

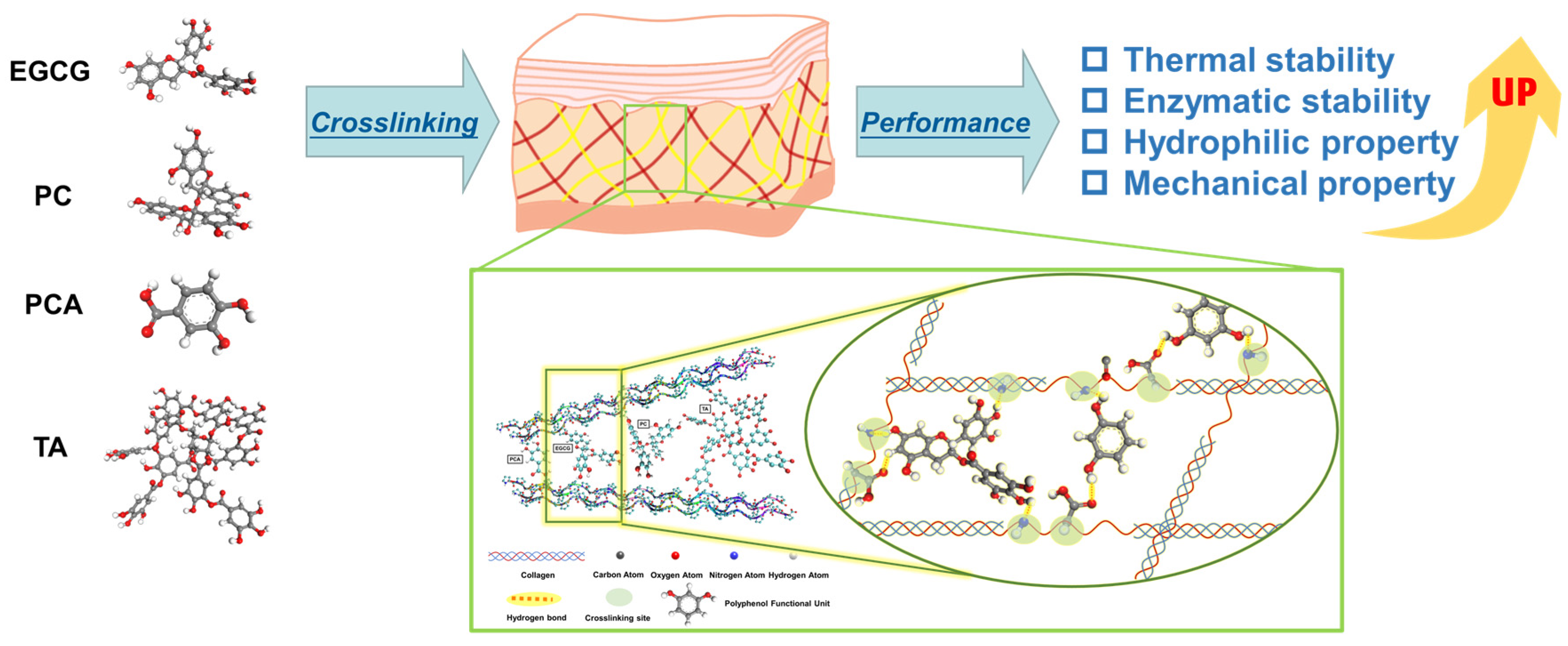
Influences of Polyphenols on the Properties of Crosslinked Acellular Fish Swim Bladders: Experiments and Molecular Dynamic Simulations



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Crosslinking of AFSBs
2.3. Simulation Details
3. Results and Discussion
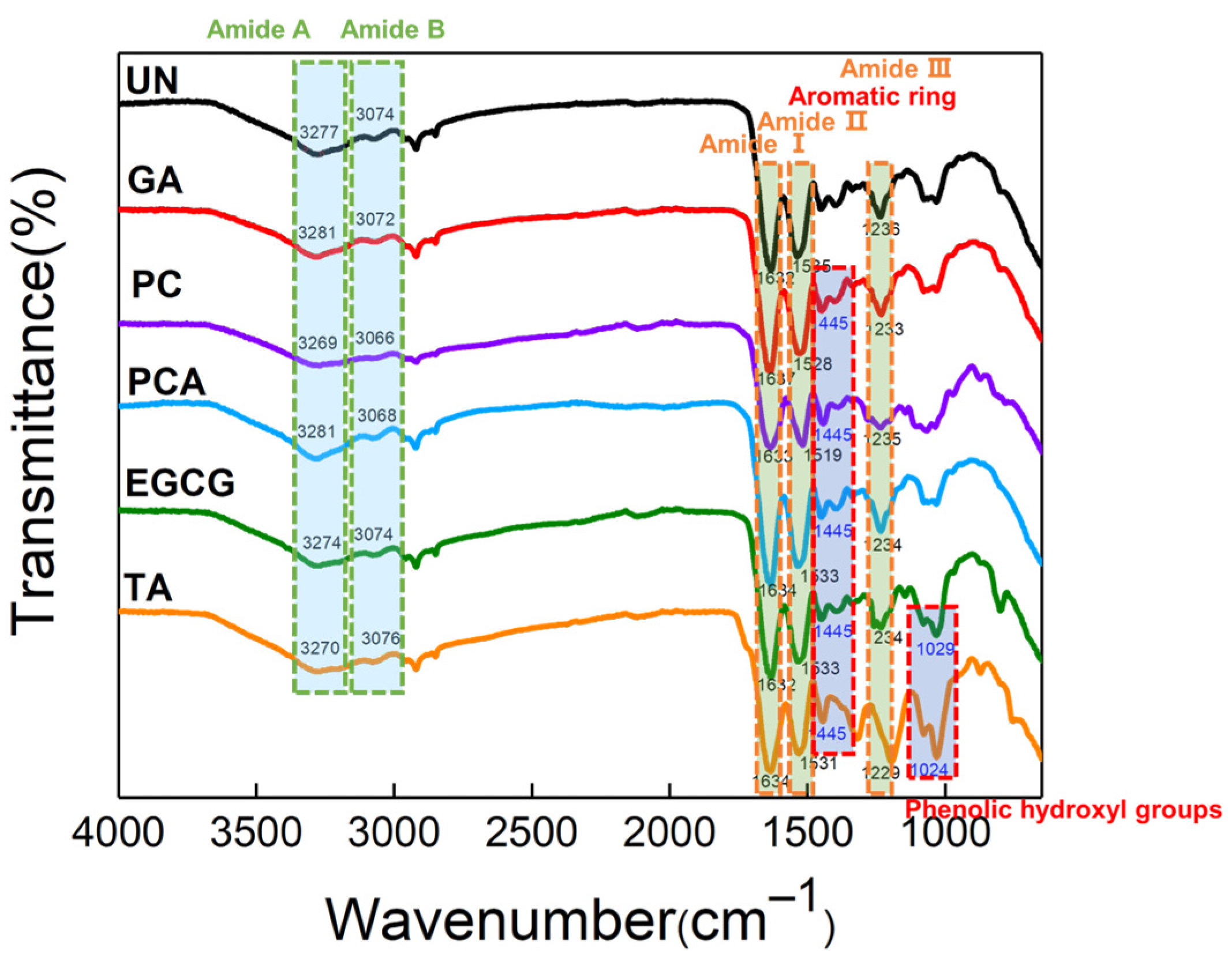
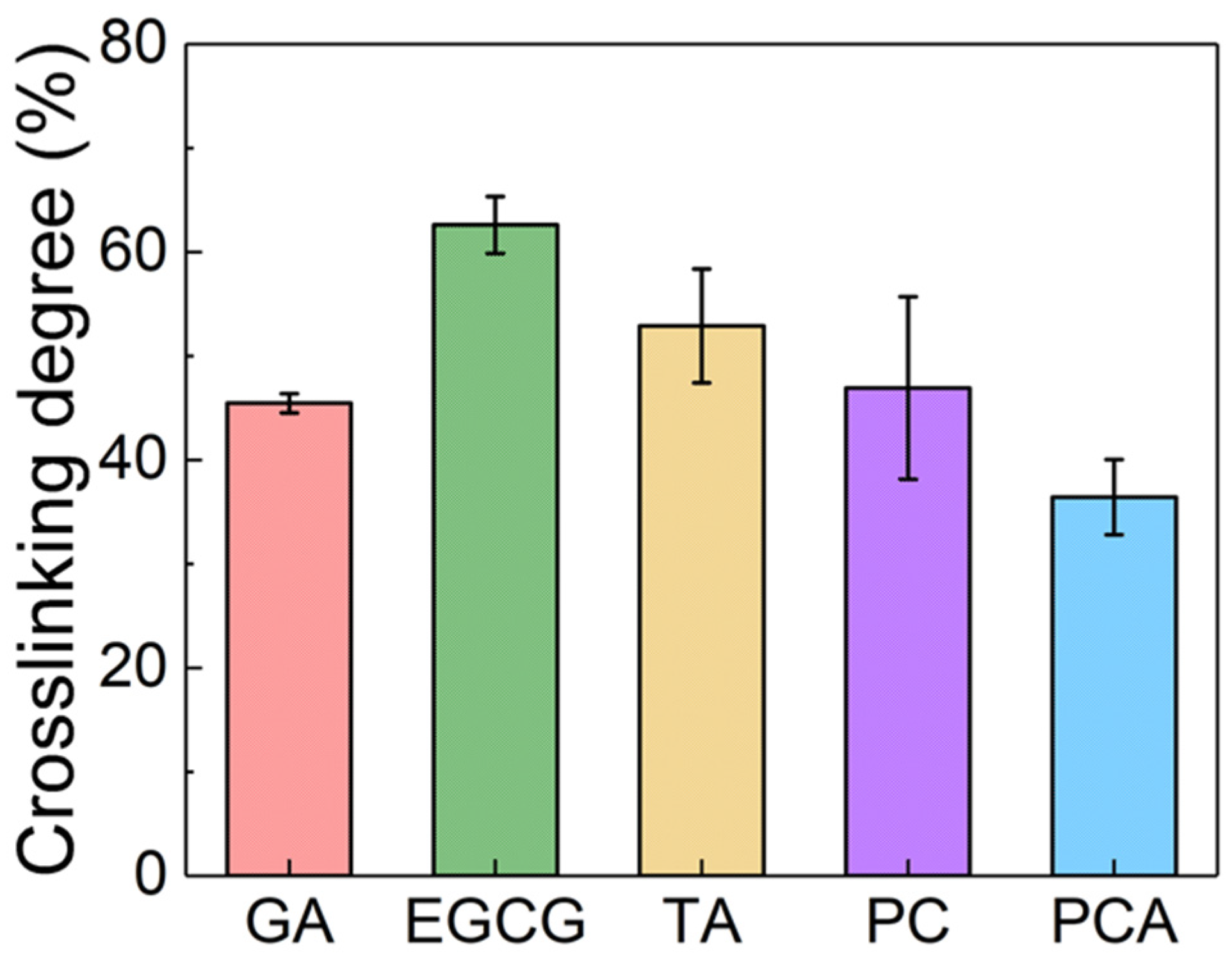
3.1. Structural Characterization of AFSBs
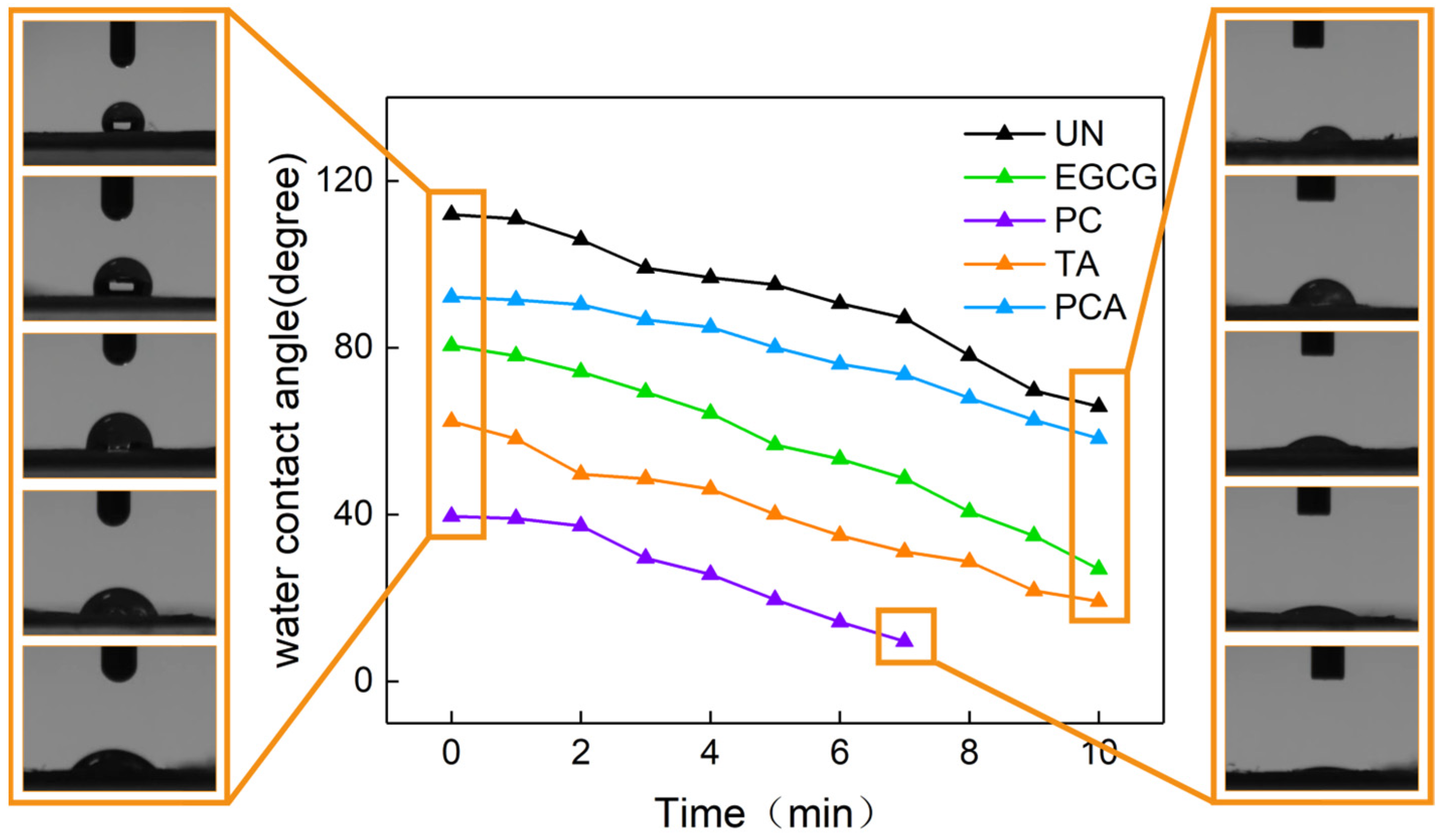
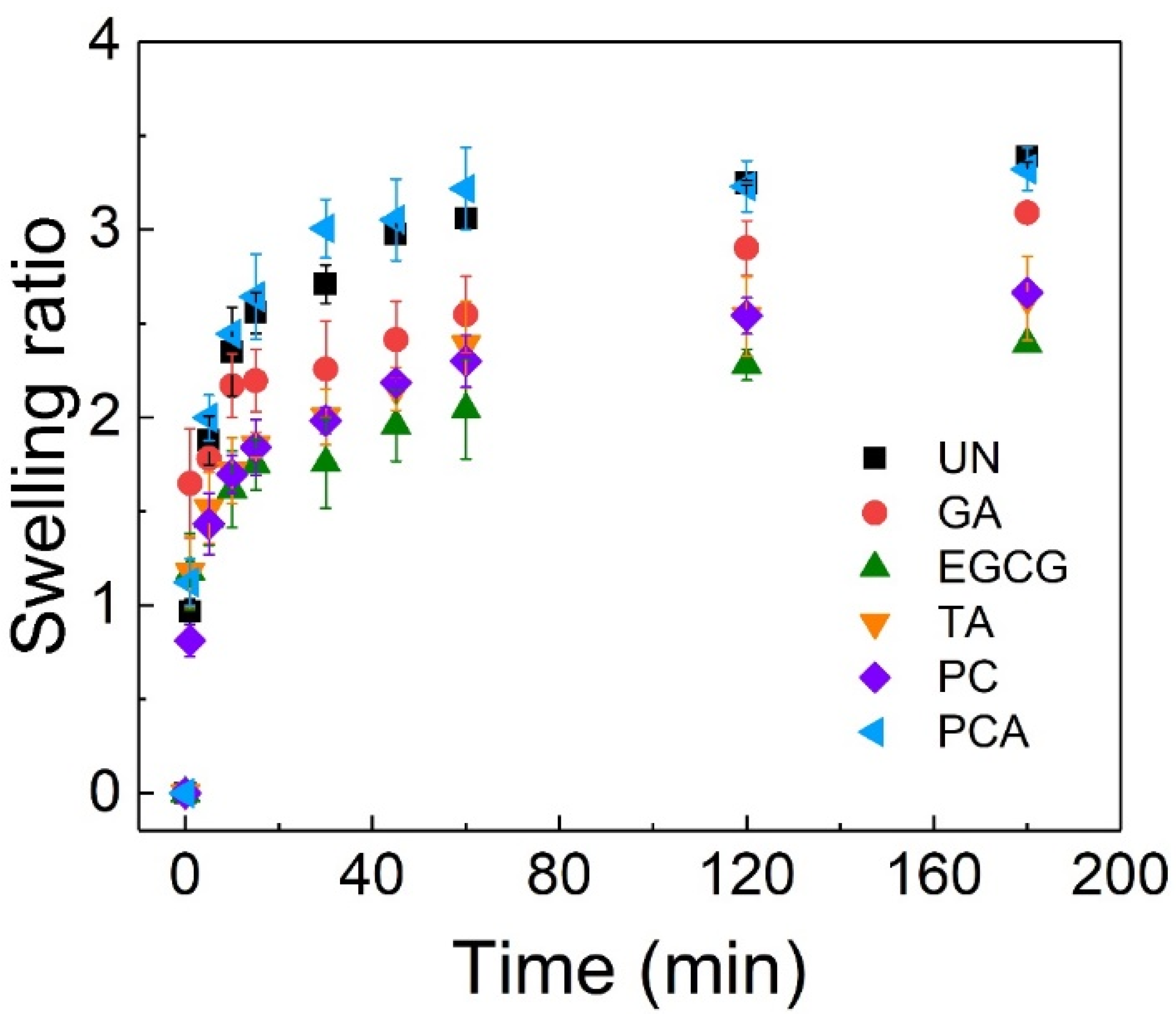
3.2. Surface Wettability and Swelling Behavior
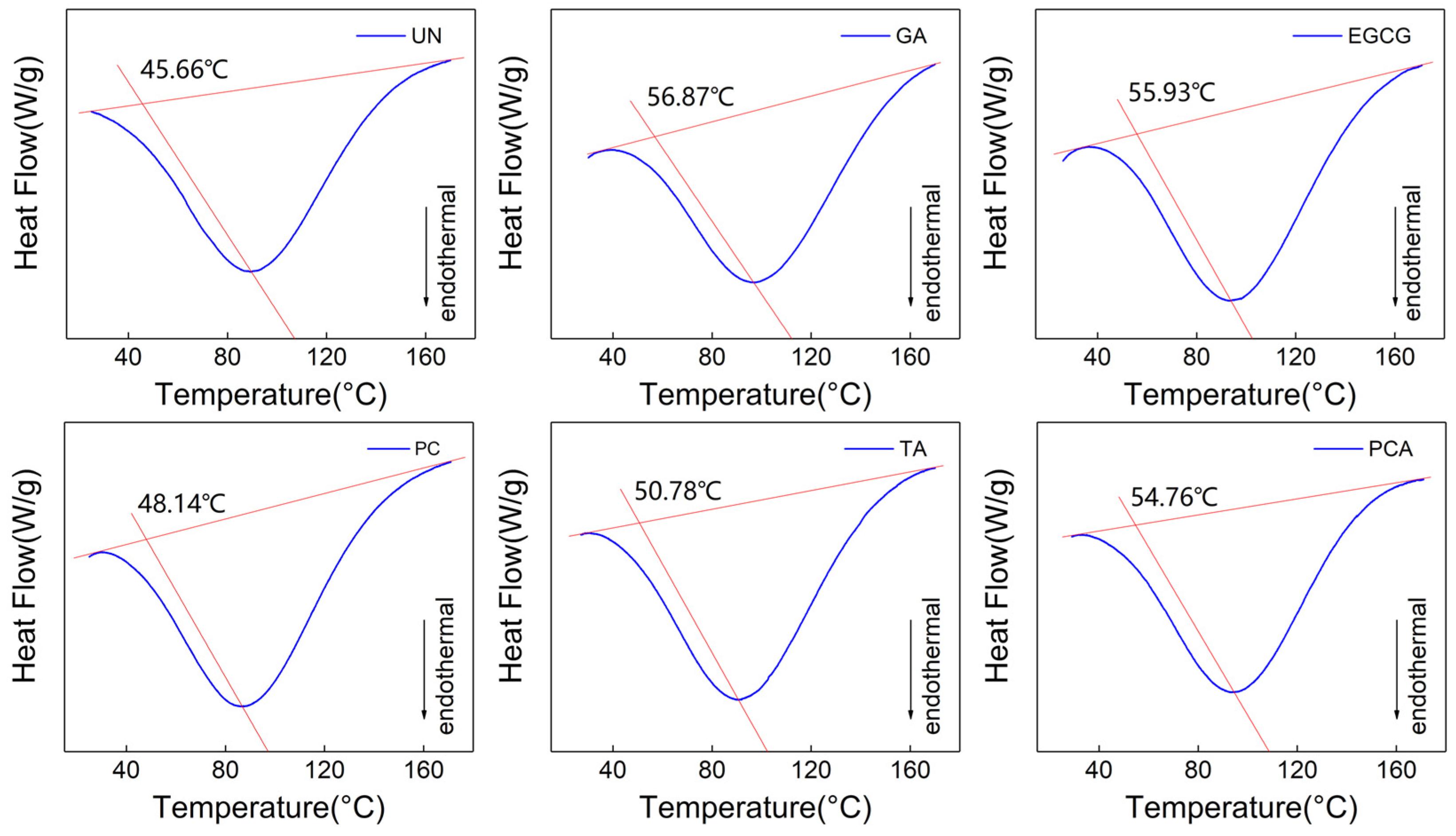
3.3. Thermomechanical Properties
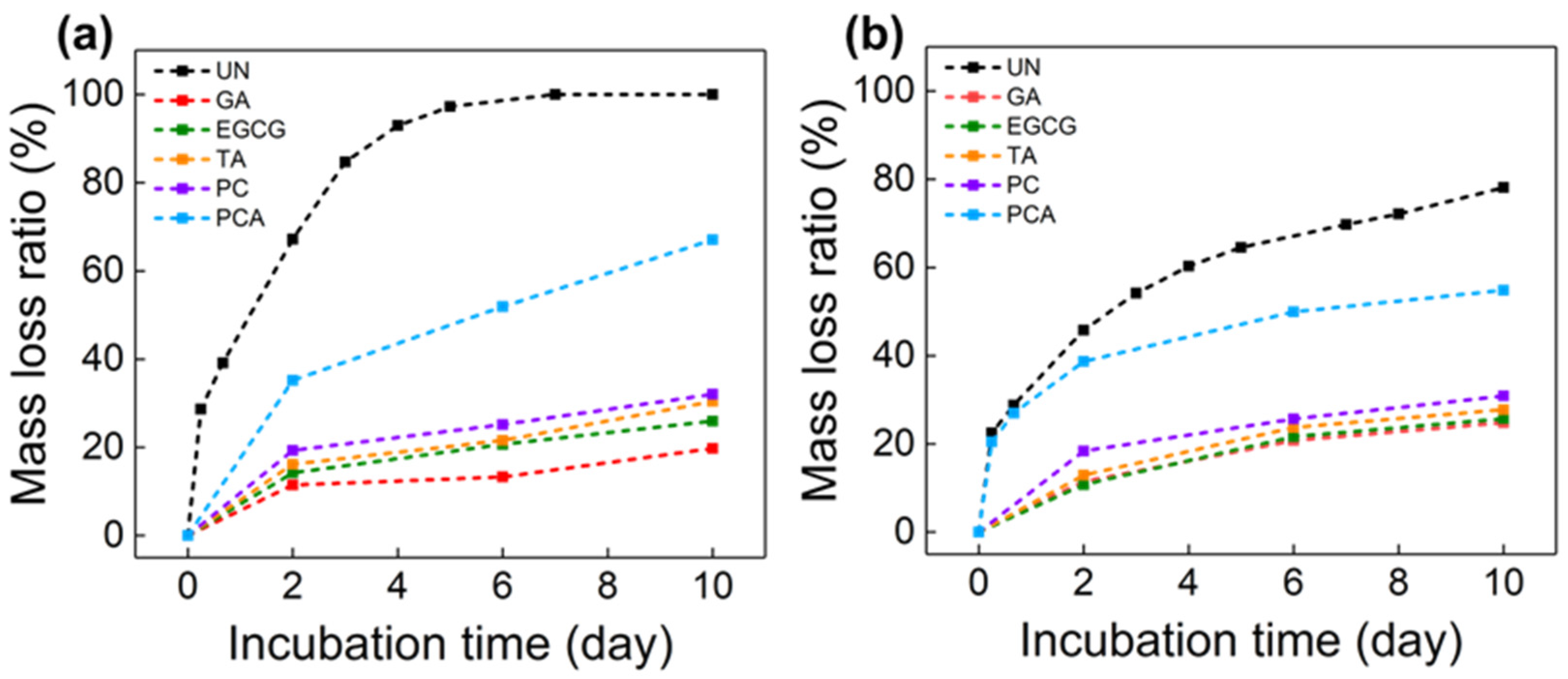
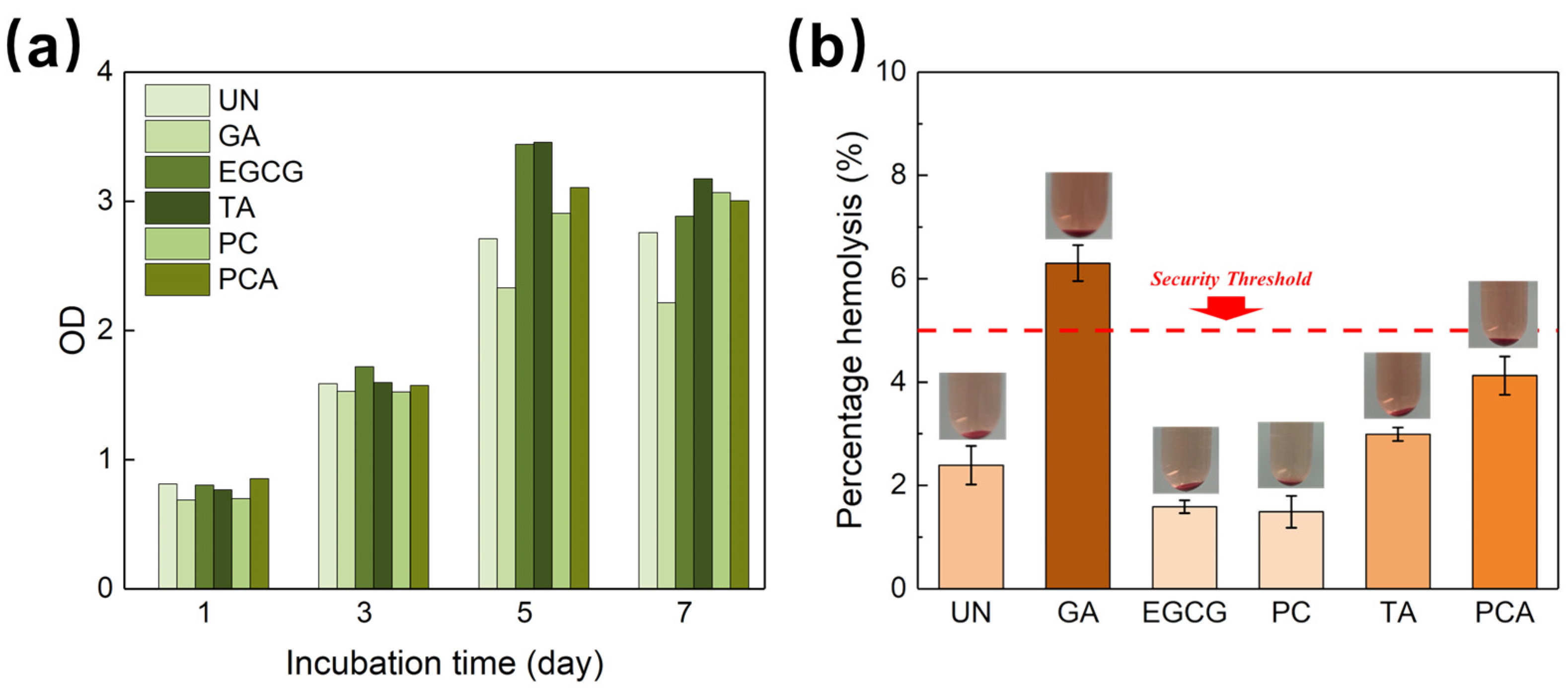
3.4. In Vitro Enzymatic Stability and Biocompatibility
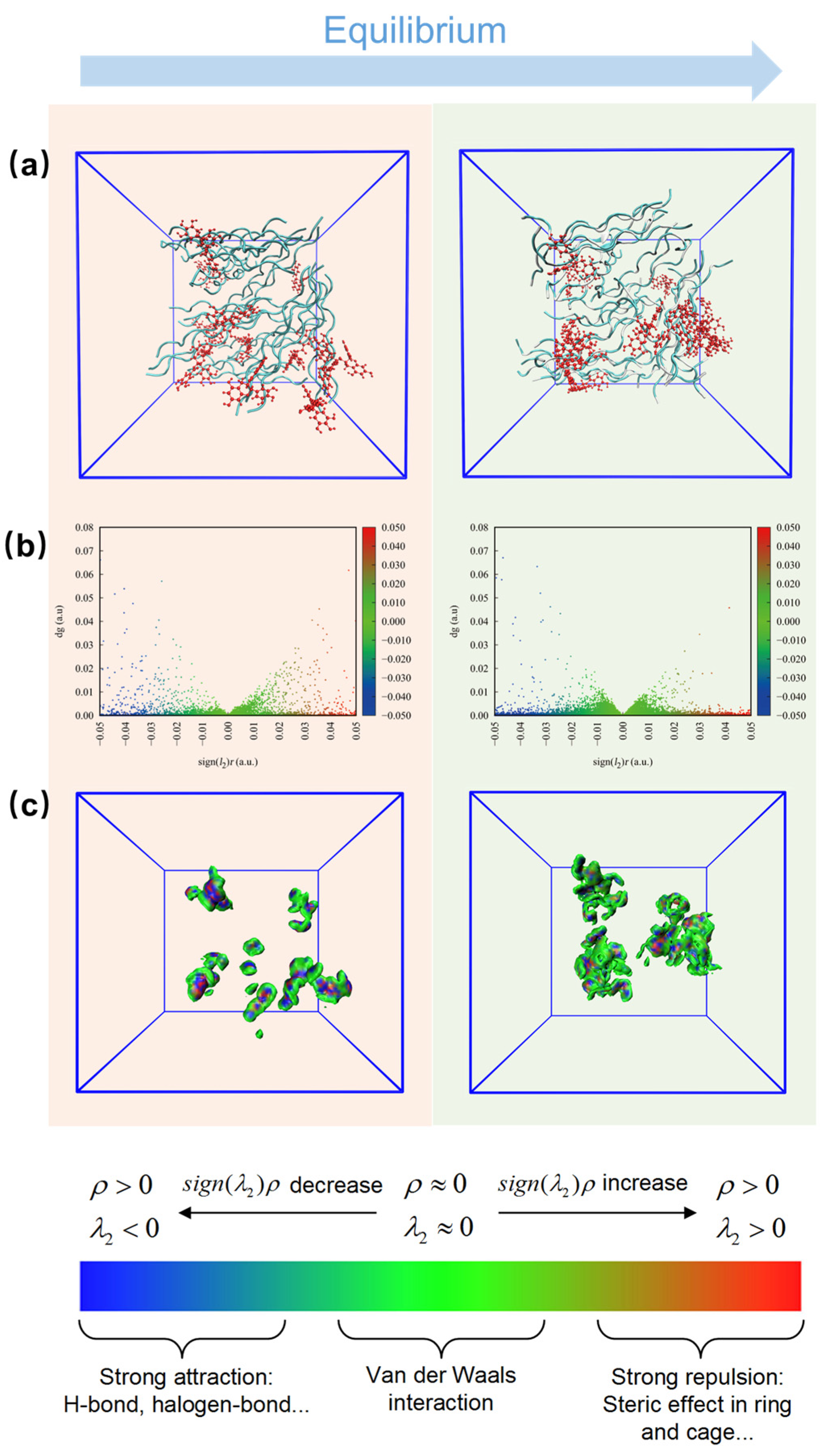
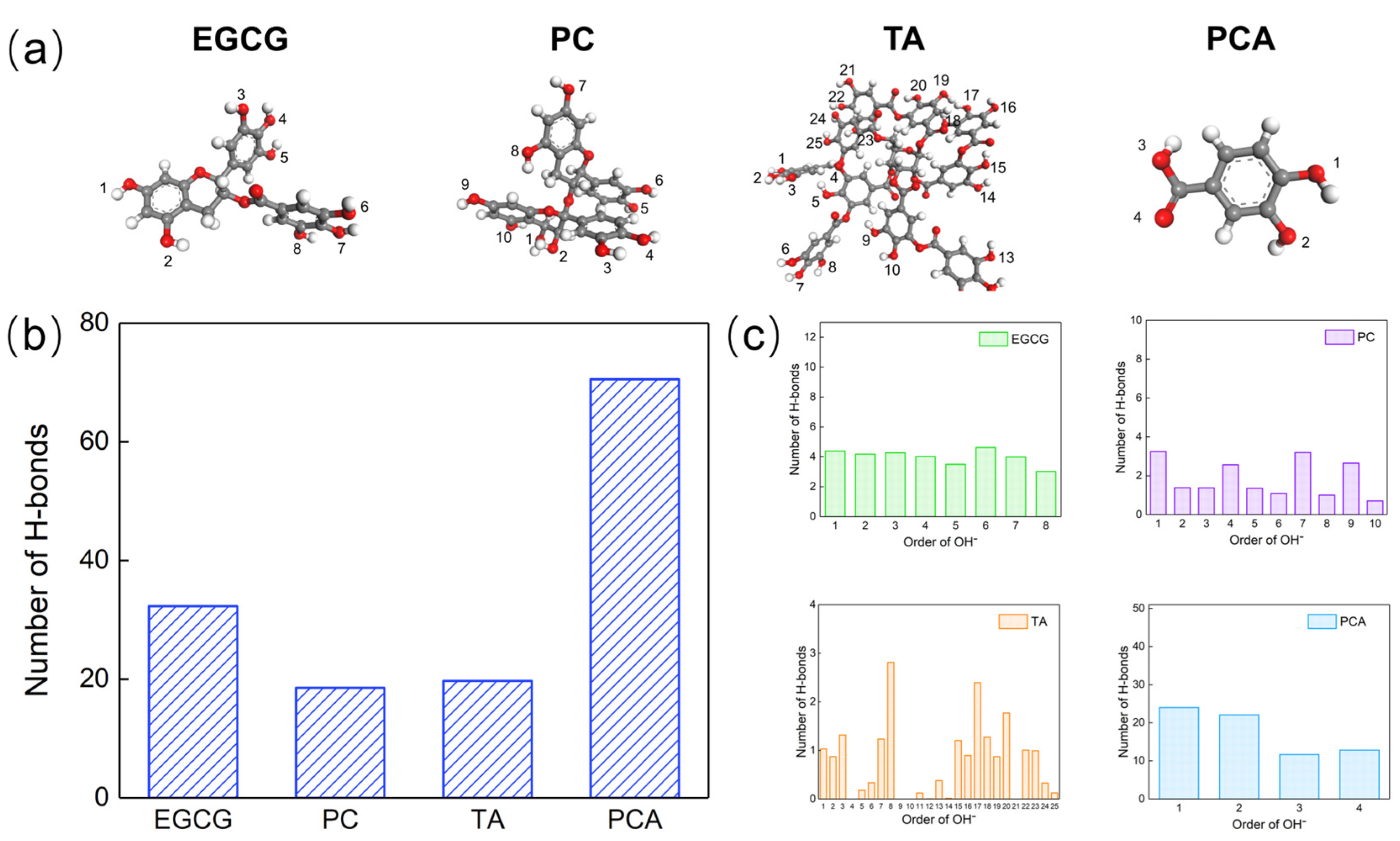
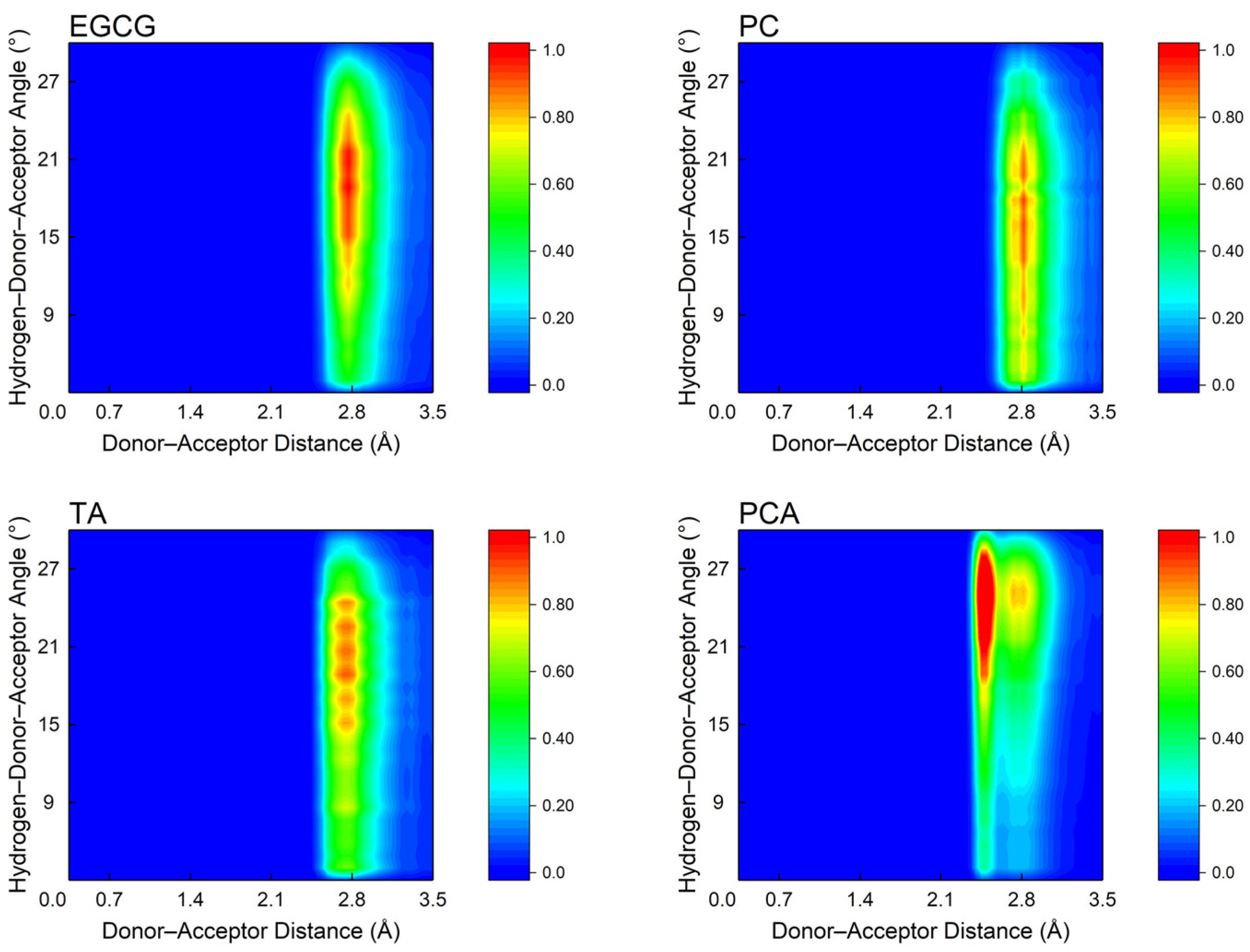
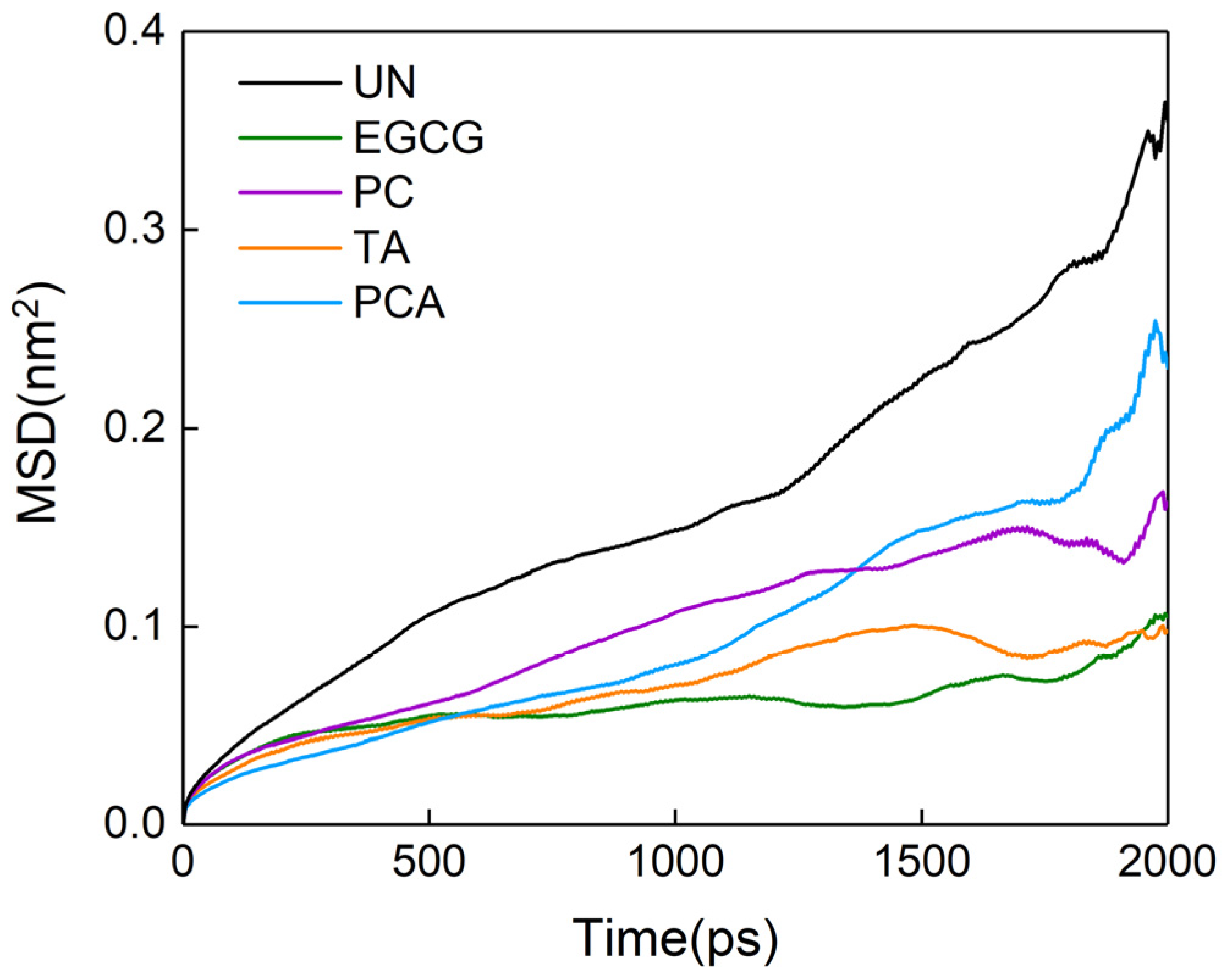
3.5. Microscopic Interactions between Polyphenols and AFSBs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rose, C.; Mandal, A.B.; Joseph, K.T. Characterization of collagen from the Swimbladder of Catfish (Tachysurus maculates). Asian Fish. Sci. 1998, 11, 1–10. [Google Scholar]
- Li, Q.; Zhang, F.; Wang, H.; Pan, T. Preparation and characterization of a novel acellular swim bladder as dura mater substitute. Neurol. Res. 2019, 41, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, B.; Jing, H.; Wu, Y.; Kong, D.; Leng, X.; Wang, Z. Swim Bladder as a Novel Biomaterial for Cardiovascular Materials with Anti-Calcification Properties. Adv. Healthc. Mater. 2020, 9, 1901154. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Sun, P.; Wu, H.; Wei, S.; Xie, B.; Wang, W.; Hou, Y.; Li, J.A.; Dardik, A.; Li, Z. The application of tissue-engineered fish swim bladder vascular graft. Commun. Biol. 2021, 4, 1153. [Google Scholar] [CrossRef]
