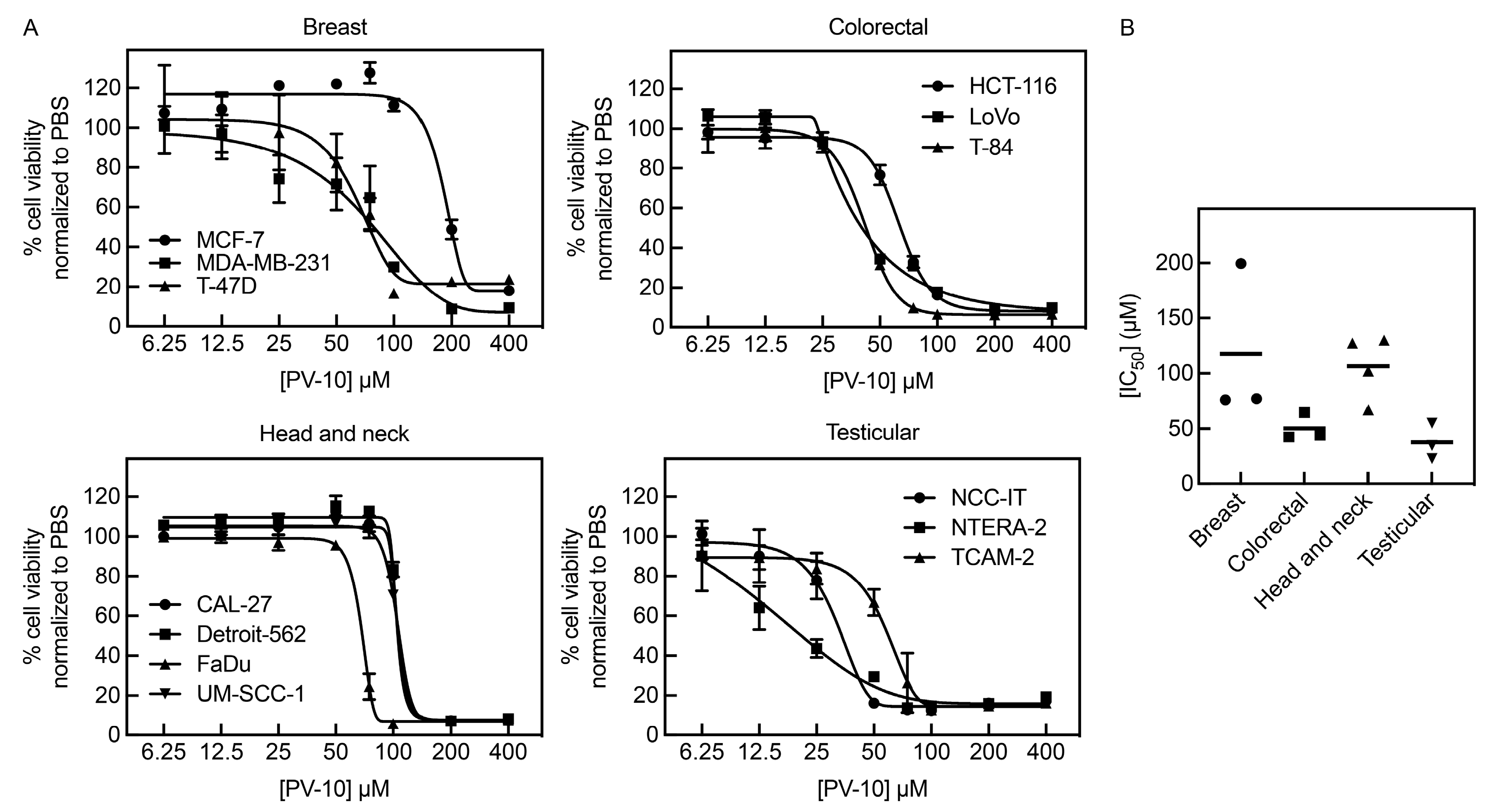
3.1. PV-10 Inhibits Growth of Diverse Adult Solid Tumor Cell Lines
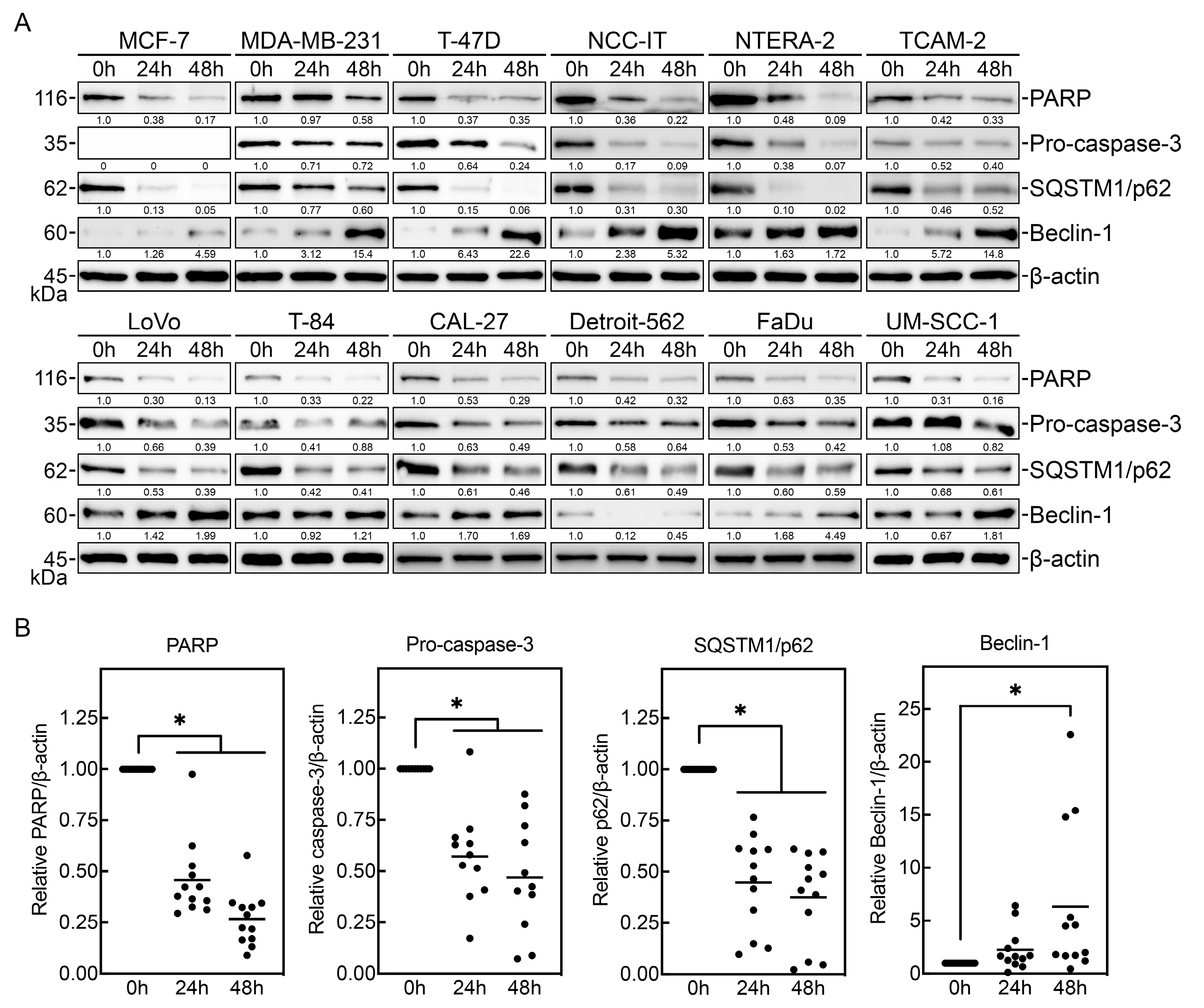
3.2. PV-10 Induces Cancer Cell Apoptosis and Autophagy
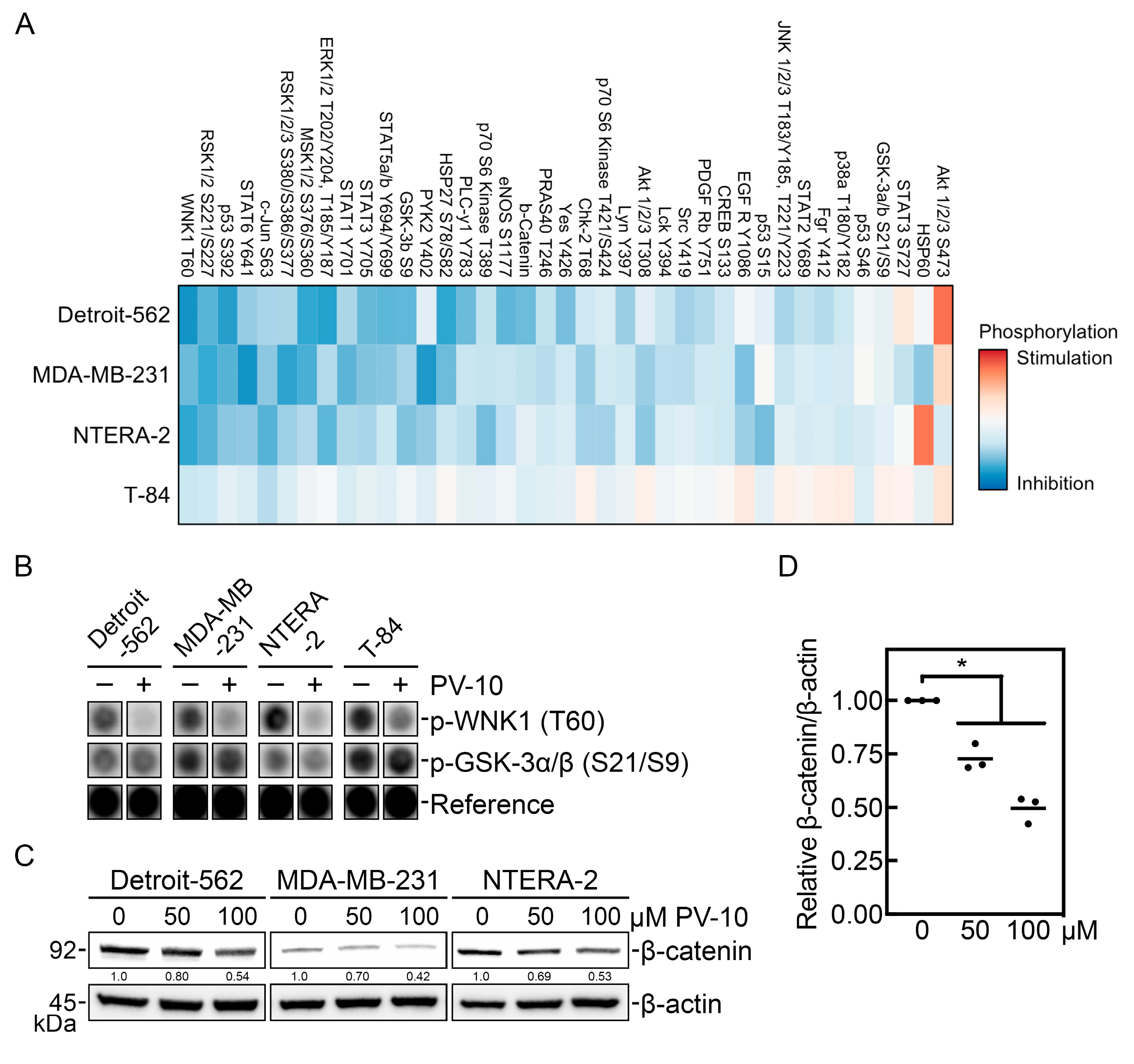
3.3. PV-10 Downregulates Key Phospho-Kinase Signaling Proteins
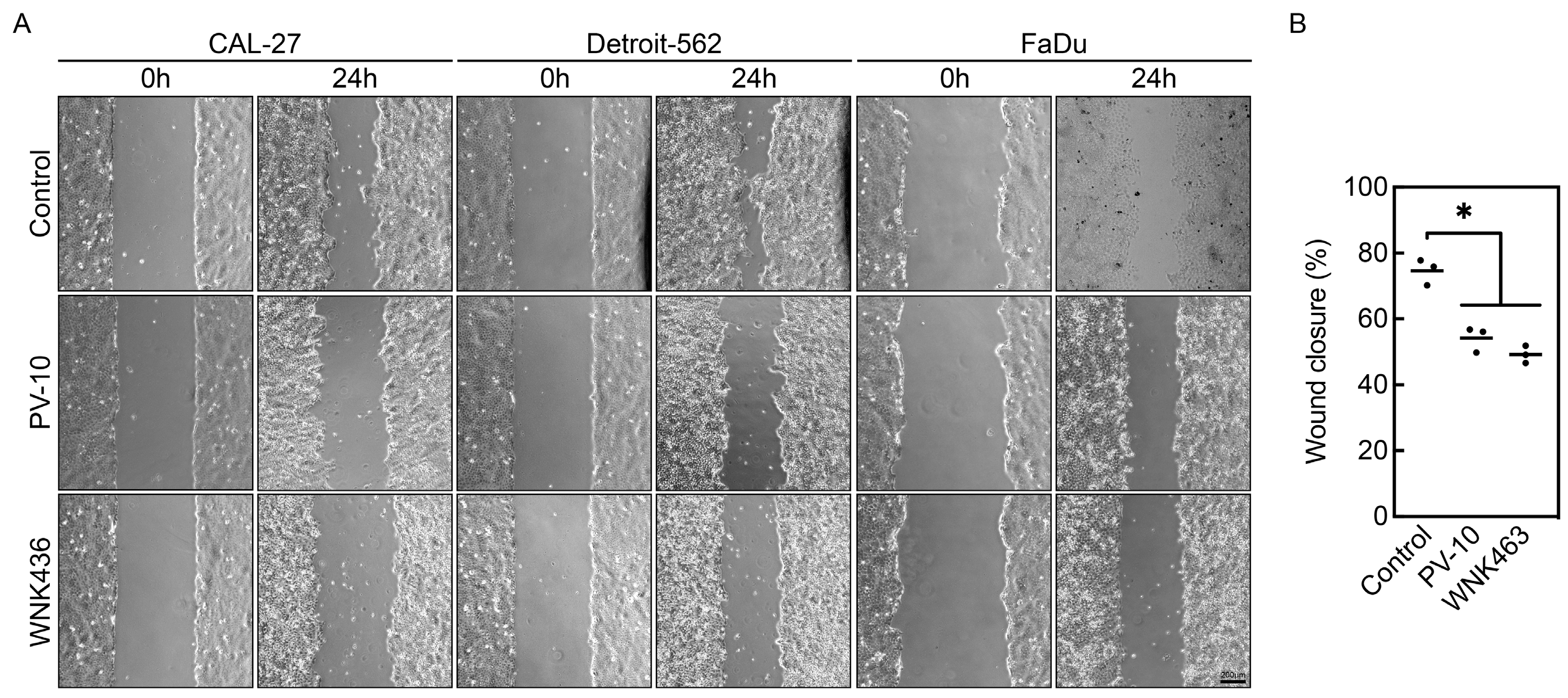
3.4. PV-10 Inhibits Cancer Cell Migration
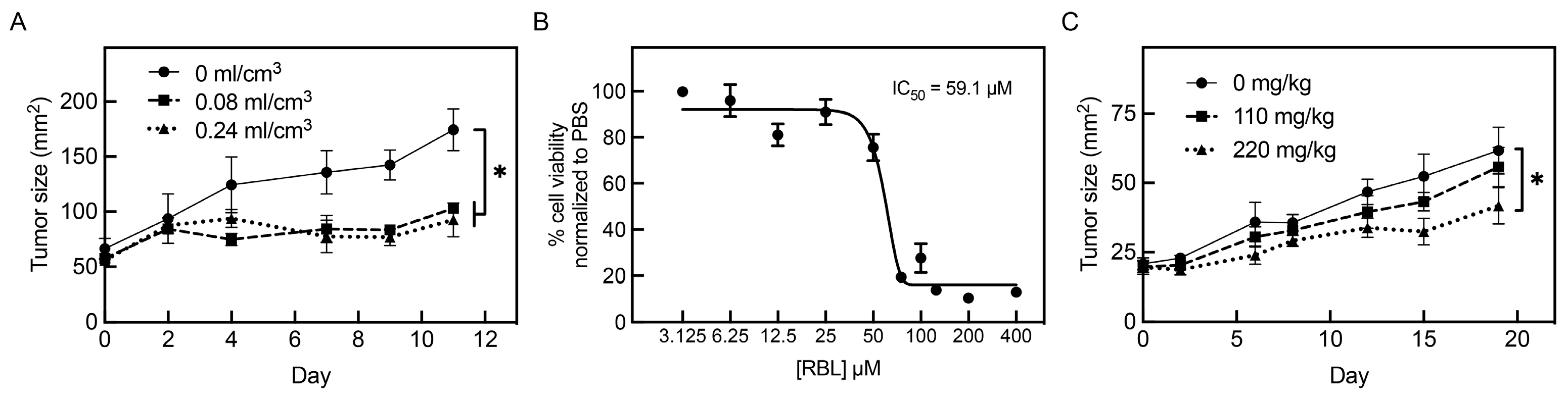
3.5. Intralesional and Systemic Administration of PV-10 Decreases Tumor Growth In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demartis, S.; Obinu, A.; Gavini, E.; Giunchedi, P.; Rassu, G. Nanotechnology-Based Rose Bengal: A Broad-Spectrum Biomedical Tool. Dyes Pigments 2021, 188, 109236. [Google Scholar] [CrossRef]
- Chen, H.-J.; Zhou, X.-B.; Wang, A.-L.; Zheng, B.-Y.; Yeh, C.-K.; Huang, J.-D. Synthesis and Biological Characterization of Novel Rose Bengal Derivatives with Improved Amphiphilicity for Sono-Photodynamic Therapy. Eur. J. Med. Chem. 2018, 145, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.N.; Bekmukhametova, A.; Antony, A.; Barman, S.K.; Houang, J.; Wu, M.J.; Hook, J.M.; George, L.; Wuhrer, R.; Mawad, D.; et al. Encapsulated Rose Bengal Enhances the Photodynamic Treatment of Triple-Negative Breast Cancer Cells. Molecules 2024, 29, 546. [Google Scholar] [CrossRef] [PubMed]
