
Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review


 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
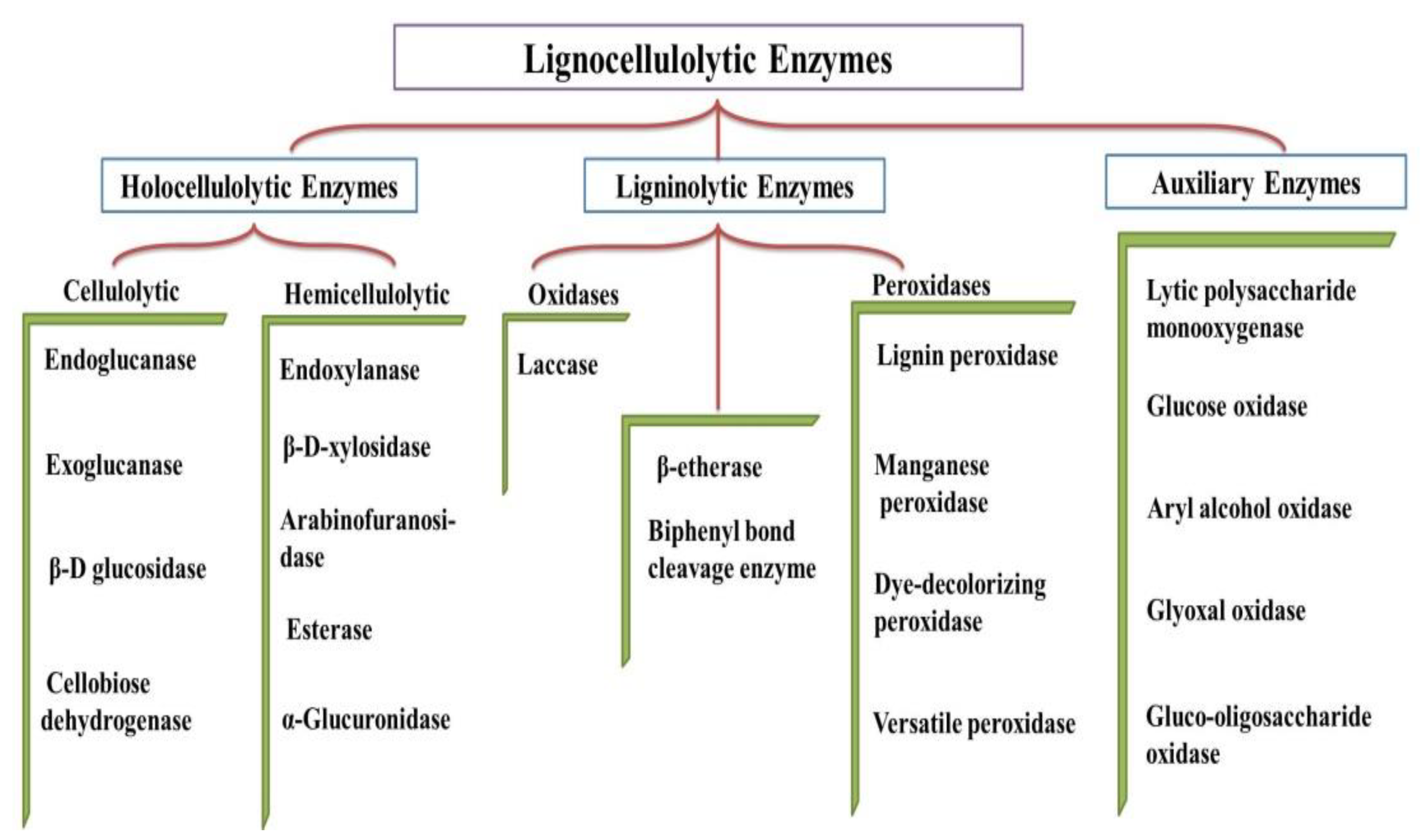
2. Lignocellulolytic Enzymes
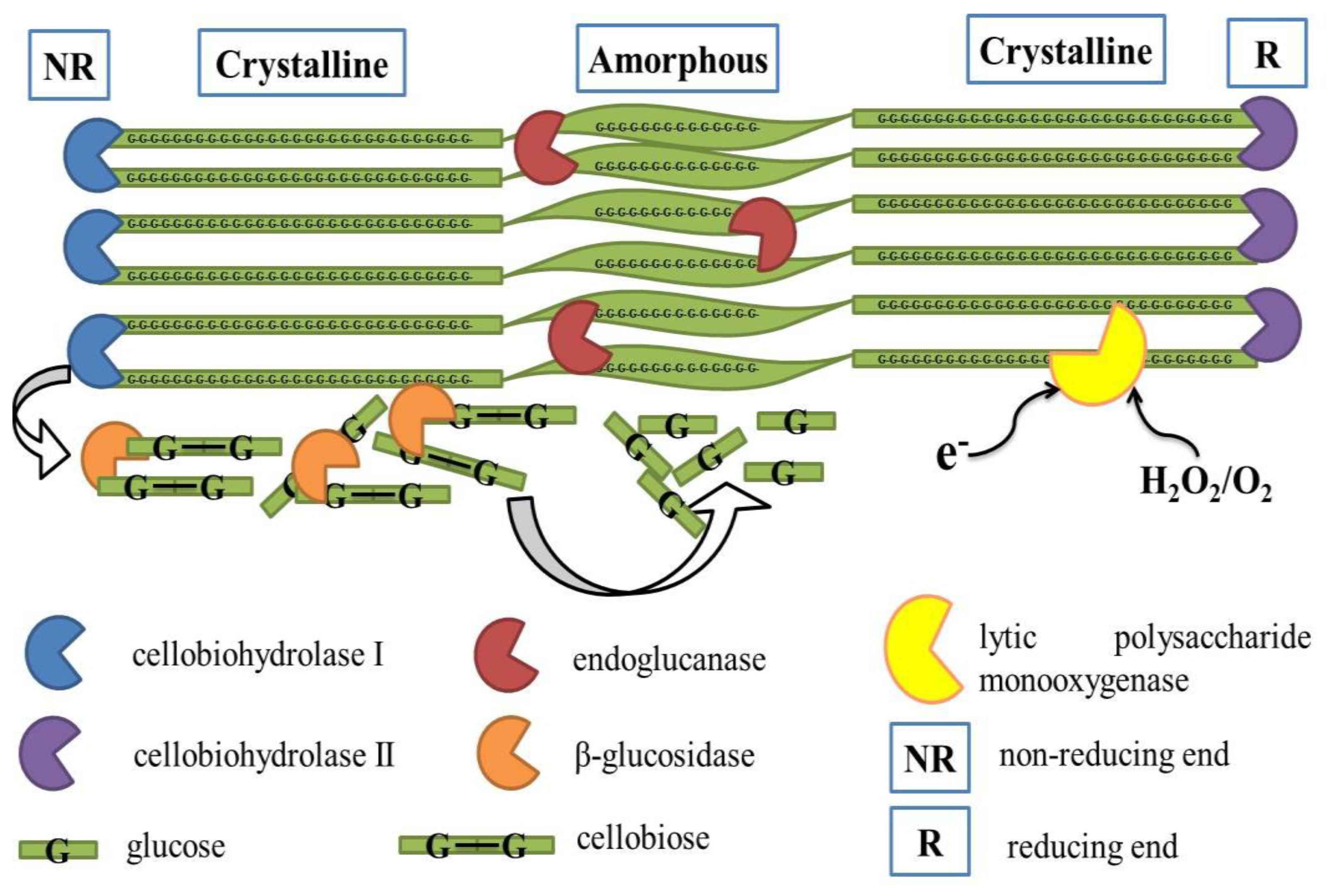
2.1. Cellulase
2.1.1. Endoglucanase
2.1.2. Exoglucanase
2.1.3. β-D Glucosidase
2.1.4. Cellobiose Dehydrogenase
2.1.5. Lytic Polysaccharide Monooxygenase
2.2. Hemicellulases
2.2.1. Endoxylanase
2.2.2. β-D-Xylosidase
2.2.3. Arabinofuranosidase
2.2.4. Esterase
2.2.5. α–Glucuronidase
2.3. Ligninolytic Enzymes
2.3.1. Laccase
2.3.2. Manganese Peroxidase
2.3.3. Lignin Peroxidase
2.3.4. Dye-Decolorizing Peroxidase
2.3.5. β-Etherase
2.3.6. Biphenyl Bond Cleavage Enzyme
3. Microbial Production of Lignocellulolytic Enzymes
3.1. Fungal Lignocellulolytic Enzymes
3.1.1. Soft-Rot Fungi
3.1.2. Brown-Rot Fungi
3.1.3. White-Rot Fungi
3.2. Bacterial Lignocellulolytic Enzymes
3.3. Lignocellulolytic Enzyme Production from Recombinant Microorganisms

{kind=link}
{kind=link}
{kind=link}
| Microorganism | Enzyme | Lignocellulosic Feedstock | Significant Results | Reference |
|---|---|---|---|---|
| Fungi | ||||
| Aspergillus niger ITV02 | Cellulase (β-glucosidase and endoglucanase) | Wheat straw | Glucose yield 24.58 ± 0.08 g/L with a conversion rate of 40.2 ± 0.14%; xylose 8.32 ± 0.02 g/L with a conversion rate of 77.54 ± 0.2% | [118] |
| Penicillium aurantiogriseum | Endoglucanase, cellobiohydrolase and β-glucosidase | Corn stover | Methane yield (281 mLN/g oTS); | [119] |
| Geobacillus sp. | Laccase | Corn stover and bagasse | Corn stover hydrolysis increased by 1.31–2.28 folds (used along with commercial enzymes); bagasse hydrolysis increased by 1.32–2.02 folds (used along with commercial enzymes) | [120] |
| Trametes hirsuta F13 | Laccase and manganese-dependent peroxidase | Beechwood sawdust | 63.58 ± 1.47 mg/mL fermentable sugar from 18-days treated substrate | [121] |
| Aspergillus niger CBS 513.88 | Lytic polysaccharide monooxygenases | Rice straw | 2.31 times more reducing sugar yield together with commercial cellulase | [122] |
| Trichoderma asperellum BPLMBT1 | Laccase | Sweet sorghum stover | Lignin removal of 76.93%; biohydrogen production 402.01 mL | [123] |
| P. citrinum LMI01, Aspergillus sp. LMI03, T. reesei QM9414 | (CMCase), Endoglucanase, β-glucosidase and xylanase | Cellulose pulp and cassava peel | Cellulose pulp-hydrolytic efficiency of 93%; cassava peel-hydrolytic efficiency of 78% | [124] |
| Lentinus squarrosulus MR13, Trichoderma reesei Rut C30 | Laccase and cellulase | Saacharum spontaneum or Kans grass | 81.67% delignification and reducing sugar yield of 500.30 mg/g | [125] |
| Pycnoporus sanguineus MCA 16 | Exoglucanase, endoglucanase, β-glucosidase, xylanase, β-xylosidase, manganese peroxidase laccase and lignin peroxidase | Sugarcane bagasse | Glucose yield-7.32 g/L; total phenolic reduction-82.3% | [126] |
| Marasmiellus palmivorus VE111, Penicillium echinulatum S1M29 | Laccases, peroxidase, cellulase and xylanase | Eucalyptus globulus wood | 31% decrease in the lignin content; 10% increase in the glucose yield; 15% increase in xylose yield | [127] |
| Aspergillus aculeatus PN14 | Cellulase and xylanase | Parthenium hysterophorus | Reducing sugar yield-213.89 mg/g biomass | [26] |
| Aspergillus assiutensis VS34 | Cellulase and xylanase | Sugarcane bagasse | Total reducing sugar yield- 224 mg/g biomass | [30] |
| Bacteria | ||||
| Ruminiclostridium thermocellum M3 | Endoglucanase, exoglucanase, β-glucosidase, and xylanase | Corn straw, corn cobs, rice straw, poplar sawdust | High oligosaccharide yields: corn cobs (77.8 mg/g), corn straw (89.4 mg/g), rice straw (107.8 mg/g), poplar sawdust (52.7 mg/g) | [128] |
| Chromohalobacter salexigens | Laccase | Almond shell | Delignification efficiency strengthened up to 58% | [129] |
| Streptomyces ipomoeae (SilA) | Wheat straw | Decreased phenol content by up to 35% | [130] | |
| Acinetobacter sp. B213, | Manganese peroxidase, lignin peroxidase | Corn straw | 98.51% cellulose retention rate and 12.02%, lignin degradation rate. | [131] |
| Micromonospora sp. G7, Streptomyces sp. H1, Saccharomonospora sp., Mycobacterium sp. | Xylanase, CMCase, lignin peroxidase, manganese peroxidase, laccase | Rice straw, corn straw, wheat straw, soybean straw | Degradation ratio increase: cellulose-46.2%, hemicellulose-22.5%, Lignin-28.9% | [132] |
| Bacillus sp. CX6 | Cellulase and xylanase | Wheat straw | Glucose-6.03 ± 0.12 mg/mL and xylose-6.16 ± 0.07 mg/mL | [133] |
| Cellulomonas sp. CX4 | Cellulase and xylanase | Wheat straw | Glucose-2.36 ± 0.06 mg/mL and xylose-2.43 ± 0.06 mg/mL | [133] |
| Paenibacillus illinoisensis CX11 | Cellulase and xylanase | Wheat straw | Glucose-2.56 ± 0.09 mg/mL and xylose-2.64 ± 0.03 mg/mL [133] | [133] |
| Bacillus cereus CX15 | Cellulase and xylanase | Wheat straw | Glucose-3.39 ± 0.19 mg/mL and xylose-3.61 ± 0.03 mg/mL | [133] |
| Paenibacillus barcinonensis CX17 | Cellulase and xylanase | Wheat straw | Glucose-3.97 ± 0.07 mg/mL and xylose-4.2 ± 0.11 mg/mL | [133] |
| Bacillus ligniniphilus L1 | Laccase | Rice straw | Lignin content decrease-8.93%; phenolic content decrease-44.8% | [134] |
| Thermus sp. 2.9 | Laccase | Eucalyptus | Reducing sugar- 0.96 ± 0.05 mg/mL | [135] |
4. Solid State Fermentation for Enzyme Production
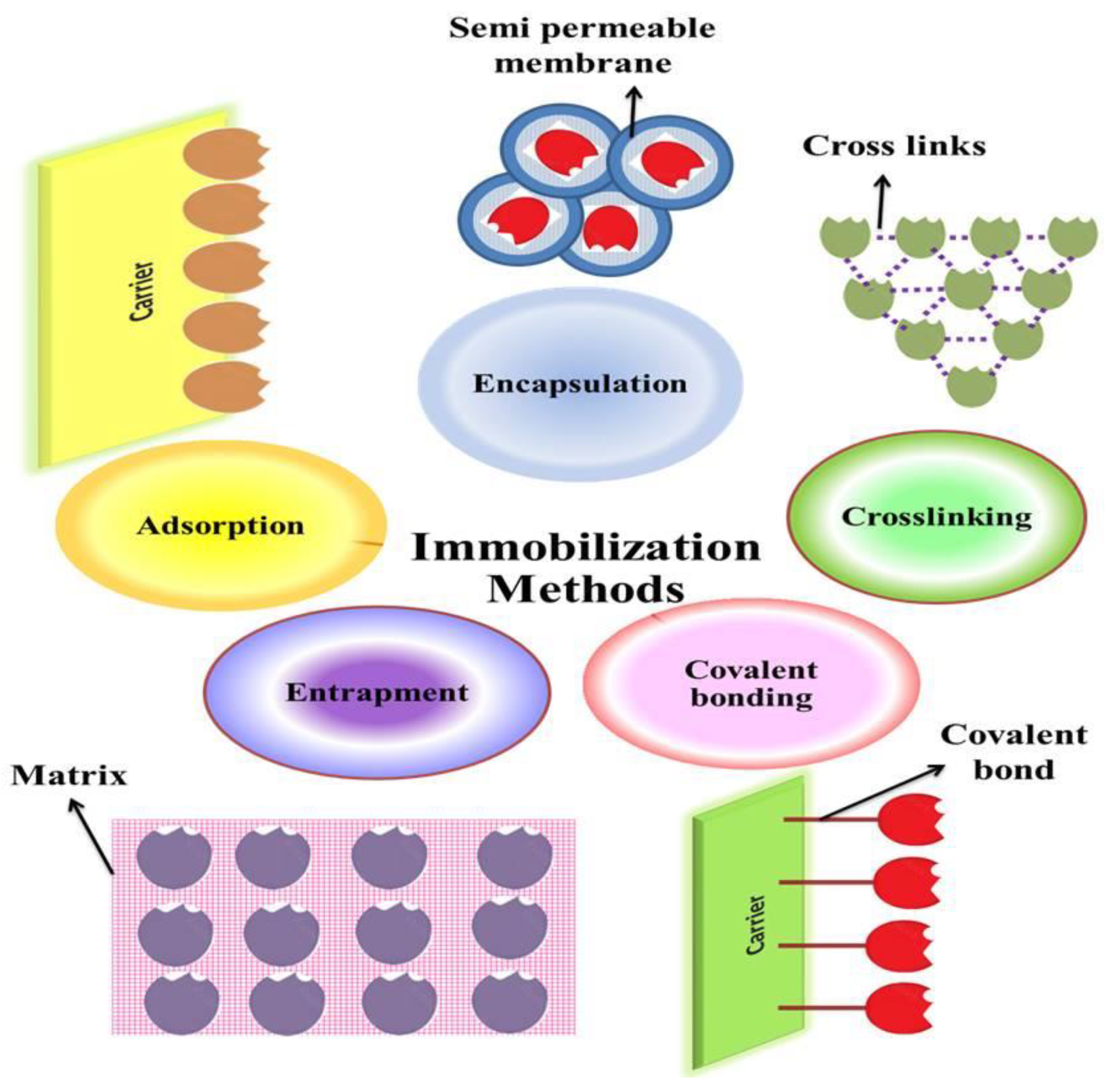
5. Enzyme Immobilization
6. Future Prospects and Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koop, S.H.A.; van Leeuwen, C.J. The Challenges of Water, Waste and Climate Change in Cities. Environ. Dev. Sustain. 2017, 19, 385–418. [Google Scholar] [CrossRef] [Green Version]
- Leong, H.Y.; Chang, C.-K.; Khoo, K.S.; Chew, K.W.; Chia, S.R.; Lim, J.W.; Chang, J.-S.; Show, P.L. Waste Biorefinery towards a Sustainable Circular Bioeconomy: A Solution to Global Issues. Biotechnol. Biofuels 2021, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Le Page, M. Eight Billion Strong. New Sci. 2022, 256, 14–15. [Google Scholar] [CrossRef]
- Taherzadeh, M.J. Bioengineering to Tackle Environmental Challenges, Climate Changes and Resource Recovery. Bioengineered 2019, 10, 698–699. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Tsai, M.-L.; Chen, C.-W.; Sun, P.-P.; Patel, A.K.; Singhania, R.R.; Nargotra, P.; Dong, C.-D. Deep Eutectic Solvents as Promising Pretreatment Agents for Sustainable Lignocellulosic Biorefineries: A Review. Bioresour. Technol. 2022, 360, 127631. [Google Scholar] [CrossRef]
