
Claudin11 Promoter Hypermethylation Is Frequent in Malignant Melanoma of the Skin, but Uncommon in Nevus Cell Nevi


Abstract
:1. Introduction
2. Results
2.1. Epigenetic Inactivation of CLDN11 in Malignant Melanoma


2.2. CLDN11 Promoter Hypermethylation Occurs Frequently in Melanoma, Is a Rare Event in Nevi


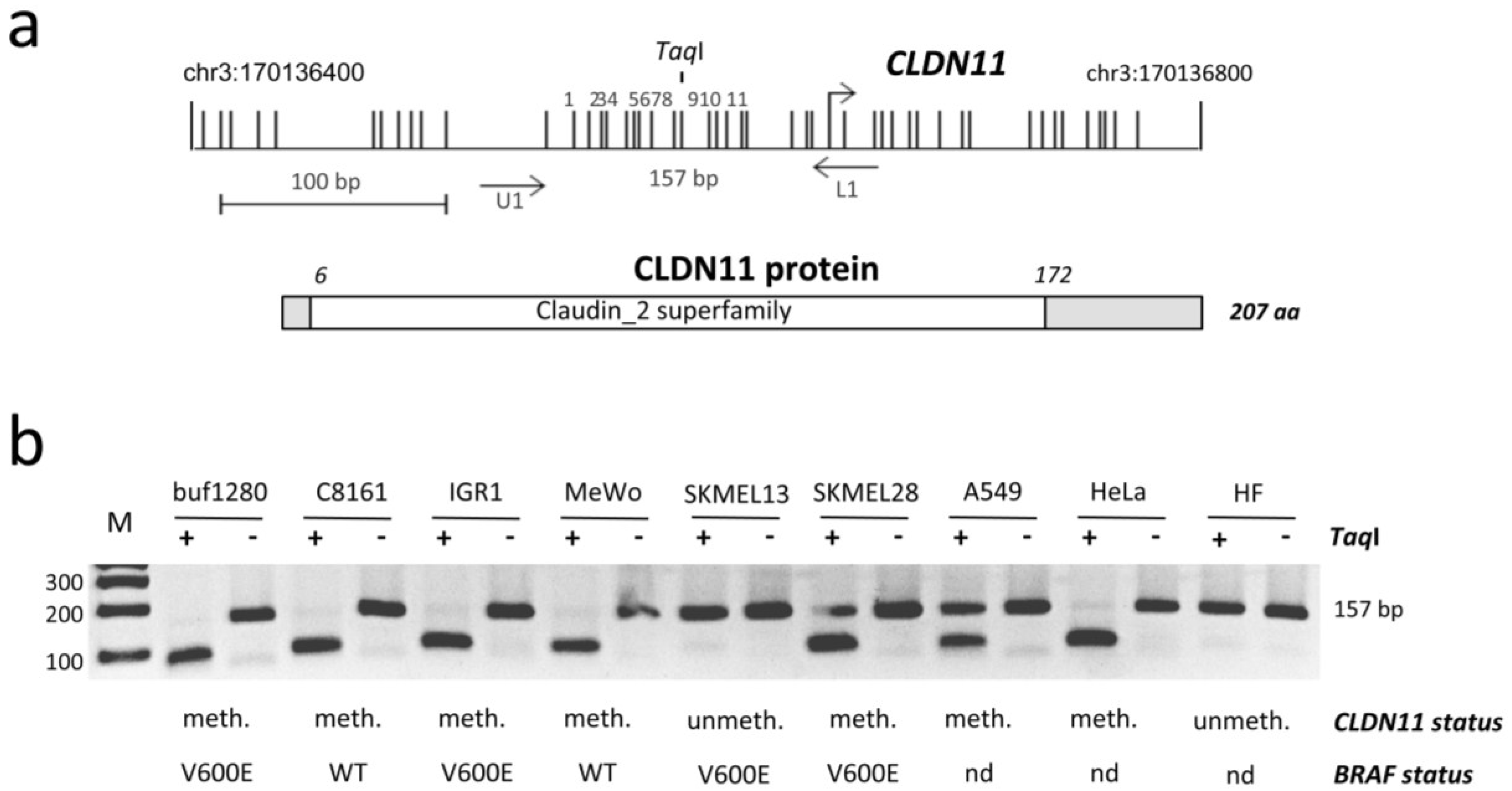{kind=link}
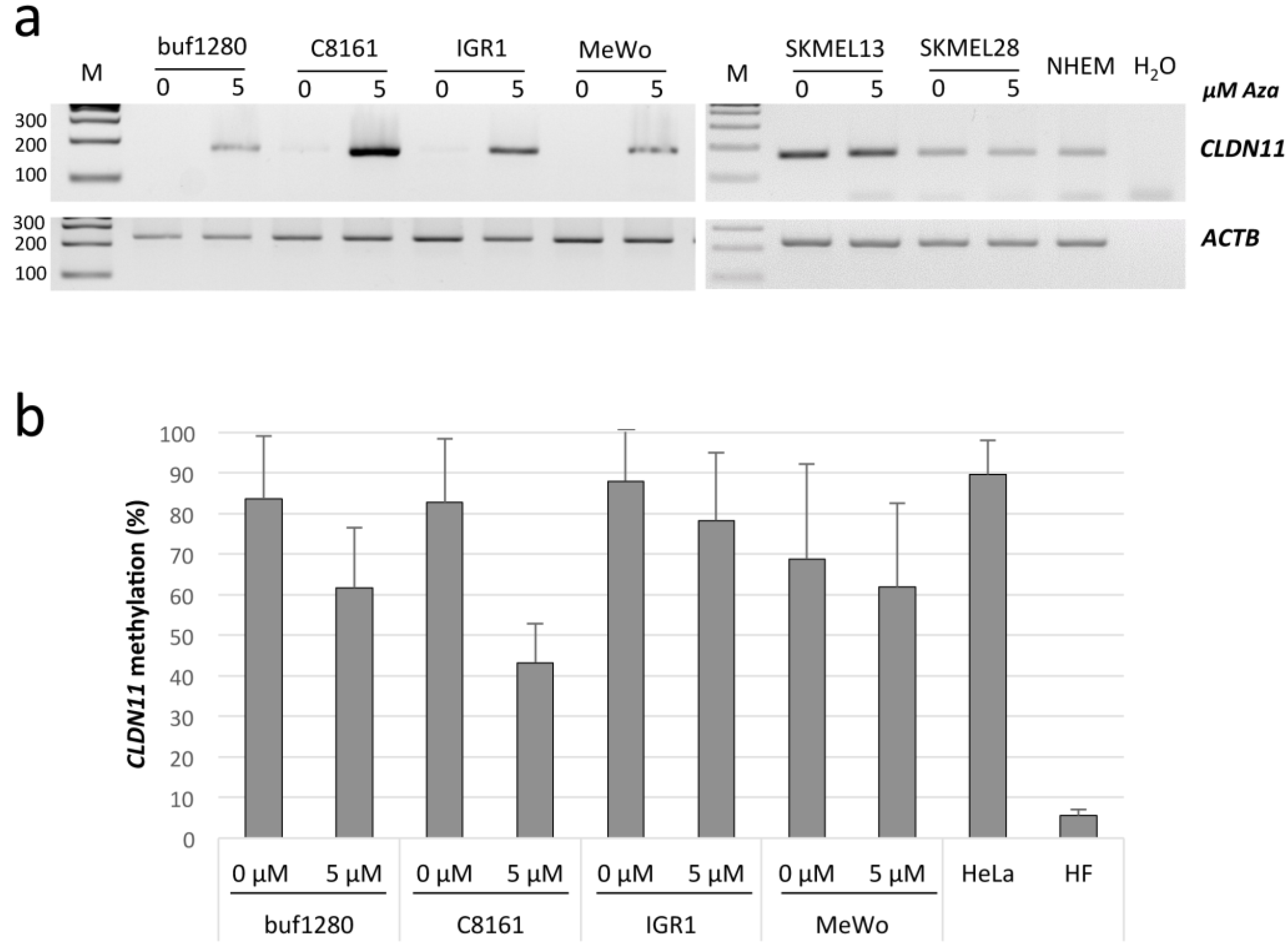{kind=link}
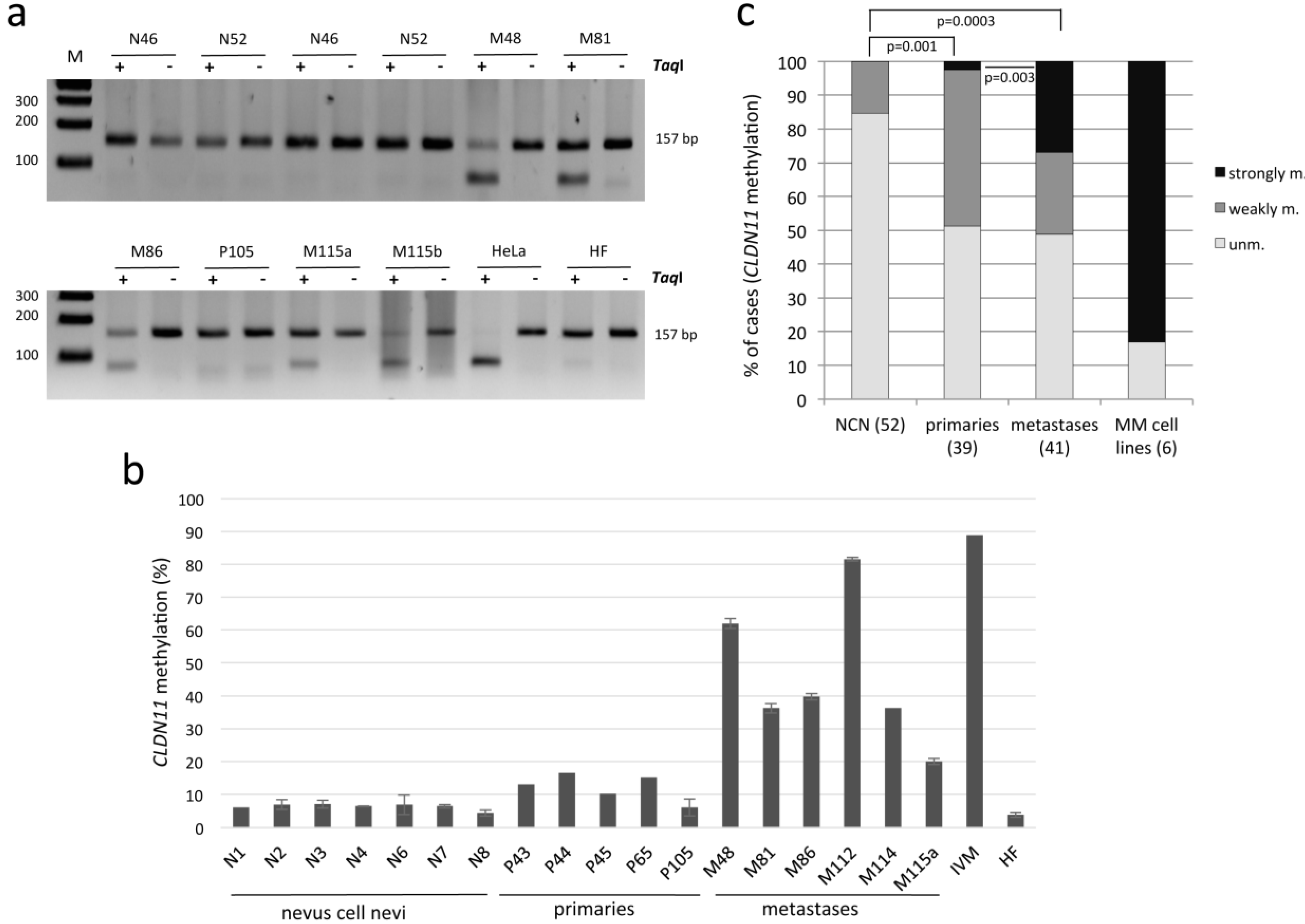{kind=link}
| Case | Unmethylated a | Weakly Methylated b | Strongly Methylated c |
|---|---|---|---|
| Nevus cell nevus | |||
| dysplastic (14) | 86% (12) | 14% (2) | 0% |
| Non-dysplastic (38) | 84% (32) | 16% (6) | 0% |
| Total (52) | 85% (44) | 15% (8) | 0% |
| Primary melanoma (39) | 51% (20) | 46% (18) | 3% (1) |
| Melanoma metastasis | |||
| Skin (19) | 21% (4) | 37% (7) | 42% (8) |
| Brain (16) | 68.75% (11) | 18.75% (3) | 12.5% (2) |
| Lymph node (4) | 75% (3) | 0% | 25% (1) |
| Others (2) | 100% (2) | 0% | 0% |
| Total (41) | 49% (20) | 24% (10) | 27% (11) |
| MM cell lines (6) | 17% (1) | 0% | 83% (5) |
3. Discussion
4. Experimental Section
4.1. Tissue and Cell Lines
4.2. Methylation Analysis
4.3. Expression Analysis
4.4. Statistical Evaluation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CLDN11 | Claudin 11 |
| COBRA | combined bisulfite restriction analysis |
| Aza | 5-Aza-2′-deoxycytidine |
| MM | malignant melanoma |
| NCN | nevus cell nevus |
| RASSF | Ras Association Domain Family |
| ACTB | Beta-Actin |
