
Characterization and Structural Insights of a Novel Arylsulfatase from Pseudoalteromonas atlantica T6c

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
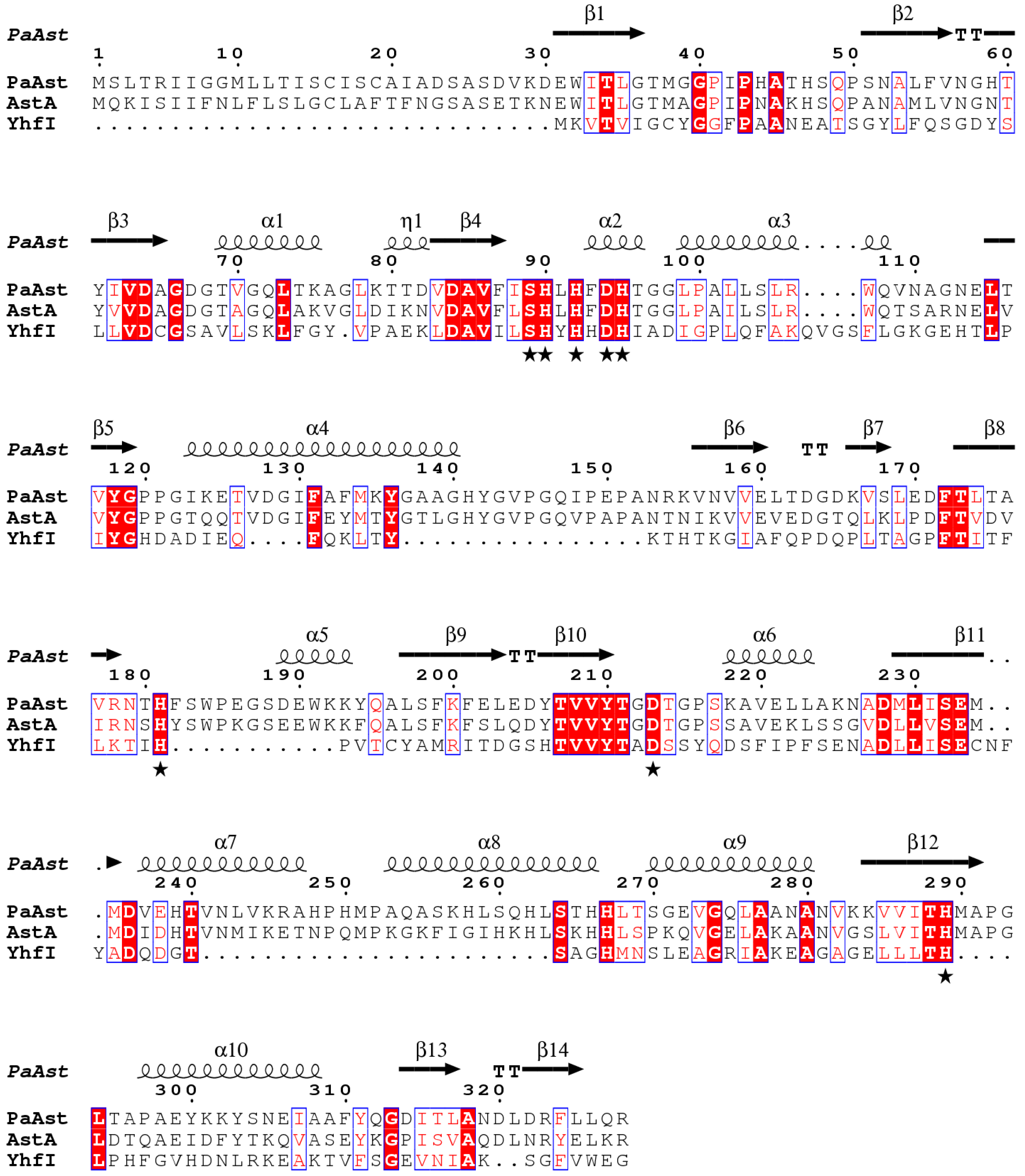
2.1. Sequence Analysis of PaAst
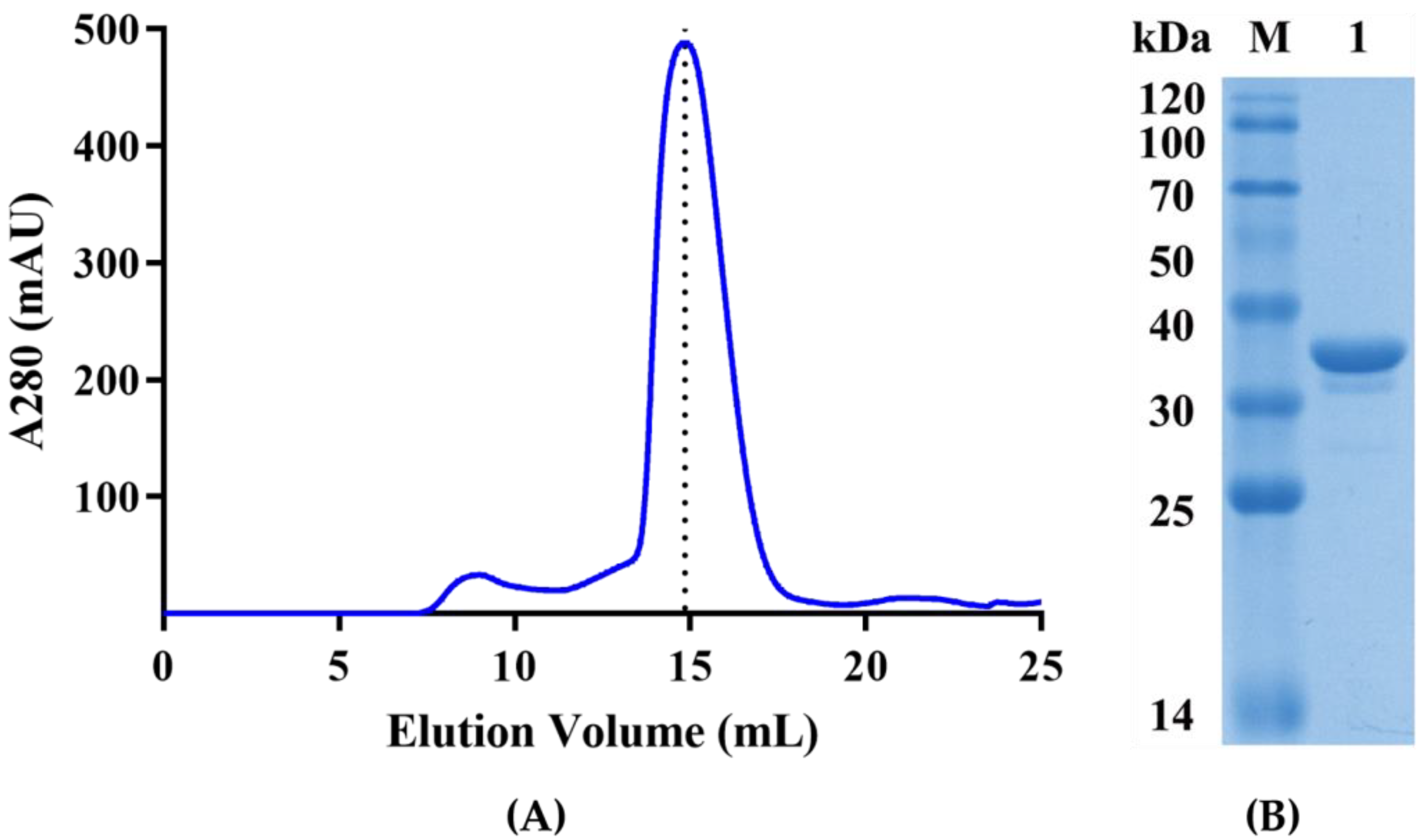
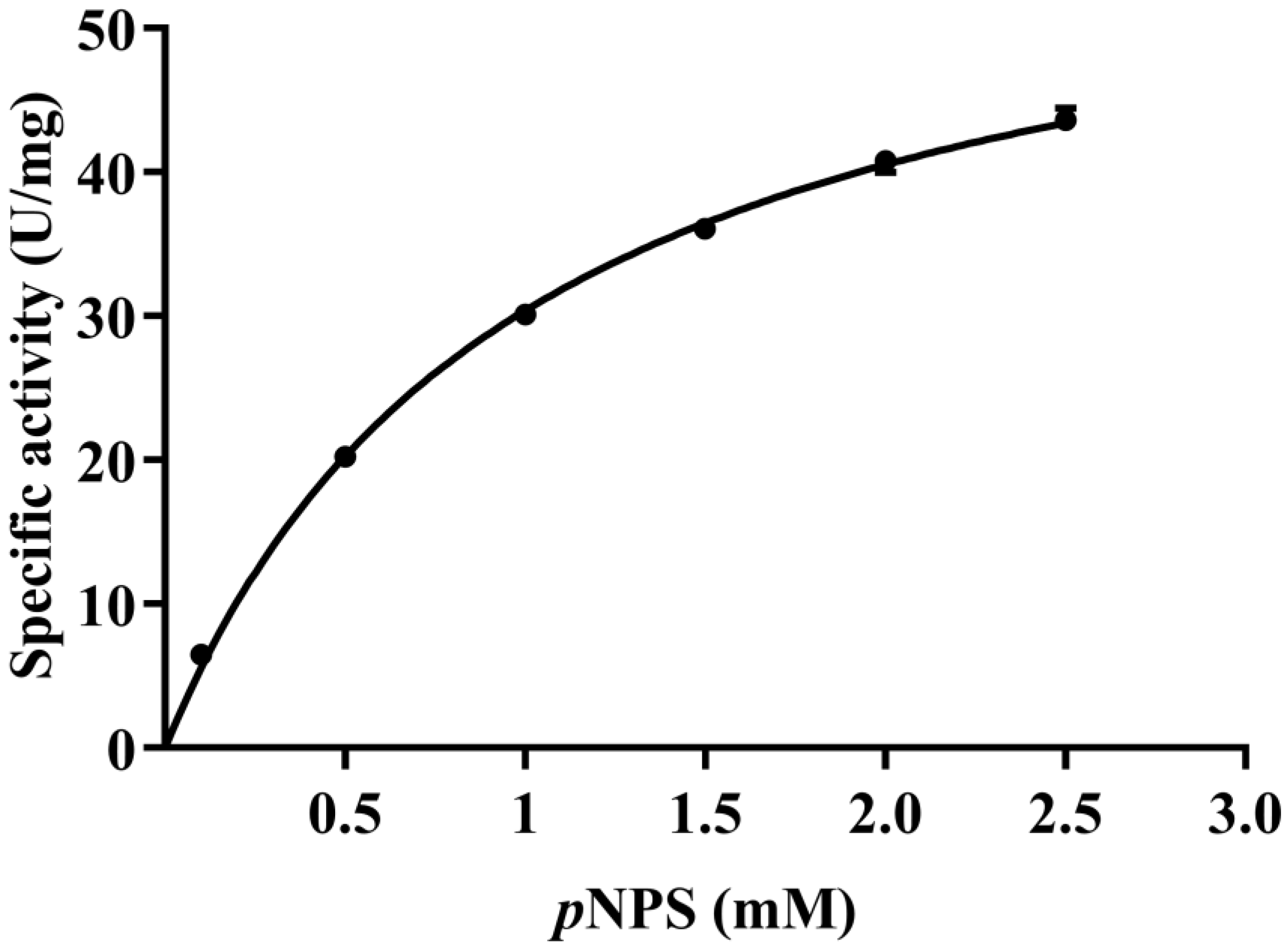
2.2. Expression, Purification, and Characterization of Recombinant PaAst
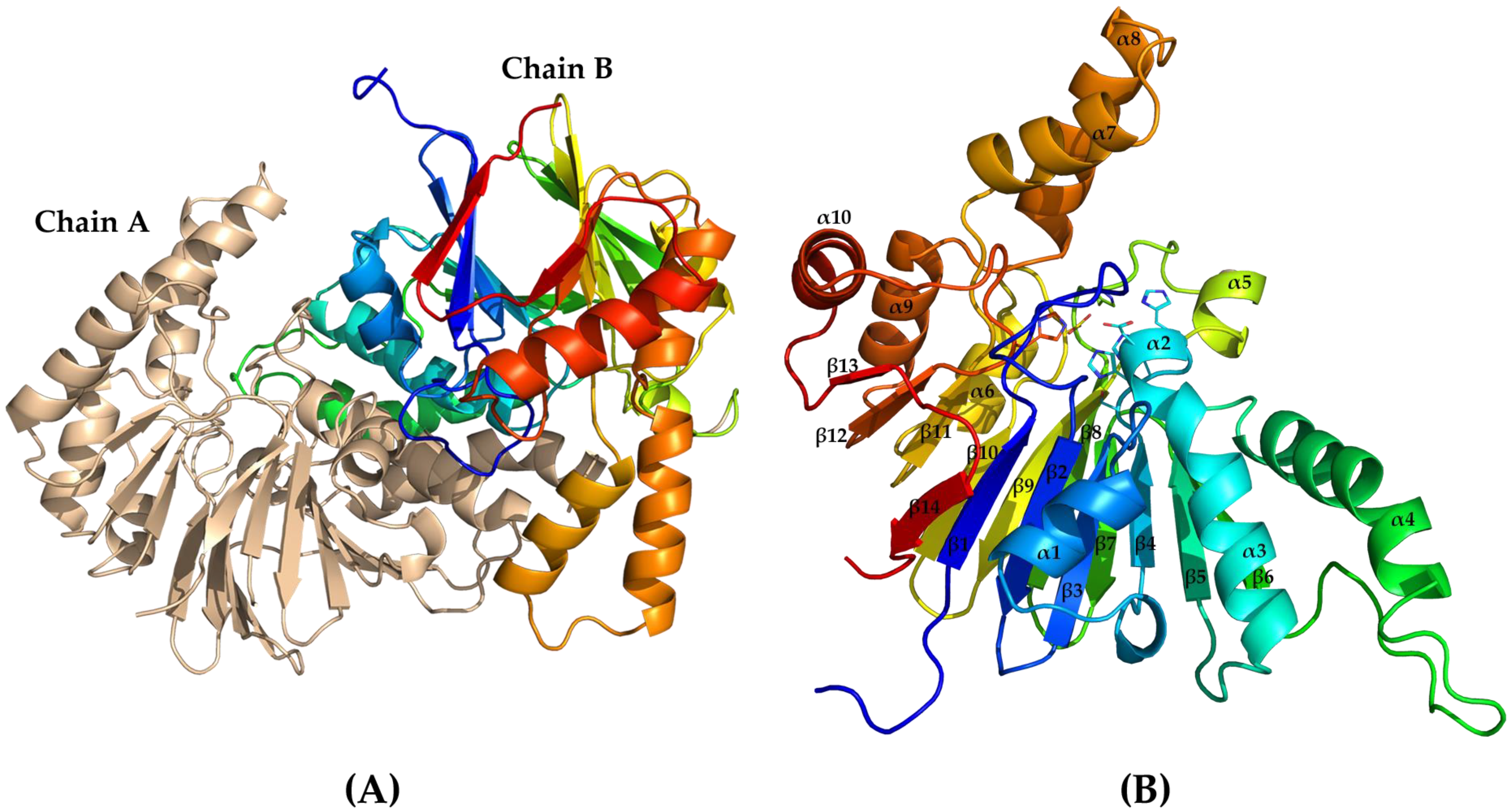
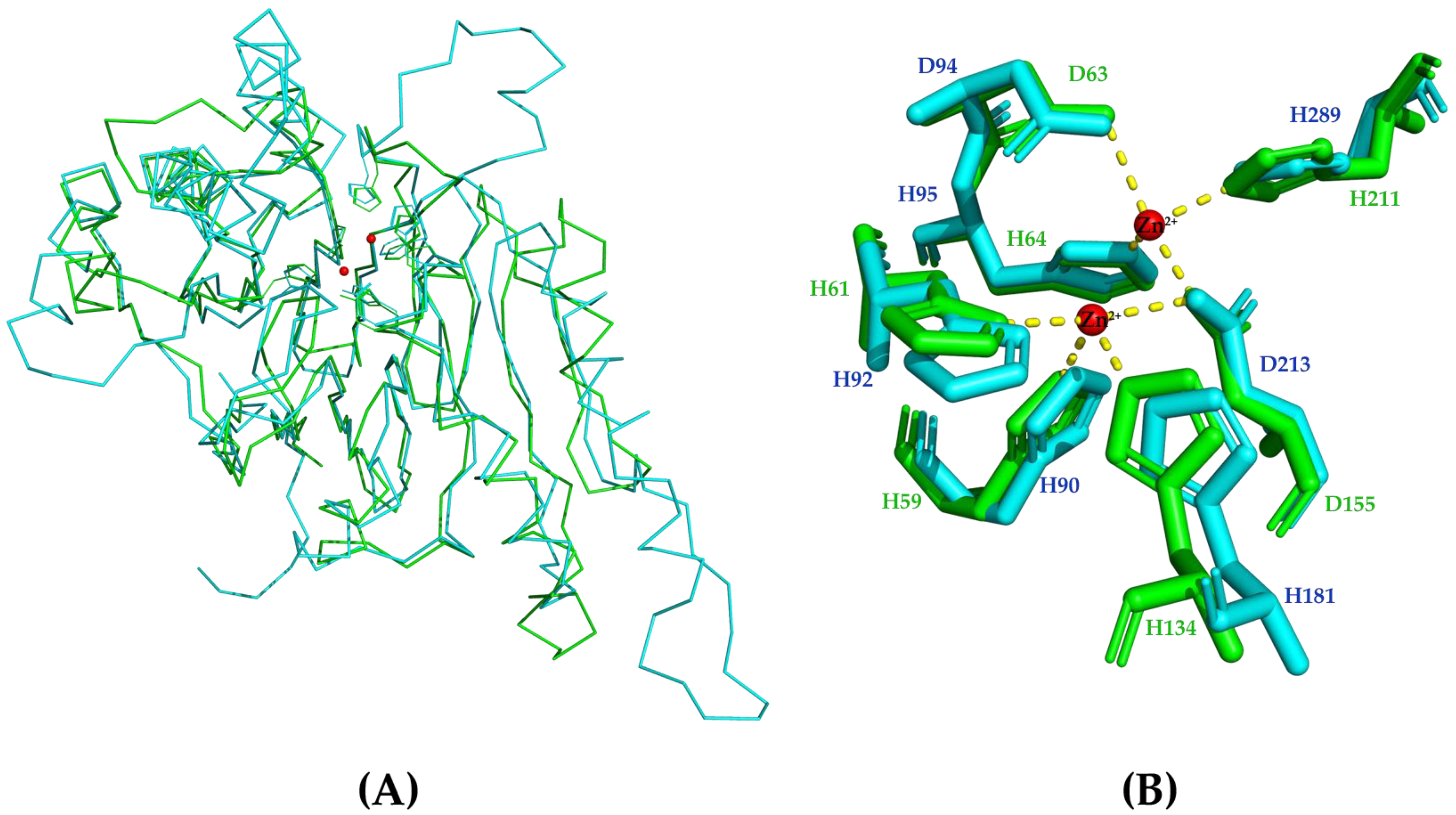
2.3. Crystal Structure of PaAst
3. Materials and Methods
3.1. Strains, Plasmids, and Reagents
3.2. Sequence Analysis
3.3. Protein Expression and Purification
3.4. Enzymatic Activity Assay
3.5. Crystallization
3.6. Structure Determination and Refinement
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value(s) |
|---|---|
| Data collection | |
| Beamline | SSRF BL18U1 |
| Wavelength (Å) | 0.97915 |
| Space group | P 1 21 1 |
| Unit cell parameters | |
| a, b, c (Å) | 55.17, 61.38, 89.60 |
| α, β, γ (°) | 90, 97.88, 90 |
| Resolution range (Å) | 38.61–1.55 (1.59–1.55) |
| Completeness (%) | 99.7 |
| No. of unique reflections measured | 85,833 |
| Redundancy | 6.1 (6.5) |
| Rmerge | 0.117 (1.077) |
| <I/σ(I)> | 8.3 (1.8) |
| CC1/2 | 0.995 (0.528) |