- Sun, P.; Wu, H.; Bai, X.; Zhang, L.; Zhang, C.; Wang, X.; Lou, C.; Li, B.; Li, Z.; Bai, H. Decellularized fish swim bladder patch loaded with mesenchymal stem cells inhibits neointimal hyperplasia. J. Biomed. Mater. Res. Part B 2023, 111, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jing, H.; Sun, Z.; Wang, X.; Kong, D.; Liu, J.; Leng, X.; Wang, Z. Comprehensive analyses and prioritization of various swim bladder-derived extracellular matrix in the application of heart valve prosthesis. Smart Mater. Med. 2021, 2, 209–218. [Google Scholar] [CrossRef]
- Howaili, F.; Mashreghi, M.; Shahri, N.M.; Kompany, A.; Jalal, R. Development and evaluation of a novel beneficent antimicrobial bioscaffold based on animal waste-fish swim bladder (FSB) doped with silver nanoparticles. Environ. Res. 2020, 188, 109823. [Google Scholar] [CrossRef]
- Mahesh, R.; Sreenu, M.; Kishore, P.V.S.; Kumar, R.V.S. Acellular Matrix of Swim Bladder for Cervical Oesophagoplasty in Rabbits. Trends Biomater. Artif. Organs. 2013, 27, 146–152. [Google Scholar]
- Lavanya, K.; Sreenu, M.; Prasad, V.D.; Suresh, K. Haemato-biochemical studies on acellular matrix of swim bladder for cystoplasty in rabbits. Indo-Am. J. Agric. Vet. Sci. 2014, 48, 459–463. [Google Scholar]
- Damink, L.H.H.O.; Dijkstra, P.J.; van Luyn, M.J.A.; van Wachem, P.B.; Nieuwenhuis, P.; Feijen, J. In vitro degradation of dermal sheep collagen cross-linked using a water-soluble carbodiimide. Biomaterials 1996, 17, 679–684. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, N.; Singh, H.; Gangwar, A.K.; Dewangan, R.; Kumar, A.; Rai, R.B. Effects of crosslinking treatments on the physical properties of acellular fish swim bladder. Trends Biomater. Artif. Organs. 2013, 27, 93–101. [Google Scholar]
- Li, H.-C.; Sun, X.-M.; Huang, Y.-R.; Peng, Y.-H.; Liu, J.; Ren, L. Synthetic Crosslinker Based on Amino–yne Click to Enhance the Suture Tension of Collagen-Based Corneal Repair Materials. ACS Appl. Polym. Mater. 2022, 4, 4495–4507. [Google Scholar] [CrossRef]
