
Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy



{kind=link}
{kind=link}
Abstract
:1. Introduction

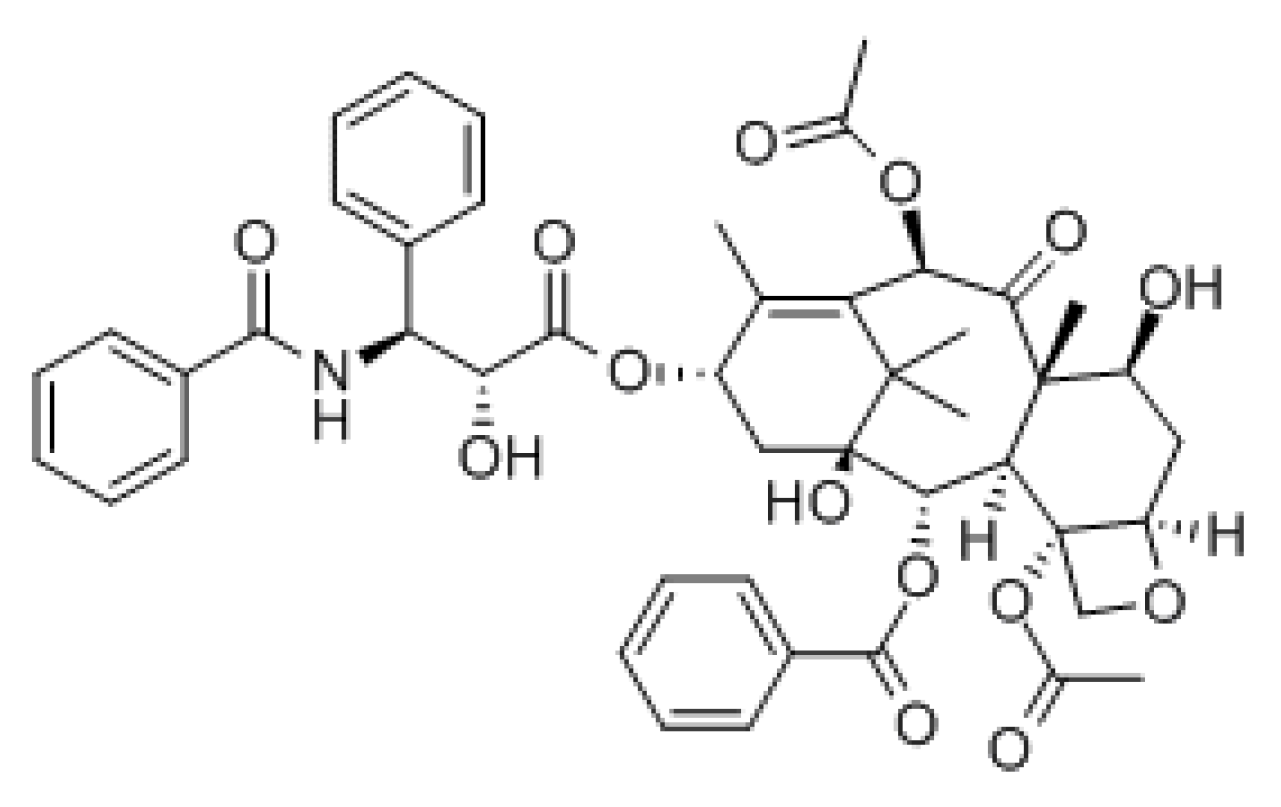
2. The Pharmacology of Paclitaxel
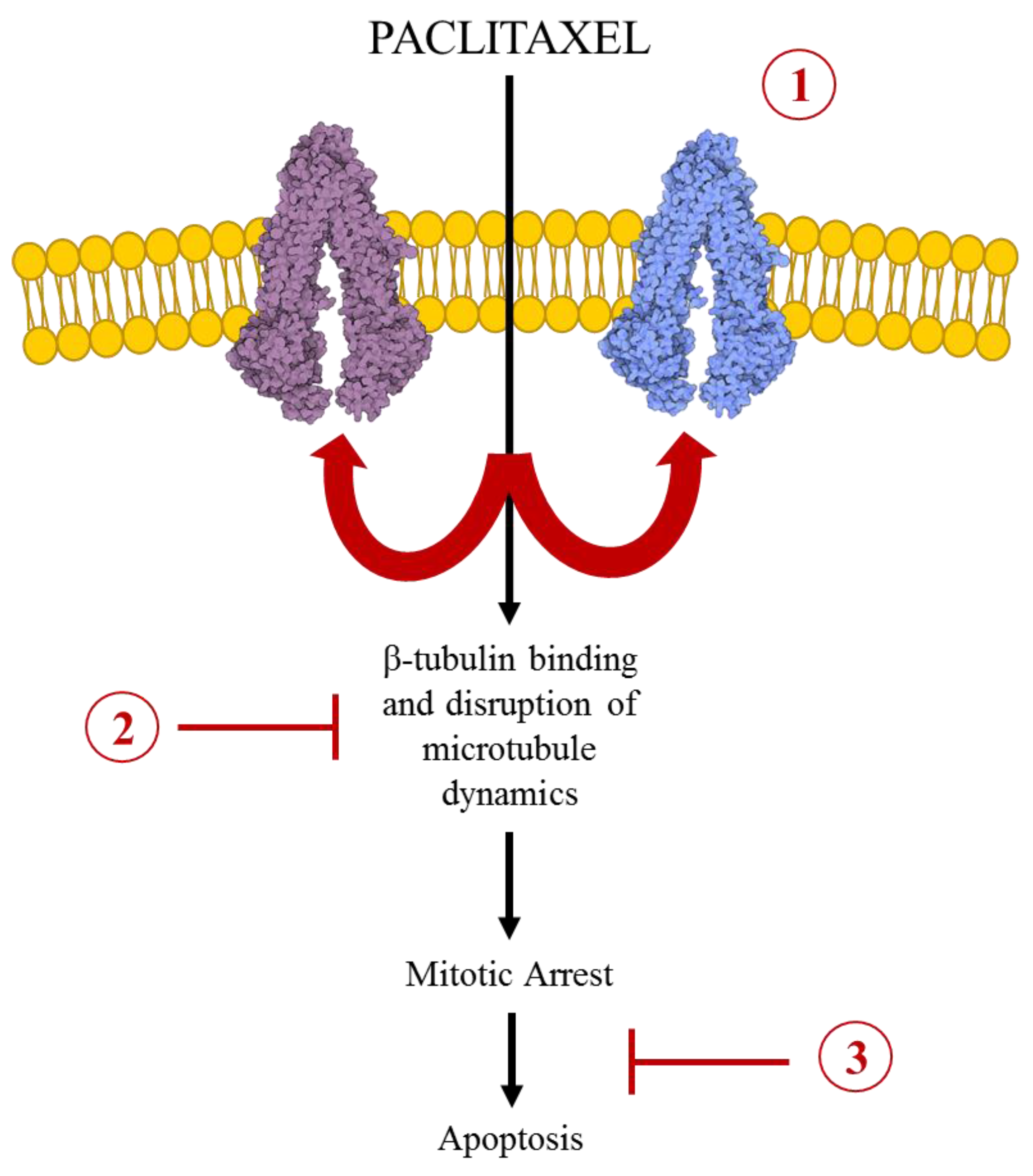
3. Paclitaxel in Drug Resistance

3.1. ABC Transport Mediated MDR in Paclitaxel Chemotherapy
3.2. Alterations of the Microtubule Dynamics in Paclitaxel Resistance
4. Paclitaxel in Chemoradiotherapy
5. Conclusions
Conflicts of Interest
References
- Wani, M.C.; Horwitz, S.B. Nature as a Remarkable Chemist: A personal story of the discovery and development of Taxol®. Anti-Cancer Drugs 2014, 25, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, S.B.; Lothstein, L.; Manfredi, J.J.; Mellado, W.; Parness, J.; Roy, S.N.; Schiff, P.B.; Sorbara, L.; Zeheb, R. Taxol: Mechanisms of action and resistance. Ann. N. Y. Acad. Sci. 1986, 466, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by Taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Rowinsky, E.K.; Cazenave, L.A.; Donehower, R.C. Taxol: A novel investigational antimicrotubule