
Effects of Tetrahydrolipstatin on Glioblastoma in Mice: MRI-Based Morphologic and Texture Analysis Correlated with Histopathology and Immunochemistry Findings—A Pilot Study
 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Xenograft Transplantation
2.2. In Vivo Treatment with the FASN Inhibitor Orlistat
2.3. Animal Preparation for MRI Examination and MRI Protocol
2.4. MR Image Evaluation and Texture Analysis
2.5. Pathological Analysis
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
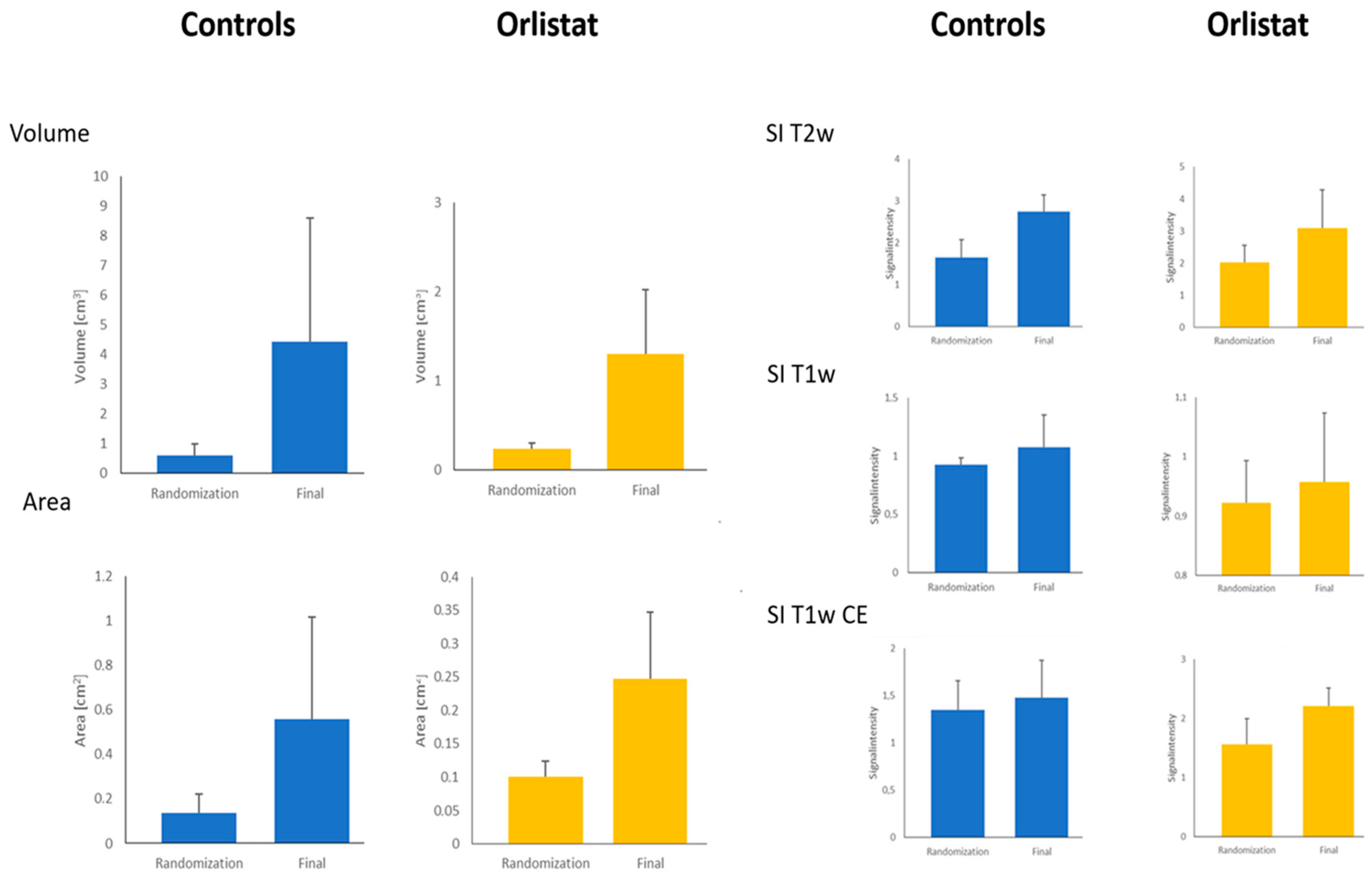
3.1. MR Image Evaluation and Texture Analysis
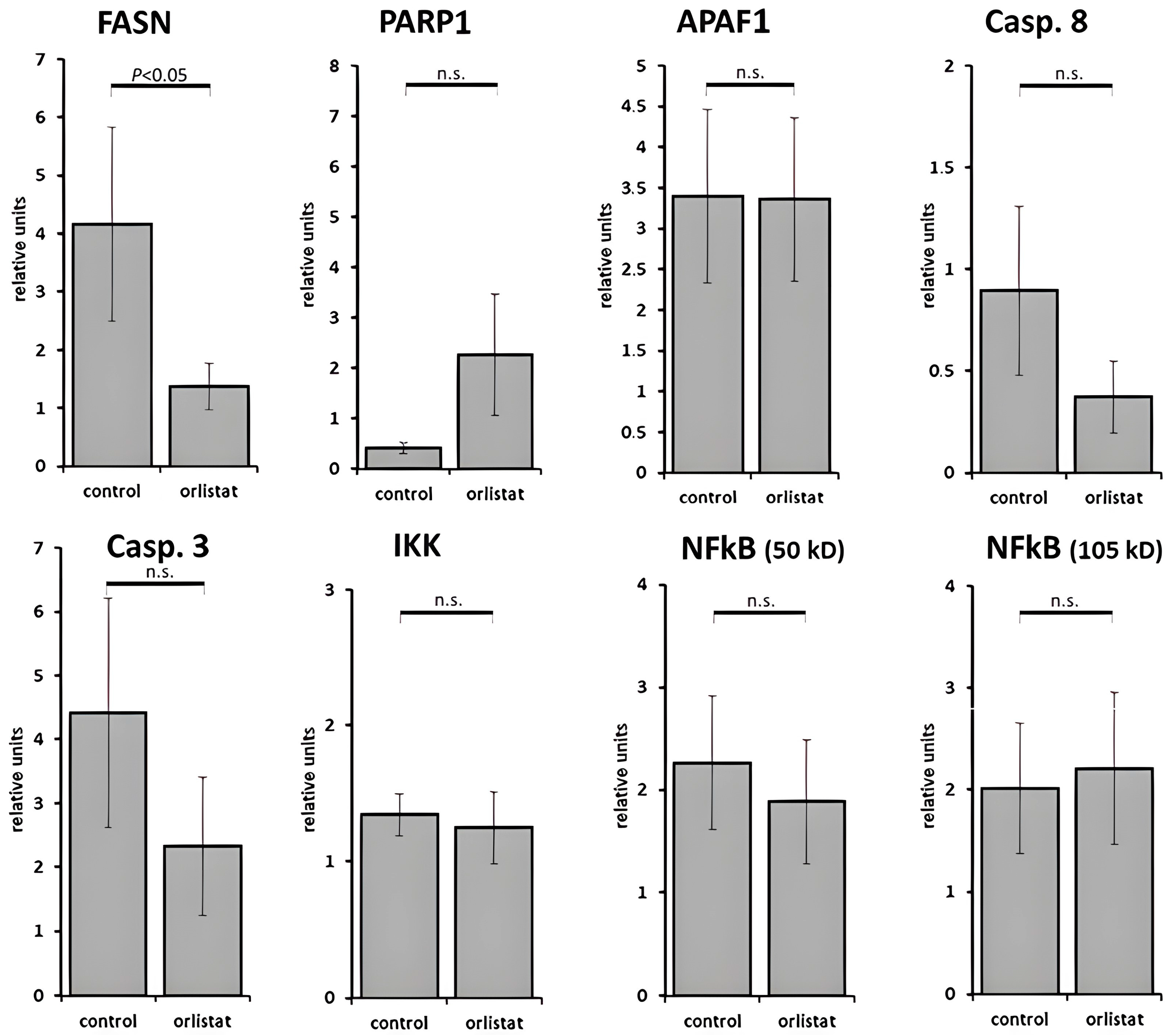
3.2. Histological and Laboratory Examination
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CE | contrast-enhanced |
| FASN | fatty acid synthase |
| FFA | free fatty acids |
| GBM | glioblastoma multiforme |
| Gd DTPA | gadolinium diethylenetriaminepentaacetic acid |
| GFAP | glial fibrillary acidic protein |
| MR | magnetic resonance |
| MRI | magnetic resonance imaging |
| ROI | region of interest |
| SI | signal intensity |
| T | Tesla |
| TSE | turbo spin echo |
| T1w | T1-weighted |
| T2w | T2-weighted |
References
- Ostrom, Q.T.; Gittleman, H.; Farah, P.; Ondracek, A.; Chen, Y.; Wolinsky, Y.; Stroup, N.E.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2006–2010. Neuro. Oncol. 2013, 15 (Suppl. S2), ii1–ii56. [Google Scholar] [CrossRef]
- Smoll, N.R.; Schaller, K.; Gautschi, O.P. Long-term survival of patients with glioblastoma multiforme (GBM). J. Clin. Neurosci. 2013, 20, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Sottoriva, A.; Spiteri, I.; Piccirillo, S.G.; Touloumis, A.; Collins, V.P.; Marioni, J.C.; Curtis, C.; Watts, C.; Tavaré, S. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Pessina, F.; Navarria, P.; Cozzi, L.; Ascolese, A.M.; Simonelli, M.; Santoro, A.; Clerici, E.; Rossi, M.; Scorsetti, M.; Bello, L. Maximize surgical resection beyond contrast-enhancing boundaries in newly diagnosed glioblastoma multiforme: Is it useful and safe? A single institution retrospective experience. J. Neurooncol. 2017, 135, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Szopa, W.; Burley, T.A.; Kramer-Marek, G.; Kaspera, W. Diagnostic and therapeutic biomarkers in glioblastoma: Current status and future perspectives. Biomed. Res. Int. 2017, 2017, 8013575. [Google Scholar] [CrossRef]
- Aum, D.J.; Kim, D.H.; Beaumont, T.L.; Leuthardt, E.C.; Dunn, G.P.; Kim, A.H. Molecular and cellular heterogeneity: The hallmark of glioblastoma. Neurosurg. Focus. 2014, 37, E11. [Google Scholar] [CrossRef] [PubMed]
- Seymour, L.; Bogaerts, J.; Perrone, A.; Ford, R.; Schwartz, L.H.; Mandrekar, S.; Lin, N.U.; Litière, S.; Dancey, J.; Chen, A.; et al. RECIST working group. iRECIST: Guidelines for response criteria for use in trials testing immunotherapeutics. Immunother. Lancet Oncol. 2017, 18, e143–e152. [Google Scholar] [CrossRef]
- Wen, P.Y.; Macdonald, D.R.; Reardon, D.A.; Cloughesy, T.F.; Sorensen, A.G.; Galanis, E.; Degroot, J.; Wick, W.; Gilbert, M.R.; Lassman, A.B.; et al. Updated response assessment criteria for high-grade gliomas: Response assessment in neuro-oncology working group. J. Clin. Oncol. 2010, 28, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Lanfermann, H.; Eichner, G.; Gufler, H. Radiation injury versus malignancy after stereotactic radiosurgery for brain metastases: Impact of time-dependent changes in lesion morphology on MRI. Neuro. Oncol. 2017, 19, 586–594. [Google Scholar] [CrossRef]
