
Structural Characterization of Pectic Polysaccharides in the Cell Wall of Stevens Variety Cranberry Using Highly Specific Pectin-Hydrolyzing Enzymes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Alcohol Insoluble Solids
2.3. Sequential Extraction of Polysaccharides
2.4. Fractionation of Polysaccharides by Anion Exchange Chromatography
2.5. Purification of Polysaccharides by Gel Filtration Chromatography
2.6. Enzymatic Fragmentation of Selected Polysaccharides
2.7. Sugar Content and Monosaccharide Profile
2.8. Molecular Weight Distribution Analysis
2.9. Preparation of Methylated PMP-Monosaccharides
2.10. Sugar Linkage Analysis by Liquid Chromatography-Mass Spectrometry
3. Results and Discussion
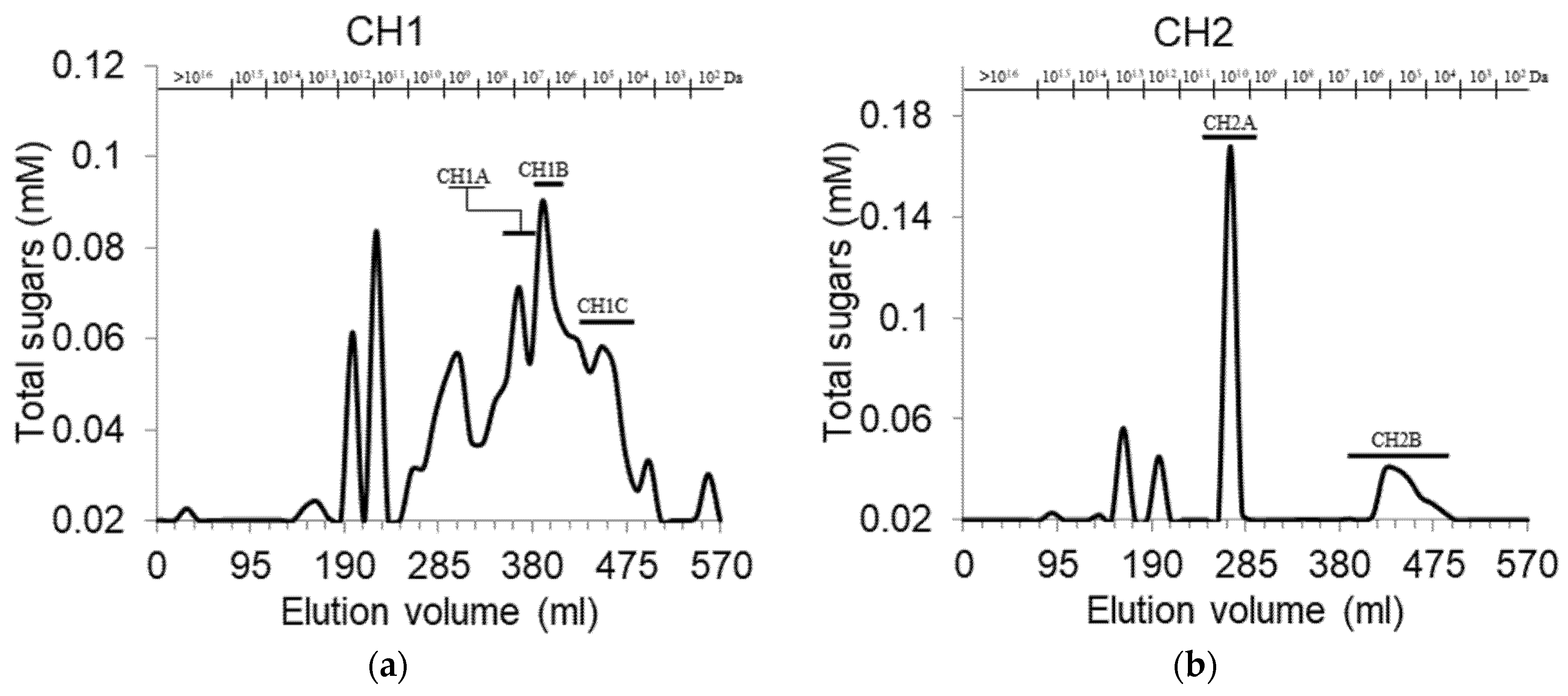
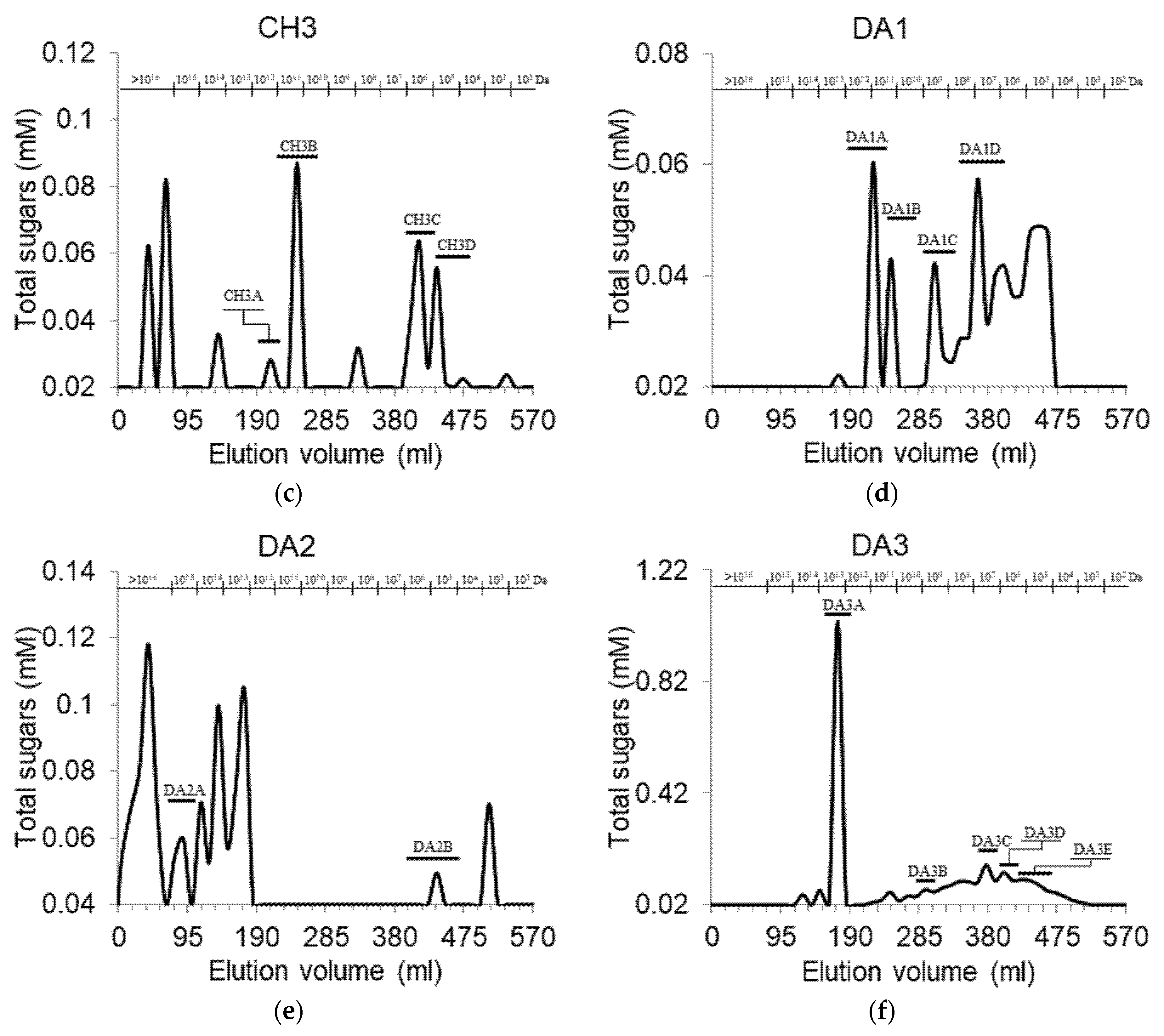
3.1. Fractionation and Purification of Cell Wall Pectic Polysaccharides Fractions
3.2. Monosaccharide Profile of Purified Pectic Polysaccharides
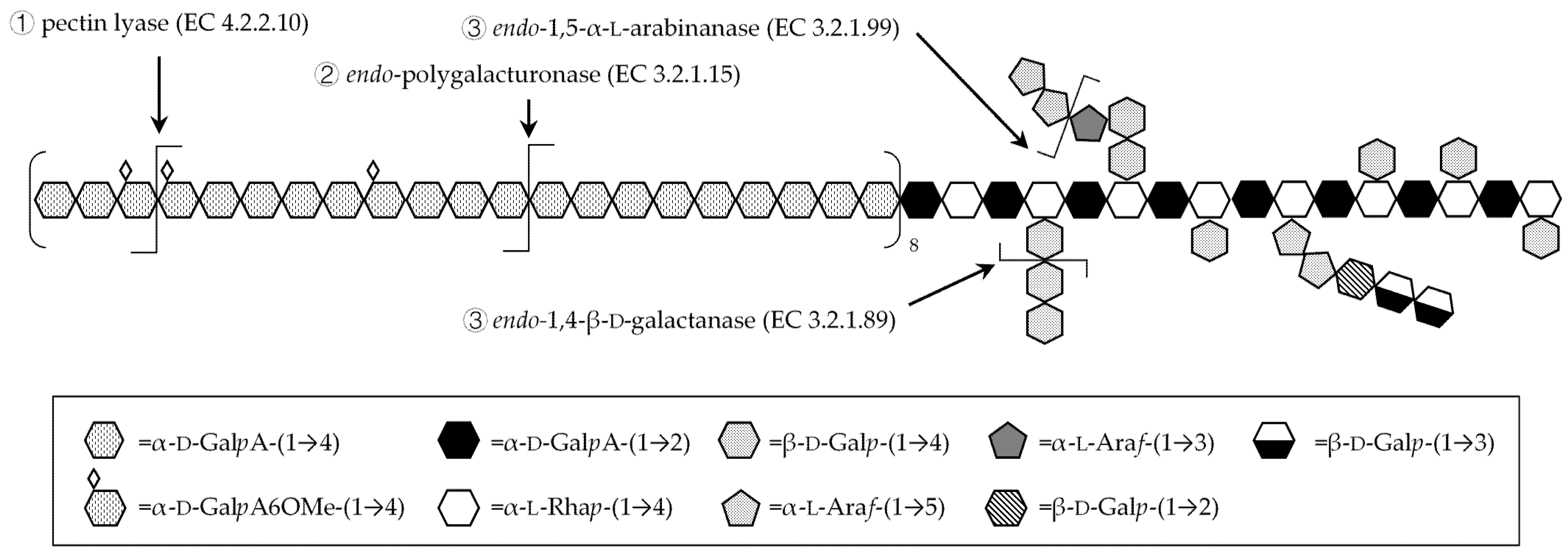
3.3. Fragmentation with Homogalacturonan-Degrading Enzymes
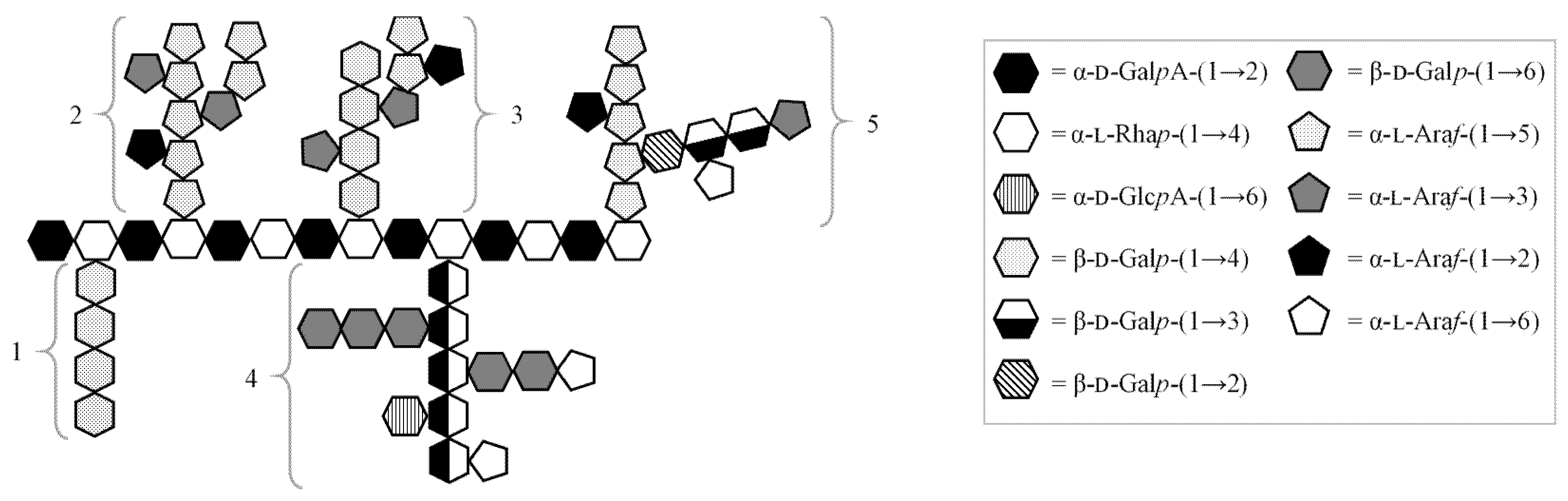
3.4. Fragmentation with Debranching Glycosyl-Hydrolase Enzymes
3.5. Glycoside Linkage Analysis of Polysaccharide Fragments Recovered upon Debranching
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kratchanova, M.; Nikolova, M.; Pavlova, E.; Yanakieva, I.; Kussovski, V. Composition and properties of biologically active pectic polysaccharides from leek (Allium porrum). J. Sci. Food Agric. 2010, 90, 2046–2051. [Google Scholar] [CrossRef]
- Oosterveld, A.; Beldman, G.; Schols, H.A.; Voragen, A.G. Arabinose and ferulic acid rich pectic polysaccharides extracted from sugar beet pulp. Carbohydr. Res. 1996, 288, 143–153. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Bonnin, E.; Garnier, C.; Ralet, M.-C. Pectin-modifying enzymes and pectin-derived materials: Applications and impacts. Appl. Microbiol. Biotechnol. 2013, 98, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Yapo, B.M. Rhamnogalacturonan-I: A Structurally Puzzling and Functionally Versatile Polysaccharide from Plant Cell Walls and Mucilages. Polym. Rev. 2011, 51, 391–413. [Google Scholar] [CrossRef]