agent. J. Natl. Cancer Inst. 1990, 82, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L. Microtubules as drug receptors: Pharmacological properties of microtubule protein. Ann. N. Y. Acad. Sci. 1975, 253, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Ferlini, C.; Raspaglio, G.; Mozzetti, S.; Distefano, M.; Filippetti, F.; Martinelli, E.; Ferrandina, G.; Gallo, D.; Ranelletti, F.O.; Scambia, G. Bcl-2 down-regulation is a novel mechanism of paclitaxel resistance. Mol. Pharmacol. 2003, 64, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Fransson, M.; Green, H.; Litton, J.E.; Friberg, L.E. Influence of Cremophor EL and genetic polymorphisms on the pharmacokinetics of paclitaxel and its metabolites using a mechanism-based model. Drug Metab. Dispos. 2011, 39, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Hennenfent, K.L.; Govindan, R. Novel formulations of taxanes: A review. Old wine in a new bottle? Ann. Oncol. 2006, 17, 735–749. [Google Scholar]
- Li, Y.; Chen, N.; Palmisano, M.; Zhou, S. Pharmacologic sensitivity of paclitaxel to its delivery vehicles drives distinct clinical outcomes of paclitaxel formulations. Mol. Pharm. 2015, 12, 1308–1317. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, U.; Parchment, R.E.; Jasti, B.R.; Hussain, M. Taxanes: An overview of the pharmacokinetics and pharmacodynamics. Urology 1999, 54, 22–29. [Google Scholar] [CrossRef]
- Monsarrat, B.; Alvinerie, P.; Wright, M.; Dubois, J.; Gueritte-Voegelein, F.; Guenard, D.; Donehower, R.C.; Rowinsky, E.K. Hepatic metabolism and biliary excretion of Taxol in rats and humans. J. Natl. Cancer Inst. Monogr. 1993, 15, 39–46. [Google Scholar] [PubMed]
- Yamaguchi, H.; Hishinuma, T.; Endo, N.; Tsukamoto, H.; Kishikawa, Y.; Sato, M.; Murai, Y.; Hiratsuka, M.; Ito, K.; Okamura, C.; et al. Genetic variation in ABCB1 influences paclitaxel pharmacokinetics in Japanese patients with ovarian cancer. Int. J. Gynecol. Cancer 2006, 16, 979–985. [Google Scholar] [CrossRef] [PubMed]
- NIH National Cancer Institute. Cancer Treatment, Paclitaxel. Available online: http://www.cancer.gov/about-cancer/treatment/drugs/paclitaxel (accessed on 1 September 2015).