- Haralick, R.M. Statistical and structural approaches to texture. Proc. IEEE 1979, 5, 786–804. [Google Scholar] [CrossRef]
- Li, X.; Zhu, H.; Qian, X.; Chen, N.; Lin, X.Z. MRI Texture Analysis for Differentiating Nonfunctional pancreatic Neuroendocrine Neoplasms from Solid Pseudopapillary Neoplasms of the Pancreas. Acad. Radiol. 2020, 27, 815–823. [Google Scholar] [CrossRef]
- Schick, U.; Lucia, F.; Dissaux, G.; Visvikis, D.; Badic, B.; Masson, I.; Pradier, O.; Bourbonne, V.; Hatt, M. MRI-derived radiomics: Methodology and clinical applications in the field of pelvic oncology. Br. J. Radiol. 2019, 92, 20190105. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, N.; Hartmann, S.K.; Chang, Y.H.; Delfanti, R.; White, N.S.; Karunamuni, R.; Seibert, T.M.; Dale, A.M.; Hattangadi-Gluth, J.A.; Piccioni, D.; et al. Molecular classification of patients with grade II/III glioma using quantitative MRI characteristics. J. Neurooncol 2018, 139, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Kuhajda, F.P.; Jenner, K.; Wood, F.D.; Hennigar, R.A.; Jacobs, L.B.; Dick, J.D.; Pasternack, G.R. Fatty axid synthesis: A potential selective target for antineoplastic therapy. Proc. Natl. Acad. Sci. USA 1994, 91, 6379–6383. [Google Scholar] [CrossRef] [PubMed]
- Grube, S.; Dünisch, P.; Freitag, D.; Klausnitzer, M.; Sakr, Y.; Walter, J.; Kalff, R.; Ewald, C. Overexpression of fatty acid synthase in human gliomas correlates with the WHO tumor grade and inhibition with Orlistat reduces cell viability and triggers apoptosis. J. Neurooncol. 2014, 118, 277–287. [Google Scholar] [CrossRef] [PubMed]
- VASARI Project. Available online: https://wiki.cancerimagingarchive.net/ (accessed on 9 June 2016).