| Refinement | |
| Resolution range (Å) | 29.59–2.0 |
| Completeness (%) | 99.5 |
| No. of unique reflections refined | 40,070 |
| Rwork/Rfree (%) | 20.04/24.35 |
| No. of molecules per asymmetric unit | 2 |
| No. of residues | Chain A: 291 Chain B: 305 |
| No. of atoms | 5202 |
| Ramachandran Plot (%) | |
| Favored regions | 97.63 |
| Allowed regions | 2.03 |
| Outliers | 0.34 |
| Mean B value, overall (Å2) | 23.9 |
| PDB ID | 8GYG |
| Purification Procedures | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Factor | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 136.07 | 5000.57 | 36.75 | 1 | 100 |
| Affinity chromatography | 48.88 | 2099.67 | 42.96 | 1.17 | 41.98 |
| Size-exclusion chromatography | 6.39 | 500.15 | 78.32 | 2.13 | 10.00 |
References
- Pogorevc, M.; Faber, K. Purification and characterization of an inverting stereo- and enantioselective sec-alkylsulfatase from the gram-positive bacterium Rhodococcus ruber DSM 44541. Appl. Environ. Microbiol. 2003, 69, 2810–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.; Ruan, L.; Li, F.; Yu, Z.; Xu, X. Heterologous expression and characterization of a recombinant thermostable alkylsulfatase (sdsAP). Extremophiles 2011, 15, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, H.; Qiao, C.; Li, L.; Jiang, Z.; Xiao, A.; Ni, H. Characterization of an arylsulfatase from a mutant library of Pseudoalteromonas carrageenovora arylsulfatase. Int. J. Biol. Macromol. 2017, 96, 370–376. [Google Scholar] [CrossRef]
- Hagelueken, G.; Adams, T.M.; Wiehlmann, L.; Widow, U.; Kolmar, H.; Tummler, B.; Heinz, D.W.; Schubert, W.D. The crystal structure of SdsA1, an alkylsulfatase from Pseudomonas aeruginosa, defines a third class of sulfatases. Proc. Natl. Acad. Sci. USA 2006, 103, 7631–7636. [Google Scholar] [CrossRef] [Green Version]
- Berteau, O.; Guillot, A.; Benjdia, A.; Rabot, S. A new type of bacterial sulfatase reveals a novel maturation pathway in prokaryotes. J. Biol. Chem. 2006, 281, 22464–22470. [Google Scholar] [CrossRef] [Green Version]
- Sogi, K.M.; Gartner, Z.J.; Breidenbach, M.A.; Appel, M.J.; Schelle, M.W.; Bertozzi, C.R. Mycobacterium tuberculosis Rv3406 is a type II alkyl sulfatase capable of sulfate scavenging. PLoS ONE 2013, 8, e65080. [Google Scholar] [CrossRef]