- Peña, M.J.; Carpita, N.C. Loss of highly branched arabinans and debranching of rhamnogalacturonan i accompany loss of firm texture and cell separation during prolonged storage of apple. Plant Physiol. 2004, 135, 1305–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schols, H.A.; Bakx, E.J.; Schipper, D.; Voragen, A.G. A xylogalacturonan subunit present in the modified hairy regions of apple pectin. Carbohydr. Res. 1995, 279, 265–279. [Google Scholar] [CrossRef]
- Oechslin, R.; Lutz, M.V.; Amadò, R. Pectic substances isolated from apple cellulosic residue: Structural characterisation of a new type of rhamnogalacturonan I. Carbohydr. Polym. 2003, 51, 301–310. [Google Scholar] [CrossRef]
- Sakamoto, T.; Sakai, T. Analysis of structure of sugar-beet pectin by enzymatic methods. Phytochemistry 1995, 39, 821–823. [Google Scholar] [CrossRef]
- Yu, L.; Yu, C.; Zhu, M.; Cao, Y.; Yang, H.; Zhang, X.; Ma, Y.; Zhou, G. Structural analysis of galactoarabinan from duckweed. Carbohydr. Polym. 2015, 117, 807–812. [Google Scholar] [CrossRef]
- Vidal, S.; Doco, T.; Williams, P.; Pellerin, P.; York, W.S.; O’Neill, M.A.; Glushka, J.; Darvill, A.G.; Albersheim, P. Structural characterization of the pectic polysaccharide rhamnogalacturonan II: Evidence for the backbone location of the aceric acid-containing oligoglycosyl side chain. Carbohydr. Res. 2000, 326, 277–294. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.; Seymour, G.B. Fruit Softening: Revisiting the Role of Pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Popper, Z.A.; Fry, S.C. Xyloglucan−pectin linkages are formed intra-protoplasmically, contribute to wall-assembly, and remain stable in the cell wall. Planta 2008, 227, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Theuwissen, E.; Mensink, R.P. Water-soluble dietary fibers and cardiovascular disease. Physiol. Behav. 2008, 94, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Spadoni Andreani, E.; Li, M.; Ronholm, J.; Karboune, S. Feruloylation of polysaccharides from cranberry and characterization of their prebiotic properties. Food Biosci. 2021, 101071. [Google Scholar] [CrossRef]