- Strzelecki, M.; Szczypinski, P.; Materka, A.; Klepaczko, A. A software tool for automatic classification and segmentation of 2D/3D medical images. Nucl. Instrum. Methods Phys. Res. A 2013, 702, 137–140. [Google Scholar] [CrossRef]
- Matsunaga de Angelis, C.; Alcides de Lima-Souza, R.; Figueira Scarini, J.; Said Abu Egal, E.; Kleber do Amaral-Silva, G.; de Oliveira Gondak, R.; de Almeida, O.P.; Chone, C.T.; Kowalski, L.P.; Altemani, A.; et al. Immunohistochemical Expression of Fatty Acid Synthase (FASN) is Correlated to Tumor Aggressiveness and Cellular Differentiation in Salivary Gland Carcinomas. Head Neck Pathology. 2021, 15, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Menendez, J.A.; Lupu, R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat. Rev. Cancer. 2007, 7, 763–777. [Google Scholar]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V., Jr. Cellular fatty acid metabolism and cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef]
- Rohrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Rev. Cancer. 2016, 16, 732–749. [Google Scholar] [CrossRef]
- Haberl, E.M.; Weiss, T.S.; Peschel, G.; Weigand, K.; Köhler, N.; Pauling, J.K.; Wenzel, J.J.; Höring, M.; Krautbauer, S.; Liebisch, G.; et al. Liver Lipids of Patients with Hepatitis B and C and Associated hepatocellular carcinoma. Int. J. Mol. Sci. 2021, 22, 5297. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Aslanidis, C. Role of lipids in pathophysiology, diagnosis and therapy of hepatocellular carcinoma. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2020, 1865, 158658. [Google Scholar] [CrossRef] [PubMed]
- Sounni, N.E.; Cimino, J.; Blacher, S.; Primac, I.; Truong, A.; Mazzucchelli, G.; Paye, A.; Calligaris, D.; Debois, D.; De Tullio, P.; et al. Blocking lipid synthesis overcomes tumor regrowth and metastasis after angiogenetic therapy withdrawal. Cell Metab. 2014, 20, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Tucci, J.; Chen, T.; Margulis, K.; Orgel, E.; Paszkiewicz, R.L.; Cohen, M.D.; Oberley, M.J.; Wahhab, R.; Jones, A.E.; Divakaruni, A.S.; et al. Adipocytes Provide Fatty Acids to Acute Lymphoblastic Leukemia Cells. Front. Oncol. 2021, 11, 665763. [Google Scholar] [CrossRef] [PubMed]
- Stamatakos, S.; Beretta, G.L.; Vergani, E.; Dugo, M.; Corno, C.; Corna, E.; Tinelli, S.; Frigerio, S.; Ciusani, E.; Rodolfo, M.; et al. Deregulated FASN Expression in BRAF Inhibitor-Resistant Melanoma Cells Unveils New Targets for Drug Combinations. Cancers 2021, 13, 2284. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.Y.; Moreira, F.D.S.; Santos, G.A.S.D.; Cuadra Zelaya, F.J.M.; Ortiz, C.A.; Agostini, M.; Mariano, F.S.; Bastos, D.C.; Daher, U.R.N.; Kowalski, L.P.; et al. FASN inhibition sensitizes metastatic OSCC cells to cisplatin and paclitaxel by downregulating cyclin B1. Oral Dis. 2023, 29, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, W.; Zheng, Y.; Dai, W.; Ji, J.; Wu, L.; Cheng, Z.; Zhang, J.; Li, J.; Xu, X.; et al. Targeting fatty acid synthase modulates sensitivity of hepatocellular carcinoma to sorafenib via ferroptosis. J. Exp. Clin. Cancer Res. 2023, 42, 6. [Google Scholar] [CrossRef] [PubMed]