- Sharma, V.; Tsai, M.-L.; Nargotra, P.; Chen, C.-W.; Kuo, C.-H.; Sun, P.-P.; Dong, C.-D. Agro-Industrial Food Waste as a Low-Cost Substrate for Sustainable Production of Industrial Enzymes: A Critical Review. Catalysts 2022, 12, 1373. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Sindhu, R.; Sirohi, R.; Kumar, V.; Ahluwalia, V.; Binod, P.; Juneja, A.; Kumar, D.; Yan, B.; Sarsaiya, S.; et al. Agricultural Waste Biorefinery Development towards Circular Bioeconomy. Renew. Sustain. Energy Rev. 2022, 158, 112122. [Google Scholar] [CrossRef]
- Rojas, L.F.; Zapata, P.; Ruiz-Tirado, L. Agro-Industrial Waste Enzymes: Perspectives in Circular Economy. Curr. Opin. Green Sustain. Chem. 2022, 34, 100585. [Google Scholar] [CrossRef]
- Devi, A.; Bajar, S.; Kour, H.; Kothari, R.; Pant, D.; Singh, A. Lignocellulosic Biomass Valorization for Bioethanol Production: A Circular Bioeconomy Approach. Bioenergy Res. 2022. [Google Scholar] [CrossRef]
- Devi, M.M.; Aggarwal, N.; Saravanamurugan, S. Rice Straw: A Major Renewable Lignocellulosic Biomass for Value-Added Carbonaceous Materials. Curr. Green Chem. 2020, 7, 290–303. [Google Scholar] [CrossRef]
- Kaur, N.; Singh, G.; Khatri, M.; Arya, S.K. Review on Neoteric Biorefinery Systems from Detritus Lignocellulosic Biomass: A Profitable Approach. J. Clean. Prod. 2020, 256, 120607. [Google Scholar] [CrossRef]
- Sharma, V.; Bhat, B.; Gupta, M.; Vaid, S.; Sharma, S.; Nargotra, P.; Singh, S.; Bajaj, B.K. Role of Systematic Biology in Biorefining of Lignocellulosic Residues for Biofuels and Chemicals Production. In Sustainable Biotechnology- Enzymatic Resources of Renewable Energy; Singh, O.V., Chandel, A.K., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 5–55. ISBN 978-3-319-95480-6. [Google Scholar]
- Vaid, S.; Bhat, N.; Nargotra, P.; Bajaj, B.K. Combinatorial Application of Ammonium Carbonate and Sulphuric Acid Pretreatment to Achieve Enhanced Sugar Yield from Pine Needle Biomass for Potential Biofuel–Ethanol Production. Energy Ecol. Environ. 2018, 3, 126–135. [Google Scholar] [CrossRef]
- Sharma, V.; Nargotra, P.; Sharma, S.; Bajaj, B.K. Efficacy and Functional Mechanisms of a Novel Combinatorial Pretreatment Approach Based on Deep Eutectic Solvent and Ultrasonic Waves for Bioconversion of Sugarcane Bagasse. Renew. Energy 2021, 163, 1910–1922. [Google Scholar] [CrossRef]
- Nargotra, P.; Sharma, V.; Sharma, S.; Bangotra, R.; Bajaj, B.K. Purification of an Ionic Liquid Stable Cellulase from Aspergillus aculeatus PN14 with Potential for Biomass Refining. Environ. Sustain. 2022, 5, 313–323. [Google Scholar] [CrossRef]
- Nargotra, P.; Sharma, V.; Sharma, S.; Kapoor, N.; Bajaj, B.K. Development of Consolidated Bioprocess for Biofuel-Ethanol Production from Ultrasound-Assisted Deep Eutectic Solvent Pretreated Parthenium hysterophorus Biomass. Biomass Conv. Bioref. 2020. [Google Scholar] [CrossRef]
- Saini, S.; Sharma, K.K. Fungal Lignocellulolytic Enzymes and Lignocellulose: A Critical Review on Their Contribution to Multiproduct Biorefinery and Global Biofuel Research. Int. J. Biol. Macromol. 2021, 193, 2304–2319. [Google Scholar] [CrossRef] [PubMed]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Ideal Feedstock and Fermentation Process Improvements for the Production of Lignocellulolytic Enzymes. Processes 2021, 9, 38. [Google Scholar] [CrossRef]
- Ferreira-Leitão, V.S.; Cammarota, M.C.; Gonçalves Aguieiras, E.C.; Vasconcelos de Sá, L.R.; Fernandez-Lafuente, R.; Freire, D.M.G. The Protagonism of Biocatalysis in Green Chemistry and Its Environmental Benefits. Catalysts 2017, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Saldarriaga-Hernández, S.; Velasco-Ayala, C.; Leal-Isla Flores, P.; de Jesús Rostro-Alanis, M.; Parra-Saldivar, R.; Iqbal, H.M.N.; Carrillo-Nieves, D. Biotransformation of Lignocellulosic Biomass into Industrially Relevant Products with the Aid of Fungi-Derived Lignocellulolytic Enzymes. Int. J. Biol. Macromol. 2020, 161, 1099–1116. [Google Scholar] [CrossRef]
- Cheng, P.-Y.; Liao, H.-Y.; Kuo, C.-H.; Liu, Y.-C. Enhanced Erinacine A Production by Hericium erinaceus Using Solid-State Cultivation. Fermentation 2021, 7, 182. [Google Scholar] [CrossRef]
- Sharma, S.; Nargotra, P.; Sharma, V.; Bangotra, R.; Kaur, M.; Kapoor, N.; Paul, S.; Bajaj, B.K. Nanobiocatalysts for Efficacious Bioconversion of Ionic Liquid Pretreated Sugarcane Tops Biomass to Biofuel. Bioresour. Technol. 2021, 333, 125191. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.-H.; Nargotra, P.; Kuo, C.-H.; Liu, Y.-C. Immobilization of Recombinant Endoglucanase (CelA) from Clostridium thermocellum on Modified Regenerated Cellulose Membrane. Catalysts 2022, 12, 1356. [Google Scholar] [CrossRef]
- Nargotra, P.; Vaid, S.; Bajaj, B.K. Cellulase Production from Bacillus subtilis SV1 and Its Application Potential for Saccharification of Ionic Liquid Pretreated Pine Needle Biomass under One Pot Consolidated Bioprocess. Fermentation 2016, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Nargotra, P.; Sharma, V.; Bajaj, B.K. Consolidated Bioprocessing of Surfactant-Assisted Ionic Liquid-Pretreated Parthenium hysterophorus L. Biomass for Bioethanol Production. Bioresour. Technol. 2019, 289, 121611. [Google Scholar] [CrossRef]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from Enzymatic Degradation of Cellulose and Hemicellulose to Fermentable Sugars—A Review. Biomass Bioenergy 2020, 134, 105481. [Google Scholar] [CrossRef]
- Huang, C.; Li, R.; Tang, W.; Zheng, Y.; Meng, X. Improve Enzymatic Hydrolysis of Lignocellulosic Biomass by Modifying Lignin Structure via Sulfite Pretreatment and Using Lignin Blockers. Fermentation 2022, 8, 558. [Google Scholar] [CrossRef]
- Vaid, S.; Nargotra, P.; Bajaj, B.K. Consolidated Bioprocessing for Biofuel-Ethanol Production from Pine Needle Biomass. Environ. Prog. Sustain. Energy 2018, 37, 546–552. [Google Scholar] [CrossRef]
- Sharma, V.; Nargotra, P.; Bajaj, B.K. Ultrasound and Surfactant Assisted Ionic Liquid Pretreatment of Sugarcane Bagasse for Enhancing Saccharification Using Enzymes from an Ionic Liquid Tolerant Aspergillus assiutensis VS34. Bioresour. Technol. 2019, 285, 121319. [Google Scholar] [CrossRef]
