1. Introduction
We have previously shown that α-lactalbumin vaccination mediates protection against the development of murine breast cancer in the absence of any detectable inflammatory changes in all normal non-lactating tissues examined [
1]. Based on these results, we have proposed that vaccination of healthy, cancer-free, adult women against α-lactalbumin may provide safe and effective immunoprevention of breast cancer [
2]. Since all current prophylactic vaccines target pathogens, several unique issues inherently arise regarding the overall feasibility of applying an autoimmune strategy for primary immunoprevention of breast cancer [
3]. These issues include the following: (1) whether adult women would be immunologically responsive to human α-lactalbumin, (2) whether a history of lactation would create an insurmountable tolerance that would preclude generating effective immunity against α-lactalbumin, (3) whether α-lactalbumin is immunologically available in human breast tumors, and (4) whether expression of α-lactalbumin in normal non-breast tissues would predispose to systemic autoimmune complications.
Here we provide experimental data that directly address these concerns. We found that in vitro priming of peripheral blood mononuclear cells (PBMC) from healthy adult women results in frequencies of α-lactalbumin-specific proinflammatory T cells consistent with those associated with protection against murine breast tumors. We also found that the frequencies of T cells secreting interferon-gamma (IFNγ) and the level of protection from the development of breast tumors are virtually identical whether α-lactalbumin vaccination occurs in parous mice with a history of lactation and breastfeeding or in non-parous mice with no such history.
In addition, we experimentally confirmed the results of several database searches indicating highly significant overexpression of α-lactalbumin in human triple negative breast cancer (TNBC) [
4,
5]. This confirmation involved several experimental approaches including RT-PCR, Western blot, and immunohistochemical analysis of human TNBC tissues, as well as longitudinal visualization of α-lactalbumin gene expression during
in vivo growth of human TNBC in immunodeficient mice. This