- Bax, D.V.; Davidenko, N.; Hamaia, S.W.; Farndale, R.W.; Best, S.M.; Cameron, R.E. Impact of UV- and carbodiimide-based crosslinking on the integrin-binding properties of collagen-based materials. Acta Biomater. 2019, 100, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.W.; Siddiq, M. Quantum Mechanics of In Situ Synthesis of Metal Nanoparticles within Anionic Microgels. J. Theor. Chem. 2013, 2013, 410417. [Google Scholar] [CrossRef]
- Broguiere, N.; Formica, F.; Barreto, G.; Zenobi-Wong, M. Sortase A as a cross-linking enzyme in tissue engineering. Acta Biomater. 2018, 77, 182–190. [Google Scholar] [CrossRef]
- Veríssimo, D.M.; Leitão, R.F.C.; Ribeiro, R.A.; Figueiró, S.D.; Sombra, A.S.B.; Góes, J.C.; Brito, G.A.C. Polyanionic collagen membranes for guided tissue regeneration: Effect of progressive glutaraldehyde cross-linking on biocompatibility and degradation. Acta Biomater. 2010, 6, 4011–4018. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Yue, W.; Li, J.; Yu, X.; Ying, T.; Liu, R.; You, J.; Xiong, S.; Hu, Y. Effects of Phenolics on the Physicochemical and Structural Properties of Collagen Hydrogel. Polymers 2023, 15, 4647. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Mu, C.; Shi, J.; Zhang, Q.; Shi, B.; Lin, W. Modification of collagen with a natural cross-linker, procyanidin. Int. J. Biol. Macromol. 2011, 48, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Galicka, A.; Nazaruk, J. Stimulation of collagen biosynthesis by flavonoid glycosides in skin fibroblasts of osteogenesis imperfecta type I and the potential mechanism of their action. Int. J. Mol. Med. 2007, 20, 889–895. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhai, W.; Chang, J.; Lü, X.; Wang, Z. Procyanidins-crosslinked heart valve matrix: Anticalcification effect. J. Biomed. Mater. Res. Part B 2009, 90, 913–921. [Google Scholar] [CrossRef]