- Ashokkumar, V.; Venkatkarthick, R.; Jayashree, S.; Chuetor, S.; Dharmaraj, S.; Kumar, G.; Chen, W.-H.; Ngamcharussrivichai, C. Recent Advances in Lignocellulosic Biomass for Biofuels and Value-Added Bioproducts—A Critical Review. Bioresour. Technol. 2022, 344, 126195. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Raj, T.; Chen, C.-W.; Ponnusamy, V.K.; Tahir, N.; Kim, S.-H.; Dong, C.-D. Lignin Valorisation via Enzymes: A Sustainable Approach. Fuel 2022, 311, 122608. [Google Scholar] [CrossRef]
- Zerva, A.; Pentari, C.; Ferousi, C.; Nikolaivits, E.; Karnaouri, A.; Topakas, E. Recent Advances on Key Enzymatic Activities for the Utilisation of Lignocellulosic Biomass. Bioresour. Technol. 2021, 342, 126058. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The Carbohydrate-Active Enzyme Database: Functions and Literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-Active Enzymes Database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomazetto, G.; Pimentel, A.C.; Wibberg, D.; Dixon, N.; Squina, F.M. Multi-omic directed discovery of cellulosomes, polysaccharide utilization loci, and lignocellulases from an enriched rumen anaerobic consortium. Appl. Environ. Microbiol. 2020, 86, e00199-20. [Google Scholar] [CrossRef]
- Oates, N.C.; Abood, A.; Schirmacher, A.M.; Alessi, A.M.; Bird, S.M.; Bennett, J.P.; Leadbeater, D.R.; Li, Y.; Dowle, A.A.; Liu, S.; et al. A multi-omics approach to lignocellulolytic enzyme discovery reveals a new ligninase activity from Parascedosporium putredinis NO1. Proc. Natl. Acad. Sci. USA 2021, 118, e2008888118. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Kumar, G.; Vickram, A.S.; Mohan, M.; Singhania, R.R.; Patel, A.K.; Dong, C.-D.; Anbarasu, K.; Thanigaivel, S.; Ponnusamy, V.K. Nanotechnology-Assisted Production of Value-Added Biopotent Energy-Yielding Products from Lignocellulosic Biomass Refinery—A Review. Bioresour. Technol. 2022, 344, 126171. [Google Scholar] [CrossRef]
- Arpit Singh, T.; Sharma, M.; Sharma, M.; Dutt Sharma, G.; Kumar Passari, A.; Bhasin, S. Valorization of Agro-Industrial Residues for Production of Commercial Biorefinery Products. Fuel 2022, 322, 124284. [Google Scholar] [CrossRef]
- Siqueira, J.G.W.; Rodrigues, C.; Vandenberghe, L.P.d.S.; Woiciechowski, A.L.; Soccol, C.R. Current Advances in On-Site Cellulase Production and Application on Lignocellulosic Biomass Conversion to Biofuels: A Review. Biomass Bioenergy 2020, 132, 105419. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.B.; Binod, P.; Sirohi, R.; Tarafdar, A.; Reshmy, R.; Kumar Awasthi, M.; Sindhu, R. Design of Novel Enzyme Biocatalysts for Industrial Bioprocess: Harnessing the Power of Protein Engineering, High Throughput Screening and Synthetic Biology. Bioresour. Technol. 2021, 325, 124617. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, K.-D.; Zhang, P.-Y.; Zhou, X.; Ma, X.-Q.; Li, F.-L. Synergistic Cellulose Hydrolysis Dominated by a Multi-Modular Processive Endoglucanase from Clostridium cellulosi. Front. Microbiol. 2016, 7, 932. [Google Scholar] [CrossRef] [Green Version]
- Jayasekara, S.; Ratnayake, R. Microbial Cellulases: An Overview and Applications. In Cellulose; Rodríguez Pascual, A., Eugenio Martín, M.E., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-83968-056-4. [Google Scholar]
- Warner, C.D.; Camci-Unal, G.; Pohl, N.L.B.; Ford, C.; Reilly, P.J. Substrate Binding by the Catalytic Domain and Carbohydrate Binding Module of Ruminococcus flavefaciens FD-1 Xyloglucanase/Endoglucanase. Ind. Eng. Chem. Res. 2013, 52, 30–36. [Google Scholar] [CrossRef]
- Akram, F.; Haq, I.u.; Imran, W.; Mukhtar, H. Insight Perspectives of Thermostable Endoglucanases for Bioethanol Production: A Review. Renew. Energy 2018, 122, 225–238. [Google Scholar] [CrossRef]
- Raghuwanshi, S.; Deswal, D.; Karp, M.; Kuhad, R.C. Bioprocessing of Enhanced Cellulase Production from a Mutant of Trichoderma asperellum RCK2011 and Its Application in Hydrolysis of Cellulose. Fuel 2014, 124, 183–189. [Google Scholar] [CrossRef]
- Hiras, J.; Wu, Y.-W.; Deng, K.; Nicora, C.D.; Aldrich, J.T.; Frey, D.; Kolinko, S.; Robinson, E.W.; Jacobs, J.M.; Adams, P.D.; et al. Comparative Community Proteomics Demonstrates the Unexpected Importance of Actinobacterial Glycoside Hydrolase Family 12 Protein for Crystalline Cellulose Hydrolysis. mBio 2016, 7, e01106-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annamalai, N.; Rajeswari, M.V.; Sivakumar, N. Cellobiohydrolases: Role, Mechanism, and Recent Developments. In Microbial Enzymes in Bioconversions of Biomass; Gupta, V.K., Ed.; Biofuel and Biorefinery Technologies; Springer International Publishing: Cham, Switzerland, 2016; pp. 29–35. ISBN 978-3-319-43679-1. [Google Scholar]
- Yao, G.; Wu, R.; Kan, Q.; Gao, L.; Liu, M.; Yang, P.; Du, J.; Li, Z.; Qu, Y. Production of a High-Efficiency Cellulase Complex via β-Glucosidase Engineering in Penicillium oxalicum. Biotechnol. Biofuels 2016, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naraian, R.; Gautam, R.L. Penicillium Enzymes for the Saccharification of Lignocellulosic Feedstocks. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 121–136. ISBN 978-0-444-63501-3. [Google Scholar]
- Singhania, R.R.; Patel, A.K.; Pandey, A.; Ganansounou, E. Genetic Modification: A Tool for Enhancing Beta-Glucosidase Production for Biofuel Application. Bioresour. Technol. 2017, 245, 1352–1361. [Google Scholar] [CrossRef]
- Srivastava, N.; Rathour, R.; Jha, S.; Pandey, K.; Srivastava, M.; Thakur, V.K.; Sengar, R.S.; Gupta, V.K.; Mazumder, P.B.; Khan, A.F.; et al. Microbial Beta Glucosidase Enzymes: Recent Advances in Biomass Conversation for Biofuels Application. Biomolecules 2019, 9, 220. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.T.; Ruiz-Dueñas, F.J.; Camarero, S.; Serrano, A.; Linde, D.; Lund, H.; Vind, J.; Tovborg, M.; Herold-Majumdar, O.M.; Hofrichter, M.; et al. Oxidoreductases on Their Way to Industrial Biotransformations. Biotechnol. Adv. 2017, 35, 815–831. [Google Scholar] [CrossRef] [Green Version]
- Sulej, J.; Osińska-Jaroszuk, M.; Jaszek, M.; Grąz, M.; Kutkowska, J.; Pawlik, A.; Chudzik, A.; Bancerz, R. Antimicrobial and Antioxidative Potential of Free and Immobilised Cellobiose Dehydrogenase Isolated from Wood Degrading Fungi. Fungal Biol. 2019, 123, 875–886. [Google Scholar] [CrossRef]
