


Poly(lactic acid) Degradation by Recombinant Cutinases from Aspergillus nidulans


Abstract
1. Introduction
2. Materials and Methods
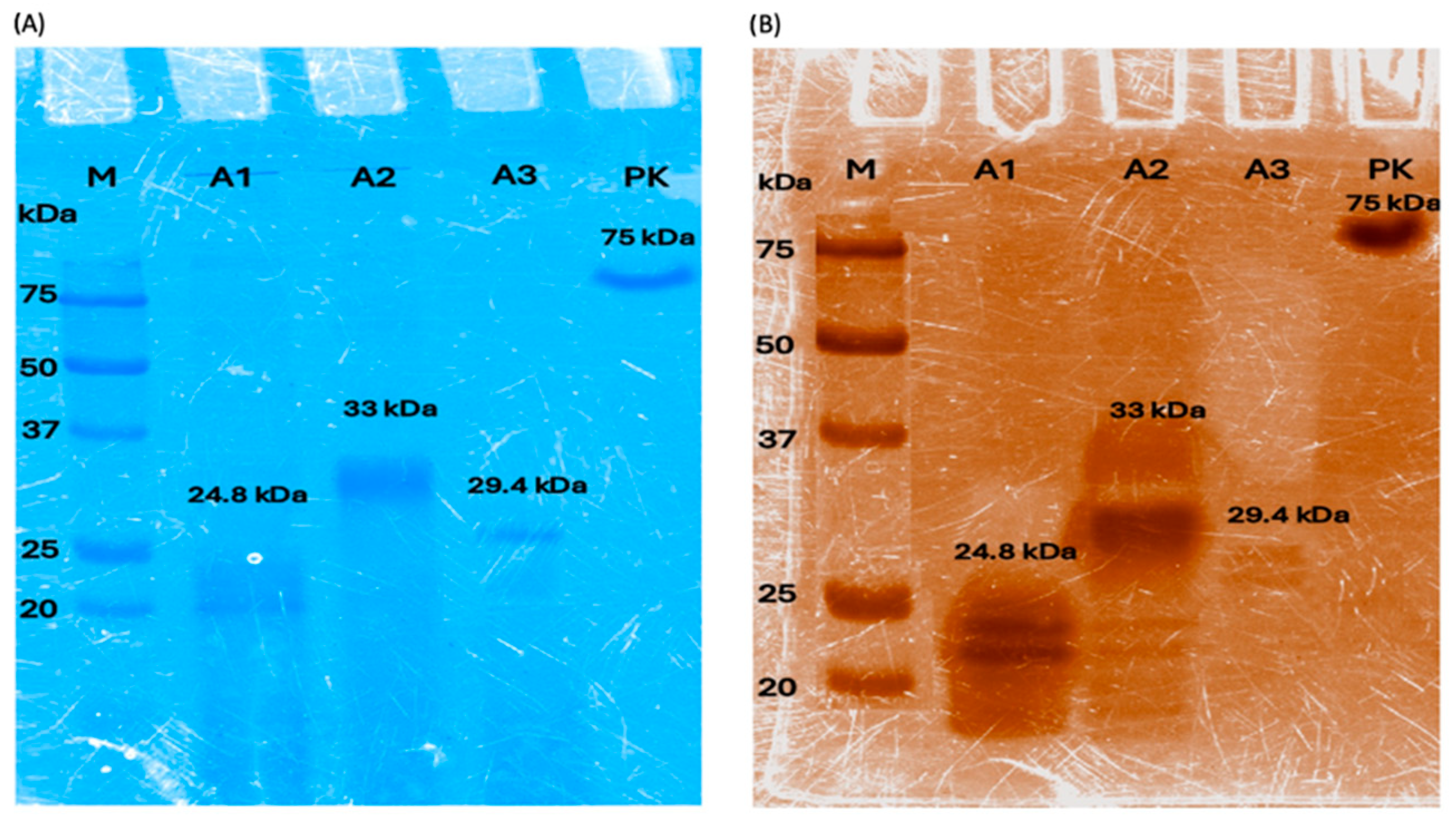
2.1. Recombinant Enzyme Production
2.2. Poly L-Lactic Acid (PLLA) Hydrolysis Reaction with Recombinant Cutinases
2.3. PLLA Hydrolysis with Proteinase K
2.4. Assay of Lactic Acid after Enzyme Hydrolysis of PLLA
2.5. Assay of Lactic Acid after Enzyme Hydrolysis of PDLA
2.6. Weight Loss
2.7. Morphological Changes Detected by Scanning Electronic Microscopy (SEM)
3. Results
3.1. Enzyme Production and Activity Assay
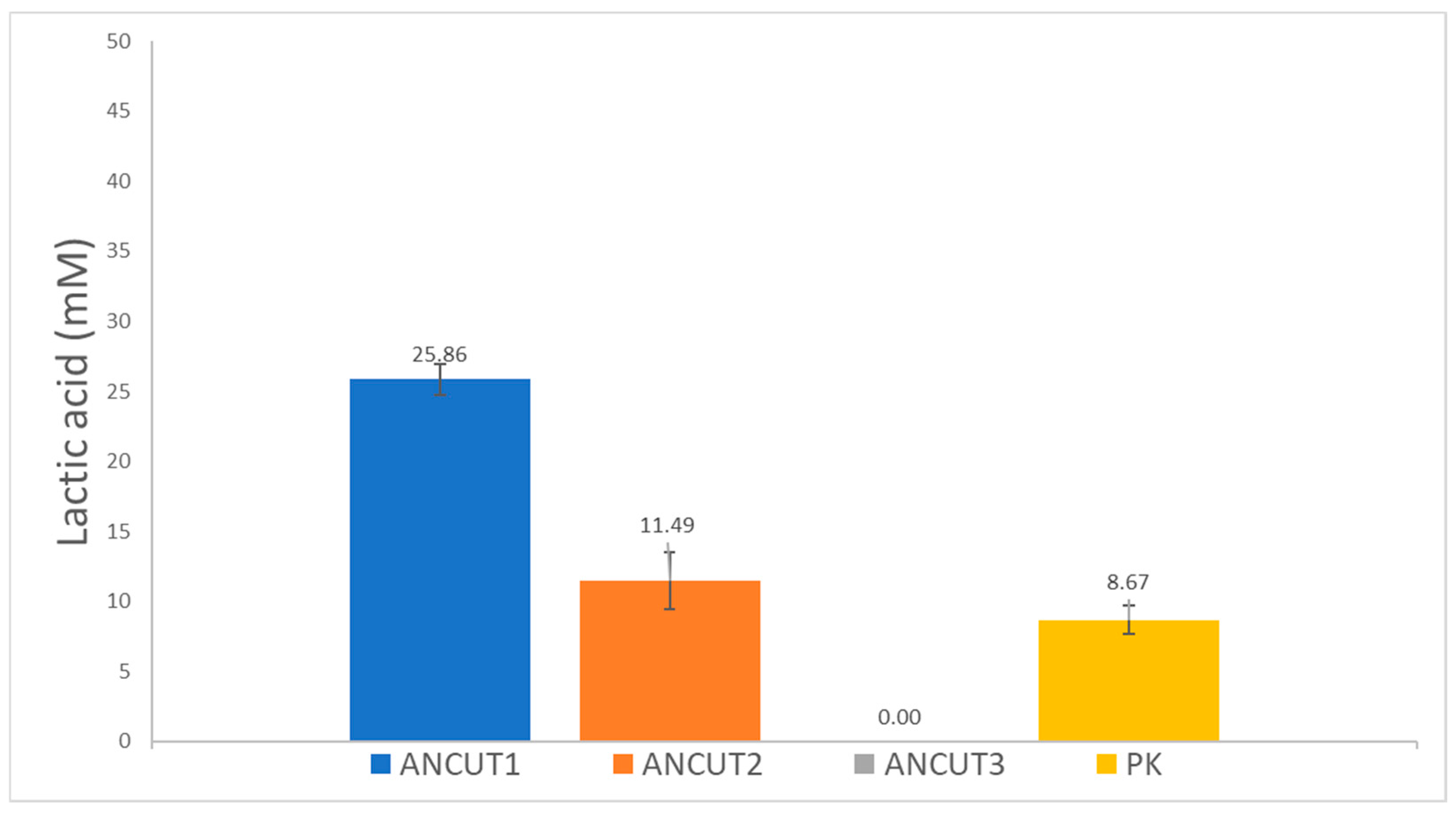
3.2. PLLA Degradation Assayed through L-Lactic Acid Release
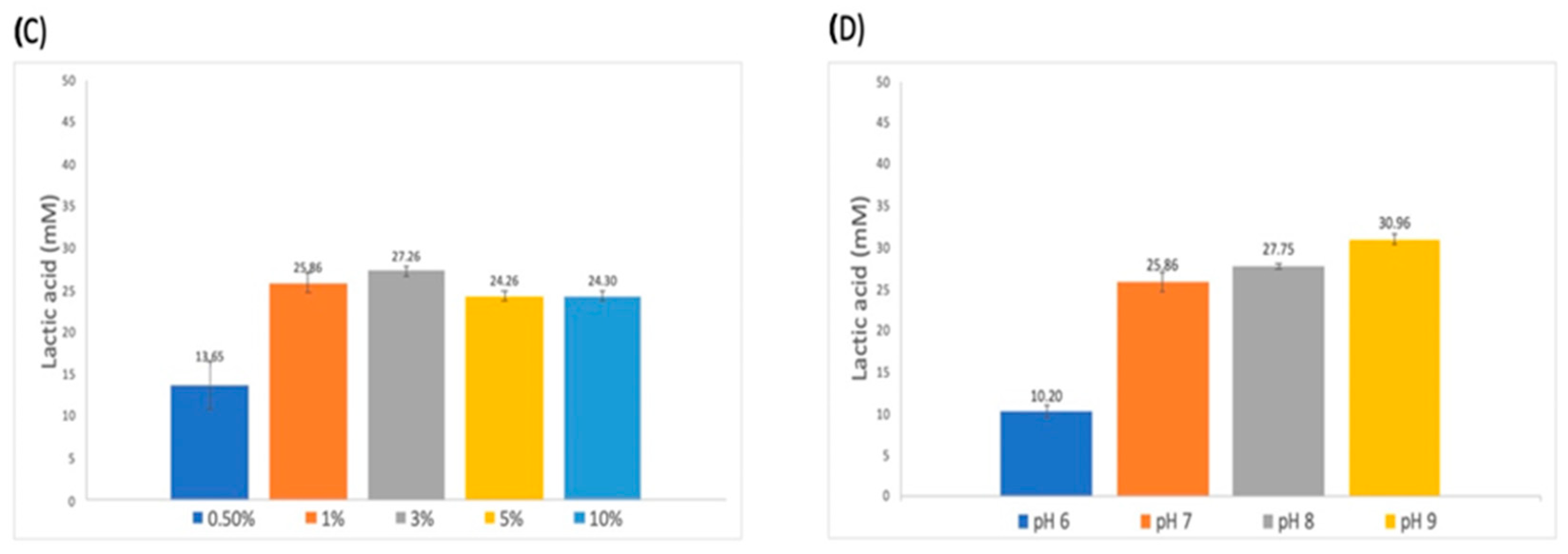
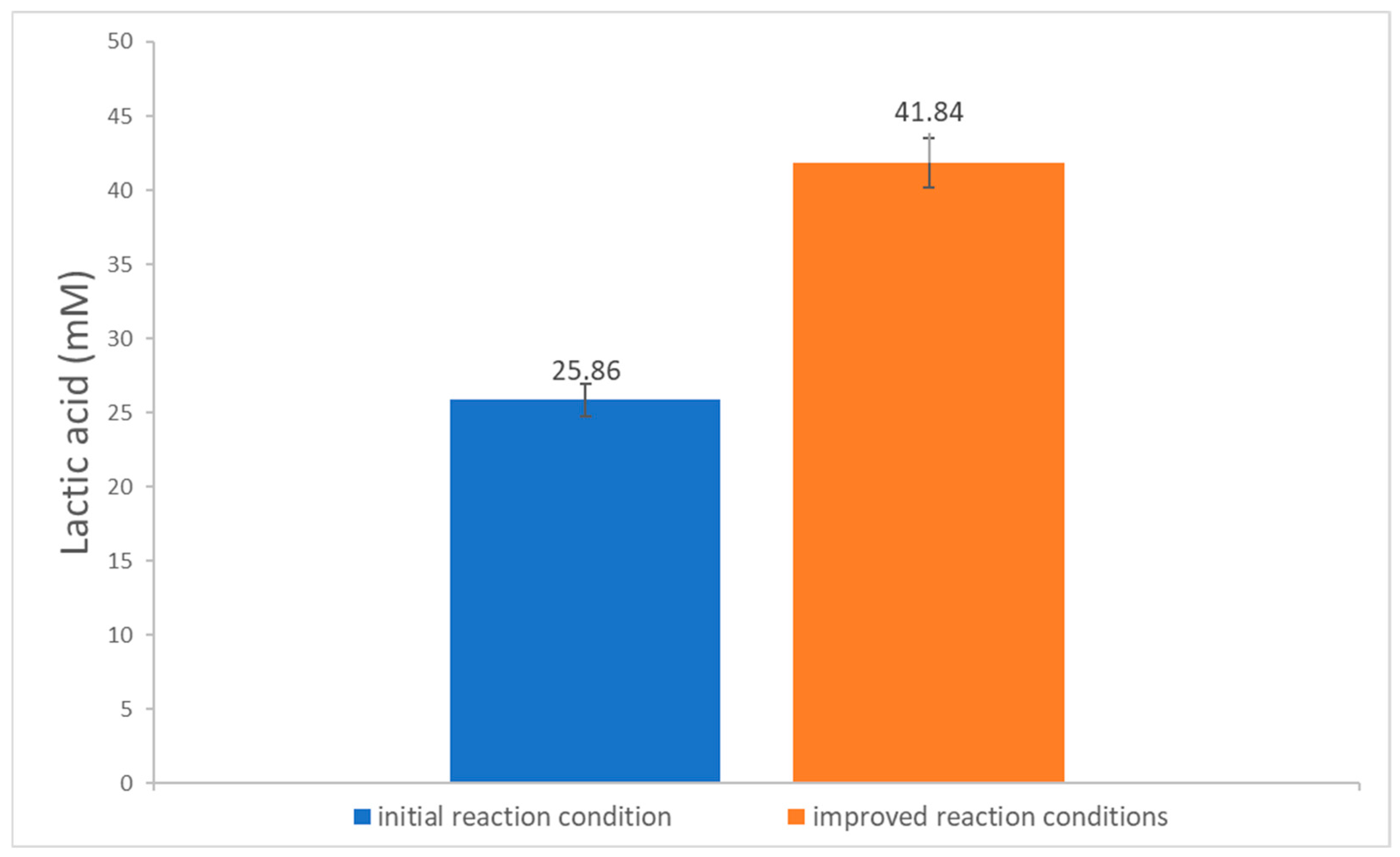
3.3. Establishment of Conditions for the Enzymatic Degradation Reaction
3.4. PLLA Degradation Determined by Weight Loss
3.5. Morphological Changes Evaluated by Scanning Electronic Microscopy (SEM)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tripathi, A.D.; Srivastava, S.K.; Yadav AJ, A.Y. Biopolymers potential biodegradable packaging material for food industry. In Polymers for Packaging Applications; Alavi, S., Thomas, S., Sandeepd, O., Kalarikkal, N., Varghese, J., Yaragalla, S., Eds.; Apple Academic Press: Waretown, NJ, USA, 2014; Volume 153, p. 486. [Google Scholar]