- Zhai, W.; Chang, J.; Lin, K.; Wang, J.; Zhao, Q.; Sun, X. Crosslinking of decellularized porcine heart valve matrix by procyanidins. Biomaterials 2006, 27, 3684–3690. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Lü, X.; Chang, J.; Zhou, Y.; Zhang, H. Quercetin-crosslinked porcine heart valve matrix: Mechanical properties, stability, anticalcification and cytocompatibility. Acta Biomater. 2010, 6, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, B.; Jing, H.; Qin, Y.; Wu, Y.; Kong, D.; Leng, X.; Wang, Z. Curcumin-crosslinked acellular bovine pericardium for the application of calcification inhibition heart valves. Biomed. Mater. 2020, 15, 045002. [Google Scholar] [CrossRef] [PubMed]
- Russo, N.; Cassinelli, C.; Torre, E.; Morra, M.; Iviglia, G. Improvement of the physical properties of guided bone regeneration membrane from porcine pericardium by polyphenols-rich pomace extract. Materials 2019, 12, 2564. [Google Scholar] [CrossRef]
- Manjari, M.S.; Aaron, K.P.; Muralidharan, C.; Rose, C. Highly biocompatible novel polyphenol cross-linked collagen scaffold for potential tissue engineering applications. React. Funct. Polym. 2020, 153, 104630. [Google Scholar] [CrossRef]
- Qu, W.; Xiong, T.; Wang, B.; Li, Y.; Zhang, X. The modification of pomegranate polyphenol with ultrasound improves mechanical, antioxidant, and antibacterial properties of tuna skin collagen-chitosan film. Ultrason. Sonochem. 2022, 85, 105992. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Shao, H.; Fang, Z.; Zhao, Y.; Cao, C.Y.; Li, Q. Mechanism and effects of polyphenol derivatives for modifying collagen. ACS Biomater. Sci. Eng. 2019, 5, 4272–4284. [Google Scholar] [CrossRef]
- Haslam, E.; Lilley, T.; Cai, Y.; Martin, R.; Mangnolato, D. Traditional herbal medicines-the role of polyphenols. Planta Med. 1989, 55, 1–8. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Schmid, N.; Eichenberger, A.P.; Choutko, A.; Riniker, S.; Winger, M.; Mark, A.E.; van Gunsteren, W.F. Definition and testing of the GROMOS force-field versions 54A7 and 54B7. Eur. Biophys. J. Biophys. Lett. 2011, 40, 843. [Google Scholar] [CrossRef]