- Muller, I.; Kahnert, A.; Pape, T.; Sheldrick, G.M.; Meyer-Klaucke, W.; Dierks, T.; Kertesz, M.; Uson, I. Crystal structure of the alkylsulfatase AtsK: Insights into the catalytic mechanism of the Fe(II) alpha-ketoglutarate-dependent dioxygenase superfamily. Biochemistry 2004, 43, 3075–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahnert, A.; Kertesz, M.A. Characterization of a sulfur-regulated oxygenative alkylsulfatase from Pseudomonas putida S-313. J. Biol. Chem. 2000, 275, 31661–31667. [Google Scholar] [CrossRef] [Green Version]
- Davison, J.; Brunel, F.; Phanopoulos, A.; Prozzi, D.; Terpstra, P. Cloning and sequencing of Pseudomonas genes determining sodium dodecyl sulfate biodegradation. Gene 1992, 114, 19–24. [Google Scholar] [CrossRef]
- Sun, L.; Chen, P.; Su, Y.; Cai, Z.; Ruan, L.; Xu, X.; Wu, Y. Crystal structure of thermostable alkylsulfatase SdsAP from Pseudomonas sp. S9. Biosci. Rep. 2017, 37, BSR20170001. [Google Scholar] [CrossRef] [Green Version]
- Knaus, T.; Schober, M.; Kepplinger, B.; Faccinelli, M.; Pitzer, J.; Faber, K.; Macheroux, P.; Wagner, U. Structure and mechanism of an inverting alkylsulfatase from Pseudomonas sp. DSM6611 specific for secondary alkyl sulfates. FEBS J. 2012, 279, 4374–4384. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Kim, K.H.; Bae, Y.J.; Lee, J.H.; Jang, Y.H.; Nam, S.W. Purification and characterization of the recombinant arylsulfatase cloned from Pseudoalteromonas carrageenovora. Protein Expr. Purif. 