- Wei, R.; Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: How far are we? Microb. Biotechnol. 2017, 10, 1308–1322. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kumar, R.; Singh, J.; Kumar, P. (Eds.) Contaminants in Agriculture and Environment: Health Risks and Remediation; Agro Environ Media, Publication Cell of AESA, Agriculture and Environmental Science Academy: Haridwar, India, 2019; Volume 1. [Google Scholar]
- Jem, K.J.; Tan, B. The development and challenges of poly (lactic acid) and poly (glycolic acid). Adv. Ind. Eng. Polym. Res. 2020, 3, 60–70. [Google Scholar] [CrossRef]
- Sengupta, S.; Manna, S.; Roy, U.; Das, P. Manufacturing of Biodegradable Poly Lactic Acid (PLA): Green Alternatives to Petroleum Derived Plastics. In Reference Module in Materials Science and Materials Engineering; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Ikura, Y.; Kudo, T. Isolation of a microorganism capable of degrading poly-(L-lactide). J. Gen. Appl. Microbiol. 1999, 45, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tomita, T.; Abe, N.; Kamio, Y. Purification and Characterization of an Extracellular Poly(L-Lactic Acid) Depolymerase from a Soil Isolate, Amycolatopsis sp. Strain K104-1. Appl. Environ. Microbiol. 