- Anjani, Q.K.; Demartis, S.; Volpe-Zanutto, F.; Li, H.; Sabri, A.H.B.; Gavini, E.; Donnelly, R.F. Fluorescence-Coupled Techniques for Determining Rose Bengal in Dermatological Formulations and Their Application to Ex Vivo Skin Deposition Studies. Pharmaceutics 2023, 15, 408. [Google Scholar] [CrossRef] [PubMed]
- Kuryanova, A.S.; Savko, M.A.; Kaplin, V.S.; Aksenova, N.A.; Timofeeva, V.A.; Chernyak, A.V.; Glagolev, N.N.; Timashev, P.S.; Solovieva, A.B. Effect of Chitosan and Amphiphilic Polymers on the Photosensitizing and Spectral Properties of Rose Bengal. Molecules 2022, 27, 6796. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.F.; Hersey, P.; Wachter, E. Chemoablation of Metastatic Melanoma Using Intralesional Rose Bengal. Melanoma Res. 2008, 18, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Lippey, J.; Bousounis, R.; Behrenbruch, C.; McKay, B.; Spillane, J.; Henderson, M.A.; Speakman, D.; Gyorki, D.E. Intralesional PV-10 for In-transit Melanoma—A Single-center Experience. J. Surg. Oncol. 2016, 114, 380–384. [Google Scholar] [CrossRef]
- Read, T.A.; Smith, A.; Thomas, J.; David, M.; Foote, M.; Wagels, M.; Barbour, A.; Smithers, B.M. Intralesional PV-10 for the Treatment of In-transit Melanoma Metastases—Results of a Prospective, Non-randomized, Single Center Study. J. Surg. Oncol. 2018, 117, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.F.; Agarwala, S.S.; Smithers, B.M.; Ross, M.I.; Scoggins, C.R.; Coventry, B.J.; Neuhaus, S.J.; Minor, D.R.; Singer, J.M.; Wachter, E.A. Phase 2 Study of Intralesional PV-10 in Refractory Metastatic Melanoma. Ann. Surg. Oncol. 