in vivo visualization of α-lactalbumin gene expression was facilitated by measuring bioluminescence from growing human HCC1937 TNBC cells stably transfected with a lentivirus designed to express firefly luciferase under regulation of the human α-lactalbumin promoter. Finally, we found substantial published evidence indicating negative immunohistochemical staining for α-lactalbumin in 78 normal human tissues examined, thereby confirming the widely held view that α-lactalbumin expression in normal human tissues is confined exclusively to the lactating breast [
6].
Thus far, our studies have resulted in the issuance of two US patents [
7,
8]. Collectively, our results support the view that α-lactalbumin vaccination has substantial potential for providing therapy against TNBC recurrence as well as primary immmunoprevention of TNBC, the most aggressive form of breast cancer and the most common variant occurring in women with
BRCA1 mutations, a high risk population with the greatest need for primary immunoprevention [
9,
10].
3. Discussion
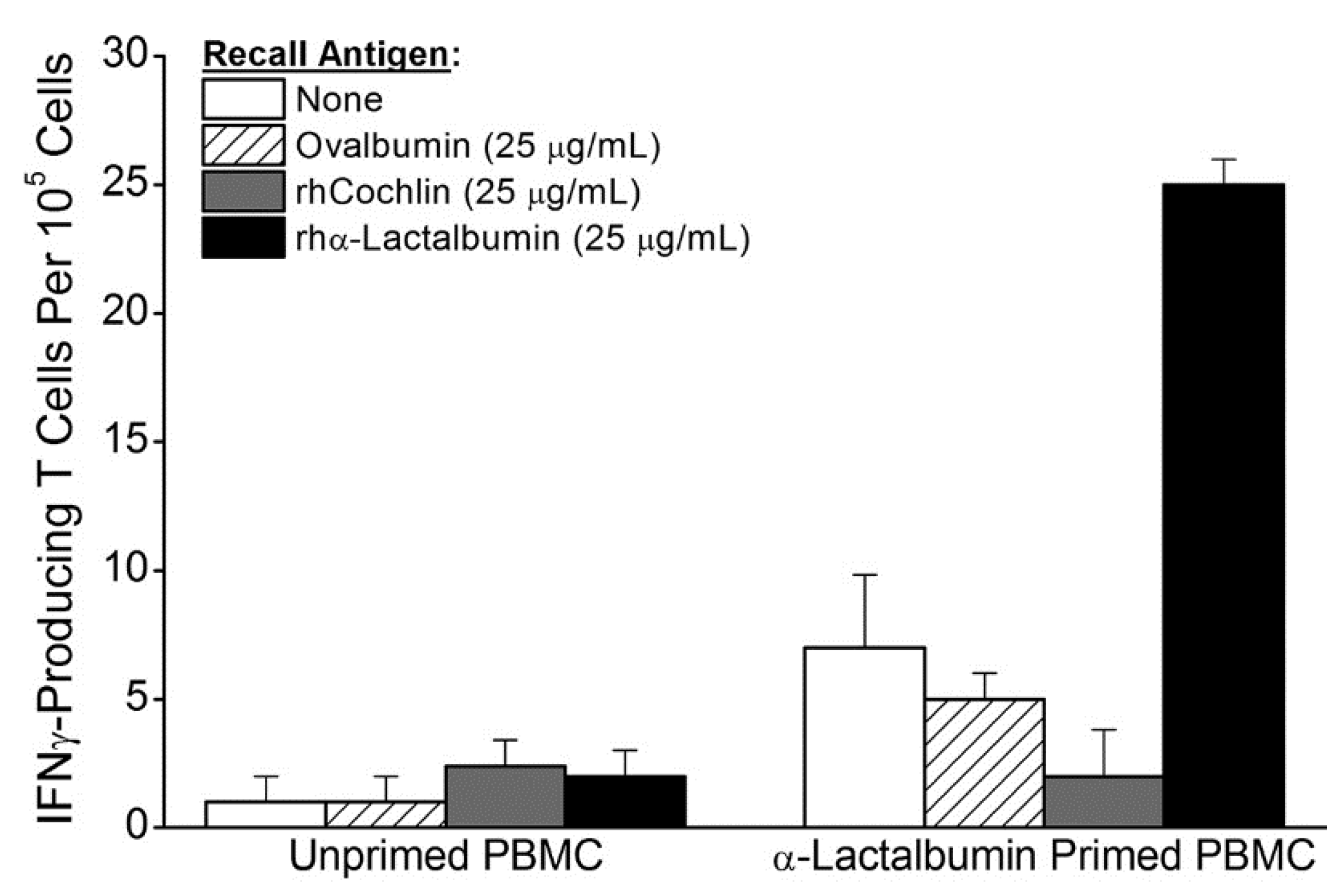
In the current study we show that adult women have an available proinflammatory T cell repertoire capable of responding to rhα-lactalbumin at frequencies that are comparable to those observed following active immunization of mice with rmα-lactalbumin in CFA [
1]. The observed frequency of 1 per 20,000 α-lactalbumin-specific type-1 proinflammatory T cells induced by
in vitro priming is a frequency comparable to those obtained
in vivo and shown to induce effective breast tumor immunity following active α-lactalbumin immunization of mice (
Figure 2a) [
1]. Thus, α-lactalbumin appears to be sufficiently immunogenic to induce effective tumor immunity in women. Moreover, if α-lactalbumin vaccination occurs in combination with an adjuvant capable of orchestrating a robust adaptive proinflammatory T cell response, any inherent immune tolerance to this self-protein will likely be overcome. Such adjuvants include CFA for use in animal experimentation and other adjuvants more suitable for human vaccination including the many DNA vaccines that induce IFNγ through CpG-Toll-like receptor 9 ligand interactions.