2001, 67, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Masaki, K.; Kamini, N.; Ikeda, H.; Jefugi, H. Cutinase-Like Enzyme from the Yeast Cryptococcus sp. Strain S-2 Hydrolyzes Poly (lactic Acid) and Other Biodegradable Plastics. Appl. Env. Microbiol. 2005, 71, 7548–7550. [Google Scholar] [CrossRef] [PubMed]
- Biundo, A.; Ribitsch, D.; Guebitz, G.M. Surface engineering of polyester-degrading enzymes to improve efficiency and tune specificity. Appl. Microbiol. Biotechnol. 2018, 102, 3551–35599. [Google Scholar] [CrossRef] [PubMed]
- Satti, S.M.; Shah, A.A. Polyester-based biodegradable plastics: An approach towards sustainable development. Lett. Appl. Microbiol. 2020, 70, 413–430. [Google Scholar] [CrossRef]
- Kawai, F.; Nakadai, K.; Nishioka, E.; Nakajima, H.; Ohara, H.; Masaki, K.; Iefuji, H. Different enantio selectivity of two types of poly lactic acid) depolymerases toward poly (L-lactic acid) and poly (D-lactic acid). Polym. Degrad. Stab. 2011, 96, 1342–1348. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Wang, Z.; Lyu, S. A poly (lactic acid) degrading lipase from Bacillus safensis: Characterization and structural analysis. Int. J. Biol. Macromol. 2024, 268, 131916. [Google Scholar] [CrossRef]