2015, 22, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Foote, M.; Read, T.; Thomas, J.; Wagels, M.; Burmeister, B.; Smithers, B.M. Results of a Phase II, Open-label, Non-comparative Study of Intralesional PV-10 Followed by Radiotherapy for the Treatment of In-transit or Metastatic Melanoma. J. Surg. Oncol. 2017, 115, 891–897. [Google Scholar] [CrossRef]
- Thompson, J.F.; Saw, R.P.M.; Dalton, J.M.; Stretch, J.R.; Spillane, A.J.; Osborne, N.S.; Williams, G.J.; Lo, S.N. Treatment of In-Transit Melanoma Metastases Using Intralesional PV-10. Melanoma Res. 2021, 31, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Wiener, M.; Damian, D.L.; Thompson, J.F. Systemic Phototoxicity Following Intralesional Rose Bengal for Subcutaneous Mel-anoma Metastases. Dermatology 2008, 216, 361–362. [Google Scholar] [CrossRef] [PubMed]
- Toomey, P.; Kodumudi, K.; Weber, A.; Kuhn, L.; Moore, E.; Sarnaik, A.A.; Pilon-Thomas, S. Intralesional Injection of Rose Ben-gal Induces a Systemic Tumor-Specific Immune Response in Murine Models of Melanoma and Breast Cancer. PLoS ONE 2013, 8, e68561. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Kunda, N.; Qiao, G.; Calata, J.F.; Pardiwala, K.; Prabhakar, B.S.; Maker, A.V. Colon Cancer Cell Treatment with Rose Bengal Generates a Protective Immune Response via Immunogenic Cell Death. Cell Death Dis. 2017, 8, e2584. [Google Scholar] [CrossRef] [PubMed]
