Abstract
In higher plants, the lipoxygenase enzymatic pathway combined actions of several enzymes to convert lipid substrates into signaling and defense molecules called phytooxylipins including short chain volatile aldehydes, alcohols, and esters, known as green leaf volatiles (GLVs). GLVs are synthesized from C18:2 and C18:3 fatty acids that are oxygenated by lipoxygenase (LOX) to form corresponding hydroperoxides, then the action of hydroperoxide lyase (HPL) produces C6 or C9 aldehydes that can undergo isomerization, dehydrogenation, and esterification. GLVs are commonly used as flavors to confer a fresh green odor of vegetable to perfumes, cosmetics, and food products. Given the increasing demand in these natural flavors, biocatalytic processes using the LOX pathway reactions constitute an interesting application. Vegetable oils, chosen for their lipid profile are converted in natural GLVs with high added value. This review describes the enzymatic reactions of GLVs biosynthesis in the plant, as well as the structural and functional properties of the enzymes involved. The various stages of the biocatalytic production processes are approached from the lipid substrate to the corresponding aldehyde or alcoholic aromas, as well as the biotechnological improvements to enhance the production potential of the enzymatic catalysts.
1. Introduction
Green leaf volatiles (GLVs) are important contributors to the characteristic flavors of fruits, vegetables, and green leaves [1,2,3]. GLVs include short carbon chain aldehydes, alcohols, and esters produced in higher plants during the lipoxygenase (LOX) pathway [1,2,3,4,5,6]. The LOX pathway is activated during plant development or in response to biotic or abiotic stresses and implements the sequential action of several enzymes to transform polyunsaturated fatty acids (PUFAs) into signaling and defense molecules called phytooxylipins [5,6,7,8,9]. Phytooxylipins include, in particular jasmonates, traumatic acid, epoxy-hydroxy fatty acids, divinyl ethers, as well as GLVs [2,3,6,8,10,11]. GLVs are metabolized from C18-PUFAs. First, the LOX catalyzes regiospecific dioxygenation of C18-PUFAs, including linoleic and linolenic acids, to form 9- or 13-PUFA hydroperoxides. Subsequently, hydroperoxide lyase (HPL) acts on these PUFA hydroperoxides to produce volatile C6 or C9 aldehydes and a 9- or 12-oxo acid. Finally, isomerization, dehydrogenation, and esterification reactions of aldehydes can occur to produce aldehydes with different isomeric forms, alcohols, and esters.
Saturated and unsaturated volatile C6 and C9 aldehydes, alcohols, and their esters are associated with the green note odor and are widely used as aromas and food additives in the cosmetics and perfumes industry as well as in the food industry.
Their industrial production by chemical synthesis is environmentally unfriendly. Indeed, the processes use metal catalysts and are based on naphtha which arise from fractional distillation petroleum. Thus, it is desirable to switch to bioproduction including extraction from natural sources and bioconversion of natural precursor using enzymes. In addition, today’s consumers have a strong preference for naturally synthesized additives and flavors.
Given the high demand for such natural flavors and the difficulty of extracting compounds from plants, efficient biocatalytic processes using the enzymes of the LOX pathway have to be developed for large scale production. In these processes, vegetal oils containing C18:2 and C18:3 fatty acids are hydrolyzed by a lipase, and then the PUFAs released are converted by LOX and HPL into green note aldehydes. Large-scale conversion of triacylglycerols from vegetable oils into hydroperoxides has been successfully accomplished [12,13]. Since lipases and LOXs are commercially available, genetic engineering approaches focus on the production of recombinant HPL, the limiting enzyme for biocatalytic processes.
Thus, interest in the lipoxygenase pathway and its products is growing, not only from a physiological point of view but also from a technological point of view. In this review, first the steps of the biosynthesis of GLV by the LOX pathway in the plant is studied as well as the enzymes involved, and then to approach the use of enzymes of the LOX pathway as biotechnological tools for the industrial production of natural GLVs.
2. The Lipoxygenase Pathway in Higher Plants: Activation and Description of the Enzymatic Cascade
The presence in plants of an enzymatic mechanism responsible for the oxidation of lipids has been known since 1932 [14]. This metabolic pathway is now commonly referred as the lipoxygenase pathway.
The first step, in the LOX pathway is the action of a lipoxygenase on polyunsaturated fatty acids, preferably linoleic acid (LA, 18:2, ω6) and α-linolenic acid (ALA, 18:3, ω3) [15,16]. Generally, these PUFAs have been previously released from membrane phospholipids or galactolipids by phospholipases or galactolipases [3,8,17]. LOX catalyzes regiospecific dioxygenation of the position 9 or 13, of the C18 PUFAs, either LA or ALA to form relevant 9- or 13-hydroperoxyoctadecadi(tri)enoic acids [6,18]. PUFA hydroperoxides are highly reactive molecules and are readily converted by various enzymes into a series of signaling and defense molecules called phytooxylipins [7,10,11,15,19] (Figure 1).
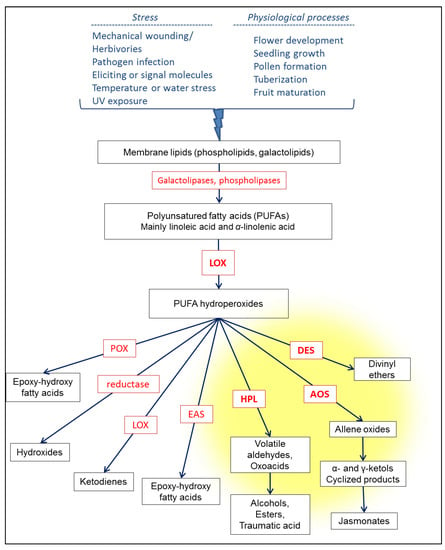
Figure 1.
The lipoxygenase pathway in plants. The substrates and products of the reactions are represented in black, and the enzymes involved appear in red (allene oxide synthase (AOS); divinyl ether synthase (DES); epoxide alcohol synthase (EAS); hydroperoxide lyase (HPL); lipoxygenase (LOX); and peroxygenase (POX)). The reactions catalyzed by the enzymes of the CYP74 subfamily are stained yellow.
PUFA hydroperoxides are mainly metabolized by three enzymes: Allene oxide synthase (AOS), divinyl ether synthase (DES), and hydroperoxide lyase, grouped together in the CYP74 subfamily of cytochromes P450 (Figure 2). AOS dehydrates the PUFA hydroperoxides to unstable allene oxides, which are cyclized by allene oxide cyclase (AOC), or spontaneously hydrolyzed into α- and γ-ketols [20,21,22]. The cyclized product of AOC derived from ALA, 12-oxo-(10Z,15Z)-phytodienoic acid (12-OPDA), can be reduced and then undergo three cycles of β-oxidation to form jasmonic acid (JA) and its derivatives, including methyl jasmonate (MeJA) [20,21,22,23,24]. DES synthesizes divinyl ethers of PUFAs such as colneleic or colnelenic acids and etheroleic or etherolenic acids [25,26]. The action of HPL leads to the cleavage of the C-C backbone of the PUFA hydroperoxides yielding C6 or C9 volatile aldehydes and 9-oxononanoic acid or 12-oxo-(9Z)-dodecenoic acid, a precursor of traumatin [20,27,28,29]. The volatile aldehydes can then be converted into the corresponding alcohols or esters. Other enzymes also metabolize PUFA hydroperoxides produced by LOX: A reductase can reduce the PUFA hydroperoxides to fatty acid hydroxides [30], which are less reactive, a peroxygenase (POX) or an epoxy alcohol synthase (EAS) can transform the PUFA hydroperoxides to epoxy-hydroxy fatty acids [31], and, under oxygen-poor conditions, LOX can catalyze cleavage of the O-O bond of the PUFA hydroperoxides to form alkoxy radicals which rearrange themselves in ketodienes [32].
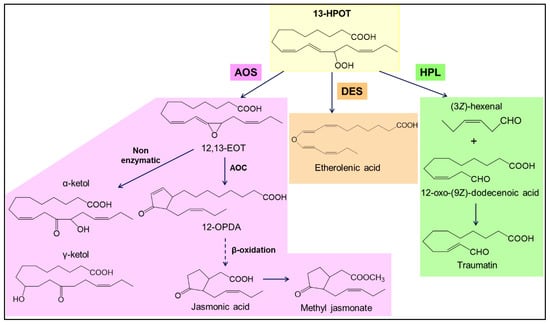
Figure 2.
Phytooxylipins produced from 13-hydroperoxy-(9Z,11E,15Z)-octadecatrienoic acid (13-HPOT) by the enzymes of the CYP74 family. 13-HPOT can be metabolized by CYP74 family enzymes. AOS produces an unstable allene oxide: (12,13)-epoxyoctadecatrienoic acid (12,13-EOT), which can hydrolyze spontaneously to α- and γ-ketols, or be cyclized by allene oxide cyclase (AOC) to 12-Oxo-(10Z,15Z)-phytodienoic acid (12-OPDA), which can be converted into jasmonates after three cycles of β-oxidation. DES generates the etherolenic acid. HPL produces an aldehyde: (3Z)-hexenal, and an oxoacid: 12-oxo-(9Z)-dodecenoic acid which is the precursor of traumatin.
The regulation of these various enzymatic branches of the LOX pathway competing for the same PUFA hydroperoxides constitutes an important control point for the plant. The reaction cascade triggered in response to these stimuli leads to the production of phytooxylipins adapted to fight the stress undergone.
In the plant, the LOX pathway can be induced by a variety of biotic stresses such as wounding caused by herbivorous insects or animals, and fungal or bacterial pathogen infection [3,5,6,9,30,33,34,35]. Plants can detect the presence of a pathogen or herbivore through the intermediary of specific eliciting molecules [36]. Abiotic stresses including mechanical wounding, water deficiency and osmotic stress, UV exposure, suboptimal light, and temperature, can also trigger the biosynthesis of oxylipins [37,38,39,40,41,42].
Several studies have highlighted the role of oxylipins in plant defense against a variety of pathogens. Prost et al. [43] screened numerous oxylipins for antimicrobial activity and found that most of the oxylipins were active against eukaryotic microbes. C6 and C9 aldehydes, especially (2E)-hexenal and (3Z)-hexenal, have bactericidal activity against both Gram-positive and Gram-negative bacteria [44]. Croft et al. [45] showed the lethal effect of (2E)-hexenal on the phytopathogenic bacterium Pseudomonas syringae pv phaseolicola. Other molecules such as epoxy, hydroxy, divinyl ethers, and alcohols have antimicrobial properties [8,46]. The volatile aldehydes produced by HPL also exhibit antifungal activity. Matsui et al. [47] and Kishimoto et al. [46] have demonstrated the role of these molecules in defense against Botrytis cinerea and Fusarium oxysporum. After mechanical wounding or attack by herbivores the injured leaves activate the oxylipin pathway [48]. Traumatin and traumatic acid also act as wound hormones to stimulate cell multiplication around the wound, resulting in the formation of protective calluses that impede the entry of pathogens [49]. In response to herbivories, plants produce direct defense molecules, which act as toxins and repellents, as well as indirect defense molecules [50]. The jasmonates are indispensable metabolites in mediating the wound-induced expression of direct defense molecules such as proteinase inhibitors that inactivate the proteolytic enzymes of the digestive tract of herbivores [10,51,52]. On the other hand, C6 aldehydes and alcohols and hexenyl acetate, a C6 aldehyde-derived compound, are wound-inducible volatile signals that mediate indirect defense responses by directing tritrophic (plant-herbivore-natural enemy) interactions [3,50]. In fact, attack by herbivores induces plants to emit sets of volatile compounds that differ depending on the herbivore species [53,54]. These volatile compounds attract natural enemies of the herbivore, such as carnivorous insects or parasitic wasps [36,55]. Phytooxylipins can also act as signal molecules to induce genes associated with the defense response, and thus provide a feedback loop for amplification of the signal [5]. Thus, JA and MeJA can induce upregulation of the LOX pathway [56]. C6 and C9 volatiles and MeJA serve as signals of stress within a plant as well as between neighboring plants within a plant community [57]. Under stress-related conditions such as herbivories, pathogen attack, and abiotic stimuli, these volatiles can be formed very rapidly at the wound site and travel through the air thereby potentially serving as signal molecules to distal sites in the plant, or acting as allelopathic molecules activating the expression of genes related to direct and indirect defenses of nearby plants [5,58,59,60,61,62]. Treatment of Arabidopsis tissue with LOX-derived C6-volatiles induce defense genes, suggesting that these compounds might be a signal for alerting other plants in the surroundings [5,63]. The profiles of oxylipins vary depending on the stimulus perceived by the plant [6]. These molecules will be thereby adapted to fight directly or indirectly against the source of the stimulus.
Furthermore, phytooxylipins produced by the LOX pathway especially JA and meJA from the AOS branch, are phytohormones involved in physiological processes such as flower development, seedling growth, pollen formation tuberization, and fruit maturation [3,9,64]. Traumatin and traumatic acid also act as growth promoters.
The C6 and C9 aldehydes, alcohols and their esters are known as ‘green-leaf volatiles’, because they embody the typical odor of damaged leave and have a characteristic green and fruity smell. These volatile compounds are important constituents of the flavor and fresh green aroma in fruits and vegetables and impart a characteristic odor to each plant.
3. Biosynthesis of GLVs: The HPL Branch of the LOX Pathway
3.1. Lipase Activity
The degradation of membrane lipids occurs under normal conditions during physiological and developmental processes in plants and may be initiated by biotic and abiotic stresses [52,65,66]. Polyunsaturated fatty acids converted into oxylipins by the LOX pathway are generally derived from the hydrolysis of membrane phospholipids by phospholipases or galactolipids by galactolipases [3,17,67] Phospholipids and galactolipids, especially monogalactosyldiacylglycerols (MGDG) and digalactosyldiacylglycerols (DGDG) present in chloroplast membranes, constitute the major part of the plant cell membrane. Phospholipase A (PLA) activity was found to be induced by wounding [68] and phospholipase D was found to play regulatory roles in diverse plant processes and stress responses [69]. Some studies have elucidated the role of PLA enzymes in the release of free PUFAs, which are the substrates for the oxylipin pathway. For example, defective anther dehiscence 1 (DAD1) and dongle (DGL) possess PLA1 activity and release ALA for the biosynthesis of JA in Arabidopsis [17,70]. Yang et al. [71] reported that a family of patatin-related phospholipases A (pPLA) is involved in the release of free fatty acids and monoacylglycerol through the hydrolysis of membrane glycerolipids. Glycerolipase A1 (GLA1) in Nicotiana attenuata is thought to be involved in JA biosynthesis during herbivorous and mechanical wounding in leaves, but not during Phytophthora parasitica infection [72]. Accordingly, it is assumed that there must be stimuli-specific lipases that are responsible for JA formation [64].
Although the involvement of lipolytic enzymes in the oxylipin synthesis pathway is not questioned, oxylipins linked to complex lipids have, however, also been discovered; thus, a novel oxylipin pathway which does not require the action of a lipase is envisaged. MGDG or DGDG containing 12-OPDA molecules (called arabidopsides) have been identified in Arabidopsis thaliana [73]. Galactolipids containing divinyl ether residues (linolipins) or precursors of traumatic acid have also been detected [74,75]. These particular oxylipins would result from the direct action of enzymes of the LOX pathway (LOX, AOS, or HPL) on esterified PUFAs without prior release by a lipolytic enzyme [22,74,76]. The phospholipases or the galactolipases could act in a later phase, to release the oxylipins linked to the complex lipids [75].
3.2. LOX Activity
LOXs (linoleate oxygen oxidoreductase; EC 1.13.11.12) are nonheme iron-containing dioxygenases widely distributed in plants and animals [16,77]. In plants, they have been identified in different organs such as: Leaves, fruits, roots, and seeds [78,79]. LOXs can be localized in chloroplasts, either in the stroma or associated within the envelope, as well as in lipid bodies, in vacuole, or in cytosol [6,79,80].
LOXs catalyze the regio- and stereo-specific hydroperoxidation of PUFAs with a (1Z, 4Z)-pentadiene system to produce corresponding fatty acid hydroperoxides [2,81,82]. Generally, these enzymes act on free PUFAs [82], but studies have shown that they are also able to oxygenate esterified PUFAs within complex lipids like galactolipids, phospholipids, triacylglycerols [16,22,83,84], or sterol esters [85]. In plants, LA and ALA are the most common substrates for LOX [77].
Plant LOXs are usually classified into 9-LOX and 13-LOX, depending on the regio-specific oxygenation, which can occur at carbon-9 or at carbon-13 of the hydrocarbon backbone of linoleic acid, to generate the corresponding 9- or 13-hydroperoxides [86] (Figure 3). A few non-traditional/conventional plant LOXs with dual positional specificity produce 9-hydroperoxides and 13-hydroperoxides (both in melange) with a ratio ranging from 10:1 to 1:10, and were therefore named respectively 9/13-LOXs or 13/9-LOXs [35,87,88,89]. LA or ALA are oxygenated to produce linoleic acid hydroperoxides, such as 13-hydroperoxy-(9Z,11E)-octadecadienoic acid (13-HPOD) and 9-hydroperoxy-(10E,12Z)-octadecadienoic acid (9-HPOD), or linolenic acid hydroperoxides such as 13-hydroperoxy-(9Z,11E,15Z)-octadecatrienoic acid (13-HPOT) and 9-hydroperoxy-(10E,12Z,15Z)-octadecatrienoic acid (9-HPOT) [79].