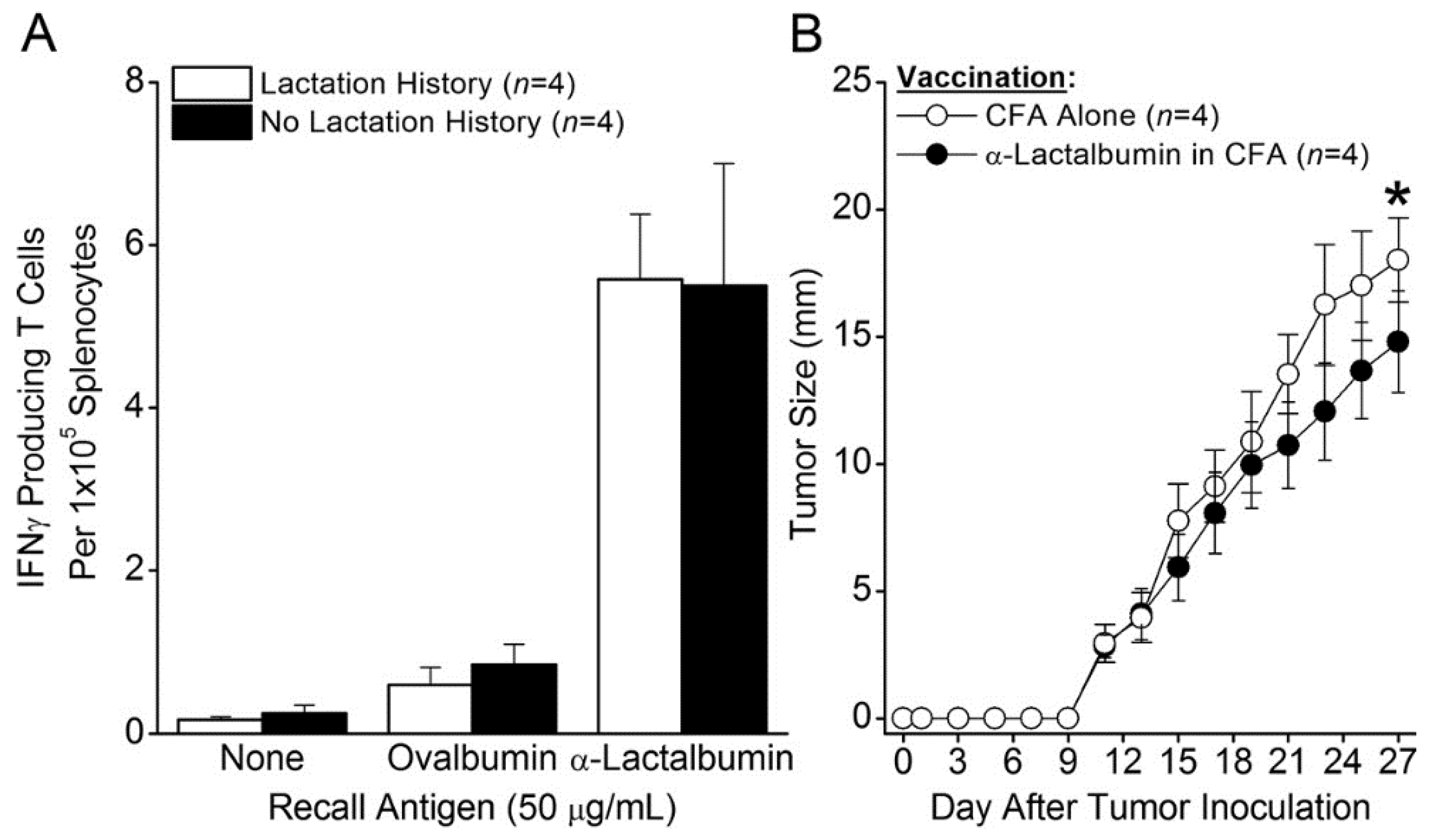
The likelihood of inducing proinflammatory immunity in women vaccinated with α-lactalbumin was further reassured when α-lactalbumin vaccinated parous mice with an established history of lactation and breastfeeding showed frequencies of proinflammatory T cells and tumor immunity indistinguishable from those occurring in non-parous mice with no such history. This outcome indicates that the sudden burst exposure of the adult immune system to massive levels of α-lactalbumin through the process of lactation does not irreversibly tolerize. These results indicate the feasibility of inducing a clinically relevant proinflammatory immune response to α-lactalbumin as long as the protein is properly and efficiently presented to the adult immune system.