- Swift, L.; Zhang, C.; Trippett, T.; Narendran, A. Potent in Vitro and Xenograft Antitumor Activity of a Novel Agent, PV-10, against Relapsed and Refractory Neuroblastoma. OncoTargets Ther. 2019, 12, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Rabe, S.Z.T.; Mousavi, S.H.; Tabasi, N.; Rastin, M.; Rabe, S.Z.T.; Siadat, Z.; Mahmoudi, M. Rose Bengal Suppresses Gastric Cancer Cell Proliferation via Apoptosis and Inhibits Nitric Oxide Formation in Macrophages. J. Immunotoxicol. 2014, 11, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Koevary, S.B. Selective Toxicity of Rose Bengal to Ovarian Cancer Cells in Vitro. Int. J. Physiol. Pathophysiol. Pharmacol. 2012, 4, 99–107. [Google Scholar] [PubMed]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of Poly(ADP-Ribose) Polymerase (PARP) Cleavage in Apoptosis. Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef]
- Bjørkøy, G.; Lamark, T.; Pankiv, S.; Øvervatn, A.; Brech, A.; Johansen, T. Chapter 12 Monitoring Autophagic Degradation of P62/SQSTM1. Methods Enzym. 2009, 452, 181–197. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T. How to Interpret LC3 Immunoblotting. Autophagy 2007, 3, 542–545. [Google Scholar] [CrossRef]
- Li, D.-D.; Wang, L.-L.; Deng, R.; Tang, J.; Shen, Y.; Guo, J.-F.; Wang, Y.; Xia, L.-P.; Feng, G.-K.; Liu, Q.Q.; et al. The Pivotal Role of C-Jun NH2-Terminal Kinase-Mediated Beclin 1 Expression during Anticancer Agents-Induced Autophagy in Cancer Cells. Oncogene 2009, 28, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Kankanamalage, S.G.; Lee, A.-Y.; Wichaidit, C.; Lorente-Rodriguez, A.; Shah, A.M.; Stippec, S.; Whitehurst, A.W.; Cobb, M.H. Multistep Regulation of Autophagy by WNK1. Proc. Natl. Acad. Sci. USA 2016, 113, 14342–14347. [Google Scholar] [CrossRef] [PubMed]