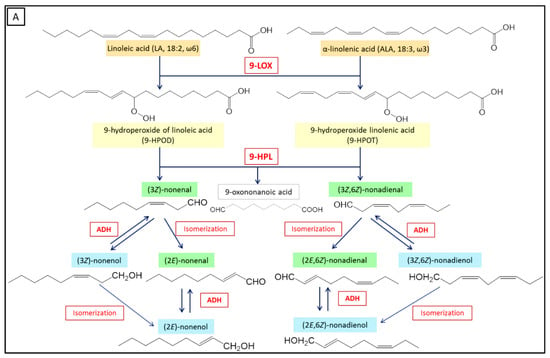
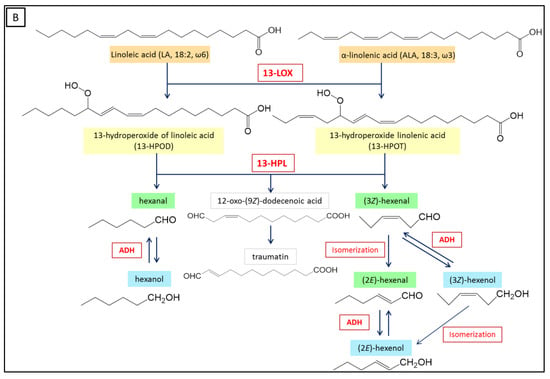
Figure 3.
Biosynthesis of green leaf volatile compounds following the LOX pathway. (A) 9-LOXs produce 9-hydroperoxy-(10E,12Z)-octadecadienoic acid (9-HPOD) and 9-hydroperoxy-(10E,12Z,15Z)-octadecatrienoic acid (9-HPOT). 9-HPLs act on 9-hydroperoxides to form C9 aldehydes and a 9-oxoacid. (B) 13-LOXs produce 13-hydroperoxy-(9Z,11E)-octadecadienoic acid (13-HPOD) and 13-HPOT. 13-HPLs act on the 13-hydroperoxides to form C6 aldehydes and a 12-oxoacid. Finally, isomerization, dehydrogenation, and esterification reactions of aldehydes can occur to produce aldehydes with different isomeric forms, alcohols, and esters. Isomerization can occur spontaneously or through the action of an isomerase. The enzymes involved appear in red, the fatty acids are colored in orange, the hydroperoxides are colored in yellow, and the oxoacids are colored gray. The aldehydes and alcohols produced are colored respectively in green and in blue. ADH: Alcohol dehydrogenase.
In most plants, several isoforms of LOX have been detected (for example six isoforms, LOX-1 to LOX-6, in Arabidopsis) [90]. LOX isoforms are distinguished by their optimal reaction pH or their specificity of products and substrates [91].
From a structural point of view, plant LOXs are monomers of approximately 100 kDa, folded into two domains, (i) an N-terminal domain composed of four β-sheets organized into a β-barrel potentially involved in membrane binding, and (ii) a larger C-terminal domain including the catalytic site and composed predominantly of α-helices. The iron atom is located in the C-terminal domain and coordinated by five highly conserved amino acid residues (three histidine, an isoleucine, and an asparagine) and a water molecule in the absence of substrate [92,93]. LOX in its native form contains Fe2+(OH2) and must first be activated by oxidation by its own hydroperoxide product to the LOX Fe3+(OH−).
The reaction mechanism of plant LOXs begins with the stereospecific abstraction of a hydrogen atom carried by the C11 of LA or ALA [7,86] (Figure 4). This step correspond to a proton-coupled electron transfer: The proton is transferred from the substrate to the oxygen of the OH− while the electron is transferred from the C9–C13 pentadiene system to Fe3+ in a concerted proton tunneling-electron tunneling process [94]. Concomitantly, the Fe3+ is reduced to Fe2+ and a water molecule is produced. The intermediate fatty acid radical formed by the abstraction of hydrogen on the central carbon is a pentadienyl radical whose electrons are delocalized throughout the pentadiene system. Then, a molecule of dioxygen binds to the pentadiene radical to form a hydroperoxyl radical. The oxygen can be inserted only at position {+2} or {−2} [7,82]. Consequently, when hydrogen is abstracted at C-11, oxygen insertion occurs at C-13 or C-9 of the C18 PUFA, to form a peroxyl radical [6]. The two double bonds are conjugated, acquiring the configuration (Z, E) [18]. The peroxyl radical is then reduced to the peroxide anion by an electron from Fe2+ which resumes its active and oxidized Fe3+ form [82]. The anion is finally protonated and a 9- or 13-hydroperoxide of LA or ALA is released from the active site of the enzyme [77,81].
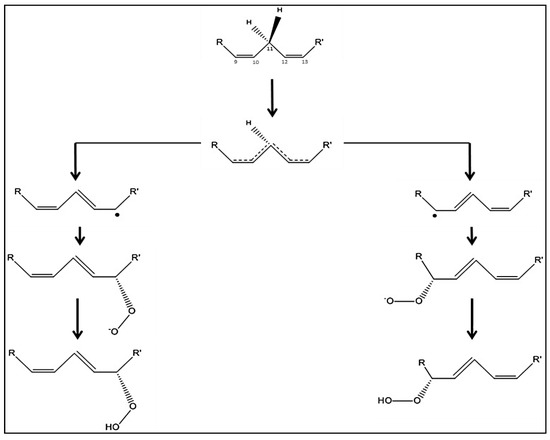
Figure 4.
Reaction mechanism of vegetable lipoxygenases. R and R‘ respectively represent the carboxylate and methyl ends of the fatty acid. The abstraction of hydrogen on the C11 of the (1Z, 4Z)-pentadiene system of linoleic or α-linolenic acid, induces an electronic delocalization on the five carbons of the pentadiene system. The antarafacial insertion of molecular dioxygen at the position {n+2} (left part) or {n−2} (right part) leads to the formation of a peroxide anion, which is finally protonated to give a 9- or 13-hydroperoxide.
Two models were proposed to explain the reaction mechanism of positional specificity of LOXs: A space-related hypothesis assumes that the fatty acid substrate would penetrate the active site generally with its methyl end first. Then, the depth of the substrate-binding pocket (related with the steric hindrance of amino acids at the bottom of the active site) limits its penetration and thus determines the relative alignment of the pentadiene system with the catalytic iron atom. So, the positional specificity of molecular oxygen insertion depends on the position of the substrate in the active site [95,96,97]. According to a second hypothesis, the regiospecificity of the LOXs would be dependent on how the substrate enters the active site, i.e., by its carboxylate for 9-LOXs or methyl end for 13-LOXs [98,99]. Consequently, a radical rearrangement at either {n+2} or {n−2}, respectively, may be facilitated in both cases by the same mechanism within the active site. At least for some plant LOXs, a combined version of both models could occur because the inverse orientation of the substrate is determined by the space available in the substrate-binding pocket [6,99].
The carbon carrying the hydroperoxide function is an asymmetric carbon. There are therefore two stereoisomers, (S) or (R), of the PUFA hydroperoxide formed. In plants, LOXs synthesize high levels of (S) diastereoisomer (95%), by contrast with spontaneous auto-oxidation, which is not stereospecific and produces racemic (R, S) mixture of isomers [100,101,102]. However, LOXs responsible for the formation of the (R) epimer have been described [16,87,89].
3.3. HPL Activity
Hydroperoxide lyase is a member of the CYP74 subfamily of cytochrome P450. HPL has been found in many plant species, and in various organs, both in photosynthetic and non-photosynthetic tissues, such as tomato [103] and mint leaves [104], sunflower hypocotyls [105], olive [106,107,108], green bell pepper fruit [109,110] and cucumber [111], soybeans [112], and rice [113].
HPL substrates must meet several structural requirements: (i) they must be a long fatty acid chain (more than 12 carbons) with a terminal carboxyl group, (ii) they must have a hydroperoxyl group on C9 or C13, and (iii) they must have a conjugated diene system (Z, E) with the double bond adjacent to the carbon carrying the hydroperoxide function [1,103,114,115]. Studies have shown that only the (S) isomer of mixtures of 13(R)- and 13(S)-hydroperoxides is converted by HPL [112,116]. The number and position of unsaturations have an influence. HPL activity increases with the number of carbons for fatty acids up to 22 carbons, and decreases thereafter [114]. The hydroperoxides of γ-linolenic acid (6Z,9Z,12Z)-octadecatrienoic acid) are only slightly converted by HPL and the 15-hydroperoxides of arachidonic acid (15-hydroperoxy-(5Z,8Z,11Z,13E)-icosatetraenoic acid, 15-HPETE) is not a substrate of the enzyme [1,114,117]. The presence of the carboxyl group on the substrate influences HPL activity: Its substitution by a methyl ester or an alcohol halves the activity of HPL [1,117]. Finally, the 9- and 13-hydroperoxides obtained from LA (9-HPOD and 13-HPOD) or from ALA (9-HPOT or 13-HPOT) are the main natural substrates of HPLs (Figure 3). The 9-hydroperoxides are converted into a 9-oxononanoic acid and a C9 aldehyde, which is (3Z)-nonenal from 9-HPOD and (3Z,6Z)-nonadienal from 9-HPOT [7,118,119,120,121]. The 13-hydroperoxides are converted into 12-oxo-(9Z)-dodecenoic acid, a precursor of traumatin, and a C6 aldehyde, which is hexanal from 13-HPOD or (3Z)-hexenal from 13-HPOT [7,106,119,122,123,124]. HPL would also be able to convert esterified fatty acid hydroperoxides into MGDGs [74].
Depending on the specificity of the substrate, plant HPLs have been classified into three types: 9-HPL, 13-HPL, and 9/13 HPL. The majority of plant HPLs are specific for 13-hydroperoxide substrates; 13-HPLs have been identified, especially, in tomatoes [125,126], sunflowers [105], potatoes [127], sugar beet [128], watermelon [129], and olive [106,107]. On the contrary, 9-HPLs have been identified from a few plant species, such as pear [118] and almond [121]. 9/13-HPLs acting on both 9- and 13-hydroperoxides, have been found especially in rice [113], melon [120], cucumber [111], medicago spp. [130], and grape berries [131]. 13-HPLs are classified as CYP74B, while 9-HPLs and 9/13 HPLs are classified as CYP74C. The presence of 9- or 13-HPL activity can vary according to the organs of the same plant. For example, in the cucumber, cotyledons show a predominant 13-HPL activity, whereas in the roots, 9-HPL activity predominates [132].
Most HPLs are membrane-bound [4,7,15,115,133]. 13-HPLs such as those present in olive [106], tomato [134], and Arabidopsis thaliana [135] are integrated into chloroplast envelope membranes. A chloroplast transit peptide located at the N-terminal end of the enzyme have been identified in 13-HPLs from olive [107,108] and Arabidopsis thaliana [135], whereas this signal peptide is absent in tomato 13-HPL [134,136]. Maucher et al. [137] and Froehlich et al. [136] hypothesized that the presence or absence of the chloroplast transit peptide would involve different targeting pathways that route the enzymes to different membranes of the chloroplast envelope. 9- and 9/13-HPLs are found in the membranes of the Golgi apparatus and the endoplasmic reticulum [78] or in lipid bodies [121,130].
HPLs are trimers [103,110,138] or tetramers [105,139] of about 170–250 kDa [109,122,140,141]. Most of HPL genes code for a subunit of 50–60 kDa [139,141,142,143,144]. Like all CYP74, HPL is a heme protein with 2.2 mol of heme b per mol enzyme [105,138,145]. Circular dichroism analysis demonstrated that the protein secondary structure is composed of approximately 13% α-helix, 32% β-sheet, 21% turns, and 31% unordered coils [146,147].
Until now, there is no crystal structure of HPL, and there are only X-ray crystal structures of two CYP74s: Arabidopsis thaliana CYP741 [148] and Parthenium argentatum (guayule) CYP742 [149], each an AOS classified as CYP74A. Investigations of the structure-function relationships of other CYP74 enzymes, particularly HPL, are generally based on the analysis of these X-ray crystal structures and/or in silico studies using molecular modeling tools. In fact, the amino acid sequences of HPL, AOS, and DES have an identity level of between 38% and 57%, depending on the enzymes [135,150]. The catalytic activity of CYP74 enzymes can be modified by single point mutations. The S297A mutant of tomato AOS [151] and the mutant F137L of Arabidopsis thaliana AOS [148] showed HPL activity. In addition to the study of crystalline AOS structures, Lee et al. [148] and Li et al. [149] analyzed protein sequence alignments of different CYP74s and performed alignments of crystalline structures of AOS with modeled structures of other CYP74s Moreover, a tridimensional model of truncated alfalfa HPL was constructed by Hughes et al. [152], by homology with cytochrome P450, a rabbit monooxygenase (CYP25C). These studies helped to advance the understanding of the structure and reaction mechanism of CYP74s and made possible to bring out similitudes but also some functional and structural differences between CYP74s and other P450s. The six substrate recognition sites (SRS) described in P450s are found in CYP74s, both in AOS and in HPL [148,149,153]. CYP74s have a major α-domain which is predominantly alpha-helical and contains a conserved structural core around heme, and a small β-domain which is predominantly composed of beta-sheets with two α-helices A and A’ in the N-terminus, and a 310 helix B [148,149]. The α-helices and β-sheets were labelled by analogy with those of the P450s. The heme prosthetic group is located mainly between helices I and L. Cysteine acts as the fifth ligand for the heme iron. Helix I contains residues essential for catalysis [148,153,154]. In the central region of this helix, an asparagine conserved in all CYP74s pointing to the heme may interact with the 13-hydroperoxyl group of the substrate, suggesting a critical role in catalysis [148,153].
In the proposed reaction mechanism of HPL [27,28,29], the substrate displaces the water molecule that served as the sixth ligand of the heme iron (Figure 5 step 1 and 2). The iron (III) is then coordinated with the terminal oxygen atom of the hydroperoxide function of the substrate. Hydrogen bonds are established between the amide group of asparagine and the two oxygen atoms of the hydroperoxide function [148]. This results in the homolytic cleavage of the O-O bond of the hydroperoxide, giving rise to an alkoxy radical (RO•) and a protonated ferryl species Fe(IV)-OH [124] (Figure 5 step 3). The hydrogen bonds between asparagine and oxygen are maintained. A rearrangement of the double bond between C11 and C12 of the substrate creates a cyclization around the oxygen atom of the radical, resulting in the abstraction of hydrogen on the C11. A 12,13-epoxide with a carbon-centered radical (C•) at C11 is thus generated [148] (Figure 5 step 4). Since this epoxycarbinyl radical is not stabilized, the bond between C12 and C13 is split off. A double bond is established between C11 and C12, and the radical formed at C13 undergoes oxygenation by iron-catalyzed oxygen transfer [28] (Figure 5 step 5). An unstable hemiacetal is thus generated (Figure 5 step 6), and then cleaves spontaneously to form a C6 or a C9 aldehyde and an oxoacid [27,29] (Figure 5 step 7).
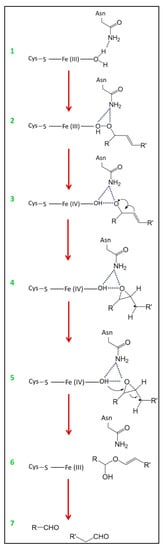
Figure 5.
Schematic diagram of the proposed reaction mechanism of HPL [148]. In the absence of substrate, the iron atom of the heme is coordinated with a cysteine residue and a molecule of water which establishes hydrogen bonds with an asparagine residue (1). The substrate picks up the molecule of water (2). After cleavage of the O-O bond of the hydroperoxide function of the substrate (3), an epoxycarbinyl radical is generated (4). The formation of a double bond between the carbons 11 and 12 and the oxygenation of the carbon 13 (5) leads to the formation of a hemiacetal (6) which cleaves spontaneously into aldehyde and oxoacid (7). R and R’ represent the carbon chains, respectively, towards the methyl and carboxyl ends of the substrate.