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Dahl, C.; Guldberg, P. The genome and epigenome of malignant melanoma. Apmis 2007, 115, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Mihm, M.C., Jr. Melanoma. N. Engl. J. Med. 2006, 355, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Rastetter, M.; Schagdarsurengin, U.; Lahtz, C.; Fiedler, E.; Marsch, W.; Dammann, R.; Helmbold, P. Frequent intra-tumoural heterogeneity of promoter hypermethylation in malignant melanoma. Histol. Histopathol. 2007, 22, 1005–1015. [Google Scholar] [PubMed]
- Lahtz, C.; Stranzenbach, R.; Fiedler, E.; Helmbold, P.; Dammann, R.H. Methylation of pten as a prognostic factor in malignant melanoma of the skin. J. Investig. Dermatol. 2010, 130, 620–622. [Google Scholar] [CrossRef] [PubMed]
- Helmbold, P.; Richter, A.M.; Walesch, S.; Skorokhod, A.; Marsch, W.; Enk, A.; Dammann, R.H. RASSF10 promoter hypermethylation is frequent in malignant melanoma of the skin but uncommon in nevus cell nevi. J. Investig. Dermatol. 2012, 132, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Baylin, S.B. The epigenomics of cancer. Cell 2007, 128, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, T.; Bosserhoff, A.K. Epigenetic events in malignant melanoma. Pigment Cell Res. 2007, 20, 92–111. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; van den Hurk, K.; Moerkerk, P.T.; Goeman, J.J.; Beck, S.; Gruis, N.A.; van den Oord, J.J.; Winnepenninckx, V.J.; van Engeland, M.; van Doorn, R. Promoter cpg island hypermethylation in dysplastic nevus and melanoma: Cldn11 as an epigenetic biomarker for malignancy. J. Investig. Dermatol. 2014, 134, 2957–2966. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, K.J.; Agarwal, R.; Morin, P.J. The claudin gene family: Expression in normal and neoplastic tissues. BMC Cancer 2006, 6, 186. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Anderson, J.M. Claudin interactions in and out of the tight junction. Tissue Barriers 2013, 1, e25247. [Google Scholar] [CrossRef] [PubMed]
- Jetten, A.M.; Suter, U. The peripheral myelin protein 22 and epithelial membrane protein family. Progr. Nucleic Acid Res. Mol. Biol. 2000, 64, 97–129. [Google Scholar]
- Mineta, K.; Yamamoto, Y.; Yamazaki, Y.; Tanaka, H.; Tada, Y.; Saito, K.; Tamura, A.; Igarashi, M.; Endo, T.; Takeuchi, K.; et al. Predicted expansion of the claudin multigene family. FEBS Lett. 2011, 585, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Mori, Y.; Cheng, Y.; Jin, Z.; Olaru, A.V.; Hamilton, J.P.; David, S.; Selaru, F.M.; Yang, J.; Abraham, J.M.; et al. Silencing of claudin-11 is associated with increased invasiveness of gastric cancer cells. PLoS ONE 2009, 4, e8002. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Smit, M.A.; van den Oord, J.J.; Goeman, J.J.; Verdegaal, E.M.; van der Burg, S.H.; Stas, M.; Beck, S.; Gruis, N.A.; Tensen, C.P.; et al. Genome-wide promoter methylation analysis identifies epigenetic silencing of mapk13 in primary cutaneous melanoma. Pigment Cell Melanoma Res. 2013, 26, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Mezzanotte, J.J.; Hill, V.; Schmidt, M.L.; Shinawi, T.; Tommasi, S.; Krex, D.; Schackert, G.; Pfeifer, G.P.; Latif, F.; Clark, G.J. Rassf6 exhibits promoter hypermethylation in metastatic melanoma and inhibits invasion in melanoma cells. Epigenetics 2014, 9, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Colombino, M.; Capone, M.; Lissia, A.; Cossu, A.; Rubino, C.; de Giorgi, V.; Massi, D.; Fonsatti, E.; Staibano, S.; Nappi, O.; et al. Braf/nras mutation frequencies among primary tumors and metastases in patients with melanoma. J. Clin. Oncol. 2012, 30, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Strunnikova, M.; Schagdarsurengin, U.; Kehlen, A.; Garbe, J.C.; Stampfer, M.R.; Dammann, R. Chromatin inactivation precedes de novo DNA methylation during the progressive epigenetic silencing of the rassf1a promoter. Mol. Cell Biol. 2005, 25, 3923–3933. [Google Scholar] [CrossRef] [PubMed]
- Di Cello, F.; Cope, L.; Li, H.; Jeschke, J.; Wang, W.; Baylin, S.B.; Zahnow, C.A. Methylation of the claudin 1 promoter is associated with loss of expression in estrogen receptor positive breast cancer. PLoS ONE 2013, 8, e68630. [Google Scholar] [CrossRef] [PubMed]
- Ogoshi, K.; Hashimoto, S.; Nakatani, Y.; Qu, W.; Oshima, K.; Tokunaga, K.; Sugano, S.; Hattori, M.; Morishita, S.; Matsushima, K. Genome-wide profiling of DNA methylation in human cancer cells. Genomics 2011, 98, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.; Roth, M.J.; Abnet, C.C.; Dawsey, S.P.; Qiao, Y.L.; Wang, G.Q.; Wei, W.Q.; Lu, N.; Dawsey, S.M.; Woodson, K. Promoter methylation in cytology specimens as an early detection marker for esophageal squamous dysplasia and early esophageal squamous cell carcinoma. Cancer Prev. Res. 2008, 1, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Yang, Y.D.; Fu, L.; Xu, W.; Liu, D.; Liang, Q.; Zhang, X.; Xu, L.; Guan, X.Y.; Wu, B.; et al. Cldn3 inhibits cancer aggressiveness via wnt-emt signaling and is a potential prognostic biomarker for hepatocellular carcinoma. Oncotarget 2014, 5, 7663–7676. [Google Scholar] [PubMed]