- Arfi, Y.; Shamshoum, M.; Rogachev, I.; Peleg, Y.; Bayer, E.A. Integration of Bacterial Lytic Polysaccharide Monooxygenases into Designer Cellulosomes Promotes Enhanced Cellulose Degradation. Proc. Natl. Acad. Sci. USA 2014, 111, 9109–9114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bissaro, B.; Røhr, Å.K.; Müller, G.; Chylenski, P.; Skaugen, M.; Forsberg, Z.; Horn, S.J.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Oxidative Cleavage of Polysaccharides by Monocopper Enzymes Depends on H2O2. Nat. Chem. Biol. 2017, 13, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Kont, R.; Bissaro, B.; Eijsink, V.G.H.; Väljamäe, P. Kinetic Insights into the Peroxygenase Activity of Cellulose-Active Lytic Polysaccharide Monooxygenases (LPMOs). Nat. Commun. 2020, 11, 5786. [Google Scholar] [CrossRef]
- Vu, V.V.; Beeson, W.T.; Span, E.A.; Farquhar, E.R.; Marletta, M.A. A family of starch-active polysaccharide monooxygenases. Proc. Natl. Acad. Sci. USA 2014, 111, 13822–13827. [Google Scholar] [CrossRef] [Green Version]
- Sabbadin, F.; Hemsworth, G.R.; Ciano, L.; Henrissat, B.; Dupree, P.; Tryfona, T.; Marques, R.D.S.; Sweeney, S.T.; Besser, K.; Elias, L.; et al. An Ancient Family of Lytic Polysaccharide Monooxygenases with Roles in Arthropod Development and Biomass Digestion. Nat. Commun. 2018, 9, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco Cairo, J.P.L.; Almeida, D.V.; Damasio, A.; Garcia, W.; Squina, F.M. The Periplasmic Expression and Purification of AA15 Lytic Polysaccharide Monooxygenases from Insect Species in Escherichia coli. Protein Expr. Purif. 2022, 190, 105994. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C. Solid State Fermentation for Production of Microbial Cellulases: Recent Advances and Improvement Strategies. Int. J. Biol. Macromol. 2016, 86, 656–669. [Google Scholar] [CrossRef]
- Mohanram, S.; Amat, D.; Choudhary, J.; Arora, A.; Nain, L. Novel Perspectives for Evolving Enzyme Cocktails for Lignocellulose Hydrolysis in Biorefineries. Sustain. Chem. Process. 2013, 1, 15. [Google Scholar] [CrossRef] [Green Version]
- Adsul, M.; Sandhu, S.K.; Singhania, R.R.; Gupta, R.; Puri, S.K.; Mathur, A. Designing a Cellulolytic Enzyme Cocktail for the Efficient and Economical Conversion of Lignocellulosic Biomass to Biofuels. Enzym. Microb. Technol. 2020, 133, 109442. [Google Scholar] [CrossRef]
- Sethi, A.; Scharf, M.E. Biofuels: Fungal, Bacterial and Insect Degraders of Lignocellulose. In eLS; John Wiley & Sons, Ltd.; Wiley: Hoboken, NJ, USA, 2013; ISBN 978-0-470-01617-6. [Google Scholar]
- Albornoz-Palma, G.; Ching, D.; Valerio, O.; Mendonça, R.T.; Pereira, M. Effect of Lignin and Hemicellulose on the Properties of Lignocellulose Nanofibril Suspensions. Cellulose 2020, 27, 10631–10647. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, V.; Nargotra, P.; Bajaj, B.K. Process Desired Functional Attributes of an Endoxylanase of GH10 Family from a New Strain of Aspergillus terreus S9. Int. J. Biol. Macromol. 2018, 115, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sharma, V.; Nargotra, P.; Bajaj, B.K. Bioprocess Development for Production of a Process-Apt Xylanase with Multifaceted Application Potential for a Range of Industrial Processes. SN Appl. Sci. 2020, 2, 739. [Google Scholar] [CrossRef] [Green Version]
- Kubicek, C.P. The tools—Part 2: Enzymology of hemicellulose degradation. In Fungi and Lignocellulosic Biomass, 1st ed.; Wiley: Hoboken, NJ, USA, 2012; ISBN 978-0-470-96009-7. [Google Scholar]
- Najjarzadeh, N.; Matsakas, L.; Rova, U.; Christakopoulos, P. Effect of Oligosaccharide Degree of Polymerization on the Induction of Xylan-Degrading Enzymes by Fusarium oxysporum f. sp. Lycopersici. Molecules 2020, 25, 5849. [Google Scholar] [CrossRef] [PubMed]
- Østby, H.; Hansen, L.D.; Horn, S.J.; Eijsink, V.G.H.; Várnai, A. Enzymatic Processing of Lignocellulosic Biomass: Principles, Recent Advances and Perspectives. J. Ind. Microbiol. Biotechnol. 2020, 47, 623–657. [Google Scholar] [CrossRef] [PubMed]
- Carli, S.; Meleiro, L.P.; Rosa, J.C.; Moraes, L.A.B.; Jorge, J.A.; Masui, D.C.; Furriel, R.P.M. A Novel Thermostable and Halotolerant Xylanase from Colletotrichum graminicola. J. Mol. Catal. B Enzym. 2016, 133, S508–S517. [Google Scholar] [CrossRef]
- Luo, L.; Cai, J.; Wang, C.; Lin, J.; Du, X.; Zhou, A.; Xiang, M. Purification and Characterization of an Alkaliphilic Endo-Xylanase from Streptomyces althioticus LMZM and Utilization in the Pulp Paper Industry. J. Chem. Technol. Biotechnol. 2016, 91, 1093–1098. [Google Scholar] [CrossRef]
- Alokika; Singh, B. Production, Characteristics, and Biotechnological Applications of Microbial Xylanases. Appl. Microbiol. Biotechnol. 2019, 103, 8763–8784. [Google Scholar] [CrossRef]
- Santibáñez, L.; Henríquez, C.; Corro-Tejeda, R.; Bernal, S.; Armijo, B.; Salazar, O. Xylooligosaccharides from Lignocellulosic Biomass: A Comprehensive Review. Carbohydr. Polym. 2021, 251, 117118. [Google Scholar] [CrossRef]
- Binod, P.; Gnansounou, E.; Sindhu, R.; Pandey, A. Enzymes for Second Generation Biofuels: Recent Developments and Future Perspectives. Bioresour. Technol. Rep. 2019, 5, 317–325. [Google Scholar] [CrossRef]
- Rohman, A.; Dijkstra, B.W.; Puspaningsih, N.N.T. β-Xylosidases: Structural Diversity, Catalytic Mechanism, and Inhibition by Monosaccharides. Int. J. Mol. Sci. 2019, 20, 5524. [Google Scholar] [CrossRef]
- Contesini, F.J.; Liberato, M.V.; Rubio, M.V.; Calzado, F.; Zubieta, M.P.; Riaño-Pachón, D.M.; Squina, F.M.; Bracht, F.; Skaf, M.S.; Damasio, A.R. Structural and Functional Characterization of a Highly Secreted α-l-Arabinofuranosidase (GH62) from Aspergillus nidulans Grown on Sugarcane Bagasse. Biochim. Biophys. Acta. Proteins Proteom. BBA-Proteins Proteom. 2017, 1865, 1758–1769. [Google Scholar] [CrossRef] [PubMed]
- Karnaouri, A.; Antonopoulou, I.; Zerva, A.; Dimarogona, M.; Topakas, E.; Rova, U.; Christakopoulos, P. Thermophilic Enzyme Systems for Efficient Conversion of Lignocellulose to Valuable Products: Structural Insights and Future Perspectives for Esterases and Oxidative Catalysts. Bioresour. Technol. 2019, 279, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Sista Kameshwar, A.K.; Qin, W. Understanding the Structural and Functional Properties of Carbohydrate Esterases with a Special Focus on Hemicellulose Deacetylating Acetyl Xylan Esterases. Mycology 2018, 9, 273–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malgas, S.; Mafa, M.S.; Mkabayi, L.; Pletschke, B.I. A Mini Review of Xylanolytic Enzymes with Regards to Their Synergistic Interactions during Hetero-Xylan Degradation. World J. Microbiol. Biotechnol. 2019, 35, 187. [Google Scholar] [CrossRef]