Based on alignments of AOS and HPL sequences, Lee et al. [148] found the substitution of a phenylalanine in AOS (F137 Arabidopsis thaliana AOS) by leucine in HPL. The involvement of the residue at this position in the catalytic mechanism and the reactive specificity of AOS and HPL has been discussed [148]. Indeed, the first steps (Figure 5 steps 1 to 4) of the catalytic mechanisms of the two enzymes are identical and both lead to the formation of an epoxide with a radical at C11 [148,154]. However, they differ subsequently, mainly depending on the presence of the phenylalanine or leucine residue. Lee et al. [148] suggest that in AOS, the presence of the phenylalanine residue having an aromatic ring stabilizes the radical at C11 and makes possible its oxidation. The carbocation formed is stabilized by cation-π interactions with the phenylalanine residue, which favors the formation of an allene oxide. On the other hand, in HPL, the presence of a leucine residue does not lead to stabilization of the radical, leading to the cleavage of the bond between the carbons C12-C13 of the epoxy group, and thus orienting the mechanism towards the formation of an unstable hemiacetal. The substitution of phenylalanine 137 residue of Arabidopsis thaliana AOS by a leucine residue (mutant F137L) exhibit HPL activity [148]. Similarly, the substitution of phenylalanine 92 by leucine in rice AOS (mutant F92L), allowed the production of 12-oxoacid by the enzyme [148,155].
The action of HPL on HPOs is carried out at an optimum pH of between 5.5 and 8 [20,106,109,118,127,140,156], and at an optimal temperature ranging from 20 to 45 °C [107,127,133,144]. When the amount of HPOs is in excess of that of HPL, the reaction velocity gradually deceased and finally stopped before all available substrate was used [125,126]. Only a secondary addition of fresh enzyme induced renewed reaction [125]. This is typical of mechanism-based inhibition or suicide inactivation which is a phenomenon commonly found in cytochromes P450. Studies by Matsui et al. [157], Matsui et al. [125], and Suurmeijer et al. [126] concluded that hydroperoxides specifically recognized by the active site of HPL are transformed into radical species responsible for the oxidation of the sulfhydryl group of the cysteine involved in binding with the catalytic iron atom. Dithiothreitol (DTT) and organic antioxidants such as α-tocopherol, butylhydroxyanisole, and butylhydroxytoluene protect HPL against this inactivation [125,157].
HPL is not inhibited by the direct products of its action (such as hexanal or (3Z)-hexenal) but by the α,β-unsaturated aldehydes obtained after isomerization (in particular (2E)-hexenal), which can bind to the sulfhydryl group [126,144].
HPL can be inhibited by metal chelators such as salicylic acid and deferoxamine mesylate (Desferal), known to inhibit heme enzymes [105,125,126]. Nordihydroguaiaretic acid (NDGA) inactivates HPL by reducing the active form Fe (III) of catalytic iron to its inactive form Fe (II) [144,158].
3.4. Isomerization, Dehydrogenation and Esterification Reactions of HPL Products
The C6 and C9 aldehydes produced by the HPL branch of the LOX pathway, as well as the corresponding alcohols and esters are commonly called green leaf volatiles. When produced by HPL, the short-chain aldehydes are only into Z conformation, but then they can be isomerized to the isomeric form E. Isomerization can occur spontaneously depending on the physical factors of the medium (pH, temperature), or be promoted by an isomerase activity [7,81,159] (Figure 3). The aldehydes can also be reduced to alcohols by an NAD-dependent alcohol dehydrogenase (ADH, EC 1.1.1.1) [75,133,160,161].
The action of a 13-HPL on a 13-HPOD leads to the formation of hexanal, which can be converted into hexanol by ADH [21]. The action of a 13-HPL on a 13-HPOT results in the release of (3Z)-hexenal, which can be reduced to (3Z)-hexenol by ADH, or isomerized in (2E)-hexenal, and then converted by ADH to (2E)-hexenol [1,10].
From 9-HPOD, 9-HPL produces (3Z)-nonenal that can be reduced to (3Z)-nonenol by ADH, or isomerized to (2E)-nonenal, then converted by ADH into (2E)-nonenol. If the substrate is 9-HPOT, (3Z,6Z)-nonadienal is produced and then reduced to (3Z,6Z)-nonadienol by the ADH, or isomerized at (3E,6Z)-nonadienal, then converted by ADH to (3E,6Z)-nonadienol [21,81,162].
The short-chain carbon alcohols thus produced can be esterified by an alcohol acetyl transferase (AAT), using acetyl coenzyme A [61,75]. AAT catalyzes particularly the synthesis of (3Z)-hexenyl acetate from (3Z)-hexenol [61].
4. Industrial Purpose and Use of GLVs
4.1. Flavoring Interest of GLVs
In addition to their biological activities on plants, herbivores, and microorganisms, GLVs including C6 or C9 aldehydes, alcohols, and esters, have interesting flavoring properties, which make them widely used in industry (Table 1).

Table 1.
Odor qualities attributed to green leaf volatiles (GLVs) formed by the lipoxygenase pathway.
These volatile compounds are responsible for the fresh smell of green leaves and cut grass referred as “green note” in the field of aromas and fragrances [2,162]. They are important constituents of the flavor and fresh green aroma in fruits and vegetables and they give a distinctive odor to every plant and consequently to processed agri-food products [2,133,163].
The key aromatic compounds that contribute to the green and grassy odor note of guava fruit are (3Z)-hexenal and hexanal [164]. The volatile alcohols, (3Z)-hexenol and (2E)-hexenol, and the aldehydes, hexanal and (2E)-hexenal, with their green, grassy, sweet, fruity odor, are the major contributors of the sensory characteristics in different tea varieties [165,166]. C6 compounds, particularly hexanal, (2E)-hexenal and hexanol, are the most abundant volatiles in either red or white grapes and give a grassy smell to grapes, which is crucial to grape berry flavor [131]. When grape berries get injured or crushed, GLVs are quickly generated and released. Thus, such steps as destemming and crushing in the process of winemaking can directly affect the generation of C6 compounds and consequently the flavor of the produced wine [167]. C6 aldehydes (hexanal, (3Z)-hexenal and (2E)-hexenal) and alcohols (hexanol, (3Z)-hexenol and (2E)-hexenol), and their acetyl esters (hexyl acetate and (3Z)-hexenyl acetate), are the most important quantitative (80% of total volatile compounds) and qualitative compounds in virgin olive oil aroma, with (2E)-hexenal being the most prominent component [168,169,170]. The biosynthesis of these C6 compounds depend upon the availability of PUFAs, especially ALA, and the enzymatic activity of the LOX/HPL system when tissues are disrupted during physical procedures (milling, malaxation) of the oil extraction process [106,107,160,171,172]. Thus, the GLVs synthesized during the olive fruit crushing are incorporated into the oil conferring their fresh green odor.
4.2. Industrial Use of GLVs
Because of the blend of green, fresh and fruity aromas, GLVs are widely used in perfumes and flagrances, food products like chewing gums, soft drinks, and also in prepared meals, to impart a green character as well as a feeling of freshness. The natural ‘fresh green’ aroma of fruits and vegetables, which is lost during industrial processing or sterilization processes, can be reconstituted through the addition of HPL-generated volatiles. GLVs are also used to improve the shelf-life and safety of processed foods due to their antifungal and antibacterial activities against both Gram-negative and Gram-positive bacteria [173]. Among GLVs, (2E)-hexenal is highly bactericidal, essentially because of its α,β-unsaturated carbonyl moiety that has high reactivity [174,175].
Each of the compounds has a characteristic fragrance and a specific use (Table 1). The separate use of one or more of these flavors is highly sought after.
GLVs share a major part of the global flavors and fragrances market (F&F market), which is valued at approximately US $26.3 billion in the worldwide market in 2017*. This amount represents a growth of 1.1% compared to the previous year and it is expected that F&F market will continue to increase steadily year in year out. The production of GLVs is therefore an important issue for the industry.
4.3. Production of GLVs: Synthetic versus Natural Flavors
The GLVs market mainly comprises the synthesis of artificial flavors, of which mainly C6 aldehydes and alcohols [176]. Indeed, chemical synthesis is the easiest way to produce large amounts of stable C6 or C9 aldehydes and alcohols. The production method is inexpensive and provides high yields for a final product 95% pure. The synthesis begins with the production of C6 alcohols from a combination of ethylene molecules and directed oxidation. Ethylene is produced from naphtha from fractional distillation of petroleum. Additional directed oxidation is then performed to obtain the aldehydes. After distillation, pure mixtures with more than 95% of aldehydes or alcohols can be marketed at about 25–50$ per liter, depending on the molecule synthesized.
However, in recent years, consumers are increasingly demanding natural products including flavor, which they consider more healthy and safer than synthetic ones [19,176]. Furthermore, this approach also allows substitution of petrochemical resources by renewable agrochemical resources since natural processes are based on natural raw materials or renewable resources and are therefore environmentally friendly and sustainable.
The EU regulation (EC) No 1334/2008 on flavorings and food ingredients with flavoring properties defines in the articles 3.2(c) “natural flavoring substance” as “a flavoring substance obtained by appropriate physical, enzymatic or microbiological processes from material of vegetable, animal or microbiological origin either in the raw state or after processing for human consumption by one or more of the traditional food preparation processes (including drying, roasting and fermentation)”. Biotechnologically produced flavors are also covered by the term “natural”. The regulation also states that “natural flavoring substances correspond to substances that are naturally present and have been identified in nature”. All flavorings considered natural in the EU should also be considered natural in the USA; however, the reverse is not necessarily true [177].
Actually, natural GLVs are the most valuable flavor class commonly sold in natural food industry but its large-scale synthesis remains a great challenge for producers. Natural GLVs are high value-added compounds which can be marketed at about 750–3000$ per liter [176,178], depending on the molecule, but their production is more complex and expensive than that of artificial flavors.
To produce a natural aroma, the initial substrate must be of plant or animal origin. This means that extracts from natural sources as well compounds produced by biotechnological processes may be considered as natural aromas.
Due to the growing demand and the low abundance of compounds in plants, the extraction of GLVs directly from plants is unprofitable and very expensive.
Therefore, in order to meet market requirements, efficient biotechnological processes of GLVs synthesis are developed for large scale production. These processes reproduce in vitro the reactions of the LOX pathway for GLVs biosynthesis (Figure 6). Vegetable oils rich in C18 PUFAs are the best initial substrates for GLVs production because they are inexpensive and available in large quantities. The enzymatic reactions of the LOX pathway are carried out successively on this substrate in order to obtain the GLVs: The triacylglycerols from the vegetable oil are first hydrolyzed by the action of a lipase, then the released PUFAs (C18:2 and C18:3) are converted by the actions of a LOX and then a HPL into GLVs. Plant material in the form of flours, or crushed leaves or pulps are often used as a source of LOX or HPL. The process ends with the extraction of the volatile molecules to obtain a sufficiently pure product.
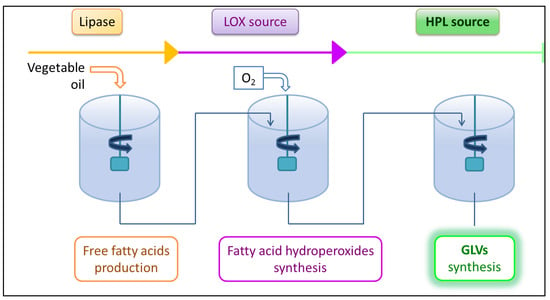
Figure 6.
Biotechnological process of GLVs production in bioreactor. Biotechnological processes are based on the reactions of the LOX pathway. First, a vegetable oil in emulsion is hydrolyzed by the action of a lipase to release the fatty acids. Then, a lipoxygenase catalyzes the dioxygenation of the polyunsaturated fatty acids into hydroperoxides. HPL acts on these hydroperoxides to produce C6 or C9 aldehydes that are GLVs.
Several patents have described the production of natural GLVs from vegetable oils using plant material containing the enzymatic activities of the lipoxygenase pathway. Formerly, the described processes used a single plant source to perform all processing steps. Kanisawa and Itoh [179] patented a method for preparing green flavors using crushed soybean seeds with or without additional lipase activity. Representative examples of substrate have been used such as soybean, flax, olive, rapeseed, maize, wheat, sesame, nut, grape seed, palm, and coconut oils. The compounds produced are considered as natural flavoring substances. In another patent, Goers et al. [180] have described a process for producing C6 aldehydes and alcohols using strawberry leaves as a source of biocatalysts. In a bioreactor, linoleic acids or hydrolyzed linseed oil are converted to GLVs by the actions of LOX, HPL, and ADH present in the leaves. However, yields remain low because, in many cases, not all the enzymes of interest have sufficient activities in the same organism at the same stage of development. The activity of HPL is, for example, very low during the development of soybeans [181] and decreases rapidly during storage, unlike LOX [182]. One of the advantages of biosynthesis, compared to the extraction of compounds, is precisely the possibility of using enzymatic activities from different plant sources to perform successive reactions. Therefore, in many biotechnological processes, soybean is preferentially used as a source of LOX and must be coupled to another source of HPL. Several biotechnological processes will be described in more details in part V.2.
Nevertheless, the use of plant extracts has disadvantages. Large quantities of plant material whose supply depends on the harvest period and climatic hazards, are necessary and a lot of waste is generated. For example, a biotechnological process requiring the use of fresh guava fruit, has to be implemented in a country where fresh guava fruits are available cheaply and freely, but even when such a site is found, availability is limited to the growing season of the fruit [183]. Another disadvantage is that the desired enzyme activities are rather dilute in the sources employed. Furthermore, plant extracts contain many enzymes that use the same substrates as LOX and HPL, leading to undesired products and loss of substrate. Besides, LOX and HPL of different regiospecificities are present, generating a mixture of C6 and C9 compounds, while industrials often seek a separate use of these flavors.
These drawbacks can be circumvented by using purified and regiospecific enzymes such as recombinant enzymes, allowing efficient and highly selective (chemo, regio, and stereo) production of the desired volatile compound(s) through directed synthesis. It also helps to overcome the dependence on plants, which offers technical advantages for plant material supply and waste treatment. The use of recombinant enzymes also allows the qualification of the molecules being produced as natural flavoring substances and the possibility of making such a claim on the product label and the list of ingredients.
5. Potential of the Use of the LOX Pathway for Biotechnological Production of Natural GLVs
Biotechnological production processes of natural GLVs involves various steps with the sequential or simultaneous (coupling) use of the enzymatic biocatalysts, in order to convert the FAs of a vegetable oil into HPOs and then into GLVs. The various steps are described in the following paragraphs and some data of the biocatalytic reactions are detailed in Table 2.
Table 2.
Comparison of the methods used and the yields obtained for reactions of biotechnological processes.
5.1. Release of Fatty Acids from Triacylgycerols
The first step in GLVs synthesis is the production of free PUFAs from complex lipids. So, it is necessary to benefit from a lipid source rich in interesting FAs. Vegetable oils such as sunflower, linseed, colza, olive, or soybean oils rich in C18 PUFAs can be used as a raw material substrate for the biotechnological processes. Fu et al. [184] showed that the more PUFAs in the oil, the faster is the hydrolysis of triacylglycerols. The choice of the oil determines the class of HPOs produced and thus the class of GLVs synthetized. Generally, biotechnological processes use lipase to hydrolyze olive oil or sunflower oil rich in LA for subsequent HPOD production, and linseed oil rich in ALA for subsequent HPOT synthesis.
Release of FAs can be performed by chemical hydrolysis of the oils. The use of water or ethanol in this hydrolysis is compatible with agri-food regulations (Directive 2009/32/EC of the European parliament and of the council). But the drastic conditions of hydrolysis require an achievement of this step separately from the later steps of the process, because they would cause denaturation and loss of activity of all the enzymes subsequently involved in the synthesis.
Enzymatic hydrolysis is an opportune choice of reaction because this process is one of those accepted for the production of natural flavoring substances. For industrial purposes, microbial lipases are often used to produce large amounts of free FAs [185]. Commercially available lipases usually obtained from microorganisms can be used as potential biocatalysts. In order to reduce the cost of the enzymatic process, several studies have successfully evaluated the effectiveness of immobilized lipases, which allows the reuse of the enzyme to be considered [186,187].
The hydrolysis of triacylglycerols from the oil produces diacylglycerols, monoacylglycerols, and glycerols with release of FAs at each step. Triacylglycerols are insoluble in water but can be solubilized in organic solvents like hexane or octane [184,188]. Several organic/aqueous biphasic systems are used for lipid hydrolysis [184,189,190,191,192]. The hydrolytic reaction catalyzed by lipases takes place generally at the interface between the organic and aqueous phases (in particular oil/water interface). The free FAs produced by the hydrolysis of triacylglycerols are poorly soluble in water and this characteristic depends on the nature of the carbon chain and environmental conditions (pH and temperature).
Nevertheless, the use of certain organic solvents is problematic for food applications, unlike the production of a perfume aroma. The alternative solution is the production of a stable oil emulsion in an aqueous solution by high speed mixing and addition of emulsifying molecules [193,194].
5.2. Transformation of Polyunsatured Fatty Acids into Hydroperoxides
The production of HPOs is the second step towards the synthesis of GLVs. HPOs synthesis involves (i) PUFAs in particular LA or ALA or a mix of both, (ii) a source of LOX, to produce 9- or 13-HPOs, and (iii) oxygen as cofactor of the reaction. The actions of lipases and LOX can be carried out successively or coupled within the same bioreactor.