- Kankanamalage, S.G.; Lee, A.-Y.; Wichaidit, C.; Lorente-Rodriguez, A.; Shah, A.M.; Stippec, S.; Whitehurst, A.W.; Cobb, M.H. WNK1 Is an Unexpected Autophagy Inhibitor. Autophagy 2017, 13, 969–970. [Google Scholar] [CrossRef] [PubMed]
- Serysheva, E.; Berhane, H.; Grumolato, L.; Demir, K.; Balmer, S.; Bodak, M.; Boutros, M.; Aaronson, S.; Mlodzik, M.; Jenny, A. Wnk Kinases Are Positive Regulators of Canonical Wnt/Β-catenin Signalling. EMBO Rep. 2013, 14, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Shimizu, M.; Goto, T.; Masuno, H.; Kagechika, H.; Tanaka, N.; Shibuya, H. WNK Regulates Wnt Signalling and β-Catenin Levels by Interfering with the Interaction between β-Catenin and GID. Commun. Biol. 2020, 3, 666. [Google Scholar] [CrossRef] [PubMed]
- Shyamasundar, S.; Lim, J.P.; Bay, B.H. MiR-93 Inhibits the Invasive Potential of Triple-Negative Breast Cancer Cells in Vitro via Protein Kinase WNK1. Int. J. Oncol. 2016, 49, 2629–2636. [Google Scholar] [CrossRef] [PubMed]
- Jaykumar, A.B.; Jung, J.-U.; Parida, P.K.; Dang, T.T.; Wichaidit, C.; Kannangara, A.R.; Earnest, S.; Goldsmith, E.J.; Pearson, G.W.; Malladi, S.; et al. WNK1 Enhances Migration and Invasion in Breast Cancer Models. Mol. Cancer Ther. 2021, 20, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Gao, L.; Yu, R.K.; Zeng, G. Down-regulation of WNK1 Protein Kinase in Neural Progenitor Cells Suppresses Cell Pro-liferation and Migration. J. Neurochem. 2006, 99, 1114–1121. [Google Scholar] [CrossRef]