- Stroet, M.; Caron, B.; Visscher, K.M.; Geerke, D.P.; Malde, A.K.; Mark, A.E. Automated topology builder version 3.0: Prediction of solvation free enthalpies in water and hexane. J. Chem. Theory Comput. 2018, 14, 5834–5845. [Google Scholar] [CrossRef] [PubMed]
- Mark, P.; Nilsson, L. Structure and dynamics of the TIP3P, SPC, and SPC/E water models at 298 K. J. Phys. Chem. B 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Martínez, L.; Andrade, R.; Birgin, E.G.; Martínez, J.M. PACKMOL: A package for building initial configurations for molecular dynamics simulations. J. Comput. Chem. 2009, 30, 2157–2164. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. Model. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Markel, D.C.; Wang, S.; Shi, T.; Mao, G.; Ren, W. Electrospun polyvinyl alcohol–collagen–hydroxyapatite nanofibers: A biomimetic extracellular matrix for osteoblastic cells. Nanotechnology 2012, 23, 115101. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.E.; Murali, J.; Muralidharan, C.; Madhulata, W.; Jayakumar, R. Interaction of collagen with corilagin. Colloid. Polym. Sci. 2003, 281, 766–770. [Google Scholar] [CrossRef]
- Bal Ram, S.; Daniel, B.D.; Fen-Ni, F.; Michael, P.F. Fourier transform infrared analysis of amide III bands of proteins for the secondary structure estimation. In Biomolecular Spectroscopy III; SPIE: Los Angeles, CA, USA, 1993; pp. 47–55. [Google Scholar]
- Mu, C.; Liu, F.; Cheng, Q.; Li, H.; Wu, B.; Zhang, G.; Lin, W. Collagen Cryogel Cross-Linked by Dialdehyde Starch. Macromol. Mater. Eng. 2010, 295, 100–107. [Google Scholar] [CrossRef]
- Madhan, B.; Subramanian, V.; Rao, J.R.; Nair, B.U.; Ramasami, T. Stabilization of collagen using plant polyphenol: Role of catechin. Int. J. Biol. Macromol. 2005, 37, 47–53. [Google Scholar] [CrossRef]