- Shueng, P.W.; Chan, H.W.; Lin, W.C.; Kuo, D.Y.; Chuang, H.Y. Orlistat Resensitizes Sorafenib-Resistance in Hepatocellular Carcinoma Cells through Modulating Metabolism. Int. J. Mol. Sci. 2022, 23, 6501. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Prins, R.M.; Dang, J.; Kuga, D.; Iwanami, A.; Soto, H.; Lin, K.Y.; Huang, T.T.; Akhavan, D.; Hock, M.B.; et al. EGFR signaling through an Akt-SREBP-1-dependent, rapamycin-resistant pathway sensitizes glioblastomas to antilipogenic therapy. Sci. Signal 2009, 2, ra82. [Google Scholar] [CrossRef]
- Harris, D.M.; Li, L.; Chen, M.; Lagunero, F.T.; Go, V.L.; Boros, L.G. Diverse mechanisms of growth inhibition by luteolin, resveratrol, and quercetin in MIA PaCa-2 cells: A comparative glucose tracer study with the fatty acid synthase inhibitor C75. Metabolomics 2012, 8, 201–210. [Google Scholar] [CrossRef]
- Alieva, M.; Leidgens, V.; Riemenschneider, M.J.; Klein, C.A.; Hau, P.; van Rheenen, J. Intravital imaging of glioma border morphology reveals distinctive cellular dynamics and contribution to tumor cell invasion. Sci. Rep. 2019, 9, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Mayerhoefer, M.E.; Materka, A.; Langs, G.; Häggström, I.; Szczypiński, P.; Gibbs, P.; Cook, G. Introduction to Radiomics. Nucl. Med. 2020, 61, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Eresen, A.; Yang, J.; Shangguan, J.; Li, Y.; Hu, S.; Sun, C.; Velichko, Y.; Yaghmai, V.; Benson, A.B., 3rd; Zhang, Z. MRI radiomics for early prediction of response to vaccine therapy in a transgenic mouse model of pancreatic ductal adenocarcinoma. J. Transl. Med. 2020, 18, 61. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Fu, S.; Zhu, Y.; Liu, Z.; Chen, S.; Lu, L.; Liang, C. Computed tomography texture analysis to facilitate therapeutic decision making in hepatocellular carcinoma. Oncotarget 2016, 7, 13248–13259. [Google Scholar] [CrossRef] [PubMed]
- Papadomanolakis, T.N.; Sergaki, E.S.; Polydorou, A.A.; Krasoudakis, A.G.; Makris-Tsalikis, G.N.; Polydorou, A.A.; Afentakis, N.M.; Athanasiou, S.A.; Vardiambasis, I.O.; Zervakis, M.E. Tumor Diagnosis against Other Brain Diseases Using T2 MRI Brain Images and CNN Binary Classifier and DWT. Brain Sci. 2023, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N. Molecular pathology of malignant gliomas. Annu. Rev. Pathol. 2006, 1, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Capper, D.; Jeibmann, A.; Habel, A.; Paulus, W.; Troost, D.; von Deimling, A. Addressing Diffuse Glioma as a Systemic Brain Disease With Single-Cell Analysis. Arch. Neurol. 2012, 69, 523–526. [Google Scholar] [PubMed]
- Claes, A.; Idema, A.J.; Wesseling, P. Diffuse glioma growth: A guerilla war. Acta Neuropathol. 2007, 114, 443–458. [Google Scholar] [CrossRef]
- Kim, J.; Lee, I.H.; Cho, H.J.; Park, C.K.; Jung, Y.S.; Kim, Y.; Nam, S.H.; Kim, B.S.; Johnson, M.D.; Kong, D.S.; et al. Spatiotemporal Evolution of the Primary Glioblastoma Genome. Cancer Cell. 2015, 28, 318–328. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T2-SPACE | T2-SPACE FS | T1-VIBE | |
|---|---|---|---|
| Dimension | 3D | 3D | 3D |