- Irdawati, I.; Syamsuardi, S.; Agustien, A.; Rilda, Y. Screening of Thermophilic Bacteria Produce Xylanase from Sapan Sungai Aro Hot Spring South Solok. IOP Conf. Ser. Mater. Sci. Eng. 2018, 335, 012021. [Google Scholar] [CrossRef]
- Chadha, B.S.; Kaur, B.; Basotra, N.; Tsang, A.; Pandey, A. Thermostable Xylanases from Thermophilic Fungi and Bacteria: Current Perspective. Bioresour. Technol. 2019, 277, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Chio, C.; Sain, M.; Qin, W. Lignin Utilization: A Review of Lignin Depolymerization from Various Aspects. Renew. Sustain. Energy Rev. 2019, 107, 232–249. [Google Scholar] [CrossRef]
- Suseela, V. Potential Roles of Plant Biochemistry in Mediating Ecosystem Responses to Warming and Drought. In Ecosystem Consequences of Soil Warming; Elsevier: Amsterdam, The Netherlands, 2019; pp. 103–124. ISBN 978-0-12-813493-1. [Google Scholar]
- Weng, C.; Peng, X.; Han, Y. Depolymerization and Conversion of Lignin to Value-Added Bioproducts by Microbial and Enzymatic Catalysis. Biotechnol. Biofuels 2021, 14, 84. [Google Scholar] [CrossRef]
- Mayr, S.A.; Subagia, R.; Weiss, R.; Schwaiger, N.; Weber, H.K.; Leitner, J.; Ribitsch, D.; Nyanhongo, G.S.; Guebitz, G.M. Oxidation of Various Kraft Lignins with a Bacterial Laccase Enzyme. Int. J. Mol. Sci. 2021, 22, 13161. [Google Scholar] [CrossRef]
- Datta, R.; Kelkar, A.; Baraniya, D.; Molaei, A.; Moulick, A.; Meena, R.S.; Formanek, P. Enzymatic Degradation of Lignin in Soil: A Review. Sustainability 2017, 9, 1163. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Rea, D.; Jamshidi, S.; Fülöp, V.; Bugg, T.D.H. Structure of Thermobifida fusca DyP-Type Peroxidase and Activity towards Kraft Lignin and Lignin Model Compounds. Arch. Biochem. Biophys. 2016, 594, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Gonzalo, G.; Colpa, D.I.; Habib, M.H.M.; Fraaije, M.W. Bacterial Enzymes Involved in Lignin Degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Bugg, T.D.H.; Williamson, J.J.; Rashid, G.M.M. Bacterial Enzymes for Lignin Depolymerisation: New Biocatalysts for Generation of Renewable Chemicals from Biomass. Curr. Opin. Chem. Biol. 2020, 55, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Picart, P.; Domínguez de María, P.; Schallmey, A. From Gene to Biorefinery: Microbial β-Etherases as Promising Biocatalysts for Lignin Valorization. Front. Microbiol. 2015, 6, 916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-Degrading Enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef] [PubMed]
- Kracher, D.; Ludwig, R. Cellobiose Dehydrogenase: An Essential Enzyme for Lignocellulose Degradation in Nature—A Review/Cellobiosedehydrogenase: Ein Essentielles Enzym Für Den Lignozelluloseabbau in Der Natur—Eine Übersicht. Die Bodenkultur J. Land Manag. Food Environ. 2016, 67, 145–163. [Google Scholar] [CrossRef] [Green Version]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose Degradation: An Overview of Fungi and Fungal Enzymes Involved in Lignocellulose Degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Madadi, M.; Abbas, A. Lignin Degradation by Fungal Pretreatment: A Review. J. Plant Pathol. Microbiol. 2017, 8, 398–404. [Google Scholar] [CrossRef]
- Altgen, M.; Kyyrö, S.; Paajanen, O.; Rautkari, L. Resistance of Thermally Modified and Pressurized Hot Water Extracted Scots Pine Sapwood against Decay by the Brown-Rot Fungus Rhodonia placenta. Eur. J. Wood Prod. 2020, 78, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Nurika, I.; Suhartini, S.; Barker, G.C. Biotransformation of Tropical Lignocellulosic Feedstock Using the Brown Rot Fungus Serpula lacrymans. Waste Biomass Valor. 2020, 11, 2689–2700. [Google Scholar] [CrossRef]
- Monrroy, M.; Ortega, I.; Ramírez, M.; Baeza, J.; Freer, J. Structural Change in Wood by Brown Rot Fungi and Effect on Enzymatic Hydrolysis. Enzym. Microb. Technol. 2011, 49, 472–477. [Google Scholar] [CrossRef]
- Hermosilla, E.; Rubilar, O.; Schalchli, H.; da Silva, A.S.; Ferreira-Leitao, V.; Diez, M.C. Sequential White-Rot and Brown-Rot Fungal Pretreatment of Wheat Straw as a Promising Alternative for Complementary Mild Treatments. Waste Manag. 2018, 79, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Marinović, M.; Aguilar-Pontes, M.V.; Zhou, M.; Miettinen, O.; de Vries, R.P.; Mäkelä, M.R.; Hildén, K. Temporal Transcriptome Analysis of the White-Rot Fungus Obba rivulosa Shows Expression of a Constitutive Set of Plant Cell Wall Degradation Targeted Genes during Growth on Solid Spruce Wood. Fungal Genet. Biol. 2018, 112, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zabed, H.M.; Akter, S.; Yun, J.; Zhang, G.; Awad, F.N.; Qi, X.; Sahu, J.N. Recent Advances in Biological Pretreatment of Microalgae and Lignocellulosic Biomass for Biofuel Production. Renew. Sustain. Energy Rev. 2019, 105, 105–128. [Google Scholar] [CrossRef]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Pretreatment of Radiata Pine Using Two White Rot Fungal Strains Stereum hirsutum and Trametes versicolor. Energy Convers. Manag. 2017, 142, 13–19. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, J.; Gu, X.; Yu, H.; Chen, S. A Comprehensive Study of the Promoting Effect of Manganese on White Rot Fungal Treatment for Enzymatic Hydrolysis of Woody and Grass Lignocellulose. Biotechnol. Biofuels 2021, 14, 176. [Google Scholar] [CrossRef]
- Ding, C.; Wang, X.; Li, M. Evaluation of Six White-Rot Fungal Pretreatments on Corn Stover for the Production of Cellulolytic and Ligninolytic Enzymes, Reducing Sugars, and Ethanol. Appl. Microbiol. Biotechnol. 2019, 103, 5641–5652. [Google Scholar] [CrossRef]
- Chauhan, P.S. Role of Various Bacterial Enzymes in Complete Depolymerization of Lignin: A Review. Biocatal. Agric. Biotechnol. 2020, 23, 101498. [Google Scholar] [CrossRef]
- Limaye, L.; Patil, R.; Ranadive, P.; Kamath, G. Application of Potent Actinomycete Strains for Bio-Degradation of Domestic Agro-Waste by Composting and Treatment of Pulp-Paper Mill Effluent. Adv. Appl. Microbiol. 2017, 7, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Artzi, L.; Bayer, E.A.; Moraïs, S. Cellulosomes: Bacterial Nanomachines for Dismantling Plant Polysaccharides. Nat. Rev. Microbiol. 2017, 15, 83–95. [Google Scholar] [CrossRef]