- Kleszcz, R.; Szymańska, A.; Krajka-Kuźniak, V.; Baer-Dubowska, W.; Paluszczak, J. Inhibition of CBP/β-Catenin and Porcupine Attenuates Wnt Signaling and Induces Apoptosis in Head and Neck Carcinoma Cells. Cell. Oncol. 2019, 42, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.H.; Lee, S.H.; Lim, Y.C. Wnt/β-Catenin/Slug Pathway Contributes to Tumor Invasion and Lymph Node Metastasis in Head and Neck Squamous Cell Carcinoma. Clin. Exp. Metastasis 2021, 38, 163–174. [Google Scholar] [CrossRef]
- Yamada, K.; Park, H.-M.; Rigel, D.F.; DiPetrillo, K.; Whalen, E.J.; Anisowicz, A.; Beil, M.; Berstler, J.; Brocklehurst, C.E.; Burdick, D.A.; et al. Small-Molecule WNK Inhibition Regulates Cardiovascular and Renal Function. Nat. Chem. Biol. 2016, 12, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Innamarato, P.P.; Kodumudi, K.; Weber, A.; Nemoto, S.; Robinson, J.L.; Crago, G.; McCardle, T.; Royster, E.; Sarnaik, A.A.; et al. Intralesional Rose Bengal in Melanoma Elicits Tumor Immunity via Activation of Dendritic Cells by the Release of High Mobility Group Box 1. Oncotarget 2016, 7, 37893–37905. [Google Scholar] [CrossRef] [PubMed]
- Jänicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 Is Required for DNA Fragmentation and Morphological Changes Associated with Apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [PubMed]