2005, 39, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liang, M.; Li, H.; Ni, H.; Li, L.; Li, Q.; Jiang, Z. A mutant of Pseudoalteromonas carrageenovora arylsulfatase with enhanced enzyme activity and its potential application in improvement of the agar quality. Food Chem. 2020, 320, 126652. [Google Scholar] [CrossRef]
- Jung, K.T.; Kim, H.W.; You, D.J.; Nam, S.W.; Kim, B.W.; Jeon, S.J. Identification of the first archaeal arylsulfatase from Pyrococcus furiosus and its application to desulfatation of agar. Biotechnol. Bioprocess Eng. 2013, 17, 1140–1146. [Google Scholar] [CrossRef]
- Shahbazi, R.; Kasra-Kermanshahi, R.; Gharavi, S.; Moosavi-Nejad, Z.; Borzooee, F. Screening of SDS-degrading bacteria from car wash wastewater and study of the alkylsulfatase enzyme activity. Iran. J. Microbiol. 2013, 5, 153–158. [Google Scholar] [PubMed]
- Zhang, C.; An, D.; Xiao, Q.; Chen, F.Q.; Zhang, Y.H.; Weng, H.F.; Xiao, A.F. Convenient Agarose Preparation with Hydrogen Peroxide and Desulfation Process Analysis. Mar. Drugs 2021, 19, 297. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Deng, X.; Lin, J.; Yang, J. Characterization and substrate-accelerated thermal inactivation kinetics of a new serine-type arylsulfatase. Enzyme Microb. Technol. 2022, 154, 109961. [Google Scholar] [CrossRef] [PubMed]
- Na, H.W.; Namgung, B.; Song, W.S.; Yoon, S.I. Structural and biochemical analyses of the metallo-beta-lactamase fold protein YhfI from Bacillus subtilis. Biochem. Biophys. Res. Commun. 2019, 519, 35–40. [Google Scholar] [CrossRef]
- Kertesz, M.A. Riding the sulfur cycle--metabolism of sulfonates and sulfate esters in gram-negative bacteria. FEMS Microbiol. Rev. 2000, 24, 135–175. [Google Scholar] [CrossRef] [Green Version]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Holm, L. Dali server: Structural unification of protein families. Nucleic Acids Res. 2022, 50, W210–W215. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, O.; Li de la Sierra-Gallay, I.; Piton, J.; Gilet, L.; Condon, C. Activation of tRNA maturation by downstream uracil residues in B. subtilis. Structure 2012, 20, 1769–1777. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Chen, Y.; Isupov, M.N.; Littlechild, J.A.; Sun, L.; Liu, X.; Wang, Q.; Gong, H.; Dong, P.; Zhang, N.; et al. Structural Insights into a Novel Esterase from the East Pacific Rise and Its Improved Thermostability by a Semirational Design. J. Agric. Food Chem. 2021, 69, 1079–1090. [Google Scholar] [CrossRef]