- Artzi, L.; Dadosh, T.; Milrot, E.; Moraïs, S.; Levin-Zaidman, S.; Morag, E.; Bayer, E.A. Colocalization and Disposition of Cellulosomes in Clostridium clariflavum as Revealed by Correlative Superresolution Imaging. mBio 2018, 9, e00012-18. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, S.; Kurosaki, M.; Kokuzawa, T.; Hirano, K.; Takano, H.; Ueda, K.; Haruki, M.; Hirano, N. Comparative Biochemical Analysis of Cellulosomes Isolated from Clostridium clariflavum DSM 19732 and Clostridium thermocellum ATCC 27405 Grown on Plant Biomass. Appl. Biochem. Biotechnol. 2019, 187, 994–1010. [Google Scholar] [CrossRef]
- Plácido, J.; Capareda, S. Ligninolytic Enzymes: A Biotechnological Alternative for Bioethanol Production. Bioresour. Bioprocess. 2015, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Granja-Travez, R.S.; Persinoti, G.F.; Squina, F.M.; Bugg, T.D.H. Functional Genomic Analysis of Bacterial Lignin Degraders: Diversity in Mechanisms of Lignin Oxidation and Metabolism. Appl. Microbiol. Biotechnol. 2020, 104, 3305–3320. [Google Scholar] [CrossRef]
- Mei, J.; Shen, X.; Gang, L.; Xu, H.; Wu, F.; Sheng, L. A Novel Lignin Degradation Bacteria-Bacillus amyloliquefaciens SL-7 Used to Degrade Straw Lignin Efficiently. Bioresour. Technol. 2020, 310, 123445. [Google Scholar] [CrossRef]
- Xu, W.; Fu, S.; Yang, Z.; Lu, J.; Guo, R. Improved Methane Production from Corn Straw by Microaerobic Pretreatment with a Pure Bacteria System. Bioresour. Technol. 2018, 259, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Rashid, G.M.M.; Durán-Peña, M.J.; Rahmanpour, R.; Sapsford, D.; Bugg, T.D.H. Delignification and Enhanced Gas Release from Soil Containing Lignocellulose by Treatment with Bacterial Lignin Degraders. J. Appl. Microbiol. 2017, 123, 159–171. [Google Scholar] [CrossRef]
- Davison, S.A.; den Haan, R.; van Zyl, W.H. Exploiting Strain Diversity and Rational Engineering Strategies to Enhance Recombinant Cellulase Secretion by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2020, 104, 5163–5184. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Du, J.-L.; Zhan, Y.-J.; Li, J.-A.; Zuo, R.-R.; Tian, S. Consolidated Bioprocessing for Cellulosic Ethanol Conversion by Cellulase–Xylanase Cell-Surfaced Yeast Consortium. Prep. Biochem. Biotechnol. 2018, 48, 653–661. [Google Scholar] [CrossRef]
- Mhatre, A.; Kalscheur, B.; Mckeown, H.; Bhakta, K.; Sarnaik, A.P.; Flores, A.; Nielsen, D.R.; Wang, X.; Soundappan, T.; Varman, A.M. Consolidated Bioprocessing of Hemicellulose to Fuels and Chemicals through an Engineered Bacillus Subtilis-Escherichia Coli Consortium. Renew. Energy 2022, 193, 288–298. [Google Scholar] [CrossRef]
- Infanzón-Rodríguez, M.I.; Ragazzo-Sánchez, J.A.; del Moral, S.; Calderón-Santoyo, M.; Aguilar-Uscanga, M.G. Enzymatic Hydrolysis of Lignocellulosic Biomass Using Native Cellulase Produced by Aspergillus niger ITV02 under Liquid State Fermentation. Biotechnol. Appl. Biochem. 2022, 69, 198–208. [Google Scholar] [CrossRef]
- Kovács, E.; Szűcs, C.; Farkas, A.; Szuhaj, M.; Maróti, G.; Bagi, Z.; Rákhely, G.; Kovács, K.L. Pretreatment of Lignocellulosic Biogas Substrates by Filamentous Fungi. J. Biotechnol. 2022, 360, 160–170. [Google Scholar] [CrossRef]
- Rai, R.; Bibra, M.; Chadha, B.S.; Sani, R.K. Enhanced Hydrolysis of Lignocellulosic Biomass with Doping of a Highly Thermostable Recombinant Laccase. Int. J. Biol. Macromol. 2019, 137, 232–237. [Google Scholar] [CrossRef]
- Jović, J.; Hao, J.; Kocić-Tanackov, S.; Mojović, L. Improvement of Lignocellulosic Biomass Conversion by Optimization of Fungal Ligninolytic Enzyme Activity and Molasses Stillage Supplementation. Biomass Conv. Bioref. 2022, 12, 2749–2765. [Google Scholar] [CrossRef]
- Du, L.; Ma, L.; Ma, Q.; Guo, G.; Han, X.; Xiao, D. Hydrolytic Boosting of Lignocellulosic Biomass by a Fungal Lytic Polysaccharide Monooxygenase, AnLPMO15g from Aspergillus niger. Ind. Crops Prod. 2018, 126, 309–315. [Google Scholar] [CrossRef]
- Shanmugam, S.; Hari, A.; Ulaganathan, P.; Yang, F.; Krishnaswamy, S.; Wu, Y.R. Potential of Biohydrogen Generation Using the Delignified Lignocellulosic Biomass by a Newly Identified Thermostable Laccase from Trichoderma asperellum Strain BPLMBT1. Int. J. Hydrogen Energy 2018, 43, 3618–3628. [Google Scholar] [CrossRef]
- Pimentel, P.S.S.R.; de Oliveira, J.B.; Astolfi-Filho, S.; Pereira, N. Enzymatic Hydrolysis of Lignocellulosic Biomass Using an Optimized Enzymatic Cocktail Prepared from Secretomes of Filamentous Fungi Isolated from Amazonian Biodiversity. Appl. Biochem. Biotechnol. 2021, 193, 3915–3935. [Google Scholar] [CrossRef]
- Chandra Rajak, R.; Banerjee, R. Enzyme Mediated Biomass Pretreatment and Hydrolysis: A Biotechnological Venture towards Bioethanol Production. RSC Adv. 2016, 6, 61301–61311. [Google Scholar] [CrossRef]
- Pereira Scarpa, J.d.C.; Paganini Marques, N.; Alves Monteiro, D.; Martins, G.M.; Paula, A.V.d.; Boscolo, M.; da Silva, R.; Gomes, E.; Alonso Bocchini, D. Saccharification of Pretreated Sugarcane Bagasse Using Enzymes Solution from Pycnoporus sanguineus MCA 16 and Cellulosic Ethanol Production. Ind. Crops Prod. 2019, 141, 111795. [Google Scholar] [CrossRef]
- Schneider, W.D.H.; Fontana, R.C.; Baudel, H.M.; de Siqueira, F.G.; Rencoret, J.; Gutiérrez, A.; de Eugenio, L.I.; Prieto, A.; Martínez, M.J.; Martínez, Á.T.; et al. Lignin Degradation and Detoxification of Eucalyptus Wastes by On-Site Manufacturing Fungal Enzymes to Enhance Second-Generation Ethanol Yield. Appl. Energy 2020, 262, 114493. [Google Scholar] [CrossRef]
- Sheng, T.; Zhao, L.; Gao, L.F.; Liu, W.Z.; Cui, M.H.; Guo, Z.C.; Ma, X.D.; Ho, S.H.; Wang, A.J. Lignocellulosic Saccharification by a Newly Isolated Bacterium, Ruminiclostridium thermocellum M3 and Cellular Cellulase Activities for High Ratio of Glucose to Cellobiose. Biotechnol. Biofuels 2016, 9, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafari, N.; Rezaei, S.; Rezaie, R.; Dilmaghani, H.; Khoshayand, M.R.; Faramarzi, M.A. Improved Production and Characterization of a Highly Stable Laccase from the Halophilic Bacterium Chromohalobacter salexigens for the Efficient Delignification of Almond Shell Bio-Waste. Int. J. Biol. Macromol. 2017, 105, 489–498. [Google Scholar] [CrossRef] [PubMed]