- Jänicke, R.U. MCF-7 Breast Carcinoma Cells Do Not Express Caspase-3. Breast Cancer Res. Treat. 2009, 117, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Hirata, N.; Tanaka, T.; Suizu, F.; Nakajima, H.; Chiorini, J.A. Autophagy as a Modulator of Cell Death Machin-ery. Cell Death Dis. 2020, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-J.; Lee, S.-H.; Lee, C.-H.; Jang, J.-Y.; Chung, J.; Kwon, M.-H.; Kim, Y.-S. Upregulation of Beclin-1 Expression and Phos-phorylation of Bcl-2 and P53 Are Involved in the JNK-Mediated Autophagic Cell Death. Biochem. Biophys. Res. Commun. 2009, 382, 726–729. [Google Scholar] [CrossRef]
- Hung, J.-Y.; Yen, M.-C.; Jian, S.-F.; Wu, C.-Y.; Chang, W.-A.; Liu, K.-T.; Hsu, Y.-L.; Chong, I.-W.; Kuo, P.-L. Secreted Protein Acidic and Rich in Cysteine (SPARC) Induces Cell Migration and Epithelial Mesenchymal Transition through WNK1/Snail in Non-Small Cell Lung Cancer. Oncotarget 2017, 8, 63691–63702. [Google Scholar] [CrossRef] [PubMed]
- Fulford, L.; Milewski, D.; Ustiyan, V.; Ravishankar, N.; Cai, Y.; Le, T.; Masineni, S.; Kasper, S.; Aronow, B.; Kalinichenko, V.V.; et al. The Transcription Factor FOXF1 Promotes Prostate Cancer by Stimulating the Mitogen-Activated Protein Kinase ERK5. Sci. Signal. 2016, 9, ra48. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor | Cell Line | IC50 (μM) |
|---|---|---|
| Breast | MCF-7 | 199.5 |
| MDA-MB-231 | 76.99 | |
| T-47D | 75.98 | |
| Colorectal | HCT-116 | 64.79 |