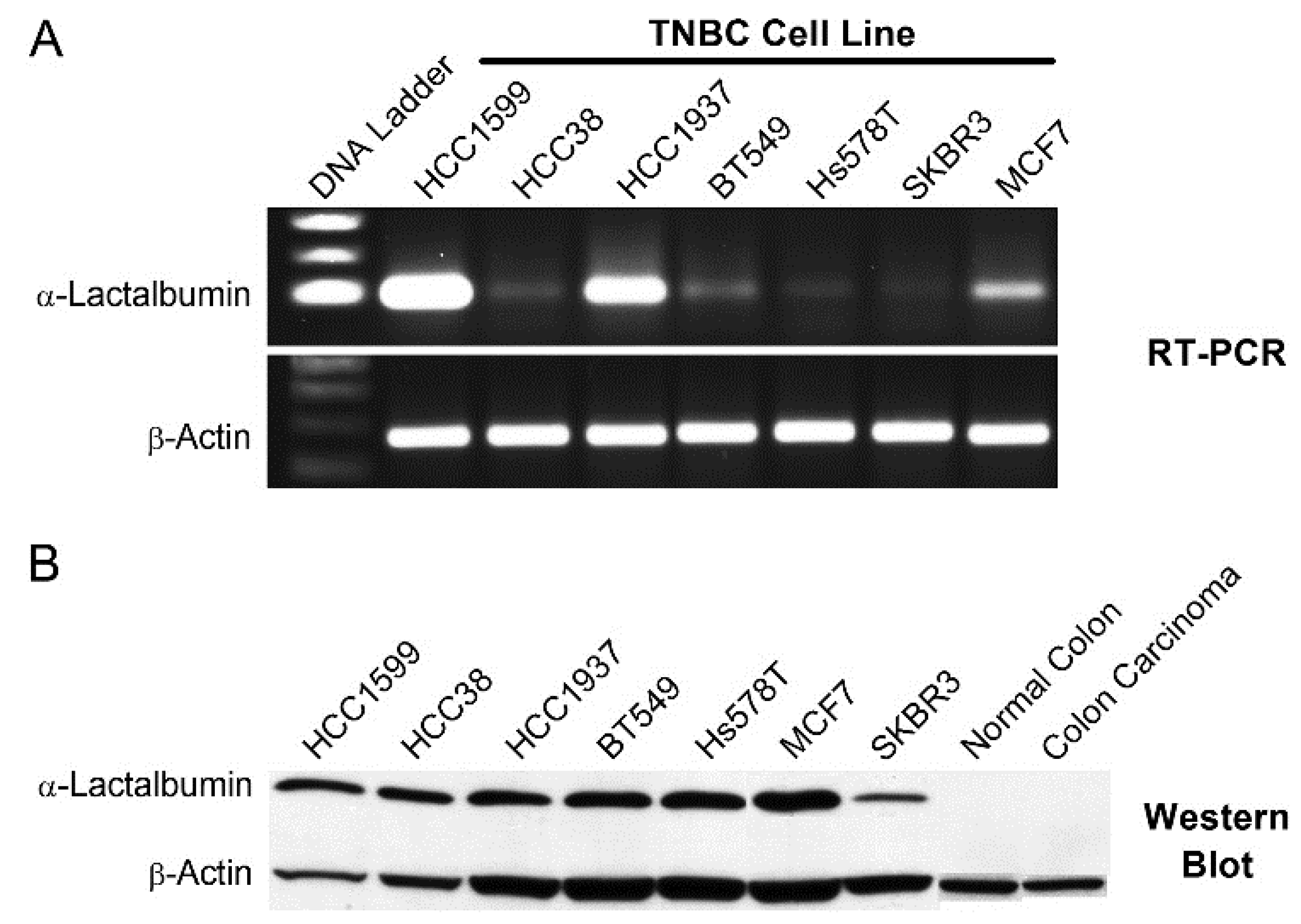
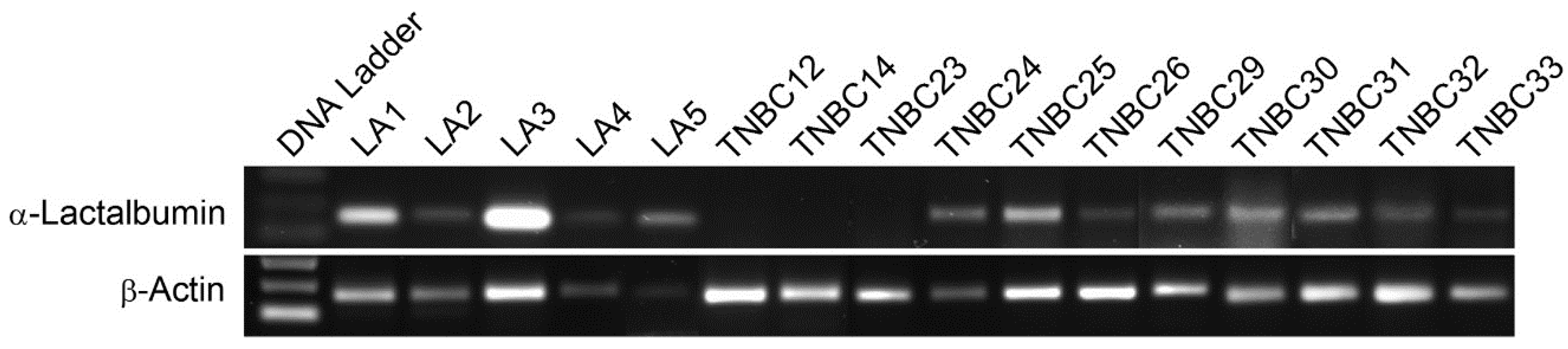
We also found that α-lactalbumin gene expression occurs at substantial levels in the vast majority of human TNBC cell lines and in 72% of primary TNBC tissues examined. Moreover, the levels of α-lactalbumin gene expression occurring in TNBC cell lines and tumor tissues was similar to the levels detected in human lactating adenomas and correlated with Western blot detection of α-lactalbumin protein. In addition, we found that the α-lactalbumin gene is expressed constitutively and stably during both the early and later stages of TNBC tumor growth in vivo. Thus, the availability of α-lactalbumin protein in most human TNBC tumors makes it a suitable vaccine target for immunotherapy and primary immunoprevention of human TNBC.
It is well-established that α-lactalbumin gene and protein expression in normal tissues is confined to the breast during late pregnancy and lactation and is not detectable in any of 78 other normal human tissues examined [
6]. Thus, it seems likely that this highly confined tissue expression precludes any systemic autoimmune complications in women vaccinated with α-lactalbumin. Thus, we believe that α-lactalbumin vaccination may be both safe and effective in preventing TNBC in women with
BRCA1 mutations who often have >80% lifetime risk for developing breast cancer with TNBC being the most common variant of this disease occurring in this high risk population [
9,
10].
Although it is not completely understood why α-lactalbumin is overexpressed in so many TNBC tumors, one explanation may be related to the loss of default inhibition that homeostatic levels of estrogen and progesterone signaling provide. Indeed, the rapid drop at parturition in production of estrogen, and perhaps more importantly progesterone, removes the impact of their homeostatic default inhibition of prolactin transcription that serves as the primary form of constitutive regulation of α-lactalbumin production [
11,
12,
13,
14,
15]. Thus, we propose that breast tumors with deficient constitutive estrogen or progesterone signaling may be unable to provide sufficient default transcriptional inhibition of critical lactation proteins like α-lactalbumin. Experiments are ongoing to test this hypothesis.
Finally, it is worth reflecting on the aggressive nature of human TNBC tumors. This aggressiveness may best be appreciated by the ability of human TNBC tumors to grow in the complete absence of cognate signaling by the potent breast cancer growth factors, estrogen and progesterone. It seems ironic that the acquisition of such a prominent evolutionary advantage to grow without any need for readily available growth factors occurs with a tradeoff that the tumor may no longer be capable of sufficiently inhibiting α-lactalbumin synthesis, a tradeoff that may be exploited using α-lactalbumin targeted immune strategies.