- Olano-Martin, E.; Gibson, G.; Rastall, R. Comparison of the in vitro bifidogenic properties of pectins and pectic-oligosaccharides. J. Appl. Microbiol. 2002, 93, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Manderson, K.; Pinart, M.; Tuohy, K.; Grace, W.E.; Hotchkiss, A.T.; Widmer, W.; Yadhav, M.P.; Gibson, G.R.; Rastall, R.A. In Vitro Determination of Prebiotic Properties of Oligosaccharides Derived from an Orange Juice Manufacturing By-Product Stream. Appl. Environ. Microbiol. 2005, 71, 8383–8389. [Google Scholar] [CrossRef] [Green Version]
- Li, P.-J.; Xia, J.-L.; Nie, Z.-Y.; Shan, Y. Pectic oligosaccharides hydrolyzed from orange peel by fungal multi-enzyme complexes and their prebiotic and antibacterial potentials. LWT 2016, 69, 203–210. [Google Scholar] [CrossRef]
- Li, S.; Li, T.; Zhu, R.; Wang, N.; Song, Y.; Wang, S.; Guo, M. Antibacterial Action of Haw Pectic Oligosaccharides. Int. J. Food Prop. 2013, 16, 706–712. [Google Scholar] [CrossRef]
- Sun, J.; Marais, J.P.; Khoo, C.; LaPlante, K.; Vejborg, R.M.; Givskov, M.; Tolker-Nielsen, T.; Seeram, N.P.; Rowley, D.C. Cranberry (Vaccinium macrocarpon) oligosaccharides decrease biofilm formation by uropathogenic Escherichia coli. J. Funct. Foods 2015, 17, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotchkiss, J.A.T.; Nuñez, A.; Strahan, G.D.; Chau, H.K.; White, A.K.; Marais, J.P.J.; Hom, K.; Vakkalanka, M.S.; Di, R.; Yam, K.L.; et al. Cranberry Xyloglucan Structure and Inhibition of Escherichia coli Adhesion to Epithelial Cells. J. Agric. Food Chem. 2015, 63, 5622–5633. [Google Scholar] [CrossRef]
- Spadoni Andreani, E.; Karboune, S.; Liu, L. Extraction and characterization of Cell Wall Polysaccharides from cranberry (Vaccinium macrocarpon var. Stevens) pomace. Carbohydr. Polym. 2021, 267, 118212. [Google Scholar] [CrossRef]