| Fat suppression | no | SPAIR | no |
| TR [ms] | 2500 | 2500 | 16 |
| TE [ms] | 289 | 289 | 2.66 |
| Flip angle | variable | variable | 20.5 |
| Echo train length | 71 | 71 | - |
| Matrix | 192 | 192 | 224 |
| FOV [mm] | 50 | 50 | 54 |
| FOV phase [%] | 71.9 | 71.9 | 93.8 |
| NEX | 1 | 1 | 2 |
| Bandwidth [Hz/Px] | 145 | 145 | 280 |
| Slice thickness [mm] | 0.25 | 0.25 | 0.25 |
| Slices in stack [nr] | 120 | 120 | 128 |
| Voxel size [mm] | 0.25 × 0.3 × 0.3 | 0.25 × 0.3 × 0.3 | 0.2 × 0.2 × 0.3 |
| Acquisition time [min] | 8:47 | 8:47 | 8:44 |
| Antigen | Antibody | Species | Manufacturer | Dilution | Cat# |
|---|---|---|---|---|---|
| APAF1 | Anti-APAF1 antibody | Rabbit | Abcam, Cambridge, UK | 1:1000 | ab32372 |
| Caspase-3 | Caspase-3 antibody | Rabbit | Cell Signaling Technology Inc., Danvers, MA, USA | 1:1000 | 9332S |
| Caspase-8 | Anti-Caspase-8 antibody | Rabbit | Abcam, Cambridge, UK | 1:2000 | ab184721 |
| FASN | Purified Mouse Anti-Fatty Acid Synthase | Mouse | Becton Dickinson, Franklin Lakes, NJ, USA | 1:1000 | 610963 |
| GAPDH | Anti-GAPDH antibody | Mouse | Abcam, Cambridge, UK | 1:5000 | ab9484 |
| IκB-alpha | Anti-IKB alpha (phospho S32 + S36) | Mouse | Abcam, Cambridge, UK | 1:500 | ab12135 |
| NFκB p50 | NFκB p50 Recombinant Polyclonal Antibody (24HCLC) | Rabbit | Thermo Fisher Scientific Inc., Langenselbold, Germany | 1:100 | 710450 |
| NFκB p105 | NFκB p105 Recombinant Polyclonal Antibody | Rabbit | Thermo Fisher Scientific Inc., Langenselbold, Germany | 1:1000 | 23576-1-AP |
| PARP1 | Anti-Poly (ADP-Ribose) Polymerase | Rabbit | Roche Diagnostic GmbH, Mannheim, Germany | 1:2000 | 11835238001 |
| Antibody | Manufacturer | Dilution | Cat# |
|---|---|---|---|
| Donkey anti-rabbit IgG-HRP | Santa Cruz Biotechnology, Inc., Paso Robles, CA, USA | 1:5000 | sc-2313 |
| Goat anti-mouse IgG-HRP | Santa Cruz Biotechnology, Inc., Paso Robles, CA, USA | 1:5000 | sc-2024 |
| Timepoint of Randomization | ||||||||
| Mouse No. | Enhancement quality | Proportion of enhancement | Proportion of necrosis | SI rim T1 nativ | Thickness of contrast-enhancing rim | Pattern of tumor growth | Hemorrhage | Neovascular proliferation |
| A 1 | mild | 2/3 | 0 | iso | thick | concentric | no | no |
| A 2 | marked | 3/3 | 0 | iso | thick | concentric | no | yes |
| A 3 | mild | 3/3 | 0 | iso | thick | concentric | yes | yes |
| A 4 | mild | 3/3 | 0 | iso | thick | concentric | no | yes |
| A 5 | mild | 3/3 | 0 | hypo | thick | concentric | no | no |
| A 6 | marked | 3/3 | 0 | iso | thick | concentric | no | no |
| A 7 | n|a | n|a | 0 | iso | n|a | concentric | no | no |
| B 1 | marked | 3/3 | 0 | iso | thin | concentric | no | no |
| B 2 | n|a | n|a | 0 | iso | n|a | concentric | no | no |
| B 3 | mild | 3/3 | 0 | iso | thick | concentric | no | no |
| B 4 | mild | 3/3 | 0 | iso | thick | concentric | no | no |
| B 5 | marked | 1/3 | 0 | iso | thick | concentric | no | no |
| B 6 | marked | 2/3 | 0 | iso | thick | concentric | no | no |
| Final MR Examination | ||||||||
| Mouse No. | Enhancement quality | Proportion of enhancement | Proportion of necrosis | SI rim T1 nativ | Thickness of contrast- enhancing rim | Pattern of tumor growth | Hemorrhage | Neovascular proliferation |