- Gottesman, M. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, R.Z.; Duan, Z.; Lamendola, D.E.; Penson, R.T.; Seiden, M.V. Paclitaxel Resistance: Molecular mechanisms and pharmacologic manipulation. Curr. Cancer Drug Targets 2003, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hari, M.; Loganzo, F.; Annable, T.; Tan, X.; Musto, S.; Morilla, D.B.; Nettles, J.H.; Snyder, J.P.; Greenberger, L.M. Paclitaxel-resistant cells have a mutation in the paclitaxel-binding region of beta-tubulin (Asp26Glu) and less stable microtubules. Mol. Cancer Ther. 2006, 5, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Juliano, R.L.; Ling, V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta 1976, 455, 152–162. [Google Scholar] [CrossRef]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Vallo, S.; Michaelis, M.; Rothweiler, F.; Bartsch, G.; Gust, K.M.; Limbart, D.M.; Rödel, F.; Wezel, F.; Haferkamp, A.; Cinatl, J. Drug-resistant urothelial cancer cell lines display diverse sensitivity profiles to potential second-line therapeutics. Transl. Oncol. 2015, 8, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Xie, J.D.; Chen, X.G.; Sim, H.M.; Zhang, X.; Liang, Y.J.; Singh, S.; Talele, T.T.; Sun, Y.; Ambudkar, S.V.; et al. Neratinib reverses ATP-binding cassette B1-mediated chemotherapeutic drug resistance in vitro, in vivo, and ex vivo. Mol. Pharmacol. 2012, 82, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Liang, Y.J.; Ruan, J.W.; Ding, Y.; Wang, X.W.; Shi, Z.; Gu, L.Q.; Yang, X.P.; Fu, L.W. Reversal of P-gp mediated multidrug resistance in vitro and in vivo by FG020318. J. Pharm. Pharmacol. 2004, 56, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Sodani, K.; Tiwari, A.K.; Dai, C.L.; Abuznait, A.H.; Patel, A.; Xiao, Z.J.; Ashby, C.R.; Kaddoumi, A.; Fu, L.W.; Chen, Z.S. Sildenafil enhances the anticancer activity of paclitaxel in an ABCB1-mediated multidrug resistance xenograft mouse model. J. Cancer Res. Updates 2014, 3, 169–173. [Google Scholar]
- Lampidis, T.J.; Krishan, A.; Planas, L.; Tapiero, H. Reversal of intrinsic resistance to adriamycin in normal cells by verapamil. Cancer Drug Deliv. 1986, 3, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.J.; Aihara, M.; Blume, K.G.; Sikic, B.I. Modulation of etoposide (VP-16) cytotoxicity by verapamil or cyclosporine in multidrug-resistant human leukemic cell lines and normal bone marrow. Exp. Hematol. 1990, 18, 1193–1198. [Google Scholar] [PubMed]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Lhomme, C.; Joly, F.; Walker, J.L.; Lissoni, A.A.; Nicoletto, M.O.; Manikhas, G.M.; Baekelandt, M.M.; Gordon, A.N.; Fracasso, P.M.; Mietlowski, W.L.; et al. Phase III study of valspodar (PSC 833) combined with paclitaxel and carboplatin compared with paclitaxel and carboplatin alone in patients with stage IV or suboptimally debulked stage III epithelial ovarian cancer or primary peritoneal cancer. J. Clin. Oncol. 2008, 26, 2674–2682. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.J.; Draper, D.; Chen, C.C.; Robey, R.W.; Figg, W.D.; Piekarz, R.L.; Chen, X.; Gardner, E.R.; Balis, F.M.; Venkatesan, A.M.; et al. A Pharmacodynamic study of docetaxel in combination with the P-glycoprotein antagonist, tariquidar (XR9576) in patients with lung, ovarian, and cervical cancer. Clin. Cancer Res. 2011, 17, 569–580. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Wei, M.J. Reversing multidrug resistance with tyrosine kinase inhibitors. Chin. J. Cancer 2012, 31, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Vergara, D.; Bellomo, C.; Zhang, X.; Vergaro, V.; Tinelli, A.; Lorusso, V.; Rinaldi, R.; Lvov, Y.M.; Leporatti, S.; Maffia, M. Lapatinib/Paclitaxel polyelectrolyte nanocapsules for overcoming multidrug resistance in ovarian cancer. Nanomedicine 2012, 8, 891–899. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.W.; Vogel, P.D.; Wise, J.G. Multiple drug transport pathways through human P‑glycoprotein. Biochemistry 2015, 54, 4374–4390. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R.; Chen, Z.S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updates 2015, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, S.; Kataoka, T.; Ohara, O.; Oishi, M.; Kuo, M.T.; Ishikawa, T. Human ATP-binding cassette transporter ABCC10: Expression profile and p53-dependent upregulation. J. Exp. Ther. Oncol. 2004, 4, 239–246. [Google Scholar] [PubMed]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.S. Multidrug resistance associated proteins in multidrug resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Hopper, E.; Belinsky, M.G.; Zeng, H.; Tosolini, A.; Testa, J.R.; Kruh, G.D. Analysis of the structure and expression pattern of MRP7 (ABCC10), a new member of the MRP subfamily. Cancer Lett. 2001, 162, 181–191. [Google Scholar] [CrossRef]
- Chen, Z.S.; Hopper-Borge, E.; Belinsky, M.G.; Shchaveleva, I.; Kotova, F.; Kruh, G.D. Characterization of the transport properties of human multidrug resistance protein 7 (MRP7, ABCC10). Mol. Pharmacol. 2003, 63, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Wang, Y.J.; Ashby, C.R.; Chen, Z.S. Recent advances regarding the role of ABC subfamily C member 10 (ABCC10) in the efflux of antitumor drugs. Chin. J. Cancer 2014, 33, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Oguri, T.; Ozasa, H.; Uemura, T.; Bessho, Y.; Miyazaki, M.; Maeno, K.; Maeda, H.; Sato, S.; Ueda, R. MRP7/ABCC10 expression is a predictive biomarker for the resistance to paclitaxel in non-small cell lung cancer. Mol. Cancer Ther. 2008, 7, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Sodani, K.; Chen, K.; Patel, A.; Abuznait, A.H.; Anreddy, N.; Sun, Y.L.; Kaddoumi, A.; Ashby, C.R.; Chen, Z.S. Masitinib antagonizes ATP-binding cassette subfamily C member 10-mediated paclitaxel resistance: A preclinical study. Mol. Cancer Ther. 2014, 13, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Hopper-Borge, E.; Chen, Z.S.; Shchaveleva, I.; Belinsky, M.G.; Kruh, G.D. Analysis of the drug resistance profile of multidrug resistance protein 7 (ABCC10): Resistance to docetaxel. Cancer Res. 2004, 64, 4927–4930. [Google Scholar] [CrossRef] [PubMed]
- Hopper-Borge, E.A.; Churchill, T.; Paulose, C.; Nicolas, E.; Jacobs, J.D.; Ngo, O.; Kuang, Y.; Grinberg, A.; Westphal, H.; Chen, Z.S.; Klein-Szanto, A.J.; Belinsky, M.G.; Kruh, G.D. Contribution of Abcc10 (Mrp7) to in vivo paclitaxel resistance as assessed in Abcc10(−/−) mice. Cancer Res. 2011, 71, 3649–3657. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Dai, C.L.; Chen, J.J.; Kathawala, R.J.; Sun, Y.L.; Chen, H.F.; Fu, L.W.; Chen, Z.S. Tandutinib (MLN518) reverses multidrug resistance by inhibiting the efflux activity of the multidrug resistance protein 7 (ABCC10). Oncol. Rep. 2013, 29, 2479–2485. [Google Scholar] [PubMed]
- Kathawala, R.J.; Wei, L.; Anreddy, N.; Chen, K.; Patel, A.; Alqahtani, S.; Zhang, Y.K.; Wang, Y.J.; Sodani, K.; Kaddoumi, A.; Ashby, C.R.; Chen, Z.S. The small molecule tyrosine kinase inhibitor NVP-BHG712 antagonizes ABCC10-mediated paclitaxel resistance: A preclinical and pharmacokinetic study. Oncotarget 2015, 6, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.; Kirschner, M. Dynamic instability of microtubule growth. Nature 1984, 312, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Cabral, F. New insights into mechanisms of resistance to microtubule inhibitors. Biochim. Biophys. Acta 2011, 1816, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Yang, H.; Cabral, F. Paclitaxel dependent cell lines reveal a novel drug activity. Mol. Cancer Ther. 2010, 9, 2914–2923. [Google Scholar] [CrossRef] [PubMed]