If the LOX step is processed separately the reaction is carried out in aqueous medium. Nevertheless in order to consider lipase/LOX coupling, the ability of LOX to catalyze the oxygenation of PUFAs in a medium containing both an aqueous phase and a non-aqueous phase has been the subject of several studies [12,195,196]. Drouet et al. [12] investigated the potential of soybean LOX for the synthesis of HPOs in a biphasic medium (octane;borate buffer, pH9.6) and showed an improvement (23–45%) of the reaction yield (compared to an aqueous system) at very high concentrations of substrate (20–40 g/L). In accordance with the regulations, for perfume aromas production, biphasic reactors (containing aqueous phase and organic solvent) can be used to combine lipase and LOX reactions. In these bi-enzymatic systems in biphasic medium, the FAs released by the lipase remain in the organic phase which serves as substrate tank for the LOX, while the lipoxygenation of FAs with (1Z,4Z)-pentadiene system takes place in the aqueous phase. HPOs are hydrophilic and therefore remain in the aqueous phase [197,198]. In the coupling system, the conversion rate is essentially controlled by the mass transfer. Gargouri and Legoy [199] used a numerical integration program to model the two-enzyme system in the heterogeneous biphasic media. The model was based on results obtained in separate studies with following three phenomena: Hydrolysis in biphasic media, mass transfer, and lipoxygenation in an aqueous media. The model simulates the evolution of the component concentrations in the system as a function of time and confirms that catalysis and transfer influenced each other reciprocally. There is a catalytic advantage of the proximity between the enzymes and their substrates. The presence of the second enzyme accelerates and increases the consumption of the first substrate (triacylglycerols), since the LOX reacts with the PUFAs produced by the first reaction [188,197,199]. This system makes it possible to couple the production of free FAs and HPOs but it requires extraction of the HPOs before subsequent reactions.
Numerous studies showed that the immobilization of LOX allows a high recovery and reuse of the enzyme, a better stability and protection of the enzyme during storage, but the enzymatic activity remains lower than that of the free enzyme [200,201,202]. The limiting factor to the use of immobilized LOX would be a problem of diffusion of the substrate in active site of the enzyme [81].
Biotechnological processes that focus on C6 aldehydes production often use soybean (Glycine max) as source of LOX for 13-HPOs production from free PUFAs. Soybean has been reported to contain in its seeds sufficiently high and stable LOX activity for an industrial application [81]. Fauconnier and Marlier [13] described the conversion of ALA contained in linseed oil to 13-HPOT using soybean LOX and obtained high transformation yields (71.5%) in a 10 L bioreactor without addition of any solvent or surfactant. In soybean seeds, different isoforms of LOXs (at least six including three main ones) have been described, on the basis of differences in pH optima, substrate specificity, and their product formation [81,203]. LOX1 differs from other isoenzymes in its optimum pH (9–9.5 instead of 6–7) and at this optimum pH LOX1 produces 13-HPOs with LA as preferential substrate [203]. Soybean seeds flour is often used as a source of LOX for the production of 13-HPOs in biotechnological processes [128,204,205]. Although the LOX1 content is higher than the levels of other LOX isoforms in soybean seeds, the reaction conditions should maximize the action of the selected LOX isoenzyme. For example, to favor the action of LOX1 and thus the formation of 13-HPOs, Márczy et al. [206] as Fauconnier and Marlier [13] chose to work at low temperature and basic pH. In this way, LA (100 mM) was converted by soybean LOX1 in presence of O2 into 13-HPOD (68.7 mM) with a yield of 72%. Specific heat treatments can also reduce the activity of other LOX isoforms to maximize the production of desired HPOs [207]. Otherwise, partially purified soybean LOXs (with different degrees of purity) are commercially available and can be successfully used for the conversion of PUFAs into HPOs.
For C9 aldehydes production, LOX extracts from potato can be used to convert PUFAs into 9-HPOs since a high 9-LOX activity has been reported in potato [208,209]. Gargouri and Legoy [210] described the use of a two-enzyme system involving a lipase from a Pseudomonas sp. and an extract of potato tubers containing LOXs to convert triacylglycerols into 9-HPOs. Otherwise, partially purified potato LOXs are commercially available.
Kerler et al. [211] patented a production process of C6 and C9 GLVs using as initial substrate particularly linseed oil and sunflower oil, and placing at pH7 for the lipoxygenation step, to promote the action of the LOX2 isoform of soybean flour to produce both 9 and 13-HPOs. These HPOs were subsequently heated (in the range 90 to 180 °C) under acidic conditions to promote Hock fragmentation (or rearrangement) of the HPOs, leading to aldehydes [211,212]. The oxidation of linseed oil produced C6 aldehydes, mainly (2E)-hexenal (20,150 ppm) and its less stable isomer, (3Z)-hexenal (10,380 ppm) and C9 aldehydes including (2E,6Z)-nonadienal (8900 ppm). When the sunflower oil was used as the substrate, (3Z)-hexenal (125 ppm) and hexanal (5250 ppm) were produced.
5.3. GLVs Synthesis
First, production of specific GLVs compounds (C6 or C9 GLVs) is directed by the selection of adequate substrates (i.e., the use of a sufficiently pure substrate either 9- or 13-HPOs) and adequate HPL activity (i.e., 9- or 13-HPL activity). HPL activity is effective in various plant sources. The plant source must be selected for industrial transformation capabilities such as optimal reaction conditions (pH and temperature), availability, specificity, and concentration. Due to its low concentration in plants, seasonal variation, suicide inactivation phenomena, low storage and process stability, difficulty to purify, and unavailability of the enzyme in commerce, HPL is the limiting component for the industrial conversion of FAs into GLVs. Several plants with HPL activity have been tested and many methods of producing GLVs mixing a lipid substrate with plant material containing the biocatalysts have been described and attempted to be optimized.
Márczy et al. [206] described a process for producing natural C6 aldehydes in two steps: First, oxygenation of LA (100 mM) contained in hydrolyzed sunflower oil has been performed by LOX1 from soybean flour to form 13-HPOD (68.7 mM) (72% yield); then, the 13-HPOD (15 mM) was cleaved by HPL isolated from spinach leaf resulting in 8.2 mM hexanal (54% yield). Nemeth et al. [207] used almost the same system: (i) LOX1 from soybean flour was chosen to oxygenate ALA (100 mM) contained in hydrolyzed linseed oil to form 13-HPOT (57 mM) (62% yield); and (ii) the 13-HPOT (20 mM) was cleaved by HPL of a homogenate from green bell pepper fruits, resulting in 5.9 mM (3Z)-hexenal and 1.6 mM (2E)-hexenal (37% yield for the hexenal isomers together). Hexenals were isolated from the reaction mixture by repeated steam distillations. During distillation, the molar ratio of the two isomers (2E)-hexenal:(3Z)-hexenal increases from 0.27 to 7.86 following the heat treatment. Rabetafika et al. [128] developed a one step process in which all reactions performed in a same bioreactor. One liter of reaction mixture contained 3 g soybean flour and 3 g linseed oil previously hydrolyzed by immobilized Thermomyces lanuginosa lipase. pH was kept at 9.0 to ensure maximal activity of soybean LOX1 isoenzyme in order to favor 13-HPOs formation. After the LOX reaction, the pH was adjusted to 6.7 and a homogenate from crushed sugar beet leaves containing HPL activity was used to perform the conversion of 13-HPOT to (3Z)-hexenal with a maximum yield of 80% reached after only 2 min. The stability of (3Z)-hexenal was improved by acidifying the reaction medium. According to Brunerie [213], which used crushed radish hay as a source of HPL to produce volatile aldehydes, excellent selectivity in producing (3Z)-hexenal over (2E)-hexenal is marked for temperatures between 0 and 20 °C. Gargouri et al. [104] described a process using a mint leaf homogenate as source of HPL activity for the synthesis of hexanal and hexenals respectively from 13-HPOD and 13-HPOT (themselves obtained through the action of commercial soybean LOX on LA and ALA respectively). They observed a very limited isomerization of (3Z)-hexenal to (2E)-hexenal, since the reaction and the volatile purge were carried out successively in the same flask without delay or any contact with the atmosphere. Schade et al. [214] reported a process for the production of hexanal using immobilized enzymes extracted from different plant sources (carnation petals and strawberry and tomato leaves). These enzymes were immobilized in an alginate matrix and used as a biocatalyst in a packed-bed bioreactor. The use of immobilized enzymes extracted from one gram of tomato leaves produced 80.2 μg of hexanal at 25 °C. They report that this production was 112 times greater than the amount of hexanal extracted from 1 g of plant tissue, which shows the interest of biocatalytic processes. Continuous product recovery was achieved using a hollow-fiber ultrafiltration unit.
Once synthesized, aldehydes can be converted into the corresponding alcohols through the action of ADH. The parameters of the synthesis define the nature of the products obtained. Indeed, depending on the reaction time between the actions of HPL and ADH, the (3Z) or (2E) isomers can be selected. Several authors [104,207] have shown that the isomerization of (3Z)-aldehydes in (2E) is not instantaneous, but requires several minutes or hours. By choosing the moment of addition of the ADH, it is possible to convert one or the other product in the form of alcohol. Indeed, by delaying the addition of ADH to the action of HPL, the (3Z)-aldehyde undergoes isomerization to the (2E) isomer before the start of the ADH reaction, resulting in a predominant production of (2E)-alcohol by reduction of the corresponding aldehyde. On the contrary, the ADH activity can be mixed simultaneously with that of HPL to primarily convert (3Z)-aldehyde to (3Z)-alcohol, avoiding isomerization of the double bond. Akacha et al. [215] simulated (using the MATLAB program) the conversion of 13-HPOT to hexenal and hexenol in a two-enzyme system involving successive action of HPL from mint and yeast ADH. The aim of the model was to predict the steady-state performance of the kinetics in terms of conversion. The simulation was based on Michaelis–Menten equations and experimentally determined parameters. This model simulates the evolution of the component concentrations in the system as a function of time. The model predicts total disappearance of initial substrate in few minutes (~20 min) leading progressively to the production of hexenal that rapidly disappears in order to form hexenol. Gargouri et al. [216] and Akacha et al. [215] showed that when coupling HPL activity to ADH activity, the second reaction catalyzed by ADH could favor the first one catalyzed by HPL, resulting in an increase of the conversion rate of the initial substrate (HPOs). Akacha and Gargouri [217] proposed an enzymatic liquid/gas reactor, where the synthesis of C6 compounds was coupled to their extraction (which acted by their removal with carrier gas N2). First, 13-HPOT is produced from chemically hydrolyzed linseed oil by the action of LOX of a homogenate from soybean seeds. Then, (3Z)- and (2E)-hexenals (up to 0.36 g·kg−1 of reaction medium) are produced from 13-HPOT in presence of HPL of homogenate from olive leaves (50% yield). Finally, the hexenals are successfully reduced into their corresponding alcohols ((3Z)-hexenol up to 3.54 g·kg−1 of olive leaves with a yield of 47.7%) by adding yeast cells Saccharomyces cerevisiae containing ADH activity to the same reactor. Baker’s yeast containing ADH is the most commonly used for C6 and C9 alcohols production from aldehydes in biotechnological processes. Muller et al. [204] patented a process using sunflower or linseed oil as initial substrate, soybean flour as LOX source, guava homogenate containing HPL, and S. cerevisiae yeast with ADH activity. Steam distillation and/or extraction with an inert organic solvent is used in recovery of the aldehydes and alcohols. Depending on the lipid source providing FAs, the reaction cascade and the experimental design chosen, various GLVs can be produced: Hexanal, hexenol, (3Z)-hexenal, (3Z)-hexenol, (2E)-hexenal, or (2E)-hexenol. The yields obtained are for example 5 g·kg−1 of reaction medium for hexanal, 4.2 g·kg−1 for (3Z)-hexenol and 1.5 g·kg−1 for (2E)-hexenal [204].
For C9 GLVs production, Hausler et al. [218] patented a process that uses Candida cylindracea lipase to hydrolyze flaxseed oil, and then shreds of violet leaves to obtain C9 aldehydes, which can be reduced to alcohol using baker’s yeast cells. This process resulted in 661 mg·kg−1 of plant material for (2E,6Z)-nonadienal and 44 mg·kg−1 of plant material for (2E,6Z)-nonadienol.
Short chain alcohols can subsequently be transformed into esters which have a reduced aromatic power, but a very high chemical stability [219]. The synthesis of esters such as (3Z)-hexenyl acetate can be performed by the AAT.
Despite efforts to develop high GLVs production systems, yields remain low for industrial purposes and in many cases stability and specificity of the biocatalysts have to be optimized to improve the productivity.
In fact, one of the main problems for HPOs transformation into GLVs using plant extracts is the abundance in plants of HPOs metabolizing enzymes other than HPL, resulting in undesirable products and loss of substrate. Among these enzymes, AOS is highly concentrated in some plants organs [136]. Brunerie and Koziet [220] have optimized a method using shredded leaves and baker’s yeast to produce C9 alcohols, by adding salicylic acid. The use of salicylic acid could inhibit the secondary metabolic pathway leading to the jasmonic acid derivatives from the HPOs produced by the LOX activity, and thus favor the pathway leading to the aldehydes and alcohols. In the presence of salicylic acid (1 mM), the concentration of (3Z)-hexenol produced is of 425 ppm, which represents a 24% increase compared to the control without salicylic acid. Another alternative to circumvent the problem is to consider the (at least partial) purification of HPL. A purification method of tomato leaves HPL developed by Fauconnier et al. [103], involving selective precipitation with different PEG6000 amounts, ultracentrifugation and chromatography on DEAE column permitted to obtain a 120 fold concentrated pure extract. Suurmeijer et al. [126] obtained better results for the purification of tomato fruit HPL, successively by filtration, concentration, ultrafiltration, and purification on a Sephadex G100 column and finally, chromatography on a DEAE column, since the enzymatic extract has been purified more than 300 fold.
However, another main problem is that the biotechnological application of HPL is limited by its low stability [133,221,222]. Suurmeijer et al. [126] show that partially purified tomato HPL is inactivated after 24 h at 4 °C. Itoh and Vick [105] reported a 50% loss of sunflower HPL activity after storage for 48 h at 4 °C. After five weeks of storage at 4 °C, only 12% of the initial activity of a HPL enzymatic extract from mint leaves is found [222]. In addition, the enzyme is sensitive to freezing [222,223]. The activity of HPL from Amaranthus tricolor leaves decreased by 31.4% after 24 h storage at −20 °C [223].
Nevertheless, addition of selected stabilizing additives, including salts, sugars/polyols, and polymers to the enzymatic extracts has been considered as an efficient mean for the stabilization of biocatalysts as well as for the enhancement of their activities. Compounds such as glycerol [126,223], sucrose [126], or dithiothreitol [224], can be used as cryoprotectants. Lyophilization of HPL enzymatic extracts of mint leaves and Penicillium camemberti in the presence of KCl preserved enzymatic activity for more than four weeks [221,222]. The protective effect of several additives was attributed to the modification of the enzyme’s microenvironment, specifically its water activity and/or to the molecular interactions between proteins and additives [225,226]. Koeduka et al. [226] showed that the addition of 2 M KCl, 1 M NaCl, or 0.5 M Na2SO4 in the reaction medium increase the specific activity of barley HPL (five-fold increase with 1 M KCl). Glycine also appears to be an effective additive since its presence at 10% (w/v) increases the catalytic efficiency of a mint leaves HPL extract by 2.5 times [222].
Furthermore, immobilization could also be considered to improve enzyme stability. The thermal, operational, and storage stabilities of 13-HPL from Amaranthus tricolor leaves were significantly improved after immobilization, as well as the tolerance against its substrate and product inhibitions [227]. 13-HPL was first partially purified from A. tricolor leaves, then the 1,6-hexamethylenediamine attached chitosan-carrageenan with biomimetic hydrophobic surface was proved to be the most suitable carrier [227]. Using the immobilized enzyme as the catalyst, the maximum hexanal concentration obtained was 3560 (±130) mg/L in a packed-bed reactor [228]. In the packed-bed reactor, continuous addition of low amounts of substrate and removal of reaction mixture containing products slowed the enzyme inactivation and prolonged the operating time.
The industrial production of GLVs by biotechnological processes using plants is subject to environmental as well as technical constraints since large amounts of raw materials are required and the batch processing in large quantities generates a large amount of residual organic matter. Thereby considerable efforts have been made to clone and produce this enzyme with enhanced stability and activity. Recombinant expression is an excellent way to increase the availability of HPL used in biotechnological processes.