- Boireau, S.; Buchert, M.; Samuel, M.S.; Pannequin, J.; Ryan, J.L.; Choquet, A.; Chapuis, H.; Rebillard, X.; Avances, C.; Ernst, M.; et al. DNA-methylation-dependent alterations of claudin-4 expression in human bladder carcinoma. Carcinogenesis 2007, 28, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Fukushima, N.; Maitra, A.; Matsubayashi, H.; Yeo, C.J.; Cameron, J.L.; Hruban, R.H.; Goggins, M. Discovery of novel targets for aberrant methylation in pancreatic carcinoma using high-throughput microarrays. Cancer Res. 2003, 63, 3735–3742. [Google Scholar] [PubMed]
- Kim, B.; Kang, S.; Jeong, G.; Park, S.B.; Kim, S.J. Identification and comparison of aberrant key regulatory networks in breast, colon, liver, lung, and stomach cancers through methylome database analysis. PLoS ONE 2014, 9, e97818. [Google Scholar] [CrossRef] [PubMed]
- Kominsky, S.L.; Argani, P.; Korz, D.; Evron, E.; Raman, V.; Garrett, E.; Rein, A.; Sauter, G.; Kallioniemi, O.P.; Sukumar, S. Loss of the tight junction protein claudin-7 correlates with histological grade in both ductal carcinoma in situ and invasive ductal carcinoma of the breast. Oncogene 2003, 22, 2021–2033. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, S.; Smith, E.; de Young, N.J.; Wang, X.; Tian, Z.Q.; Liu, J.F.; Jamieson, G.G.; Drew, P.A. Methylation of cldn6, fbn2, rbp1, rbp4, tfpi2, and tmeff2 in esophageal squamous cell carcinoma. Oncol. Rep. 