- Hia, J.; Zhang, M.; Weng, Y.; Li, C. Degradation of poly(lactic acid)/polybutylene adipate-co-terephthalate by coculture of Pseudomonas mendocina and Actinomucor elegans. J. Hazard. Mater. 2021, 403, 123679. [Google Scholar] [CrossRef]
- Hu, X.; Gu, H.; Sun, X.; Wang, J.; Liu, J.; Yu, Z.; Li, Y.; Jin, J.; Wang, G. Metagenomic exploration of microbial and enzymatic traits involved in microplastic biodegradation. Chemosphere 2024, 348, 140762. [Google Scholar] [CrossRef] [PubMed]
- Krainara, S.; Mistry, A.; Malee, C.; Chavananikul, C.; Pinyakong, O.; Assavalapsakul, W.; SJitpraphai Kachenchart, B.; Luepromchai, E. Development of a plastic waste treatment process by combining deep eutectic solvent (DES) pretreatment and bioaugmentation with a plastic-degrading bacterial consortium. J. Hazard. Mater. 2023, 460, 132507. [Google Scholar] [CrossRef] [PubMed]
- Jamshidian, E.A.; Tehrany, M.; Imran, M.; Jacquot, S. Desobry, Poly-lactic acid: Production, applications, nanocomposites, and release studies, Compr. Rev. Food Sci. Food Saf. 2010, 9, 552–557. [Google Scholar] [CrossRef]
- Karimi-Avargani, M.; Bazooyar, F.; Biria, D.; Zamani, A.; Skrifvars, M. The special effect of the Aspergillus flavus and its enzymes on biological degradation of the intact polylactic acid (PLA) and PLA-Jute composite. Polym. Degrad. Stab. 2020, 179, 109295. [Google Scholar] [CrossRef]