HPLs of different plant species have been cloned and expressed in prokaryotic or eukaryotic organisms, with the aim of producing large amounts of enzyme. Alfalfa, potato, sugar beet, and olive 13-HPL and almond 9-HPL were cloned and expressed in Escherichia coli [107,121,127,133,205]. Noordermeer et al. [205] have described a biocatalytic process for the production of C6 aldehydes from hydrolyzed safflower and linseed oils using soybean flour containing 13-LOX and alfalfa recombinant 13-HPL. Culture in E. coli yielded 3000 and 8000 units of recombinant HPL per liter of culture. Molar conversion rate of 50% for hexanal and 26% for hexenal formation were obtained with recombinant HPL and in contrast to plant extracts no side products were formed. Gigot et al. [229] studied conversion of 13-HPOT by sugar beet HPL extracted from leaves or expressed by recombinant E. coli strains. With the adaptation of a fed-batch substrate addition and a continuous extraction of volatiles, at 2 L scale, 3.46 mM of C6 aldehydes were produced with the native HPL extracted, while 5.5 mM of C6 aldehydes were produced with recombinant HPL and further no side products from the LOX pathway were formed. Whitehead et al. [183] patented a method using recombinant guava HPL expressed in E. coli, producing 14 g of hexanal per liter of bacterial lysate. Recently, Jacopini et al. [108] isolated and cloned a cDNA encoding for a 13-HPL from black olive fruits (Olea europaea L., Leccino variety). Recombinant olive HPL was expressed in E. coli, then purified, characterized, and used for bioconversion of 13-HPOD and 13-HPOT. Under optimal biotransformation conditions, 5.61 mM of hexanal and 4.39 mM of (3Z)-hexenal were produced by the action of the recombinant olive HPL, corresponding to high molar conversion yields of 93.5 and 73%, respectively [108]. Recombinant olive HPL appears to be a promising efficient biocatalyst for C6 aldehydes synthesis in a biocatalytic process. The stabilization of the enzyme using selected chemical additives was also investigated. Jacopini et al. [230] showed that about 100% of the HPL activity was maintained during five weeks of storage at −20 or at −80 °C in the presence of glycerol (10%, v/v). The addition of additives such as NaCl, Na2SO4, and glycine to the reaction medium has increased the catalytic efficiency of the enzyme. During C6 aldehydes biosynthesis in the presence of these compounds, amounts of aldehydes equivalent to those obtained in their absence, and high molar conversion rates were achieved, while the amount of enzyme used was decreased from 1.5 to 2.5 fold [230].
If needed, biocatalysts can be engineered using protein and strain engineering tools to increase their performance to meet process requirements. The performance of the 13-HPL from guava, has been improved by directed evolution [231]. Engineered 13-HPL showed increased total turnover number, higher solubility, and thermal stability that translated into lower catalyst loading. Brühlmann and Bosijokovic [232] showed that the use of a ketoreductase overproduced in a microorganism instead of baker’s yeast may offer higher selectivity and increased volumetric yields at reduced catalyst loading. Finally, combining both catalysts (engineered recombinant HPL and recombinant ketoreductase) at reasonable loading in a simple one-pot cascade reaction offered the GLV (3Z)-hexenol at high isomeric purity (>99%) and high titers (8 g·L−1) [232].
Besides, recombinant yeasts have also been tested for the production of HPL. Bourel et al. [233] and Santiago-Gomez et al. [234] obtained 1200 to 1800 units of green pepper recombinant HPL per liter of culture medium when expressed in yeast Yarrowia lipolytica, and 6 mM of hexanal was obtained from 119 mM of 13-HPOs. Another yeast, Pichia pastoris was used by Atwal et al. [235] to express tomato HPL as a secreted enzyme. Buchhaupt et al. [236] have described a process for the production of aldehydes and 6-carbon alcohols by the yeast S. cerevisiae coexpressing soybean 13-LOX and watermelon 13-HPL. However, the enzyme levels produced in yeasts are much lower than those obtained in E. coli.
Another strategy is to use the phytopathogenic bacterium Agrobacterium tumefaciens to transfect the genes encoding and overexpressing the enzymes of the LOX pathway in a plant. Thus, the gene encoding watermelon 13-HPL was used to overexpress the enzyme in Nicotiana benthamiana and Nicotiana tabacum [129,237]. In the transgenic leaves, watermelon 13-HPL activity was respectively 80 and 50 higher than endogenous HPL activity in the wild-type plants.
GLVs compounds produced from HPOs may be included in mixtures in the flavor and perfume industry and the food industry.
6. Conclusions
In addition to acting as defense molecules in plants under stress conditions, GLVs have interesting flavoring properties commonly used in the flavor and fragrance industry, as well as in food technology. Nowadays, the demand for natural GLVs still growing, but large-scale synthesis remains a major challenge for producers. The use of the naturally occurring LOX pathway in plants has led to the development of efficient and environmentally friendly biotechnological processes for the production of natural GLVs. Indeed, in such processes the enzymatic activities of the LOX pathway used to perform the bioconversion of a natural raw material, renewable, and inexpensive, which is often a vegetable oil, to natural GLVs with high added value.
Enzymatic engineering technology has contributed to the improvement of process efficiency through the production of recombinant and/or engineered enzymes, and the use of optimized bioreactors for continuous synthesis. HPL is the essential and critical step of the biotechnological process, being the limiting factor for the GLVs production. Biotechnological tools are to be developed in order to improve this step. The production of recombinant HPL in large scale and its stabilization to raise HPL levels, as well as the extraction of intermediate and final products are important ways to explore.
Author Contributions
Conceptualization: S.V., V.B.-B.d.C., L.B. and J.M.; Methodology: S.V., M.M., J.-C.A., S.J. and V.B.-B.d.C.; Validation: V.B.-B.d.C., L.B. and J.M.; Formal Analysis: S.V., V.B.-B.d.C., L.B. and J.M.; Investigation: S.V., M.M., J.-C.A., S.J., V.B.-B.d.C., L.B. and J.M.; Resources: V.B.-B.d.C., L.B. and J.M.; Writing—Original Draft Preparation, S.V. and J.M.; Writing—Review & Editing, S.V., V.B.-B.d.C., L.B. and J.M.; Supervision: L.B. and J.M.; Project Administration: L.B. and J.M.; Funding Acquisition: L.B. and J.M.
Funding
This research received no external funding.
Acknowledgments
This work has been supported by the Collectivité de Corse and the FEDER project 2014–2020, AGRIEX No. CO 0009204.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hatanaka, A.; Kajiwara, T.; Sekiya, J. Biosynthetic pathway for C6-aldehydes formation from linolenic acid in green leaves. Chem. Phys. Lipids 1987, 44, 341–361. [Google Scholar] [CrossRef]
- Hatanaka, A. The biogeneration of green odour by green leaves. Phytochemistry 1993, 34, 1201–1218. [Google Scholar] [CrossRef]
- Hassan, M.N.; Zainal, Z.; Ismail, I. Green leaf volatiles: Biosynthesis, biological functions and their applications in biotechnology. Plant Biotechnol. J. 2015, 13, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Blee, E.; Joyard, J. Envelope membranes from spinach chloroplasts are a site of metabolism of fatty acid hydroperoxides. Plant Physiol. 1996, 110, 445–454. [Google Scholar] [CrossRef]
- Bate, N.J.; Rothstein, S.J. C6-volatiles derived from the lipoxygenase pathway induce a subset of defense-related genes. Plant J. 1998, 16, 561–569. [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef]
- Grechkin, A.N. Recent developments in biochemistry of the plant lipoxygenase pathway. Prog. Lipid Res. 1998, 37, 317–352. [Google Scholar] [CrossRef]
- Blee, E. Impact of phyto-oxylipins in plant defense. Trends Plant Sci. 2002, 7, 315–322. [Google Scholar] [CrossRef]
- Howe, G.A.; Schilmiller, A.L. Oxylipin metabolism in response to stress. Curr. Opin. Plant Biol. 2002, 5, 230–236. [Google Scholar] [CrossRef]
- Blee, E. Phytooxylipins and plant defense reactions. Prog. Lipid Res. 1998, 37, 33–72. [Google Scholar] [CrossRef]
- Griffiths, G. Biosynthesis and analysis of plant oxylipins. Free Radic. Res. 2015, 49, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Drouet, P.; Thomas, D.; Legoy, M.D. Production of 13(S)-hydroperoxy-9(Z),11(E)-octadecadienoic acid using soybean lipoxygenase 1 in a biphasic octane-water system. Tetrahedron Lett. 1994, 35, 3923–3926. [Google Scholar] [CrossRef]
- Fauconnier, M.L.; Marlier, M. An efficient procedure for the production of fatty acid hydroperoxides from hydrolyzed flax seed oil and soybean lipoxygenase. Biotechnol. Tech. 1996, 10, 839–844. [Google Scholar] [CrossRef]
- Andre, E.; Hou, K.-W. Présence d’une oxydase des lipides ou lipoxydase dans la graine de soja. Comptes Rendus de l’Académie des Sciences (Paris) 1932, 194, 645–647. [Google Scholar]
- Gardner, H.W. Recent investigations into the lipoxygenase pathway of plants. Biochim. Biophys. Acta 1991, 1084, 221–239. [Google Scholar] [CrossRef]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef]
- Ishiguro, S.; Kawai-Oda, A.; Ueda, J.; Nishida, I.; Okada, K. The defective in anther dehiscence1 Gene Encodes a Novel Phospholipase A1 Catalyzing the Initial Step of Jasmonic Acid Biosynthesis, Which Synchronizes Pollen Maturation, Anther Dehiscence, and Flower Opening in Arabidopsis. Plant Cell 2001, 13, 2191–2210. [Google Scholar] [CrossRef]
- Vliegenthart, J.F.G.; Veldink, G.A. Substrates and products of lipoxygenase catalysis. Stud. Nat. Prod. Chem. 1991, 9, 559–589. [Google Scholar]
- Schwab, W. Biosynthesis of Plant Flavors: Analysis and Biotechnological Approach. In Flavor Chemistry; American Chemical Society: Washington, DC, USA, 2000; Volume 756, pp. 72–86. [Google Scholar]
- Vick, B.A.; Zimmerman, D.C. Pathways of Fatty Acid hydroperoxide metabolism in spinach leaf chloroplasts. Plant Physiol. 1987, 85, 1073–1078. [Google Scholar] [CrossRef]
- Hughes, R.K.; De Domenico, S.; Santino, A. Plant cytochrome CYP74 family: Biochemical features, endocellular localisation, activation mechanism in plant defence and improvements for industrial applications. ChemBioChem 2009, 10, 1122–1133. [Google Scholar] [CrossRef]
- Mosblech, A.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009, 47, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Itoh, A.; Schilmiller, A.L.; McCaig, B.C.; Howe, G.A. Identification of a jasmonate-regulated allene oxide synthase that metabolizes 9-hydroperoxides of linoleic and linolenic acids. J. Biol. Chem. 2002, 277, 46051–46058. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, I.; Hause, B.; Maucher, H.; Pitzschke, A.; Miersch, O.; Ziegler, J.; Ryan, C.A.; Wasternack, C. Allene oxide cyclase dependence of the wound response and vascular bundle-specific generation of jasmonates in tomato - amplification in wound signalling. Plant J. 2003, 33, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Grechkin, A.N.; Hamberg, M. Divinyl ether synthase from garlic (Allium sativum L.) bulbs: Sub-cellular localization and substrate regio- and stereospecificity. FEBS Lett. 1996, 388, 112–114. [Google Scholar] [CrossRef]
- Grechkin, A.N. Hydroperoxide lyase and divinyl ether synthase. Prostaglandins Other Lipid Mediat. 2002, 68–69, 457–470. [Google Scholar] [CrossRef]
- Grechkin, A.N.; Mukhtarova, L.S.; Hamberg, M. Detection of an enol intermediate in the hydroperoxide lyase chain cleavage reaction. FEBS Lett. 2003, 549, 31–34. [Google Scholar] [CrossRef]
- Grechkin, A.N.; Hamberg, M. The “heterolytic hydroperoxide lyase” is an isomerase producing a short-lived fatty acid hemiacetal. Biochim. Biophys. Acta 2004, 1636, 47–58. [Google Scholar] [CrossRef]
- Grechkin, A.N.; Bruhlmann, F.; Mukhtarova, L.S.; Gogolev, Y.V.; Hamberg, M. Hydroperoxide lyases (CYP74C and CYP74B) catalyze the homolytic isomerization of fatty acid hydroperoxides into hemiacetals. Biochim. Biophys. Acta 2006, 1761, 1419–1428. [Google Scholar] [CrossRef]
- Gobel, C.; Feussner, I.; Schmidt, A.; Scheel, D.; Sanchez-Serrano, J.; Hamberg, M.; Rosahl, S. Oxylipin profiling reveals the preferential stimulation of the 9-lipoxygenase pathway in elicitor-treated potato cells. J. Biol. Chem. 2001, 276, 6267–6273. [Google Scholar] [CrossRef]
- Hamberg, M. An epoxy alcohol synthase pathway in higher plants: Biosynthesis of antifungal trihydroxy oxylipins in leaves of potato. Lipids 1999, 34, 1131–1142. [Google Scholar] [CrossRef]
- Kuhn, H.; Wiesner, R.; Rathmann, J.; Schewe, T. Formation of ketodienoic fatty acids by the pure pea lipoxygenase-1. Eicosanoids 1991, 4, 9–14. [Google Scholar] [PubMed]
- Conconi, A.; Miquel, M.; Browse, J.A.; Ryan, C.A. Intracellular Levels of Free Linolenic and Linoleic Acids Increase in Tomato Leaves in Response to Wounding. Plant Physiol. 1996, 111, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T. Jasmonate-induced responses are costly but benefit plants under attack in native populations. Proc. Natl. Acad. Sci. USA 1998, 95, 8113–8118. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Schwab, W. Cloning and characterization of a 9-lipoxygenase gene induced by pathogen attack from Nicotiana benthamiana for biotechnological application. BMC Biotechnol. 2011, 11, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Kost, C.; Boland, W. Herbivore-induced, indirect plant defences. Biochim. Biophys. Acta 2005, 1734, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Kramell, R.; Atzorn, R.; Schneider, G.; Miersch, O.; Brückner, C.; Schmidt, J.; Sembdner, G.; Parthier, B. Occurrence and identification of jasmonic acid and its amino acid conjugates induced by osmotic stress in barley leaf tissue. J. Plant Growth Regul. 1995, 14, 29. [Google Scholar] [CrossRef]
- Conconi, A.; Smerdon, M.J.; Howe, G.A.; Ryan, C.A. The octadecanoid signalling pathway in plants mediates a response to ultraviolet radiation. Nature 1996, 383, 826–829. [Google Scholar] [CrossRef]
- Porta, H.; Rueda-Benitez, P.; Campos, F.; Colmenero-Flores, J.M.; Colorado, J.M.; Carmona, M.J.; Covarrubias, A.A.; Rocha-Sosa, M. Analysis of lipoxygenase mRNA accumulation in the common bean (Phaseolus vulgaris L.) during development and under stress conditions. Plant Cell Physiol. 1999, 40, 850–858. [Google Scholar] [CrossRef]
- Savchenko, T.; Dehesh, K. Drought stress modulates oxylipin signature by eliciting 12-OPDA as a potent regulator of stomatal aperture. Plant Signal. Behav. 2014, 9, e28304. [Google Scholar] [CrossRef]