- De La Torre, M.; Martín-Sampedro, R.; Fillat, Ú.; Eugenio, M.E.; Blánquez, A.; Hernández, M.; Arias, M.E.; Ibarra, D. Comparison of the Efficiency of Bacterial and Fungal Laccases in Delignification and Detoxification of Steam-Pretreated Lignocellulosic Biomass for Bioethanol Production. J. Ind. Microbiol. Biotechnol. 2017, 44, 1561–1573. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Liu, J.; Chen, B.; Ran, Q.; Wang, R.; Yu, C.; Ma, X.; Song, H.; Jiang, Z. A Newly Isolated Lignin Degradation Bacterium Acinetobacter Sp. B213 Pretreatment Effectively Improved Hydrolysis of Corn Straw by Enhance Cellulose Retention and Reduce Lignin Degradation Inhibitor. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Wei, Y.; Wu, D.; Wei, D.; Zhao, Y.; Wu, J.; Xie, X.; Zhang, R.; Wei, Z. Improved Lignocellulose-Degrading Performance during Straw Composting from Diverse Sources with Actinomycetes Inoculation by Regulating the Key Enzyme Activities. Bioresour. Technol. 2019, 271, 66–74. [Google Scholar] [CrossRef]
- Ahmed, A.A.Q.; Babalola, O.O.; McKay, T. Cellulase- and Xylanase-Producing Bacterial Isolates with the Ability to Saccharify Wheat Straw and Their Potential Use in the Production of Pharmaceuticals and Chemicals from Lignocellulosic Materials. Waste Biomass Valor. 2018, 9, 765–775. [Google Scholar] [CrossRef]
- Nazar, M.; Xu, L.; Ullah, M.W.; Moradian, J.M.; Wang, Y.; Sethupathy, S.; Iqbal, B.; Nawaz, M.Z.; Zhu, D. Biological Delignification of Rice Straw Using Laccase from Bacillus Ligniniphilus L1 for Bioethanol Production: A Clean Approach for Agro-Biomass Utilization. J. Clean. Prod. 2022, 360, 132171. [Google Scholar] [CrossRef]
- Navas, L.E.; Martínez, F.D.; Taverna, M.E.; Fetherolf, M.M.; Eltis, L.D.; Nicolau, V.; Estenoz, D.; Campos, E.; Benintende, G.B.; Berretta, M.F. A Thermostable Laccase from Thermus Sp. 2.9 and Its Potential for Delignification of Eucalyptus Biomass. AMB Expr. 2019, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Leite, P.; Sousa, D.; Fernandes, H.; Ferreira, M.; Costa, A.R.; Filipe, D.; Gonçalves, M.; Peres, H.; Belo, I.; Salgado, J.M. Recent Advances in Production of Lignocellulolytic Enzymes by Solid-State Fermentation of Agro-Industrial Wastes. Curr. Opin. Green Sustain. Chem. 2021, 27, 100407. [Google Scholar] [CrossRef]
- Abu Yazid, N.; Barrena, R.; Komilis, D.; Sánchez, A. Solid-State Fermentation as a Novel Paradigm for Organic Waste Valorization: A Review. Sustainability 2017, 9, 224. [Google Scholar] [CrossRef]
- Šelo, G.; Planinić, M.; Tišma, M.; Tomas, S.; Koceva Komlenić, D.; Bucić-Kojić, A. A Comprehensive Review on Valorization of Agro-Food Industrial Residues by Solid-State Fermentation. Foods 2021, 10, 927. [Google Scholar] [CrossRef] [PubMed]
- Martău, G.-A.; Unger, P.; Schneider, R.; Venus, J.; Vodnar, D.C.; López-Gómez, J.P. Integration of Solid State and Submerged Fermentations for the Valorization of Organic Municipal Solid Waste. J. Fungi 2021, 7, 766. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, J.; Wang, R.; Liu, L.; Zhang, Y.; Bao, H.; Jang, J.M.; Wang, E.; Yuan, H. Comparative Characterization of Extracellular Enzymes Secreted by Phanerochaete chrysosporium during Solid-State and Submerged Fermentation. Int. J. Biol. Macromol. 2020, 152, 288–294. [Google Scholar] [CrossRef]
- Leite, P.; Silva, C.; Salgado, J.M.; Belo, I. Simultaneous Production of Lignocellulolytic Enzymes and Extraction of Antioxidant Compounds by Solid-State Fermentation of Agro-Industrial Wastes. Ind. Crops Prod. 2019, 137, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.E.; Blánquez, A.; Hernández, M.; Rodríguez, J.; Ball, A.S.; Jiménez-Morillo, N.T.; González-Vila, F.J.; González-Pérez, J.A. Role of a Thermostable Laccase Produced by Streptomyces ipomoeae in the Degradation of Wheat Straw Lignin in Solid State Fermentation. J. Anal. Appl. Pyrolysis 2016, 122, 202–208. [Google Scholar] [CrossRef]
- Sharma, M.; Kumar Bajaj, B. Optimization of Bioprocess Variables for Production of a Thermostable and Wide Range PH Stable Carboxymethyl Cellulase from Bacillus subtilis MS 54 under Solid State Fermentation. Environ. Prog. Sustain. Energy 2017, 36, 1123–1130. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K. Sweet Sorghum Bagasse Pretreatment by Coriolus versicolor in Mesh Tray Bioreactor for Selective Delignification and Improved Saccharification. Waste Biomass Valor. 2019, 10, 2689–2702. [Google Scholar] [CrossRef]
- Filipe, D.; Fernandes, H.; Castro, C.; Peres, H.; Oliva-Teles, A.; Belo, I.; Salgado, J.M. Improved Lignocellulolytic Enzyme Production and Antioxidant Extraction Using Solid-state Fermentation of Olive Pomace Mixed with Winery Waste. Biofuels Bioprod. Bioref. 2020, 14, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Garg, S.; Capalash, N.; Gupta, N.; Sharma, P. Production of Thermo-Alkali-Stable Laccase and Xylanase by Co-Culturing of Bacillus Sp. and B. halodurans for Biobleaching of Kraft Pulp and Deinking of Waste Paper. Bioprocess Biosyst. Eng. 2015, 38, 947–956. [Google Scholar] [CrossRef]
- David, A.; Singh Chauhan, P.; Kumar, A.; Angural, S.; Kumar, D.; Puri, N.; Gupta, N. Coproduction of Protease and Mannanase from Bacillus nealsonii PN-11 in Solid State Fermentation and Their Combined Application as Detergent Additives. Int. J. Biol. Macromol. 2018, 108, 1176–1184. [Google Scholar] [CrossRef]
- Ozcirak Ergun, S.; Ozturk Urek, R. Production of Ligninolytic Enzymes by Solid State Fermentation Using Pleurotus ostreatus. Ann. Agrar. Sci. 2017, 15, 273–277. [Google Scholar] [CrossRef]
- Khanahmadi, M.; Arezi, I.; Amiri, M.; Miranzadeh, M. Bioprocessing of Agro-Industrial Residues for Optimization of Xylanase Production by Solid- State Fermentation in Flask and Tray Bioreactor. Biocatal. Agric. Biotechnol. 2018, 13, 272–282. [Google Scholar] [CrossRef]
- Rayhane, H.; Josiane, M.; Gregoria, M.; Yiannis, K.; Nathalie, D.; Ahmed, M.; Sevastianos, R. From Flasks to Single Used Bioreactor: Scale-up of Solid State Fermentation Process for Metabolites and Conidia Production by Trichoderma asperellum. J. Environ. Manag. 2019, 252, 109496. [Google Scholar] [CrossRef] [PubMed]
- Achappa, S.; Shet, A.R.; Hombalimath, V.S.; Patil, L.R.; Desai, S.V. Bioprocessing of Agricultural Waste (Banana Pseudostem) by Solid-State Fermentation (SSF) for Production of Cellulase. J. Pharm. Res. Int. 2022, 15–26. [Google Scholar] [CrossRef]