- Bermúdez-García, E.; Peña-Montes, C.; Martins, I.; Pais, J.; Pereira, C.S.; Sánchez, S.; Farrés, A. Regulation of the cutinases expressed by Aspergillus nidulans and evaluation of their role in cutin degradation. Appl. Microbiol. Biotechnol. 2019, 103, 3863–3874. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ochoa, L.; Peña Montes, C.; Esquivel, R.; González Canto, A.; Navarro-Ocaña, A.; Farrés, A. ANCUT2, an extracellular cutinase from Aspergillus nidulans induced by olive oil. Appl. Biochem. Biotechnol. 2012, 166, 1275–1290. [Google Scholar] [CrossRef] [PubMed]
- Farrés, A.; Peña, C.; Hernández, E.; Morales, S.; Solís, I.; Sánchez, M. Cutinasas Recombinantes de Aspergillus Nidulans para Biodegradación de Poliésteres. Mexican Patent MX2016006869A, 26 May 2017. [Google Scholar]
- EasySelect Pichia expression kit (Version G)” (Invitrogen). Available online: https://www.thermofisher.com/order/catalog/product/K174001 (accessed on 28 August 2020).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Karpushova, A.; Brümmer, F.; Barth, S.; Lange, S.; Schmid, R.D. Cloning, recombinant expression and biochemical characterisation of novel esterases from Bacillus sp. associated with the marine sponge Aplysina aerophoba. Appl. Microbiol. Biotechnol. 2005, 67, 59–69. [Google Scholar] [CrossRef]