4. Materials and Methods
4.1. Generation of Recombinant Human (rh) and Recombinant Mouse (rm) Proteins
The open reading frame cDNA nucleotide sequence for mouse α-lactalbumin (NCBI Reference Sequence: NM_010679.1) and human α-lactalbumin (NCBI Reference Sequence: NM_002289.2) were modified to ensure optimized protein folding and production in prokaryotic expression systems by substituting mammalian codons with more efficient prokaryotic sequences (Dapcel, Cleveland, OH, USA). The optimized DNA sequence was synthesized
de novo and inserted into the NdeI-Bam HI sites of pET-3a expression vector (GeneArt AG, Regensburg, Germany, Darmstadt, Germany) thereby providing C-terminal 6xHis-tagged recombinant proteins. Plasmids containing these inserts were transformed in
E. coli (Lucigen, Middleton, WI, USA). High level expression colonies were selected following induction with isopropyl β-D-1-thiogalactopyranoside (IPTG; Amresco, Solon, OH, USA) and were sequenced for confirming proper orientation and alignment. The 6xHis-tagged proteins were purified under denaturing conditions using nickel-nitrilo triacetic acid (Ni-NTA) affinity chromatography (Qiagen Sciences, Germantown, MD, USA). Prior to use, each protein was further purified by reverse phase high performance liquid chromatography (HPLC) to yield endotoxin-free protein [
16]. Levels of endotoxin were <0.05 endotoxin units (<5 pg) per mg of recombinant protein.
4.2. In Vitro Priming of Human PBMC to rhα-Lactalbumin
Monocyte derived dendritic cells (DCs) were prepared from PBMC taken from a healthy, cancer-free, adult 29 year-old woman. Adherent cell selection was followed by culture in X-VIVO media (BioWhittaker, Walkersville, MD, USA) with 50 µg/mL rhGMCSF and rhIL-4 (Peprotech, Rocky Hill, NJ, USA). Six days after initiation of culture, DCs were pulsed with 75 µg/mL of purified rhα-lactalbumin, washed extensively 48 h later, and co-cultured with nylon wool purified naive T cells from the same donor at a ratio of 1:5 (DCs to T cells). After 72 h of co-culture, in vitro primed T cells and unprimed T cells from the same donor were enriched by passage through nylon wool and re-cultured in duplicate or triplicate with γ-irradiated (30 Gy) PBMC as feeder cells at a ratio of 1:10 (feeders to T cells) with 25 µg/mL rhα-lactalbumin, rhcochlin, or ovalbumin on ELISPOT plates (EMD Millipore, Billerica, MA, USA) pre-coated with mouse anti-human IFNγ capture antibody (Clone MD-1, eBioscience, San Diego, CA, USA). Frequencies of α-lactalbumin-responsive IFNγ-producing T cells were determined 48 h later using secondary biotinylated mouse anti-human IFNγ (Clone 4S.B3, eBioscience, San Diego, CA, USA). Final frequencies were determined by subtracting spots obtained in wells with 25 µg/mL ovalbumin from spots obtained in wells containing 25 µg/mL rhα-lactalbumin. Recall responses from unprimed T cells were consistently negligible.
4.3. Mice and Immunization
Eight week old BALB/cJ female mice were purchased commercially (Jackson Laboratory, Bar Harbor, ME, USA) and housed in microisolator cages in the Biologic Resources Unit of the Lerner Research Institute, Cleveland Clinic. Some mice were allowed to complete one full cycle of pregnancy, lactation, breastfeeding, and weaning prior to immunization with α-lactalbumin. Four weeks after weaning, parous female mice and age-matched non-parous female mice with no prior lactation history were immunized by subcutaneous injection in the abdominal flank with 100 μg rmα-lactalbumin in 200 μL of an emulsion of equal volumes of water and CFA containing 200 µg Mycobacteria tuberculosis H37Ra (Difco, Detroit, MI, USA). All protocols were pre-approved by the Institutional Animal Care and Use Committee of the Cleveland Clinic.
4.4. ELISPOT Analysis in Mice
Four weeks after immunization of mice with rmα-lactalbumin, splenocyte frequencies of type-1 IFNγ-producing T cells were determined by ELISPOT analysis. Duplicate or triplicate wells containing 2 × 105 splenocytes were cultured with 50 µg/mL rmα-lactalbumin or ovalbumin in ELISPOT plates (Millipore, Billerica, MA, USA) pre-coated with capture IFNγ antibody (clone AN-18; eBioscience, San Diego, CA, USA) in 200 μL/well total culture volume in DMEM (Mediatech, Manassas, VA, USA) supplemented with L-glutamine, penicillin, streptomycin, HEPES buffer (Invitrogen Life Technologies, Grand Island, NY, USA), and 10% fetal bovine serum (Hyclone, Logan, UT, USA). At 72 h of culture, wells were treated with biotinylated capture IFNγ antibody (clone R4-6A2; eBioscience, San Diego, CA, USA), and after overnight incubation and washing, spots were visualized by sequential treatment with alkaline phosphatase-conjugated streptavidin and 5-bromo-4-chloro-3-indolyl phosphate substrate (R&D Systems, Minneapolis, MN, USA). The reaction was halted after 10 minutes by repeated washing with double-distilled deionized H2O, and spots were developed and counted using an ImmunoSpot S6 analyzer with proprietary ImmunoSpot 5.1 software (Cellular Technologies Limited, Shaker Heights, OH, USA).