- Savchenko, T.V.; Zastrijnaja, O.M.; Klimov, V.V. Oxylipins and plant abiotic stress resistance. Biochemistry (Moscow) 2014, 79, 362–375. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Prost, I.; Dhondt, S.; Rothe, G.; Vicente, J.; Rodriguez, M.J.; Kift, N.; Carbonne, F.; Griffiths, G.; Esquerre-Tugaye, M.T.; Rosahl, S.; et al. Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in defense against pathogens. Plant Physiol. 2005, 139, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Hatanaka, A. Green-leaf-derived C6-aroma compounds with potent antibacterial action that act on both gram-negative and gram-positive bacteria. J. Agric. Food Chem. 2002, 50, 7639–7644. [Google Scholar] [CrossRef] [PubMed]
- Croft, K.; Juttner, F.; Slusarenko, A.J. Volatile products of the lipoxygenase pathway evolved from Phaseolus vulgaris (L.) leaves inoculated with Pseudomonas syringae pv phaseolicola. Plant Physiol. 1993, 101, 13–24. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Direct fungicidal activities of C6-aldehydes are important constituents for defense responses in Arabidopsis against Botrytis cinerea. Phytochemistry 2008, 69, 2127–2132. [Google Scholar] [CrossRef]
- Matsui, K.; Minami, A.; Hornung, E.; Shibata, H.; Kishimoto, K.; Ahnert, V.; Kindl, H.; Kajiwara, T.; Feussner, I. Biosynthesis of fatty acid derived aldehydes is induced upon mechanical wounding and its products show fungicidal activities in cucumber. Phytochemistry 2006, 67, 649–657. [Google Scholar] [CrossRef]
- Creelman, R.A.; Tierney, M.L.; Mullet, J.E. Jasmonic acid/methyl jasmonate accumulate in wounded soybean hypocotyls and modulate wound gene expression. Proc. Natl. Acad. Sci. USA 1992, 89, 4938–4941. [Google Scholar] [CrossRef]
- Zimmerman, D.C.; Coudron, C.A. Identification of traumatin, a wound hormone, as 12-Oxo-trans-10-dodecenoic acid. Plant Physiol. 1979, 63, 536–541. [Google Scholar] [CrossRef]
- Chehab, E.W.; Kaspi, R.; Savchenko, T.; Rowe, H.; Negre-Zakharov, F.; Kliebenstein, D.; Dehesh, K. Distinct roles of jasmonates and aldehydes in plant-defense responses. Public Libr. Sci. One 2008, 3, e1904. [Google Scholar] [CrossRef]
- Royo, J.; Leon, J.; Vancanneyt, G.; Albar, J.P.; Rosahl, S.; Ortego, F.; Castanera, P.; Sanchez-Serrano, J.J. Antisense-mediated depletion of a potato lipoxygenase reduces wound induction of proteinase inhibitors and increases weight gain of insect pests. Proc. Natl. Acad. Sci. USA 1999, 96, 1146–1151. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, R.; Arimura, G.; Takabayashi, J.; Shimoda, T.; Nishioka, T. Involvement of jasmonate- and salicylate-related signaling pathways for the production of specific herbivore-induced volatiles in plants. Plant Cell Physiol. 2000, 41, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Leitner, M.; Boland, W.; Mithofer, A. Direct and indirect defences induced by piercing-sucking and chewing herbivores in Medicago truncatula. New Phytol. 2005, 167, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Shimoda, T.; Nishioka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Kohlmann, M.; Bachmann, A.; Weichert, H.; Kolbe, A.; Balkenhohl, T.; Wasternack, C.; Feussner, I. Formation of lipoxygenase-pathway-derived aldehydes in barley leaves upon methyl jasmonate treatment. Eur. J. Biochem. 1999, 260, 885–895. [Google Scholar] [CrossRef]
- Baldwin, I.T. Plant volatiles. Curr. Biol. 2010, 20, R392–R397. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Ozawa, R.; Horiuchi, J.-I.; Nishioka, T.; Takabayashi, J. Plant–plant interactions mediated by volatiles emitted from plants infested by spider mites. Biochem. Syst. Ecol. 2001, 29, 1049–1061. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “Talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Pichersky, E.; Schaub, A.; Hansel, A.; Gershenzon, J. Characterization of a BAHD acyltransferase responsible for producing the green leaf volatile (Z)-3-hexen-1-yl acetate in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2007, 49, 194–207. [Google Scholar] [CrossRef]
- Allmann, S.; Baldwin, I.T. Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles. Science 2010, 329, 1075–1078. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.D. Phospholipase activity during plant growth and development and in response to environmental stress. Trends Plant Sci. 1998, 3, 419–426. [Google Scholar] [CrossRef]
- Tuteja, N.; Sopory, S.K. Chemical signaling under abiotic stress environment in plants. Plant Signal. Behav. 2008, 3, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Kurishita, S.; Hisamitsu, A.; Kajiwara, T. A lipid-hydrolysing activity involved in hexenal formation. Biochem. Soc. Trans. 2000, 28, 857–860. [Google Scholar] [CrossRef]
- Narvaez-Vasquez, J.; Florin-Christensen, J.; Ryan, C.A. Positional specificity of a phospholipase A activity induced by wounding, systemin, and oligosaccharide elicitors in tomato leaves. Plant Cell 1999, 11, 2249–2260. [Google Scholar] [CrossRef]
- Wang, C.; Zien, C.A.; Afitlhile, M.; Welti, R.; Hildebrand, D.F.; Wang, X. Involvement of phospholipase D in wound-induced accumulation of jasmonic acid in arabidopsis. Plant Cell 2000, 12, 2237–2246. [Google Scholar] [CrossRef]
- Hyun, Y.; Choi, S.; Hwang, H.J.; Yu, J.; Nam, S.J.; Ko, J.; Park, J.Y.; Seo, Y.S.; Kim, E.Y.; Ryu, S.; et al. Cooperation and functional diversification of two closely related galactolipase genes for jasmonate biosynthesis. Dev. Cell 2008, 14, 183–192. [Google Scholar] [CrossRef]
- Yang, W.Y.; Zheng, Y.; Bahn, S.C.; Pan, X.Q.; Li, M.Y.; Vu, H.S.; Roth, M.R.; Scheu, B.; Welti, R.; Hong, Y.Y.; et al. The patatin-containing phospholipase A pPLAIIalpha modulates oxylipin formation and water loss in Arabidopsis thaliana. Mol. Plant 2012, 5, 452–460. [Google Scholar] [CrossRef]
- Bonaventure, G.; Schuck, S.; Baldwin, I.T. Revealing complexity and specificity in the activation of lipase-mediated oxylipin biosynthesis: A specific role of the Nicotiana attenuata GLA1 lipase in the activation of jasmonic acid biosynthesis in leaves and roots. Plant Cell Environ. 2011, 34, 1507–1520. [Google Scholar] [CrossRef] [PubMed]
- Stelmach, B.A.; Müller, A.; Hennig, P.; Gebhardt, S.; Schubert-Zsilavecz, M.; Weiler, E.W. A novel class of oxylipins, sn1-O-(12-oxophytodienoyl)-sn2-O-(hexadecatrienoyl)-monogalactosyl Diglyceride, from Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 12832–12838. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; von Reuss, S.H.; Tasaka, H.; Nomura, M.; Mochizuki, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Boland, W.; Takabayashi, J.; et al. Traumatin- and dinortraumatin-containing galactolipids in Arabidopsis: Their formation in tissue-disrupted leaves as counterparts of green leaf volatiles. J. Biol. Chem. 2013, 288, 26078–26088. [Google Scholar] [CrossRef] [PubMed]
- Mwenda, C.M.; Matsui, K. The importance of lipoxygenase control in the production of green leaf volatiles by lipase-dependent and independent pathways. Plant Biotechnol. 2014, 31, 445–452. [Google Scholar] [CrossRef]
- Nakashima, A.; Iijima, Y.; Aoki, K.; Shibata, D.; Sugimoto, K.; Takabayashi, J.; Matsui, K. Monogalactosyl diacylglycerol is a substrate for lipoxygenase: Its implications for oxylipin formation directly from lipids. J. Plant Interact. 2011, 6, 93–97. [Google Scholar] [CrossRef]
- Siedow, J.N. Plant lipoxygenase: Structure and function. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 145–188. [Google Scholar] [CrossRef]
- Perez, A.G.; Sanz, C.; Olias, R.; Olias, J.M. Lipoxygenase and hydroperoxide lyase activities in ripening strawberry fruits. J. Agric. Food Chem. 1999, 47, 249–253. [Google Scholar] [CrossRef]
- Porta, H.; Rocha-Sosa, M. Plant lipoxygenases. Physiological and molecular features. Plant Physiol. 2002, 130, 15–21. [Google Scholar] [CrossRef]
- Feussner, I.; Hause, B.; Vörös, K.; Parthier, B.; Wasternack, C. Jasmonate-induced lipoxygenase forms are localized in chloroplasts of barley leaves (Hordeum vulgare cv. Salome). Plant J. 1995, 7, 949–957. [Google Scholar] [CrossRef]
- Fauconnier, M.-L.; Marlier, M. Les lipoxygénases du soja. Biotechnol. Agron. Société Environ. 1997, 1, 125–141. [Google Scholar]
- Ivanov, I.; Heydeck, D.; Hofheinz, K.; Roffeis, J.; O’Donnell, V.; Kuhn, H.; Walther, M. Molecular enzymology of lipoxygenases. Arch. Biochem. Biophys. 2010, 503, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Wasternack, C.; Kindl, H.; Kühn, H. Lipoxygenase-catalyzed oxygenation of storage lipids is implicated in lipid mobilization during germination. Proc. Natl. Acad. Sci. USA 1995, 92, 11849–11853. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M.A.; Weichert, H.; Fischer, A.M.; Feussner, I.; Grimes, H.D. Activity of soybean lipoxygenase isoforms against esterified fatty acids indicates functional specificity. Arch. Biochem. Biophys. 2001, 388, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Belkner, J.; Wiesner, R.; Kuhn, H.; Lankin, V.Z. The oxygenation of cholesterol esters by the reticulocyte lipoxygenase. FEBS Lett. 1991, 279, 110–114. [Google Scholar] [CrossRef]
- Liavonchanka, A.; Feussner, I. Lipoxygenases: Occurrence, functions and catalysis. J. Plant Physiol. 2006, 163, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Palmieri-Thiers, C.; Canaan, S.P.; Brunini, V.; Lorenzi, V.; Tomi, F.L.; Desseyn, J.-L.; Garscha, U.; Oliw, E.H.; Berti, L.; Maury, J. A lipoxygenase with dual positional specificity is expressed in olives (Olea europaea L.) during ripening. Biochim. Biophys. Acta 2009, 1791, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Palmieri-Thiers, C.; Alberti, J.-C.; Canaan, S.P.; Brunini, V.; Gambotti, C.; Tomi, F.L.; Oliw, E.H.; Berti, L.; Maury, J. Identification of putative residues involved in the accessibility of the substrate-binding site of lipoxygenase by site-directed mutagenesis studies. Arch. Biochem. Biophys. 2011, 509, 82–89. [Google Scholar] [CrossRef]
- Schiller, D.; Contreras, C.; Vogt, J.; Dunemann, F.; Defilippi, B.G.; Beaudry, R.; Schwab, W. A dual positional specific lipoxygenase functions in the generation of flavor compounds during climacteric ripening of apple. Hortic. Res. 2015, 2, 15003. [Google Scholar] [CrossRef]
- Mochizuki, S.; Sugimoto, K.; Koeduka, T.; Matsui, K. Arabidopsis lipoxygenase 2 is essential for formation of green leaf volatiles and five-carbon volatiles. FEBS Lett. 2016, 590, 1017–1027. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S. [53] Lipoxygenase from soybeans: EC 1.13.11.12 Linoleate:oxygen oxidoreductase. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1981; Volume 71, pp. 441–451. [Google Scholar]
- Minor, W.; Steczko, J.; Bolin, J.T.; Otwinowski, Z.; Axelrod, B. Crystallographic determination of the active site iron and its ligands in soybean lipoxygenase L-1. Biochemistry 1993, 32, 6320–6323. [Google Scholar] [CrossRef]
- Minor, W.; Steczko, J.; Stec, B.; Otwinowski, Z.; Bolin, J.T.; Walter, R.; Axelrod, B. Crystal structure of soybean lipoxygenase L-1 at 1.4 A resolution. Biochemistry 1996, 35, 10687–10701. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, N.; Solomon, E.I. Density-functional investigation on the mechanism of H-atom abstraction by lipoxygenase. J. Biol. Inorg. Chem. 2003, 8, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Gillmor, S.A.; Villasenor, A.; Fletterick, R.; Sigal, E.; Browner, M.F. The structure of mammalian 15-lipoxygenase reveals similarity to the lipases and the determinants of substrate specificity. Nat. Struct. Biol. 1997, 4, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Borngräber, S.; Browner, M.; Gillmor, S.; Gerth, C.; Anton, M.; Fletterick, R.; Kühn, H. Shape and Specificity in Mammalian 15-Lipoxygenase Active Site The Functional Interplay Of Sequence Determinants For The Reaction Specificity. J. Biol. Chem. 1999, 274, 37345–37350. [Google Scholar] [CrossRef]
- Hughes, R.K.; Lawson, D.M.; Hornostaj, A.R.; Fairhurst, S.A.; Casey, R. Mutagenesis and modelling of linoleate-binding to pea seed lipoxygenase. Eur. J. Biochem. 2001, 268, 1030–1040. [Google Scholar] [CrossRef]
- Gardner, H.W. Soybean lipoxygenase-1 enzymically forms both (9S)- and (13S)-hydroperoxides from linoleic acid by a pH-dependent mechanism. Biochim. Biophys. Acta 1989, 1001, 274–281. [Google Scholar] [CrossRef]
- Hornung, E.; Walther, M.; Kühn, H.; Feussner, I. Conversion of cucumber linoleate 13-lipoxygenase to a 9-lipoxygenating species by site-directed mutagenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 4192–4197. [Google Scholar] [CrossRef]
- Nanda, S.; Yadav, J.S. Lipoxygenase biocatalysis: A survey of asymmetric oxygenation. J. Mol. Catal. B Enzym. 2003, 26, 3–28. [Google Scholar] [CrossRef]
- Coffa, G.; Imber, A.N.; Maguire, B.C.; Laxmikanthan, G.; Schneider, C.; Gaffney, B.J.; Brash, A.R. On the relationships of substrate orientation, hydrogen abstraction, and product stereochemistry in single and double dioxygenations by soybean lipoxygenase-1 and its Ala542Gly mutant. J. Biol. Chem. 2005, 280, 38756–38766. [Google Scholar] [CrossRef]
- Coffa, G.; Schneider, C.; Brash, A.R. A comprehensive model of positional and stereo control in lipoxygenases. Biochem. Biophys. Res. Commun. 2005, 338, 87–92. [Google Scholar] [CrossRef]
- Fauconnier, M.L.; Perez, A.G.; Sanz, C.; Marlier, M. Purification and characterization of tomato leaf (Lycopersicon esculentum mill.) hydroperoxide lyase. J. Agric. Food Chem. 1997, 45, 4232–4236. [Google Scholar] [CrossRef]
- Gargouri, M.; Drouet, P.; Legoy, M.D. Hydroperoxide-lyase activity in mint leaves. Volatile C6-aldehyde production from hydroperoxy-fatty acids. J. Biotechnol. 2004, 111, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Itoh, A.; Vick, B.A. The purification and characterization of fatty acid hydroperoxide lyase in sunflower. Biochim. Biophys. Acta 1999, 1436, 531–540. [Google Scholar] [CrossRef]
- Salas, J.Ì.N.J.; Sanchez, J. Hydroperoxide lyase from olive (Olea europaea) fruits. Plant Sci. 1999, 143, 19–26. [Google Scholar] [CrossRef]
- Padilla, M.N.; Hernandez, M.L.; Perez, A.G.; Sanz, C.; Martinez-Rivas, J.M. Isolation, expression, and characterization of a 13-hydroperoxide lyase gene from olive fruit related to the biosynthesis of the main virgin olive oil aroma compounds. J. Agric. Food Chem. 2010, 58, 5649–5657. [Google Scholar] [CrossRef]
- Jacopini, S.; Mariani, M.; Brunini-Bronzini de Caraffa, V.; Gambotti, C.; Vincenti, S.; Desjobert, J.-M.; Muselli, A.; Costa, J.; Berti, L.; Maury, J. Olive recombinant hydroperoxide lyase, an efficient biocatalyst for synthesis of green leaf volatiles. Appl. Biochem. Biotechnol. 2016, 179, 671–683. [Google Scholar] [CrossRef]