- Holck, J.; Hjernø, K.; Lorentzen, A.; Vigsnæs, L.K.; Hemmingsen, L.; Licht, T.R.; Mikkelsen, J.D.; Meyer, A.S. Tailored enzymatic production of oligosaccharides from sugar beet pectin and evidence of differential effects of a single DP chain length difference on human faecal microbiota composition after in vitro fermentation. Process. Biochem. 2011, 46, 1039–1049. [Google Scholar] [CrossRef]
- Hilz, H.; Bakx, E.J.; Schols, H.A.; Voragen, A.G. Cell wall polysaccharides in black currants and bilberries—Characterisation in berries, juice, and press cake. Carbohydr. Polym. 2005, 59, 477–488. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Khodaei, N.; Karboune, S. Extraction and structural characterisation of rhamnogalacturonan I-type pectic polysaccharides from potato cell wall. Food Chem. 2013, 139, 617–623. [Google Scholar] [CrossRef]
- Huisman, M.; Schols, H.; Voragen, A. Cell wall polysaccharides from soybean (Glycine max.) meal. Isolation and characterisation. Carbohydr. Polym. 1998, 37, 87–95. [Google Scholar] [CrossRef]
- Saulnier, L.; Thibault, J.-F. Extraction and characterization of pectic substances from pulp of grape berries. Carbohydr. Polym. 1987, 7, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Baron, A.; Missang, C.E.; Renard, C.M.; Drilleau, J. Cell wall polysaccharides of bush butter (Dacryodes edulis (G Don) HJ Lam) fruit pulp and their evolution during ripening. J. Sci. Food Agric. 2001, 81, 773–780. [Google Scholar] [CrossRef]
- Ding, X.; Zhu, F.; Gao, S. Purification, antitumour and immunomodulatory activity of water-extractable and alkali-extractable polysaccharides from Solanum nigrum L. Food Chem. 2012, 131, 677–684. [Google Scholar] [CrossRef]
- Guo, Q.; Du, J.; Jiang, Y.; Goff, H.D.; Cui, S.W. Pectic polysaccharides from hawthorn: Physicochemical and partial structural characterization. Food Hydrocoll. 2019, 90, 146–153. [Google Scholar] [CrossRef]
- Yang, J.-S.; Mu, T.-H.; Ma, M.-M. Optimization of ultrasound-microwave assisted acid extraction of pectin from potato pulp by response surface methodology and its characterization. Food Chem. 2019, 289, 351–359. [Google Scholar] [CrossRef]