- Bermúdez-García, E.; Peña-Montes, C.; Castro-Rodríguez, J.A.; González-Canto, A.; Navarro, A.; Farrés, A. ANCUT2, a Thermo-alkaline Cutinase from Aspergillus nidulans and Its Potential Applications. Appl. Biochem. Biotechnol. 2017, 182, 1014–1036. [Google Scholar] [CrossRef]
- Peña-Montes, C.; Bermúdez-García, E.; Castro-Ochoa, D.; Vega-Pérez, F.; Esqueda-Domínguez, K.; Castro-Rodríguez, J.A.; González-Canto, A.; Segoviano-Reyes, L.; Navarro-Ocaña, A.; Farrés, A. ANCUT1, a novel thermoalkaline cutinase from Aspergillus nidulans and its application on hydroxycinnamic acids lipophilization. Biotechnol. Lett. 2024, 46, 409–430. [Google Scholar] [CrossRef] [PubMed]
- Kawai, F. Polylactic Acid (PLA)-Degrading Microorganisms and PLA Depolymerases. In Cheng and Gross; Green Polymer Chemistry: Biocatalysis and Biomaterials ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2010; pp. 405–414. [Google Scholar]
- Carbios, Saint—Beauzire. Method for Recycling Plastic Products. (Patente de Estados Unidos 10, 124, 512 B2. United States Patent. 2019. Available online: https://patentimages.storage.googleapis.com/f5/d4/90/82e5631a15cb7f/US10124512.pdf (accessed on 20 November 2023).