| A 1 | mild | 3/3 | 0 | iso | thick | concentric | no | yes |
| A 2 | mild | 2/3 | 1/3 | iso | thick | concentric | no | yes |
| A 3 | marked | 2/3 | 1/3 | iso | thick | concentric | yes | yes |
| A 4 | marked | 2/3 | 1/3 | iso | thick | concentric | no | yes |
| A 5 | mild | 3/3 | 0 | iso | thick | concentric | no | no |
| A 6 | marked | 2/3 | 0 | iso | thick | concentric | no | no |
| A 7 | n|a | n|a | 0 | iso | n|a | concentric | no | no |
| B 1 | marked | 2/3 | 0 | iso | thick | eccentric | no | no |
| B 2 | n|a | n|a | 0 | iso | n|a | eccentric | no | no |
| B 3 | marked | 3/3 | 0 | iso | thick | eccentric | no | no |
| B 4 | marked | 3/3 | 0 | iso | thick | eccentric | no | yes |
| B 5 | marked | 1/3 | 0 | iso | thick | eccentric | no | no |
| B 6 | marked | 2/3 | 0 | iso | thick | eccentric | no | no |
| Mouse No. | Necrosis | Vessel Proliferation | Apoptosis | Ki67 [%] | GFAP | MAP2 | p53 [%] |
|---|---|---|---|---|---|---|---|
| A 1 | no | yes | scattered | 10 | n|a | n|a | 10 |
| A 2 | yes | yes | scattered | 30 | + | + | 20 |
| A 3 | yes | yes | scattered | 40 | − | − | 10 |
| A 4 | yes | yes | scattered | 30 | − | + | 40 |
| A 5 | no | no | no | n|e | + | + | − |
| A 6 | no | no | no | 20 | − | − | 5 |
| A 7 | no | no | no | 20 | − | − | 10 |
| B 1 | no | no | no | 90 | +++ | +++ | 40 |
| B 2 | no | no | no | 70 | +++ | +++ | 40 |
| B 3 | no | no | scattered | 20 | − | + | 10 |
| B 4 | no | yes | scattered | 20 | + | − | 15 |
| B 5 | no | no | no | 20 | − | + | 5 |
| B 6 | no | no | no | 20 | − | − | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, S.; Ewald, C.; Freitag, D.; Herrmann, K.-H.; Koch, A.; Bauer, J.; Vogl, T.J.; Kemmling, A.; Gufler, H. Effects of Tetrahydrolipstatin on Glioblastoma in Mice: MRI-Based Morphologic and Texture Analysis Correlated with Histopathology and Immunochemistry Findings—A Pilot Study. Cancers 2024, 16, 1591. https://doi.org/10.3390/cancers16081591
Wagner S, Ewald C, Freitag D, Herrmann K-H, Koch A, Bauer J, Vogl TJ, Kemmling A, Gufler H. Effects of Tetrahydrolipstatin on Glioblastoma in Mice: MRI-Based Morphologic and Texture Analysis Correlated with Histopathology and Immunochemistry Findings—A Pilot Study. Cancers. 2024; 16(8):1591. https://doi.org/10.3390/cancers16081591
Chicago/Turabian StyleWagner, Sabine, Christian Ewald, Diana Freitag, Karl-Heinz Herrmann, Arend Koch, Johannes Bauer, Thomas J. Vogl, André Kemmling, and Hubert Gufler. 2024. "Effects of Tetrahydrolipstatin on Glioblastoma in Mice: MRI-Based Morphologic and Texture Analysis Correlated with Histopathology and Immunochemistry Findings—A Pilot Study" Cancers 16, no. 8: 1591. https://doi.org/10.3390/cancers16081591
APA StyleWagner, S., Ewald, C., Freitag, D., Herrmann, K.-H., Koch, A., Bauer, J., Vogl, T. J., Kemmling, A., & Gufler, H. (2024). Effects of Tetrahydrolipstatin on Glioblastoma in Mice: MRI-Based Morphologic and Texture Analysis Correlated with Histopathology and Immunochemistry Findings—A Pilot Study. Cancers, 16(8), 1591. https://doi.org/10.3390/cancers16081591