- Kamath, K.; Wilson, L.; Cabral, F.; Jordan, M.A. βIII-Tubulin induces paclitaxel resistance in association with reduced effects on microtubule dynamic instability. J. Biol. Chem. 2005, 280, 12902–12907. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Cabral, F. Molecular basis for class V beta-tubulin effects on microtubule assembly and paclitaxel resistance. J. Biol. Chem. 2009, 284, 13023–13032. [Google Scholar] [CrossRef] [PubMed]
- Orr, G.A.; Verdier-Pinard, P.; McDaid, H.; Horwitz, S.B. Mechanisms of Taxol resistance related to microtubules. Oncogene 2003, 22, 7280–7295. [Google Scholar] [CrossRef] [PubMed]
- Schibler, M.; Cabral, F. Taxol-dependent mutants of Chinese hamster ovary cells with alterations in α- and β-tubulin. J. Cell Biol. 1986, 102, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Martello, L.A.; Verdier-Pinard, P.; Shen, H.J.; He, L.; Torres, K.; Orr, G.A.; Horwitz, S.B. Elevated levels of microtubule destabilizing factors in a Taxol-resistant/dependent A549 cell line with an alpha-tubulin mutation. Cancer Res. 2003, 63, 1207–1213. [Google Scholar] [PubMed]
- Ong, V.; Liem, N.L.; Schmid, M.A.; Verrills, N.M.; Papa, R.A.; Marshall, G.M.; Mackenzie, K.L.; Kavallaris, M.; Lock, R.B. A role for altered microtubule polymer levels in vincristine resistance of childhood acute lymphoblastic leukemia xenografts. J. Pharmacol. Exp. Ther. 2008, 324, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Monzó, M.M.; Rosell, R.; Sánchez, J.J.; Lee, J.S.; O’Brate, A.; González-Larriba, J.L.; Alberola, V.; Lorenzo, J.C.; Núñez, L.; Ro, J.Y.; et al. Paclitaxel resistance in non-small-cell lung cancer associated with beta-tubulin gene mutations. J. Clin. Oncol. 1999, 17, 1786–1793. [Google Scholar] [PubMed]
- Rath, O.; Kozielski, F. Kinesins and cancer. Nat. Rev. Cancer 2012, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Yang, H.; Pedroza, M.; Bhattacharya, R.; Cabral, F. Mitotic centromere-associated kinesin (MCAK) mediates paclitaxel resistance. J. Biol. Chem. 2011, 286, 36378–36384. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Yang, H.; Cabral, F. Overexpression of mitotic centromere associated kinesin stimulates microtubule detachment and confers resistance to paclitaxel. Mol. Cancer Ther. 2011, 10, 929–937. [Google Scholar] [CrossRef]
- Terasima, T.; Tolmach, L.J. X-ray sensitivity and DNA synthesis in synchronous populations of HeLa cells. Science 1963, 140, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Tishler, R.B.; Schiff, P.B.; Geard, C.R.; Hall, E.J. Taxol: A novel radiation sensitizer. Int. J. Radiat. Oncol. Biol. Phys. 1992, 22, 613–617. [Google Scholar] [CrossRef]
- Lokeshwar, B.L.; Ferrell, S.M.; Block, N.L. Enhancement of radiation response of prostatic carcinoma by taxol: Therapeutic potential for late-stage malignancy. Anticancer Res. 1995, 15, 93–98. [Google Scholar] [PubMed]
- Cividalli, A.; Arcangeli, G.; Cruciani, G.; Livdi, E.; Cordelli, E.; Danesi, D.T. Enhancement of radiation response by paclitaxel in mice according to different treatment schedules. Int. J. Radiat. Oncol. Biol. Phys. 1998, 40, 1163–1170. [Google Scholar] [CrossRef]