- Shibata, Y.; Matsui, K.; Kajiwara, T.; Hatanaka, A. Purification and Properties of Fatty Acid Hydroperoxide Lyase from Green Bell Pepper Fruits. Plant Cell Physiol. 1995, 36, 147–156. [Google Scholar]
- Husson, F.; Belin, J.M. Purification of hydroperoxide lyase from green bell pepper (Capsicum annuum L.) fruits for the generation of C6-aldehydes in vitro. J. Agric. Food Chem. 2002, 50, 1991–1995. [Google Scholar] [CrossRef]
- Matsui, K.; Ujita, C.; Fujimoto, S.; Wilkinson, J.; Hiatt, B.; Knauf, V.; Kajiwara, T.; Feussner, I. Fatty acid 9- and 13-hydroperoxide lyases from cucumber. FEBS Lett. 2000, 481, 183–188. [Google Scholar] [CrossRef]
- Matoba, T.; Hidaka, H.; Kitamura, K.; Kaizuma, N.; Kito, M. Contribution of hydroperoxide lyase activity to n-hexanal formation in soybean. J. Agric. Food Chem. 1985, 33, 856–858. [Google Scholar] [CrossRef]
- Kuroda, H.; Oshima, T.; Kaneda, H.; Takashio, M. Identification and functional analyses of two cDNAs that encode fatty acid 9-/13-hydroperoxide lyase (CYP74C) in rice. Biosci. Biotechnol. Biochem. 2005, 69, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, A.; Kajiwara, T.; Matsui, K.; Toyota, H. Substrate specificity of tea leaf hydroperoxide lyase. Zeitschrift für Naturforschung 1992, 47c, 379–677. [Google Scholar] [CrossRef][Green Version]
- Noordermeer, M.A.; Veldink, G.A.; Vliegenthart, J.F. Fatty acid hydroperoxide lyase: A plant cytochrome p450 enzyme involved in wound healing and pest resistance. Chembiochem 2001, 2, 494–504. [Google Scholar] [CrossRef]
- Kajiwara, T.; Sekiya, J.; Asano, M.; Hatanaka, A. Enantioselectivity of enzymatic cleavage reaction of 13-hydroperoxylinolenic acid to C6-aldehyde and C12-oxo acid in tea chloroplasts. Agric. Biol. Chem. 1982, 46, 3087–3088. [Google Scholar]
- Hatanaka, A.; Kajiwara, T.; Sekiya, J.; Imoto, M.; Inouye, S. Participation and properties of lipoxygenase and hydroperoxide lyase in volatile C6-aldehyde formation from C18-Unsaturated. Fatty acids in isolated tea chloroplasts. Plant Cell Physiol. 1982, 23, 91–99. [Google Scholar] [CrossRef]
- Kim, I.S.; Grosch, W. Partial purification and properties of a hydroperoxide lyase from fruits of pear. J. Agric. Food Chem. 1981, 29, 1220–1225. [Google Scholar] [CrossRef]
- Gardner, H.W.; Weisleder, D.; Plattner, R.D. Hydroperoxide lyase and other hydroperoxide-metabolizing activity in tissues of soybean, Glycine max. Plant Physiol. 1991, 97, 1059–1072. [Google Scholar] [CrossRef][Green Version]
- Tijet, N.; Schneider, C.; Muller, B.L.; Brash, A.R. Biogenesis of volatile aldehydes from fatty acid hydroperoxides: Molecular cloning of a hydroperoxide lyase (CYP74C) with specificity for both the 9- and 13-hydroperoxides of linoleic and linolenic acids. Arch. Biochem. Biophys. 2001, 386, 281–289. [Google Scholar] [CrossRef]
- Mita, G.; Quarta, A.; Fasano, P.; De Paolis, A.; Di Sansebastiano, G.P.; Perrotta, C.; Iannacone, R.; Belfield, E.; Hughes, R.; Tsesmetzis, N.; et al. Molecular cloning and characterization of an almond 9-hydroperoxide lyase, a new CYP74 targeted to lipid bodies. J. Exp. Bot. 2005, 56, 2321–2333. [Google Scholar] [CrossRef]
- Vick, B.A.; Zimmerman, D.C. Lipoxygenase and hydroperoxide lyase in germinating watermelon seedlings. Plant Physiol. 1976, 57, 780–788. [Google Scholar] [CrossRef]
- Sekiya, J.; Kajiwara, T.; Hatanaka, A. Volatile C6-Aldehyde Formation via Hydroperoxides from C18-Unsaturated Fatty Acids in Etiolated Alfalfa and Cucumber Seedlings. Agric. Biol. Chem. 1979, 43, 969–980. [Google Scholar] [CrossRef]
- Noordermeer, M.A.; Van Dijken, A.J.; Smeekens, S.C.; Veldink, G.A.; Vliegenthart, J.F. Characterization of three cloned and expressed 13-hydroperoxide lyase isoenzymes from alfalfa with unusual N-terminal sequences and different enzyme kinetics. Eur. J. Biochem. 2000, 267, 2473–2482. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Miyahara, C.; Wilkinson, J.; Hiatt, B.; Knauf, V.; Kajiwara, T. Fatty acid hydroperoxide lyase in tomato fruits: Cloning and properties of a recombinant enzyme expressed in Escherichia coli. Biosci. Biotechnol. Biochem. 2000, 64, 1189–1196. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suurmeijer, C.N.; Perez-Gilabert, M.; van Unen, D.J.; van der Hijden, H.T.; Veldink, G.A.; Vliegenthart, J.F. Purification, stabilization and characterization of tomato fatty acid hydroperoxide lyase. Phytochemistry 2000, 53, 177–185. [Google Scholar] [CrossRef][Green Version]
- Mu, W.; Xue, Q.; Jiang, B.; Hua, Y. Molecular cloning, expression, and enzymatic characterization of Solanum tuberosum hydroperoxide lyase. Eur. Food Res. Technol. 2012, 234, 723–731. [Google Scholar] [CrossRef]
- Rabetafika, H.N.; Gigot, C.; Fauconnier, M.-L.; Ongena, M.; Destain, J.; Du Jardin, P.; Wathelet, J.-P.; Thonart, P. Sugar beet leaves as new source of hydroperoxide lyase in a bioprocess producing green-note aldehydes. Biotechnol. Lett. 2008, 30, 1115–1119. [Google Scholar] [CrossRef]
- Fukushige, H.; Hildebrand, D.F. Watermelon (Citrullus lanatus) hydroperoxide lyase greatly increases C6 aldehyde formation in transgenic leaves. J. Agric. Food Chem. 2005, 53, 2046–2051. [Google Scholar] [CrossRef]
- De Domenico, S.; Tsesmetzis, N.; Di Sansebastiano, G.P.; Hughes, R.K.; Casey, R.; Santino, A. Subcellular localisation of Medicago truncatula 9/13-hydroperoxide lyase reveals a new localisation pattern and activation mechanism for CYP74C enzymes. BMC Plant Biol. 2007, 7, 58–70. [Google Scholar] [CrossRef]
- Zhu, B.-Q.; Xu, X.-Q.; Wu, Y.-W.; Duan, C.-Q.; Pan, Q.-H. Isolation and characterization of two hydroperoxide lyase genes from grape berries: HPL isogenes in Vitis vinifera grapes. Mol. Biol. Rep. 2012, 39, 7443–7455. [Google Scholar] [CrossRef]
- Hatanaka, A.; Kajiwara, T.; Matsui, K. Concentration of hydroperoxide lyase activities in root of cucumber seedling. Zeitschrift Naturforschung 1988, 43c, 308–310. [Google Scholar] [CrossRef]
- Gigot, C.; Ongena, M.; Fauconnier, M.-L.; Wathelet, J.-P.; Du Jardin, P.; Thonart, P. The lipoxygenase metabolic pathway in plants: Potential for industrial production of natural green leaf volatiles. Biotechnol. Agron. Soc. Environ. 2010, 14, 451–460. [Google Scholar]
- Howe, G.A.; Lee, G.I.; Itoh, A.; Li, L.; DeRocher, A.E. Cytochrome P450-dependent metabolism of oxylipins in tomato. Cloning and expression of allene oxide synthase and fatty acid hydroperoxide lyase. Plant Physiol. 2000, 123, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Bate, N.J.; Sivasankar, S.; Moxon, C.; Riley, J.M.; Thompson, J.E.; Rothstein, S.J. Molecular characterization of an Arabidopsis gene encoding hydroperoxide lyase, a cytochrome P-450 that is wound inducible. Plant Physiol. 1998, 117, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, J.E.; Itoh, A.; Howe, G.A. Tomato allene oxide synthase and fatty acid hydroperoxide lyase, two cytochrome P450s involved in oxylipin metabolism, are targeted to different membranes of chloroplast envelope. Plant Physiol. 2001, 125, 306–317. [Google Scholar] [CrossRef]
- Maucher, H.; Hause, B.; Feussner, I.; Ziegler, J.R.; Wasternack, C. Allene oxide synthases of barley (Hordeum vulgare cv. Salome): Tissue specific regulation in seedling development. Plant J. 2000, 21, 199–213. [Google Scholar] [CrossRef]
- Shibata, Y.; Matsui, K.; Kajiwara, T.; Hatanaka, A. Fatty acid hydroperoxide lyase is a heme protein. Biochem. Biophys. Res. Commun. 1995, 207, 438–443. [Google Scholar] [CrossRef]
- Tijet, N.; Wäspi, U.; Gaskin, D.J.; Hunziker, P.; Muller, B.L.; Vulfson, E.N.; Slusarenko, A.; Brash, A.R.; Whitehead, I.M. Purification, molecular cloning, and expression of the gene encoding fatty acid 13-hydroperoxide lyase from guava fruit (Psidium guajava). Lipids 2000, 35, 709–720. [Google Scholar] [CrossRef]
- Schreier, P.; Lorenz, G. Separation, partial purification and characterization of a fatty acid hydroperoxide cleaving enzyme from apple and tomato Fruits. Zeitschrift Naturforschung C 1982, 37, 165–173. [Google Scholar] [CrossRef]
- Olias, J.M.; Rios, J.J.; Valle, M.; Zamora, R.; Sanz, L.C.; Axelrod, B. Fatty acid hydroperoxide lyase in germinating soybean seedlings. J. Agric. Food Chem. 1990, 38, 624–630. [Google Scholar] [CrossRef]
- Matsui, K.; Toyota, H.; Kajiwara, T.; Kakuno, T.; Hatanaka, A. Fatty acid hydroperoxide cleaving enzyme, hydroperoxide lyase, from tea leaves. Phytochemistry 1991, 30, 2109–2113. [Google Scholar] [CrossRef]
- Gomi, K.; Yamasaki, Y.; Yamamoto, H.; Akimitsu, K. Characterization of a hydroperoxide lyase gene and effect of C6-volatiles on expression of genes of the oxylipin metabolism in Citrus. J. Plant Physiol. 2003, 160, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Long, Z.; Kong, X.; Zhang, C.; Jiang, B.; Hua, Y. Purification and characterization of hydroperoxide lyase from amaranth tricolor (Amaranthus mangostanus L.) leaves. Eur. Food Res. Technol. 2010, 231, 865–871. [Google Scholar] [CrossRef]
- Matsui, K.; Shibutani, M.; Hase, T.; Kajiwara, T. Bell pepper fruit fatty acid hydroperoxide lyase is a cytochrome P450 (CYP74B). FEBS Lett. 1996, 394, 21–24. [Google Scholar] [CrossRef]
- Santiago-Gomez, M.P.; Kermasha, S.; Nicaud, J.-M.; Belin, J.-M.; Husson, F. Predicted secondary structure of hydroperoxide lyase from green bell pepper cloned in the yeast Yarrowia lipolytica. J. Mol. Catal. B Enzym. 2010, 65, 63–67. [Google Scholar] [CrossRef]
- Panagakou, I.; Touloupakis, E.; Ghanotakis, D.F. Structural characterization of hydroperoxide lyase in dodecyl maltoside by using circular dichroism. Protein J. 2013, 32, 1–6. [Google Scholar] [CrossRef]
- Lee, D.S.; Nioche, P.; Hamberg, M.; Raman, C.S. Structural insights into the evolutionary paths of oxylipin biosynthetic enzymes. Nature 2008, 455, 363–368. [Google Scholar] [CrossRef]
- Li, L.; Chang, Z.; Pan, Z.; Fu, Z.Q.; Wang, X. Modes of heme binding and substrate access for cytochrome P450 CYP74A revealed by crystal structures of allene oxide synthase. Proc. Natl. Acad. Sci. USA 2008, 105, 13883–13888. [Google Scholar] [CrossRef]
- Wan, X.-H.; Chen, S.-X.; Wang, C.-Y.; Zhang, R.-R.; Cheng, S.-Q.; Meng, H.-W.; Shen, X.-Q. Isolation, expression, and characterization of a hydroperoxide lyase gene from cucumber. Int. J. Mol. Sci. 2013, 14, 22082–22101. [Google Scholar] [CrossRef]
- Toporkova, Y.Y.; Gogolev, Y.V.; Mukhtarova, L.S.; Grechkin, A.N. Determinants governing the CYP74 catalysis: Conversion of allene oxide synthase into hydroperoxide lyase by site-directed mutagenesis. FEBS Lett. 2008, 582, 3423–3428. [Google Scholar] [CrossRef]
- Hughes, R.K.; Yousafzai, F.K.; Ashton, R.; Chechetkin, I.R.; Fairhurst, S.A.; Hamberg, M.; Casey, R. Evidence for communality in the primary determinants of CYP74 catalysis and of structural similarities between CYP74 and classical mammalian P450 enzymes. Proteins 2008, 72, 1199–1211. [Google Scholar] [CrossRef]
- Hughes, R.K.; Belfield, E.J.; Muthusamay, M.; Khan, A.; Rowe, A.; Harding, S.E.; Fairhurst, S.A.; Bornemann, S.; Ashton, R.; Thorneley, R.N.; et al. Characterization of Medicago truncatula (barrel medic) hydroperoxide lyase (CYP74C3), a water-soluble detergent-free cytochrome P450 monomer whose biological activity is defined by monomer-micelle association. Biochem. J. 2006, 395, 641–652. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brash, A.R. Mechanistic aspects of CYP74 allene oxide synthases and related cytochrome P450 enzymes. Phytochemistry 2009, 70, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Yoeun, S.; Sukhanov, A.; Han, O. Separation of enzymatic functions and variation of spin state of rice allene oxide synthase-1 by mutation of Phe-92 and Pro-430. Bioorg. Chem. 2016, 68, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Shibata, Y.; Kajiwara, T.; Hatanaka, A. Separation of 13- and 9-hydroperoxide lyase activities in cotyledons of cucumber seedling. Zeitschrift Naturforschung 1989, 44, 883–885. [Google Scholar] [CrossRef]
- Matsui, K.; Kajiwara, T.; Hatanaka, A. Inactivation of tea leaf hydroperoxide lyase by fatty acid hydroperoxide. J. Agric. Food Chem. 1992, 40, 175–178. [Google Scholar] [CrossRef]
- Kemal, C.; Louis-Flamberg, P.; Krupinski-Olsen, R.; Shorter, A.L. Reductive inactivation of soybean lipoxygenase 1 by catechols: A possible mechanism for regulation of lipoxygenase activity. Biochemistry 1987, 26, 7064–7072. [Google Scholar] [CrossRef]
- Noordermeer, M.A.; Veldink, G.A.; Vliegenthart, J.F. Alfalfa contains substantial 9-hydroperoxide lyase activity and a 3Z:2E-enal isomerase. FEBS Lett. 1999, 443, 201–204. [Google Scholar] [CrossRef]
- Salas, J.J.; Sanchez, J.; Ramli, U.S.; Manaf, A.M.; Williams, M.; Harwood, J.L. Biochemistry of lipid metabolism in olive and other oil fruits. Prog. Lipid Res. 2000, 39, 151–180. [Google Scholar] [CrossRef]
- Matsui, K.; Sugimoto, K.; Mano, J.I.; Ozawa, R.; Takabayashi, J. Differential metabolisms of Green Leaf Volatiles in injured and intact parts of a wounded leaf meet distinct ecophysiological requirements. Public Libr. Sci. ONE 2012, 7, e36433. [Google Scholar] [CrossRef]
- Hatanaka, A.; Kajiwara, T.; Sekiya, J. Fatty Acid Hydroperoxide Lyase in Plant Tissues. In Biogeneration of Aromas; American Chemical Society: Cambridge, MA, USA, 1986; Volume 317, pp. 167–175. [Google Scholar]
- Weichert, H.; Kolbe, A.; Kraus, A.; Wasternack, C.; Feussner, I. Metabolic profiling of oxylipins in germinating cucumber seedlings--lipoxygenase-dependent degradation of triacylglycerols and biosynthesis of volatile aldehydes. Planta 2002, 215, 612–619. [Google Scholar] [CrossRef]
- Steinhaus, M.; Sinuco, D.; Polster, J.; Osorio, C.; Schieberle, P. Characterization of the key aroma compounds in pink guava (Psidium guajava L.) by means of aroma re-engineering experiments and omission tests. J. Agric. Food Chem. 2009, 57, 2882–2888. [Google Scholar] [CrossRef] [PubMed]
- Schuh, C.; Schieberle, P. Characterization of the Key Aroma Compounds in the Beverage Prepared from Darjeeling Black Tea: Quantitative Differences between Tea Leaves and Infusion. J. Agric. Food Chem. 2006, 54, 916–924. [Google Scholar] [CrossRef]
- Alasalvar, C.; Topal, B.; Serpen, A.; Bahar, B.; Pelvan, E.; Gökmen, V. Flavor Characteristics of Seven Grades of Black Tea Produced in Turkey. J. Agric. Food Chem. 2012, 60, 6323–6332. [Google Scholar] [CrossRef] [PubMed]
- Kotseridis, Y.; Baumes, R. Identification of Impact Odorants in Bordeaux Red Grape Juice, in the Commercial Yeast Used for Its Fermentation, and in the Produced Wine. J. Agric. Food Chem. 2000, 48, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.T.; Alonso, M.V.; Rios, J.J.; Aparicio, R. Virgin Olive Oil Aroma: Relationship between Volatile Compounds and Sensory Attributes by Chemometrics. J. Agric. Food Chem. 1995, 43, 2925–2931. [Google Scholar] [CrossRef]