- Payne, K.J.; Veis, A. Fourier transform ir spectroscopy of collagen and gelatin solutions: Deconvolution of the amide I band for conformational studies. Biopolymers 1988, 27, 1749–1760. [Google Scholar] [CrossRef]
- Ku, C.S.; Sathishkumar, M.; Mun, S.P. Binding affinity of proanthocyanidin from waste Pinus radiata bark onto proline-rich bovine achilles tendon collagen type I. Chemosphere 2007, 67, 1618–1627. [Google Scholar] [CrossRef]
- Canon, F.; Giuliani, A.; Paté, F.; Sarni-Manchado, P. Ability of a salivary intrinsically unstructured protein to bind different tannin targets revealed by mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 815–822. [Google Scholar] [CrossRef]
- Simon, C.; Barathieu, K.; Laguerre, M.; Schmitter, J.-M.; Fouquet, E.; Pianet, I.; Dufourc, E.J. Three-Dimensional Structure and Dynamics of Wine Tannin−Saliva Protein Complexes. A Multitechnique Approach. Biochemistry 2003, 42, 10385–10395. [Google Scholar] [CrossRef]
- Zhang, X.; Do, M.D.; Casey, P.; Sulistio, A.; Qiao, G.G.; Lundin, L.; Lillford, P.; Kosaraju, S. Chemical Modification of Gelatin by a Natural Phenolic Cross-linker, Tannic Acid. J. Agric. Food Chem. 2010, 58, 6809–6815. [Google Scholar] [CrossRef] [PubMed]
- Karri, V.V.S.R.; Kuppusamy, G.; Talluri, S.V.; Mannemala, S.S.; Kollipara, R.; Wadhwani, A.D.; Mulukutla, S.; Raju, K.R.S.; Malayandi, R. Curcumin loaded chitosan nanoparticles impregnated into collagen-alginate scaffolds for diabetic wound healing. Int. J. Biol. Macromol. 2016, 93, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Zeugolis, D.I.; Khew, S.T.; Yew, E.S.Y.; Ekaputra, A.K.; Tong, Y.W.; Yung, L.-Y.L.; Hutmacher, D.W.; Sheppard, C.; Raghunath, M. Electro-spinning of pure collagen nano-fibres—Just an expensive way to make gelatin? Biomaterials 2008, 29, 2293–2305. [Google Scholar] [CrossRef] [PubMed]