- Ross, K.A.; Godfrey, D.; Fukumoto, L. The chemical composition, antioxidant activity and α-glucosidase inhibitory activity of water-extractable polysaccharide conjugates from northern Manitoba lingonberry. Cogent Food Agric. 2015, 1, 1109781. [Google Scholar] [CrossRef]
- Zykwinska, A.W.; Ralet, M.-C.J.; Garnier, C.D.; Thibault, J.-F.J. Evidence for In Vitro Binding of Pectin Side Chains to Cellulose. Plant Physiol. 2005, 139, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Broxterman, S.E.; Schols, H.A. Interactions between pectin and cellulose in primary plant cell walls. Carbohydr. Polym. 2018, 192, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S. Enzymatic extraction and linkage analysis of pectic polysaccharides from onion. Phytochemistry 1982, 21, 778–780. [Google Scholar] [CrossRef]
- Leivas, C.L.; Iacomini, M.; Cordeiro, L.M. Pectic type II arabinogalactans from starfruit (Averrhoa carambola L.). Food Chem. 2016, 199, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Prabasari, I.; Pettolino, F.; Liao, M.-L.; Bacic, A. Pectic polysaccharides from mature orange (Citrus sinensis) fruit albedo cell walls: Sequential extraction and chemical characterization. Carbohydr. Polym. 2011, 84, 484–494. [Google Scholar] [CrossRef]
- Deng, J.; Shi, Z.-J.; Li, X.-Z.; Liu, H.-M. Soluble polysaccharides isolation and characterization from rabbiteye blueberry (Vaccinium ashei) fruits. Bioresoure 2012, 8, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Øbro, J.; Harholt, J.; Scheller, H.V.; Orfila, C. Rhamnogalacturonan I in Solanum tuberosum tubers contains complex arabinogalactan structures. Phytochemistry 2004, 65, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Van De Vis, J.; Leeuwen, M.S.-V.; Siliha, H.; Kormelink, F.; Voragen, A. Purification and characterization of Endo-1,4-β-D-galactanases from Aspergillus niger and Aspergillus aculeatus: Use in combination with arabinanases from Aspergillus niger in enzymic conversion of potato arabinogalactan. Carbohydr. Polym. 1991, 16, 167–187. [Google Scholar] [CrossRef]
- Srivastava, L.M. Cell Wall, Cell Division, and Cell Growth. In Plant Growth and Development; Elsevier: Amsterdam, The Netherlands, 2002; pp. 23–74. [Google Scholar]