- Erlich, E.; McCall, A.; Potkul, R.; Walter, S.; Vaughan, A. Paclitaxel is only a weak radiosensitizer of human cervical carcinoma cell lines. Gynecol. Oncol. 1996, 60, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Sevin, B.U.; Perras, J.; Nguyen, H.N.; Pham, C.; Steren, A.J.; Koechli, O.R.; Averette, H.E. Paclitaxel: A radiation sensitizer of human cervical cancer cells. Gynecol. Oncol. 1995, 57, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Formenti, S.C.; Volm, M.; Skinner, K.A.; Spicer, D.; Cohen, D.; Perez, E.; Bettini, A.C.; Groshen, S.; Gee, C.; Florentine, B.; et al. Preoperative twice-weekly paclitaxel with concurrent radiation therapy followed by surgery and postoperative doxorubicin-based chemotherapy in locally advanced breast cancer: A phase I/II trial. J. Clin. Oncol. 2003, 21, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Liebmann, J.; Cook, J.A.; Fisher, J.; Teague, D.; Mitchell, J.B. In vitro studies of Taxol as a radiation sensitizer in human tumor cells. J. Natl. Cancer Inst. 1994, 86, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Sunwoo, J.B.; Herscher, L.L.; Kroog, G.S.; Thomas, G.R.; Ondrey, F.G.; Duffey, D.C.; Solomon, B.I.; Boss, C.; Albert, P.S.; McCullugh, L.; et al. Concurrent paclitaxel and radiation in the treatment of locally advanced head and neck cancer. J. Clin. Oncol. 2001, 19, 800–811. [Google Scholar] [PubMed]
- Zhang, Y.; Yang, Z.; Zhou, Y.; Pan, J.; Liu, Y. Efficacy of concurrent single-agent chemotherapy using radiotherapy in patients with cervical cancer: A meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 8661–8673. [Google Scholar] [PubMed]
- Rowinsky, E.K.; Donehower, R.C. Paclitaxel (taxol). N. Engl. J. Med. 1995, 332, 1004–1014. [Google Scholar] [PubMed]
- Hanna, Y.M.; Baglan, K.L.; Stromberg, J.S.; Vicini, F.A.; Decker, D. Acute and subacute toxicity associated with concurrent adjuvant radiation therapy and paclitaxel in primary breast cancer therapy. Breast J. 2002, 8, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, T.; Matsumura, Y.; Suzuki, M.; Shimizu, K.; Goda, R.; Nakamura, I.; Nakatomi, I.; Yokoyama, M.; Kataoka, K.; Kakizoe, T. NK105, a paclitaxel-incorporating micellar nanoparticle formulation, can extend in vivo antitumour activity and reduce the neurotoxicity of paclitaxel. Br. J. Cancer 2005, 92, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; van der Meel, R.; Theek, B.; Blenke, E.O.; Pieters, E.H.; Fens, M.H.; Ehling, J.; Schiffelers, R.M.; Storm, G.; van Nostrum, C.F.; et al. Complete regression of xenograft tumors upon targeted delivery of paclitaxel via Π–Π stacking stabilized polymeric micelles. ACS Nano 2015, 9, 3740–3752. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yeo, Y. Extracellular stability of nanoparticulate drug carriers. Arch. Pharm. Res. 2014, 37, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Koizumi, F.; Uchino, H.; Kuroda, J.; Kawaguchi, T.; Naito, S.; Matsumura, Y. NK105, a paclitaxel-incorporating micellar nanoparticle, is a more potent radiosensitising agent compared to free paclitaxel. Br. J. Cancer 2006, 95, 601–606. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbuti, A.M.; Chen, Z.-S. Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy. Cancers 2015, 7, 2360-2371. https://doi.org/10.3390/cancers7040897
Barbuti AM, Chen Z-S. Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy. Cancers. 2015; 7(4):2360-2371. https://doi.org/10.3390/cancers7040897
Chicago/Turabian StyleBarbuti, Anna Maria, and Zhe-Sheng Chen. 2015. "Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy" Cancers 7, no. 4: 2360-2371. https://doi.org/10.3390/cancers7040897
APA StyleBarbuti, A. M., & Chen, Z.-S. (2015). Paclitaxel Through the Ages of Anticancer Therapy: Exploring Its Role in Chemoresistance and Radiation Therapy. Cancers, 7(4), 2360-2371. https://doi.org/10.3390/cancers7040897