- Yang, W.; Sun, L.; Dong, P.; Chen, Y.; Zhang, H.; Huang, X.; Wu, L.; Chen, L.; Jing, D.; Wu, Y. Structure-guided rational design of the Geobacillus thermoglucosidasius feruloyl esterase GthFAE to improve its thermostability. Biochem. Biophys. Res. Commun. 2022, 600, 117–122. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Mitteer, D.R.; Greer, B.D.; Randall, K.R.; Briggs, A.M. Further Evaluation of Teaching Behavior Technicians to Input Data and Graph Using GraphPad Prism. Behav. Anal. 2020, 20, 81–93. [Google Scholar] [CrossRef]
- Minor, W.; Cymborowski, M.; Borek, D.; Cooper, D.R.; Chruszcz, M.; Otwinowski, Z. Optimal structure determination from sub-optimal diffraction data. Protein Sci. 2022, 31, 259–268. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- Kabsch, W. Xds. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| Purification Procedures | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Factor | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 220.32 | 2613.00 | 11.86 | 1 | 100 |
| Affinity chromatography | 22.93 | 840.21 | 36.64 | 3.08 | 32.16 |
| Size-exclusion chromatography | 9.77 | 385.62 | 39.45 | 3.33 | 14.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, P.; Yang, W.; Sun, L.; Jing, D.; Zhang, H.; Yang, J.; Wu, L.; Chen, L.; Wu, Y. Characterization and Structural Insights of a Novel Arylsulfatase from Pseudoalteromonas atlantica T6c. Catalysts 2023, 13, 759. https://doi.org/10.3390/catal13040759
Dong P, Yang W, Sun L, Jing D, Zhang H, Yang J, Wu L, Chen L, Wu Y. Characterization and Structural Insights of a Novel Arylsulfatase from Pseudoalteromonas atlantica T6c. Catalysts. 2023; 13(4):759. https://doi.org/10.3390/catal13040759
Chicago/Turabian StyleDong, Panpan, Wendi Yang, Lifang Sun, Dingding Jing, Hong Zhang, Jinbo Yang, Linjiao Wu, Leiqing Chen, and Yunkun Wu. 2023. "Characterization and Structural Insights of a Novel Arylsulfatase from Pseudoalteromonas atlantica T6c" Catalysts 13, no. 4: 759. https://doi.org/10.3390/catal13040759
APA StyleDong, P., Yang, W., Sun, L., Jing, D., Zhang, H., Yang, J., Wu, L., Chen, L., & Wu, Y. (2023). Characterization and Structural Insights of a Novel Arylsulfatase from Pseudoalteromonas atlantica T6c. Catalysts, 13(4), 759. https://doi.org/10.3390/catal13040759