- Ralet, M.-C.; Guillon, F.; Renard, C.; Thibault, J.-F. Sugar Beet Fiber. In Fiber Ingredients; CRC Press: Boca Raton, FL, USA, 2009; pp. 359–392. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Ionic Strength (M) | Total Sugars (% mol/mol in Whole Extract) | Uronic Acid (% mol) |
|---|---|---|---|
| CH1 1 | 0.31 | 33.5 ± 2.9 | 99.0 ± 0.9 a |
| CH2 1 | 0.62 | 31.5 ± 3.0 | 91.6 ± 5.2 a |
| CH3 1 | 0.97 | 27.5 ± 5.8 | 96.1 ± 4.3 a |
| DA1 2 | 0.52 | 24.2 ± 3.7 | 52.9 ± 9.9 b |
| DA2 2 | 0.81 | 28.3 ± 4.4 | 54.6 ± 11.8 b |
| DA3 2 | 1.08 | 15.4 ± 2.2 | 85.6 ± 10.9 a |
| Gel Filtration Fraction | Molecular Weight (Da) | Monosaccharide Profile (% mol/mol) | RGI Branching c | ||
|---|---|---|---|---|---|
| Uronic Acids | Pectic Neutral Sugars a | Hemicellulose Neutral Sugars b | |||
| CH1A | 6.54·107 | 88.1 ± 6.0 d,e | 8.0 ± 1.6 e,f | 3.8 ± 0.1 h | 3.4 |
| CH1B | 1.39·107 | 93.3 ± 4.4 d | 3.0 ± 0.5 g | 3.7 ± 0.8 g,h | 0.6 |
| CH1C | 2.92·105 | 74.1 ± 0.7 d,e | 18.6 ± 3.2 d | 7.2 ± 0.7 f | 2.8 |
| CH2A | >109 | 75.2 ± 11.8 d,e | 18.8 ± 2.4 d | 6.0 ± 3.0 e–h | 4.7 |
| CH2B | 6.32·105 | 91.3 ± 2.7 d,e | 5.2 ± 0.4 f | 3.5 ± 1.5 g,h | 6.4 |
| CH3A | >109 | 90.0 ± 3.8 d,e | 5.7 ± 1.9 f | 4.3 ± 1.9 f–h | 3.4 |
| CH3B | >109 | 76.8 ± 10.7 d,e | 13.8 ± 3.5 d | 9.4 ± 2.4 d–f | 1.2 |
| CH3C | 2.97·106 | 83.6 ± 6.2 d,e | 10.1 ± 1.3 e | 6.2 ± 2.4 f–h | 3.6 |
| CH3D | 6.32·105 | 70.7 ± 11.5 e | 16.4 ± 3.5 d | 12.9 ± 2.1 d | 2.2 |
| DA2A | >109 | 89.1 ± 8.6 d,e | 3.9 ± 1.4 f,g | 7.0 ± 1.3 f | 5.5 |
| DA3A | >109 | 86.0 ± 8.6 d,e | 5.1 ± 0.5 f | 8.8 ± 0.3 e | 1.6 |
| DA3C | 3.02·107 | 88.1 ± 8.2 d,e | 8.5 ± 2.8 e,f | 3.4 ± 0.5 h | 1.7 |
| DA3D | 6.43·106 | 71.7 ± 10.2 d,e | 23.0 ± 8.1 d | 5.3 ± 1.3 f,g | 1.4 |
| DA3E | 6.32·105 | 81.2 ± 7.5 d,e | 13.3 ± 3.4 d,e | 5.5 ± 2.4 f–h | 1.7 |
| Endo-α-1,5-arabinanase | Endo-β-1,4-galactanase | Endo-α-1,5-arabinanase + Endo-β-1,4-galactanase | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rhamnose | Arabinose | Galactose | Rhamnose | Arabinose | Galactose | Rhamnose | Arabinose | Galactose | |