4.5. Evaluation of Mouse Breast Tumor Growth
Four weeks after weaning, parous mice were immunized with rmα-lactalbumin in CFA or with CFA alone as described above. Three weeks after vaccination, mice were inoculated with 1 × 104 4T1 breast cancer cells purchased commercially from the American Tissue Culture Collection (ATCC CRL-2539; Manassas, VA, USA) by subcutaneous injection in 100 µL PBS. Tumors were measured by Vernier caliper every other day until tumors reached 17 mm in any direction at which time all mice were humanely euthanized. Significance in tumor growth between groups was determined by analysis of variants (ANOVA).
4.6. α-Lactalbumin Gene Expression in Human TNBC
For analyzing gene expression in human tissues, RNA was extracted from 10 µm microdissected sections of formalin-fixed paraffin embedded human TNBC or lactating adenoma tissue blocks. For analyzing gene expression in human cell lines, RNA was extracted from human TNBC cell lines [
17] obtained commercially including: HCC1599 (ATCC CRL-2331), HCC38 (ATCC CRL-2314), HCC1937 (ATCC CRL-2336), BT549 (ATCC HTB122), and Hs578T (ATCC HTB-126). Following reverse transcription, 35 cycles of gene-specific amplification were performed using purchased human α-lactalbumin and β-actin specific primer pairs (Applied Biosystems, Foster City, CA, USA) and the amplified products were electrophoresed on a 1.5% agarose gel.
4.7. α-Lactalbumin Protein Detection in Human TNBC
For Western blot detection of α-lactalbumin protein in human TNBC cell lines, cell lysates were electrophoresed on a 15% SDS polyacrylamide gel. Gels were blotted onto a polyvinylidene fluoride (PVDF) membrane and probed with a rabbit polyclonal antibody specific for human α-lactalbumin (#HPA029856; Sigma-Aldrich, St. Louis, MO, USA) and mouse monoclonal antibody for human β-actin (clone AC-15; Sigma-Aldrich, St. Louis, MO, USA) at dilutions of 1:100 and 1:8,000 respectively. Visualization of immunoreactivity was facilitated using horse radish peroxidase (HRP)-conjugated goat anti-rabbit or goat anti-mouse IgG (Southern Biotech, Birmingham, AL, USA). For immunohistochemical analysis, formalin-fixed paraffin embedded tissue sections at 5 μm underwent antigen retrieval in citrate buffer and were subsequently treated with a 1:100 dilution of the rabbit polyclonal antibody specific for human α-lactalbumin (#HPA029856; Sigma-Aldrich, St. Louis, MO, USA) followed by detection with HRP-conjugated goat anti-rabbit IgG (#4050-05; Southern Biotech, Birmingham, AL, USA).