- Angerosa, F.; Servili, M.; Selvaggini, R.; Taticchi, A.; Esposto, S.; Montedoro, G. Volatile compounds in virgin olive oil: Occurrence and their relationship with the quality. J. Chromatogr. A 2004, 1054, 17–31. [Google Scholar] [CrossRef]
- Conde, C.; Delrot, S.; Gerós, H. Physiological, biochemical and molecular changes occurring during olive development and ripening. J. Plant Physiol. 2008, 165, 1545–1562. [Google Scholar] [CrossRef]
- Olias, J.M.; Perez, A.G.; Rios, J.J.; Sanz, L.C. Aroma of virgin olive oil: Biogenesis of the “green” odor notes. J. Agric. Food Chem. 1993, 41, 2368–2373. [Google Scholar] [CrossRef]
- Padilla, M.N.; Hernández, M.L.; Sanz, C.; Martínez-Rivas, J.M. Functional characterization of two 13-lipoxygenase genes from olive fruit in relation to the biosynthesis of volatile compounds of virgin olive oil. J. Agric. Food Chem. 2009, 57, 9097–9107. [Google Scholar] [CrossRef]
- Lanciotti, R.; Gianotti, A.; Patrignani, F.; Belletti, N.; Guerzoni, M.E.; Gardini, F. Use of natural aroma compounds to improve shelf-life and safety of minimally processed fruits. Trends Food Sci. Technol. 2004, 15, 201–208. [Google Scholar] [CrossRef]
- Kubo, A.; Lunde, C.S.; Kubo, I. Antimicrobial activity of the olive oil flavor compounds. J. Agric. Food Chem. 1995, 43, 1629–1633. [Google Scholar] [CrossRef]
- Kubo, I.; Fujita, K.; Kubo, A.; Nihei, K.; Lunde, C.S. Modes of antifungal action of (2E)-alkenals against Saccharomyces cerevisiae. J. Agric. Food Chem. 2003, 51, 3951–3957. [Google Scholar] [CrossRef] [PubMed]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.A. Flavours: The Legal Framework. In Flavours and Fragrances: Chemistry, Bioprocessing and Sustainability; Berger, R.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 15–24. [Google Scholar]
- Schrader, J.; Etschmann, M.M.W.; Sell, D.; Hilmer, J.M.; Rabenhorst, J. Applied biocatalysis for the synthesis of natural flavour compounds—Current industrial processes and future prospects. Biotechnol. Lett. 2004, 26, 463–472. [Google Scholar] [CrossRef]
- Kanisawa, T.; Itoh, H. Method for Preparing Green Aroma Compounds. U.S. Patent 4769243A, 6 September 1988. [Google Scholar]
- Goers, S.K.; Ghossi, P.; Patterson, J.T.; Young, C.L. Process for Producing a Green Leaf Essence. Patent CA 1309615C, 3 November 1992. [Google Scholar]
- Sekiya, J.; Monma, T.; Kajiwara, T.; Hatanaka, A. Changes in Activities of Lipoxygenase and Hydroperoxide Lyase during Seed Development of Soybean. Agric. Biol. Chem. 1986, 50, 521–522. [Google Scholar]
- Trawatha, S.E.; TeKrony, D.M.; Hildebrand, D.F. Relationship of Soybean Seed Quality to Fatty Acid and C6-Aldehyde Levels during Storage. Crop Sci. 1995, 35, 1415–1422. [Google Scholar] [CrossRef]
- Whitehead, I.M.; Slusarenko, A.J.; Waspi, U.; Gaskin, D.J.H.; Brash, A.R.; Tijet, N. Guava (Psidium Guajava) 13-Hydroperoxide Lyase and Uses Thereof. U.S. Patent 6,780,621 B2, 24 August 2004. [Google Scholar]
- Fu, X.; Zhu, X.; Gao, K.; Duan, J. Oil and fat hydrolysis with lipase from Aspergillus sp. J. Am. Oil Chem. Soc. 1995, 72, 527–531. [Google Scholar] [CrossRef]
- Gargouri, M.; Akacha, N.B.; Kotti, F.; Ben Rejeb, I. Lipoxygenase pathway: Valorization of plant oils and aroma biosynthesis. Biotechnol. Agron. Société Environ. 2008, 12, 185–202. [Google Scholar]
- Kosugi, Y.; Tanaka, H.; Tomizuka, N. Continuous hydrolysis of oil by immobilized lipase in a countercurrent reactor. Biotechnol. Bioeng. 1990, 36, 617–622. [Google Scholar] [CrossRef]
- Xi, W.-W.; Xu, J.-H. Preparation of enantiopure (S)-ketoprofen by immobilized Candida rugosa lipase in packed bed reactor. Process Biochem. 2005, 40, 2161–2166. [Google Scholar] [CrossRef]
- Gargouri, M.; Legoy, M.D. Bienzymatic reaction for hydroperoxide production in a multiphasic system. Enzym. Microb. Technol. 1997, 21, 79–84. [Google Scholar] [CrossRef]
- Kim, K.H.; Kwon, D.Y.; Rhee, J.S. Effects of organic solvents on lipase for fat splitting. Lipids 1984, 19, 975–977. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Kawasaki, M.; Shiomori, K.; Baba, Y.; Hano, T. Hydrolysis kinetics of olive oil with lipase in a transfer cell. J. Ferment. Bioeng. 1994, 77, 283–287. [Google Scholar] [CrossRef]
- Yang, F.; Russell, A.J. A comparison of lipase-catalyzed ester hydrolysis in reverse micelles, organic solvents, and biphasic systems. Biotechnol. Bioeng. 1995, 47, 60–70. [Google Scholar] [CrossRef]
- H-Kittikun, A.; Prasertsan, P.; Sungpud, C. Continuous production of fatty acids from palm olein by immobilized lipase in a two-phase system. J. Am. Oil Chem. Soc. 2000, 77, 599–603. [Google Scholar] [CrossRef]
- Tiss, A.; Carrière, F.; Verger, R. Effects of Gum Arabic on Lipase Interfacial Binding and Activity. Anal. Biochem. 2001, 294, 36–43. [Google Scholar] [CrossRef]
- Mtibaa, H.; Fendri, A.; Sayari, A.; Ben Salah, A.; Mejdoub, H.; Gargouri, Y. La lipase de Candida rugosa: Caractérisation biochimique. Oléagineux Corps Gras Lipides 2002, 9, 43–47. [Google Scholar] [CrossRef][Green Version]
- Piazza, G.J. Lipoxygenase catalyzed hydroperoxide formation in microemulsions containing nonionic surfactant. Biotechnol. Lett. 1992, 14, 1153–1158. [Google Scholar] [CrossRef]
- Rodakiewicz-Nowak, J.; Maslakiewicz, P.; Haber, J. The effect of linoleic acid on pH inside sodium bis(2-ethylhexyl)sulfosuccinate reverse micelles in isooctane and on the enzymic activity of soybean lipoxygenase. Eur. J. Biochem. 1996, 238, 549–553. [Google Scholar] [CrossRef]
- Gargouri, M.; Drouet, P.; Hervagault, J.F.; Legoy, M.D. Investigation of behavior of an enzyme in a biphasic system: Soybean lipoxygenase-1. Biotechnol. Bioeng. 1996, 51, 573–580. [Google Scholar] [CrossRef]
- Kaewthong, W.; Sirisansaneeyakul, S.; Prasertsan, P.; H-Kittikun, A. Continuous production of monoacylglycerols by glycerolysis of palm olein with immobilized lipase. Process Biochem. 2005, 40, 1525–1530. [Google Scholar] [CrossRef]
- Gargouri, M.; Legoy, M.D. The kinetic behaviour of a two-enzyme system in biphasic media: Coupling hydrolysis and lipoxygenation. Biochim. Biophys. Acta 1997, 1337, 227–232. [Google Scholar] [CrossRef]
- Kermasha, S.; Dioum, N.; Bisakowski, B.; Vega, M. Biocatalysis by immobilized lipoxygenase in a ternary micellar system. J. Mol. Catal. B Enzym. 2002, 19–20, 305–317. [Google Scholar] [CrossRef]
- Karadag, H.; Bilgin, R.; Tukel, S. Immobilization of Soybean Lipoxygenase Onto Polyacrylamide Gel. Biotechnol. Biotechnol. Equip. 2006, 20, 95–100. [Google Scholar] [CrossRef][Green Version]
- Lakshmi, M.C.; Amrutha, N.; Tavanandi, H.A.; Raghavarao, K.S.M.S. Stabilization of Lipoxygenase-1 from Glycine max by Microencapsulation. Dry. Technol. 2015, 33, 493–501. [Google Scholar] [CrossRef]
- Christopher, J.; Axelrod, B. On the different positional specificities of peroxidation of linoleate shown by two isozymes of soybean lipoxygenase. Biochem. Biophys. Res. Commun. 1971, 44, 731–736. [Google Scholar] [CrossRef]
- Muller, B.; Gautier, A.; Dean, C.; Kuhn, J.C. Process for the Enzymatic Preparation of Aliphatic Alcohols and Aldehydes from Linoleic Acid, Linoleic Acid, or a Natural Precursor. U.S. Patent No. 5,464,761, 1995. [Google Scholar]
- Noordermeer, M.A.; Van Der Goot, W.; Van Kooij, A.J.; Veldsink, J.W.; Veldink, G.A.; Vliegenthart, J.F. Development of a biocatalytic process for the production of c6-aldehydes from vegetable oils by soybean lipoxygenase and recombinant hydroperoxide lyase. J. Agric. Food Chem. 2002, 50, 4270–4274. [Google Scholar] [CrossRef]
- Márczy, J.S.; Németh, Á.S.; Samu, Z.; Háger-Veress, Á.; Szajáni, B. Production of hexanal from hydrolyzed sunflower oil by lipoxygenase and hydroperoxide lyase enzymes. Biotechnol. Lett. 2002, 24, 1673–1675. [Google Scholar] [CrossRef]
- Nemeth, A.S.; Marczy, J.S.; Samu, Z.; Hager-Veress, A.; Szajani, B. Biocatalytic production of 2(E)-hexenal from hydrolysed linseed oil. Enzyme Microb. Technol. 2004, 34, 667–672. [Google Scholar] [CrossRef]
- Galliard, T.; Phillips, D.R. Lipoxygenase from potato tubers. Partial purification and properties of an enzyme that specifically oxygenates the 9-position of linoleic acid. Biochem. J. 1971, 124, 431–438. [Google Scholar] [CrossRef]
- Sekiya, J.; Aoshima, H.; Kajiwara, T.; Togo, T.; Hatanaka, A. Purification and Some Properties of Potato Tuber Lipoxygenase and Detection of Linoleic Acid Radical in the Enzyme Reaction. Agric. Biol. Chem. 1977, 41, 827–832. [Google Scholar]
- Gargouri, M.; Legoy, M. A two-enzyme system for the transformation of unsaturated oils to 9(S)-hydroperoxy fatty acids. Biotechnol. Lett. 2002, 24, 915–918. [Google Scholar] [CrossRef]
- Kerler, J.; Kohlen, E.; Fitz, W.; Vliet, A.V.D.; Winkel, C. Method for the Enzymatic Preparation of Aldehydes Rich Aromas c6-c10. U.S. Patent No. 6,864,072, 8 March 2005. [Google Scholar]
- Wang, Z. Hock Rearrangement. In Comprehensive Organic Name Reactions and Reagents; American Cancer Society: Atlanta, GA, USA, 2010; pp. 1438–1442. [Google Scholar]
- Brunerie, P. Procédé de synthèse du cis-3-hexene-1-ol à partir d’acide gras insaturé. Patent EP 0481147B1, 10 June 1998. [Google Scholar]
- Schade, F.; Thompson, J.; Legge, R. Use of a plant-derived enzyme template for the production of the green-note volatile hexanal. Biotechnol. Bioeng. 2003, 84, 265–273. [Google Scholar] [CrossRef]
- Akacha, N.B.; Boubaker, O.; Gargouri, M. Production of hexenol in a two-enzyme system: Kinetic study and modelling. Biotechnol. Lett. 2005, 27, 1875–1878. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, M.; Akacha, N.B.; Legoy, M.D. Coupled hydroperoxide lyase and alcohol dehydrogenase for selective synthesis of aldehyde or alcohol. Appl. Biochem. Biotechnol. 2004, 119, 171–180. [Google Scholar] [CrossRef]
- Akacha, N.B.; Gargouri, M. Enzymatic synthesis of green notes with hydroperoxide-lyase from olive leaves and alcohol-dehydrogenase from yeast in liquid/gas reactor. Process Biochem. 2009, 44, 1122–1127. [Google Scholar] [CrossRef]
- Hausler, A.; Ehret, C.; Binggeli, E. Process for the Production of Degradation Products of Fatty Acids. U.S. Patent No. 6150145, 21 November 2000. [Google Scholar]
- Tan, Y.; Siebert, K.J. Quantitative Structure−Activity Relationship Modeling of Alcohol, Ester, Aldehyde, and Ketone Flavor Thresholds in Beer from Molecular Features. J. Agric. Food Chem. 2004, 52, 3057–3064. [Google Scholar] [CrossRef]
- Brunerie, P.; Koziet, Y. Process for Producing Natural cis-3-Hexenol from Unsaturated Fatty Acids. U.S. Patent 5620879A, 15 April 1997. [Google Scholar]
- Hall, C.E.; Karboune, S.; Florence, H.; Kermasha, S. Stabilization of an enzymatic extract from Penicillium camemberti containing lipoxygenase and hydroperoxide lyase activities. Process Biochem. 2008, 43, 258–264. [Google Scholar] [CrossRef]
- Akacha, N.B.; Karboune, S.; Gargouri, M.; Kermasha, S. Activation and stabilization of the hydroperoxide lyase enzymatic extract from mint leaves (Mentha spicata) using selected chemical additives. Appl. Biochem. Biotechnol. 2010, 160, 901–911. [Google Scholar] [CrossRef]
- Long, Z.; Kong, X.; Zhang, C.; Hua, Y. Stability of hydroperoxide lyase activity from Amaranthus tricolor (Amaranthus mangostanus L.) leaves: Influence of selected additives. J. Sci. Food Agric. 2010, 90, 729–734. [Google Scholar]
- Hornostaj, A.R.; Robinson, D.S. Purification of hydroperoxide lyase from pea seeds. Food Chem. 2000, 71, 241–247. [Google Scholar] [CrossRef]
- Gianfreda, L.; Scarfi, M.R. Enzyme stabilization: State of the art. Mol. Cell. Biochem. 1991, 100, 97–128. [Google Scholar] [CrossRef] [PubMed]
- Koeduka, T.; Stumpe, M.; Matsui, K.; Kajiwara, T.; Feussner, I. Kinetics of barley FA hydroperoxide lyase are modulated by salts and detergents. Lipids 2003, 38, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Hua, Y.; Kong, X.; Zhang, C.; Chen, Y. Covalent immobilization of hydroperoxide lyase on chitosan hybrid hydrogels and production of C6 aldehydes by immobilized enzyme. J. Mol. Catal. B Enzym. 2013, 95, 89–98. [Google Scholar] [CrossRef]
- Liu, Q.; Hua, Y. Continuous synthesis of hexanal by immobilized hydroperoxide lyase in packed-bed reactor. Bioprocess Biosyst. Eng. 2015, 38, 2439–2449. [Google Scholar] [CrossRef]
- Gigot, C.; Ongena, M.; Fauconnier, M.-L.; Muhovski, Y.; Wathelet, J.-P.; du Jardin, P.; Thonart, P. Optimization and scaling up of a biotechnological synthesis of natural green leaf volatiles using Beta vulgaris hydroperoxide lyase. Process Biochem. 2012, 47, 2547–2551. [Google Scholar] [CrossRef]
- Jacopini, S.; Vincenti, S.; Mariani, M.; Brunini-Bronzini de Caraffa, V.; Gambotti, C.; Desjobert, J.M.; Muselli, A.; Costa, J.; Tomi, F.; Berti, L.; et al. Activation and Stabilization of Olive Recombinant 13-Hydroperoxide Lyase Using Selected Additives. Appl. Biochem. Biotechnol. 2017, 182, 1000–1013. [Google Scholar] [CrossRef]
- Brühlmann, F.; Bosijokovic, B.; Ullmann, C.; Auffray, P.; Fourage, L.; Wahler, D. Directed evolution of a 13-hydroperoxide lyase (CYP74B) for improved process performance. J. Biotechnol. 2013, 163, 339–345. [Google Scholar] [CrossRef]
- Brühlmann, F.; Bosijokovic, B. Efficient Biochemical Cascade for Accessing Green Leaf Alcohols. Org. Process Res. Dev. 2016, 20, 1974–1978. [Google Scholar] [CrossRef]
- Bourel, G.; Nicaud, J.-M.; Nthangeni, B.; Santiago-Gomez, P.; Belin, J.-M.; Husson, F. Fatty acid hydroperoxide lyase of green bell pepper: Cloning in Yarrowia lipolytica and biogenesis of volatile aldehydes. Enzym. Microb. Technol. 2004, 35, 293–299. [Google Scholar] [CrossRef]
- Santiago-Gomez, M.P.; Thanh, H.T.; De Coninck, J.L.; Cachon, R.M.; Kermasha, S.L.; Belin, J.-M.; Gervais, P.; Husson, F. Modeling hexanal production in oxido-reducing conditions by the yeast Yarrowia lipolytica. Process Biochem. 2009, 44, 1013–1018. [Google Scholar] [CrossRef]
- Atwal, A.S.; Bisakowski, B.; Richard, S.; Robert, N.; Lee, B. Cloning and secretion of tomato hydroperoxide lyase in Pichia pastoris. Process Biochem. 2005, 40, 95–102. [Google Scholar] [CrossRef]
- Buchhaupt, M.; Guder, J.C.; Etschmann, M.M.W.; Schrader, J. Synthesis of green note aroma compounds by biotransformation of fatty acids using yeast cells coexpressing lipoxygenase and hydroperoxide lyase. Appl. Microbiol. Biotechnol. 2012, 93, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C.; Studart-Witkowski, C.; Schwab, W. Overexpression of hydroperoxide lyase gene in Nicotiana benthamiana using a viral vector system. Plant Biotechnol. J. 2010, 8, 783–795. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).