- Pietrucha, K. Changes in denaturation and rheological properties of collagen–hyaluronic acid scaffolds as a result of temperature dependencies. Int. J. Biol. Macromol. 2005, 36, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, G.; Selvakumar, R.; Sastry, T.P.; Sadulla, S.; Mandal, A.B.; Doble, M. Experimental and theoretical studies on Gallic acid assisted EDC/NHS initiated crosslinked collagen scaffolds. Mater. Sci. Eng. C 2014, 43, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Covington, A.D.; Hancock, R. Structure–activity relationships in the hydrophobic interactions of polyphenols with cellulose and collagen. Biopolymers 2003, 70, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, B.; Chang, J. Preparation of decellularized vascular matrix by co-crosslinking of procyanidins and glutaraldehyde. Bio-Med. Mater. Eng. 2015, 26, 19–30. [Google Scholar] [CrossRef]
- Zilberman, M.; Elsner, J.J. Antibiotic-eluting medical devices for various applications. J. Control. Release 2008, 130, 202–215. [Google Scholar] [CrossRef]
- Chung, L.; Dinakarpandian, D.; Yoshida, N.; Layer-Fields, J.L.; Fields, G.B.; Visse, R.; Nagase, H. Collagenase unwinds triple-helical collagen prior to peptide bond hydrolysis. EMBO J. 2004, 23, 3020–3030. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M. Exploring a role in tanning for the gap region of the collagen fibril: Catechin-collagen interactions. J. Am. Leather Chem. Assoc. 2008, 103, 290–297. [Google Scholar]
- Fitzmaurice, S.D.; Sivamani, R.K.; Isseroff, R.R. Antioxidant therapies for wound healing: A clinical guide to currently commercially available products. Skin Pharmacol. Physiol. 2011, 24, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Chen, Q. Independent gradient model based on Hirshfeld partition: A new method for visual study of interactions in chemical systems. J. Comput. Chem. 2022, 43, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Tian Lu, F. Multiwfn: A multifunctional Wavefunction Analyzer. J. Comput. Chem. 2012, 33, 580–592. [Google Scholar]