4.8. Visualization of Human α-Lactalbumin Gene Expression during in vivo Growth of TNBC
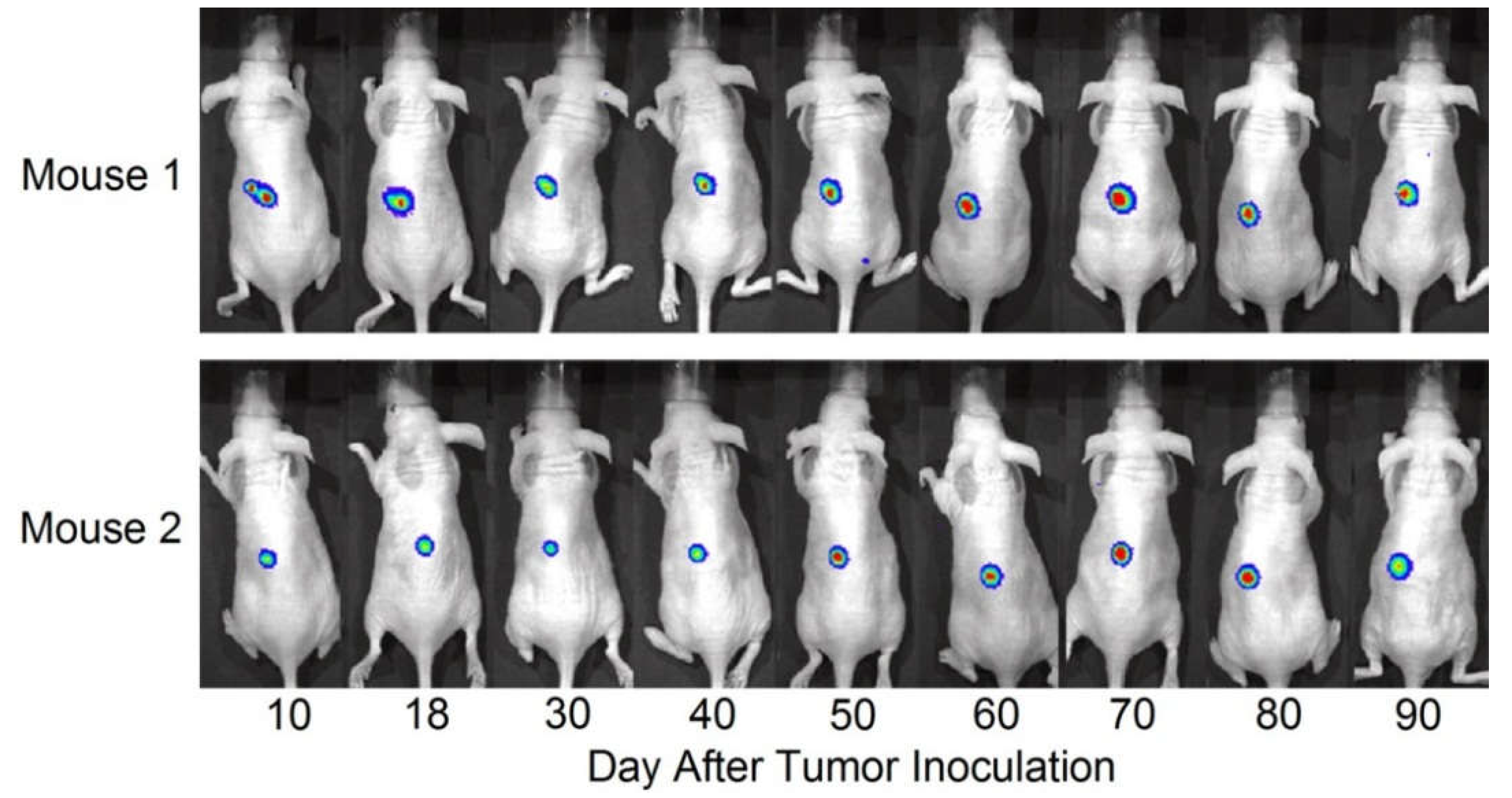
A lentiviral vector (hlac-Luc) was generated to express firefly luciferase under regulation of the human α-lactalbumin promoter (Genecopoeia, Rockville, MD, USA). The human TNBC cell line, HCC1937, was transfected with the hlac-Luc vector, and stable transfectants were established by puromycin resistance. A total of 3–5 × 106 stably transfected HCC1937-hlac-Luc cells were inoculated subcutaneously in the abdominal flanks of 6–8 week old immunodeficient, athymic nude NU/J female mice (Jackson Laboratory, Bar Harbor, ME, USA). Mice were pre-treated every other day for two weeks with intraperitoneal injections of asialo-GM1 antibody (eBioscience, San Diego, CA, USA) to deplete existing NK/NKT cells. Over a 90 day period, inoculated mice were imaged using the Xenogen in vivo imaging system (IVIS; Caliper Life Sciences, Hopkinton, MA, USA) 20 minutes after intraperitoneal injection with D-luciferin (Sigma Aldrich, St. Louis, MO, USA) at 150 mg/kg body weight.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