- Ferreira, R.G.; Azzoni, A.R.; Freitas, S. On the Production Cost of Lignocellulose-degrading Enzymes. Biofuels Bioprod. Bioref. 2021, 15, 85–99. [Google Scholar] [CrossRef]
- Ramos, M.D.N.; Milessi, T.S.; Candido, R.G.; Mendes, A.A.; Aguiar, A. Enzymatic Catalysis as a Tool in Biofuels Production in Brazil: Current Status and Perspectives. Energy Sustain. Dev. 2022, 68, 103–119. [Google Scholar] [CrossRef]
- de Lima, E.A.; Mandelli, F.; Kolling, D.; Matsusato Souza, J.; de Oliveira Filho, C.A.; Ribeiro da Silva, M.; Lobo de Mesquita Sampaio, I.; Lopes Junqueira, T.; Ferreira Chagas, M.; Teodoro, J.C.; et al. Development of an Economically Competitive Trichoderma-Based Platform for Enzyme Production: Bioprocess Optimization, Pilot Plant Scale-up, Techno-Economic Analysis and Life Cycle Assessment. Bioresour. Technol. 2022, 364, 128019. [Google Scholar] [CrossRef]
- Rajnish, K.N.; Samuel, M.S.; John J, A.; Datta, S.; Chandrasekar, N.; Balaji, R.; Jose, S.; Selvarajan, E. Immobilization of Cellulase Enzymes on Nano and Micro-Materials for Breakdown of Cellulose for Biofuel Production-a Narrative Review. Int. J. Biol. Macromol. 2021, 182, 1793–1802. [Google Scholar] [CrossRef]
- Bernal, C.; Rodríguez, K.; Martínez, R. Integrating Enzyme Immobilization and Protein Engineering: An Alternative Path for the Development of Novel and Improved Industrial Biocatalysts. Biotechnol. Adv. 2018, 36, 1470–1480. [Google Scholar] [CrossRef]
- Bashir, N.; Sood, M.; Bandral, J.D. Enzyme Immobilization and Its Applications in Food Processing: A Review. Int. J. Chem. Stud. 2020, 8, 254–261. [Google Scholar] [CrossRef]
- Ghosh, P.; Ghosh, U. Immobilization of Purified Fungal Laccase on Cost Effective Green Coconut Fiber and Study of Its Physical and Kinetic Characteristics in Both Free and Immobilized Form. Curr. Biotechnol. 2019, 8, 3–14. [Google Scholar] [CrossRef]
- Aarti, C.; Khusro, A.; Agastian, P. Saccharification of Alkali Pre-Treated Aquatic Weeds Biomass Using Partially Purified Cellulase Immobilized on Different Matrices. Biocatal. Agric. Biotechnol. 2022, 39, 102283. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.; Zhang, X.; Wu, J.; Chai, C.; Ma, D.; Chen, Q.; Xiang, D.; Ge, W. Immobilized Lignin Peroxidase on Fe3O4@SiO2@polydopamine Nanoparticles for Degradation of Organic Pollutants. Int. J. Biol. Macromol. 2019, 138, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Murguiondo, C.; Mestre, A.; Méndez-Líter, J.A.; Nieto-Domínguez, M.; de Eugenio, L.I.; Molina-Gutiérrez, M.; Martínez, M.J.; Prieto, A. Enzymatic Glycosylation of Bioactive Acceptors Catalyzed by an Immobilized Fungal β-Xylosidase and Its Multi-Glycoligase Variant. Int. J. Biol. Macromol. 2021, 167, 245–254. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, P.; Sharma, V.; Koul, S.; Kour, K.; Bajaj, B.K. Multifarious Potential Applications of keratinase of Bacillus subtilis K-5. Biocatal. Biotransformation 2014, 32, 333–342. [Google Scholar] [CrossRef]
- Sharma, S.; Tsai, M.-L.; Sharma, V.; Sun, P.-P.; Nargotra, P.; Bajaj, B.K.; Chen, C.-W.; Dong, C.-D. Environment Friendly Pretreatment Approaches for the Bioconversion of Lignocellulosic Biomass into Biofuels and Value-Added Products. Environments 2023, 10, 6. [Google Scholar] [CrossRef]
- Sharma, V.; Nargotra, P.; Sharma, S.; Bajaj, B.K. Efficient Bioconversion of Sugarcane Tops Biomass into Biofuel-Ethanol Using an Optimized Alkali-Ionic Liquid Pretreatment Approach. Biomass Conv. Bioref. 2020, 10, 1–14. [Google Scholar] [CrossRef]



| Microorganism | Enzyme | Lignocellulosic Substrate | Enzyme Activity | Reference |
|---|---|---|---|---|
| Coriolus versicolor | Lignin peroxidase, laccase, polyphenol peroxidase, manganese peroxidase, aryl alcohol oxidase and xylanase | Sweet sorghum bagasse | Laccase-115.1 ± 7.1 U/g, Lignin peroxidase-2.86 ± 0.1 U/g, Manganese peroxidase-11.1 ± 1.0 U/g, Xylanase-13.1 ± 0.9 U/g, Polyphenol peroxidase-6.2 ± 0.4 U/g, Aryl alcohol oxidase-3.1 ± 0.2 U/g, | [144] |
| Aspergillus ibericus | Xylanase, cellulase and β-glucosidase | Olive mill and winery wastes | Xylanase-96.4 U/g, Cellulase- 84.2 U/g, β-glucosidase-25.5 U/g | [145] |
| Aspergillus niger | Xylanase, cellulase and β-glucosidase | Olive mill and winery wastes | Xylanase-129.4 U/g, Cellulase-38.8 U/g, β-glucosidase-17.9 U/g | [145] |
| Bacillus halodurans FNP 135 and Bacillus sp. | Xylanase, laccase | Wheat bran | Xylanase-1685 IU/g, Laccase-2270 nkat/g | [146] |
| Bacillus nealsonii PN-11 | Mannanase | Wheat bran | Mannanase-834 U/g | [147] |
| Pleurotus ostreatus | Laccase, manganese peroxidase, lignin peroxidase and aryl alcohol oxidase | Potato peel waste | Manganese peroxidase-2503.6 ± 5 U/L, Laccase-6708.3 ± 75 U/L, Lignin peroxidase-231.2 ± 9 U/L, Aryl alcohol oxidase-677.4 ± 17 U/L | [148] |
| Aspergillus niger CCUG33991 | Xylanase | Wheat bran, sorghum stover, corn cob and soybean meal | Wheat bran-1137 ± 104 U/g, sorghum stover-257 ± 35 U/g, corn cob-380 ± 25 U/g, soybean meal-365 ± 20 U/g | [149] |
| Trichoderma asperellum TF1 | Cellulase | Mixture of jatropha cake, vine shoots, olive oil, olive pomace | Cellulase-10.68 ± 0.21 U/g biomass | [150] |
| Cellulomonas uda | Cellulase | Banana stem waste | Cellulase-6.97 IU/mL | [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nargotra, P.; Sharma, V.; Lee, Y.-C.; Tsai, Y.-H.; Liu, Y.-C.; Shieh, C.-J.; Tsai, M.-L.; Dong, C.-D.; Kuo, C.-H. Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review. Catalysts 2023, 13, 83. https://doi.org/10.3390/catal13010083
Nargotra P, Sharma V, Lee Y-C, Tsai Y-H, Liu Y-C, Shieh C-J, Tsai M-L, Dong C-D, Kuo C-H. Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review. Catalysts. 2023; 13(1):83. https://doi.org/10.3390/catal13010083
Chicago/Turabian StyleNargotra, Parushi, Vishal Sharma, Yi-Chen Lee, Yung-Hsiang Tsai, Yung-Chuan Liu, Chwen-Jen Shieh, Mei-Ling Tsai, Cheng-Di Dong, and Chia-Hung Kuo. 2023. "Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review" Catalysts 13, no. 1: 83. https://doi.org/10.3390/catal13010083
APA StyleNargotra, P., Sharma, V., Lee, Y.-C., Tsai, Y.-H., Liu, Y.-C., Shieh, C.-J., Tsai, M.-L., Dong, C.-D., & Kuo, C.-H. (2023). Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review. Catalysts, 13(1), 83. https://doi.org/10.3390/catal13010083