| CH1A | n.d. | n.d. | n.d. | 4.9 ± 0.5 c–e | 4.4 ± 0.5 b–d | 3.2 ± 0.6 b–e | 16.0 ± 2.3 b | 5.6 ± 1.1 b,c | 2.9 ± 0.2 b,c |
| CH1B | 10.6 ± 0.5 b,c | n.d. | 3.2 ± 0.6 b | 1.5 ± 0.1 e | 3.6 ± 0.6 b–d | 5.8 ± 0.3 a,b | 6.9 ± 0.8 c–e | n.d. | n.d. |
| CH1C | 1.9 ± 0.1 e | 0.1 ± 0.0 c | 0.1 ± 0.0 d | 2.8 ± 0.4 d,e | n.d. | 0.1 ± 0.0 f | 2.1 ± 0.5 e | 0.1 ± 0.0 c | 0.1 ± 0.0 d |
| CH2A | 5.8 ± 0.5 c–e | n.d. | n.d. | 15.6 ± 2.5 a,b | 0.6 ± 0.1 d | 0.3 ± 0.0 f | 5.0 ± 1.4 d,e | 0.4 ± 0.1 b,c | 0.5 ± 0.1 c,d |
| CH2B | n.d. | n.d. | n.d. | 1.0 ± 0.2 e | 2.4 ± 0.2 c,d | 1.7 ± 0.3 d–f | 2.4 ± 0.3 e | 0.1 ± 0.0 c | 0.0 ± 0.0 d |
| CH3A | 2.9 ± 0.4 d,e | 0.2 ± 0.0 c | 0.1 ± 0.0 d | 10.0 ± 0.5 b,c | 15.5 ± 3.4 a | 1.2 ± 0.1 d–f | 6.0 ± 0.4 c–e | 0.3 ± 0.0 b,c | 0.4 ± 0.1 d |
| CH3B | 29.5 ± 3.2 a | 0.3 ± 0.1 c | 0.9 ± 0.2 c,d | 10.7 ± 2.2 b,c | 0.7 ± 0.2 d | 0.6 ± 0.0 e,f | 4.0 ± 0.3 d,e | 0.4 ± 0.1 b,c | 0.5 ± 0.0 d |
| CH3C | 0.5 ± 0.1 e | 0.3 ± 0.0 c | 0.1 ± 0.0 d | 4.5 ± 0.7 c–e | 0.5 ± 0.1 d | 0.7 ± 0.0 e,f | 5.0 ± 0.3 d,e | 0.2 ± 0.1 c | 0.3 ± 0.1 d |
| CH3D | n.d. | n.d. | n.d. | 2.1 ± 0.3 e | 0.3 ± 0.0 d | n.d. | 10.6 ± 2.1 b–d | 0.5 ± 0.0 b,c | 0.9 ± 0.1 c,d |
| DA2A | 8.1 ± 1.3 b–d | 0.2 ± 0.0 c | 0.3 ± 0.0 d | n.d. | n.d. | 0.2 ± 0.0 f | 2.6 ± 0.7 e | 0.2 ± 0.0 c | 0.1 ± 0.0 d |
| DA3A | 1.0 ± 0.1 e | 1.7 ± 0.3 b | 0.8 ± 0.2 c,d | 8.9 ± 0.6 b–d | 18.0 ± 0.5 a | 2.8 ± 0.3 c–f | 4.1 ± 0.4 d,e | 6.0 ± 0.9 b | 7.2 ± 1.3 a |
| DA3C | n.d. | n.d. | n.d. | 20.7 ± 1.8 a | 3.2 ± 0.4 b–d | 4.7 ± 0.1 b,c | 12.5 ± 2.3 b,c | 2.7 ± 0.4 b,c | 2.5 ± 0.4 b,c |
| DA3D | 9.8 ± 1.7 b,c | 5.0 ± 0.3 b | 2.1 ± 0.2 b,c | 4.8 ± 0.3 c–e | 7.7 ± 1.1 b,c | 3.6 ± 0.1 b–d | 14.6 ± 0.8 b | 17.8 ± 3.5 a | 3.2 ± 0.2 b |
| DA3E | 12.7 ± 1.1 b | 10.8 ± 2.3 a | 6.4 ± 0.7 a | 18.1 ± 1.9 a | 8.6 ± 1.7 b | 8.2 ± 1.7 a | 24.0 ± 1.4 a | 4.9 ± 0.3 b,c | 0.8 ± 0.2 c,d |
| m/z | Linkage Pattern | CH1B | DA3A | DA3E | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| e-Ara | e-Gal | Both | e-Ara | e-Gal | Both | e-Ara | e-Gal | Both | ||
| 523.3 | T-Ara | 0.8 | 0.0 | 1.1 | 1.0 | 7.5 | 2.2 | 0.8 | 2.4 | 0.3 |