- Johnson, E.R.; Keinan, S.; Mori-Sánchez, P.; Contreras-García, J.; Cohen, A.J.; Yang, W. Revealing Noncovalent Interactions. J. Am. Chem. Soc. 2010, 132, 6498–6506. [Google Scholar] [CrossRef]
- Kaparekar, P.S.; Pathmanapan, S.; Anandasadagopan, S.K. Polymeric scaffold of Gallic acid loaded chitosan nanoparticles infused with collagen-fibrin for wound dressing application. Int. J. Biol. Macromol. 2020, 165, 930–947. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Young’s Modulus (MPa) | Ultimate Tensile Strength (MPa) | Elongation at Break (%) |
|---|---|---|---|
| UN | 9.68 | 2.60 | 36.9 |
| GA | 12.38 | 4.35 | 46.9 |
| EGCG | 30.62 | 8.55 | 41.4 |
| PC | 27.51 | 7.03 | 43.9 |
| TA | 27.65 | 7.34 | 43.4 |
| PCA | 17.56 | 3.61 | 30.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; Jiang, J.; Li, J.; Zhao, L.; Xi, Z. Influences of Polyphenols on the Properties of Crosslinked Acellular Fish Swim Bladders: Experiments and Molecular Dynamic Simulations. Polymers 2024, 16, 1111. https://doi.org/10.3390/polym16081111
Han Y, Jiang J, Li J, Zhao L, Xi Z. Influences of Polyphenols on the Properties of Crosslinked Acellular Fish Swim Bladders: Experiments and Molecular Dynamic Simulations. Polymers. 2024; 16(8):1111. https://doi.org/10.3390/polym16081111
Chicago/Turabian StyleHan, Yuqing, Jie Jiang, Jinjin Li, Ling Zhao, and Zhenhao Xi. 2024. "Influences of Polyphenols on the Properties of Crosslinked Acellular Fish Swim Bladders: Experiments and Molecular Dynamic Simulations" Polymers 16, no. 8: 1111. https://doi.org/10.3390/polym16081111
APA StyleHan, Y., Jiang, J., Li, J., Zhao, L., & Xi, Z. (2024). Influences of Polyphenols on the Properties of Crosslinked Acellular Fish Swim Bladders: Experiments and Molecular Dynamic Simulations. Polymers, 16(8), 1111. https://doi.org/10.3390/polym16081111