2009, 21, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Haag, T.; Herkt, C.E.; Walesch, S.K.; Richter, A.M.; Dammann, R.H. The apoptosis associated tyrosine kinase gene is frequently hypermethylated in human cancer and is regulated by epigenetic mechanisms. Genes Cancer 2014, 5, 365–374. [Google Scholar] [PubMed]
- Richter, A.M.; Walesch, S.K.; Wurl, P.; Taubert, H.; Dammann, R.H. The tumor suppressor rassf10 is upregulated upon contact inhibition and frequently epigenetically silenced in cancer. Oncogenesis 2012, 1, e18. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walesch, S.K.; Richter, A.M.; Helmbold, P.; Dammann, R.H. Claudin11 Promoter Hypermethylation Is Frequent in Malignant Melanoma of the Skin, but Uncommon in Nevus Cell Nevi. Cancers 2015, 7, 1233-1243. https://doi.org/10.3390/cancers7030834
Walesch SK, Richter AM, Helmbold P, Dammann RH. Claudin11 Promoter Hypermethylation Is Frequent in Malignant Melanoma of the Skin, but Uncommon in Nevus Cell Nevi. Cancers. 2015; 7(3):1233-1243. https://doi.org/10.3390/cancers7030834
Chicago/Turabian StyleWalesch, Sara K., Antje M. Richter, Peter Helmbold, and Reinhard H. Dammann. 2015. "Claudin11 Promoter Hypermethylation Is Frequent in Malignant Melanoma of the Skin, but Uncommon in Nevus Cell Nevi" Cancers 7, no. 3: 1233-1243. https://doi.org/10.3390/cancers7030834
APA StyleWalesch, S. K., Richter, A. M., Helmbold, P., & Dammann, R. H. (2015). Claudin11 Promoter Hypermethylation Is Frequent in Malignant Melanoma of the Skin, but Uncommon in Nevus Cell Nevi. Cancers, 7(3), 1233-1243. https://doi.org/10.3390/cancers7030834