| 509.2 | 1,n-Ara | 5.6 | 22.2 | 4.0 | 2.9 | 19.0 | 8.2 | 0.0 | 0.0 | 2.8 |
| 495.2 | 1,3,n-Ara | 6.4 | 3.6 | 4.5 | 3.4 | 0.0 | 8.4 | 17.9 | 20.9 | 6.0 |
| 481.2 | 1,2,3,5-Ara | 19.5 | 50.4 | 55.7 | 54.0 | 7.6 | 26.6 | 34.6 | 57.3 | 55.7 |
| Total Arabinose | 32.3 | 76.2 | 65.4 | 61.2 | 34.1 | 45.4 | 53.2 | 80.6 | 64.8 | |
| 567.3 | T-Gal | 0.9 | 0.0 | 0.4 | 0.8 | 8.5 | 1.3 | 1.6 | 0.0 | 0.0 |
| 553.3 | 1,n-Gal | 11.0 | 0.0 | 9.4 | 13.9 | 26.1 | 9.3 | 2.3 | 0.0 | 10.8 |
| 539.3 | 1,n,n-Gal | 19.6 | 0.0 | 8.4 | 8.1 | 0.0 | 13.5 | 6.6 | 0.0 | 8.2 |
| 525.2 | 1,3,4,6-Gal | 19.0 | 0.0 | 6.5 | 6.3 | 0.0 | 7.7 | 0.0 | 0.0 | 7.6 |
| 525.2 | 1,2,4,n-Gal | 9.6 | 9.6 | 2.7 | 3.8 | 0.0 | 8.7 | 32.8 | 5.6 | 4.4 |
| Total Galactose | 60.0 | 9.6 | 27.3 | 33.0 | 34.7 | 40.5 | 43.4 | 5.6 | 30.9 | |
| 523.3 | 1,2-Rha | 0.8 | 0.0 | 1.1 | 1.0 | 7.5 | 2.2 | 0.8 | 2.4 | 0.3 |
| 509.2 | 1,n,n-Rha | 5.6 | 8.0 | 4.3 | 2.9 | 19.0 | 8.2 | 0.0 | 0.0 | 2.8 |
| Total Rhamnose | 6.4 | 8.0 | 5.5 | 3.9 | 26.5 | 10.4 | 0.8 | 2.4 | 3.1 | |
| 567.3 | T-Glc | 0.6 | 4.8 | 1.2 | 0.6 | 4.7 | 2.5 | 2.7 | 9.7 | 0.6 |
| 495.2 | 1,3,4-Xyl | 0.0 | 0.0 | 0.0 | 0.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 567.2 | 1,n-GalA | 0.7 | 1.4 | 0.7 | 0.8 | 0.0 | 1.1 | 0.0 | 1.7 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spadoni Andreani, E.; Karboune, S.; Liu, L. Structural Characterization of Pectic Polysaccharides in the Cell Wall of Stevens Variety Cranberry Using Highly Specific Pectin-Hydrolyzing Enzymes. Polymers 2021, 13, 1842. https://doi.org/10.3390/polym13111842
Spadoni Andreani E, Karboune S, Liu L. Structural Characterization of Pectic Polysaccharides in the Cell Wall of Stevens Variety Cranberry Using Highly Specific Pectin-Hydrolyzing Enzymes. Polymers. 2021; 13(11):1842. https://doi.org/10.3390/polym13111842
Chicago/Turabian StyleSpadoni Andreani, Eugenio, Salwa Karboune, and Lan Liu. 2021. "Structural Characterization of Pectic Polysaccharides in the Cell Wall of Stevens Variety Cranberry Using Highly Specific Pectin-Hydrolyzing Enzymes" Polymers 13, no. 11: 1842. https://doi.org/10.3390/polym13111842