- Shinozaki, Y.; Morita, T.; Cao, X.H.; Yoshida, S.; Koitabashi, M.; Watanabe Watanabe, T.; Suzuki, K.; Yamashita, Y.; Kambe, T.; Fujii, T.; et al. Biodegradable plastic-degrading enzyme from Pseudozyma antarctica: Cloning, sequencing, and characterization. Appl. Microbiol. Biotechnol. 2013, 97, 2951–2959. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Mahumdar, A.; Singh Butola, B. Tailoring the biodegradability of polylactic acid (PLA) based films and ramie- PLA green composites by using selective additives. Int. J. Biol. Macromol. 2021, 181, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Kitadoro, K.; Kakara, M.; Matsui, G.; Osokoshi, R.; Thumarat, R.; Kawai, F.; Kamitani, S. Structural insights into the unique polylactate-degrading mechanism of Thermobifida alba cutinase. FEBS J. 2019, 286, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Mistry, A.N.; Kachenchart, B.; Wongthanaroj, A.; Somwangthanaroj, A.; Luepromchai, E. Rapid biodegradation of high molecular weight semi-crystalline polylactic acid at ambient temperature via enzymatic and alkaline hydrolysis by a defined bacterial consortium. Polym. Degrad. Stab. 2022, 202, 110051. [Google Scholar] [CrossRef]
- Schubert, S.W.; Thomsen, T.B.; Clausen, K.S.; Malmendal, C.; Hunt, J.; Borch, K.; Jensen, K.; Brask, A.; Meyer, S.; Westh, P. Relationships of crystallinity and reaction rates for enzymatic degradation of poly(ethyleneterephthalate), PET. ChemSisChem 2024, 17, e202301752. [Google Scholar] [CrossRef] [PubMed]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. A Publ. Protein Soc. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Protein (mg/mL) | Volumetric Activity (U/mL) | Specific Activity (U/mg) |
|---|---|---|---|
| ANCUT1 | 0.180 | 1716.23 | 17,015.30 |
| ANCUT2 | 0.152 | 6974.90 | 64,486.00 |
| ANCUT3 | 0.168 | 180.92 | 3664.10 |
| Treatment | Weight Loss (%) |
|---|---|
| PLLA with no enzyme (blank) | 0 |
| PLLA-ANCUT 2 | 7.43 ± 0.53 |
| PLLA-ANCUT3 | 6.93 ± 0.35 |
| PLLA-ANCUT1 | 45.96 ± 1.13 |
| PDLA-ANCUT 1 | 36.37 ± 1.35 |
| Commercial package–ANCUT1 | 49.7 ± 1.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado, E.; Castro, R.; Castro-Rodríguez, J.A.; Navarro, A.; Farrés, A. Poly(lactic acid) Degradation by Recombinant Cutinases from Aspergillus nidulans. Polymers 2024, 16, 1994. https://doi.org/10.3390/polym16141994
Alvarado E, Castro R, Castro-Rodríguez JA, Navarro A, Farrés A. Poly(lactic acid) Degradation by Recombinant Cutinases from Aspergillus nidulans. Polymers. 2024; 16(14):1994. https://doi.org/10.3390/polym16141994
Chicago/Turabian StyleAlvarado, Eric, Rafael Castro, José Augusto Castro-Rodríguez, Arturo Navarro, and Amelia Farrés. 2024. "Poly(lactic acid) Degradation by Recombinant Cutinases from Aspergillus nidulans" Polymers 16, no. 14: 1994. https://doi.org/10.3390/polym16141994
APA StyleAlvarado, E., Castro, R., Castro-Rodríguez, J. A., Navarro, A., & Farrés, A. (2024). Poly(lactic acid) Degradation by Recombinant Cutinases from Aspergillus nidulans. Polymers, 16(14), 1994. https://doi.org/10.3390/polym16141994