| LoVo | 43.96 | |
| T-84 | 42.37 | |
| Head and neck | CAL-27 | 127.3 |
| Detroit-562 | 129.9 | |
| FaDu | 67.09 | |
| UM-SCC-1 | 102.0 | |
| Testicular | NCC-IT | 34.54 |
| NTERA-2 | 22.77 | |
| TCAM-2 | 55.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, S.; Sipila, P.; Thakur, S.; Zhang, C.; Narendran, A. Identification and In Vivo Validation of Unique Anti-Oncogenic Mechanisms Involving Protein Kinase Signaling and Autophagy Mediated by the Investigational Agent PV-10. Cancers 2024, 16, 1520. https://doi.org/10.3390/cancers16081520
Tran S, Sipila P, Thakur S, Zhang C, Narendran A. Identification and In Vivo Validation of Unique Anti-Oncogenic Mechanisms Involving Protein Kinase Signaling and Autophagy Mediated by the Investigational Agent PV-10. Cancers. 2024; 16(8):1520. https://doi.org/10.3390/cancers16081520
Chicago/Turabian StyleTran, Son, Patrick Sipila, Satbir Thakur, Chunfen Zhang, and Aru Narendran. 2024. "Identification and In Vivo Validation of Unique Anti-Oncogenic Mechanisms Involving Protein Kinase Signaling and Autophagy Mediated by the Investigational Agent PV-10" Cancers 16, no. 8: 1520. https://doi.org/10.3390/cancers16081520
APA StyleTran, S., Sipila, P., Thakur, S., Zhang, C., & Narendran, A. (2024). Identification and In Vivo Validation of Unique Anti-Oncogenic Mechanisms Involving Protein Kinase Signaling and Autophagy Mediated by the Investigational Agent PV-10. Cancers, 16(8), 1520. https://doi.org/10.3390/cancers16081520