Large Scale Screening of Rhizospheric Allelopathic Bacteria and Their Potential for the Biocontrol of Wheat-Associated Weeds

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Rhizobacteria
2.2. Cyanide Production Assay on Strains of Rhizobacteria
2.3. Antimetabolite Assay on E. coli
2.4. Antimetabolite Assay on Lettuce (Lectuca sativa L.) Seedlings
2.5. Antimetabolite Assay on Weeds Using Presumed Allelopathic Bacteria
2.6. Antimetabolite Assay on Wheat Using Presumed Allelopathic Bacteria
2.7. Cluster Analysis for the Screening of Biological Weed Control Agents
3. Results
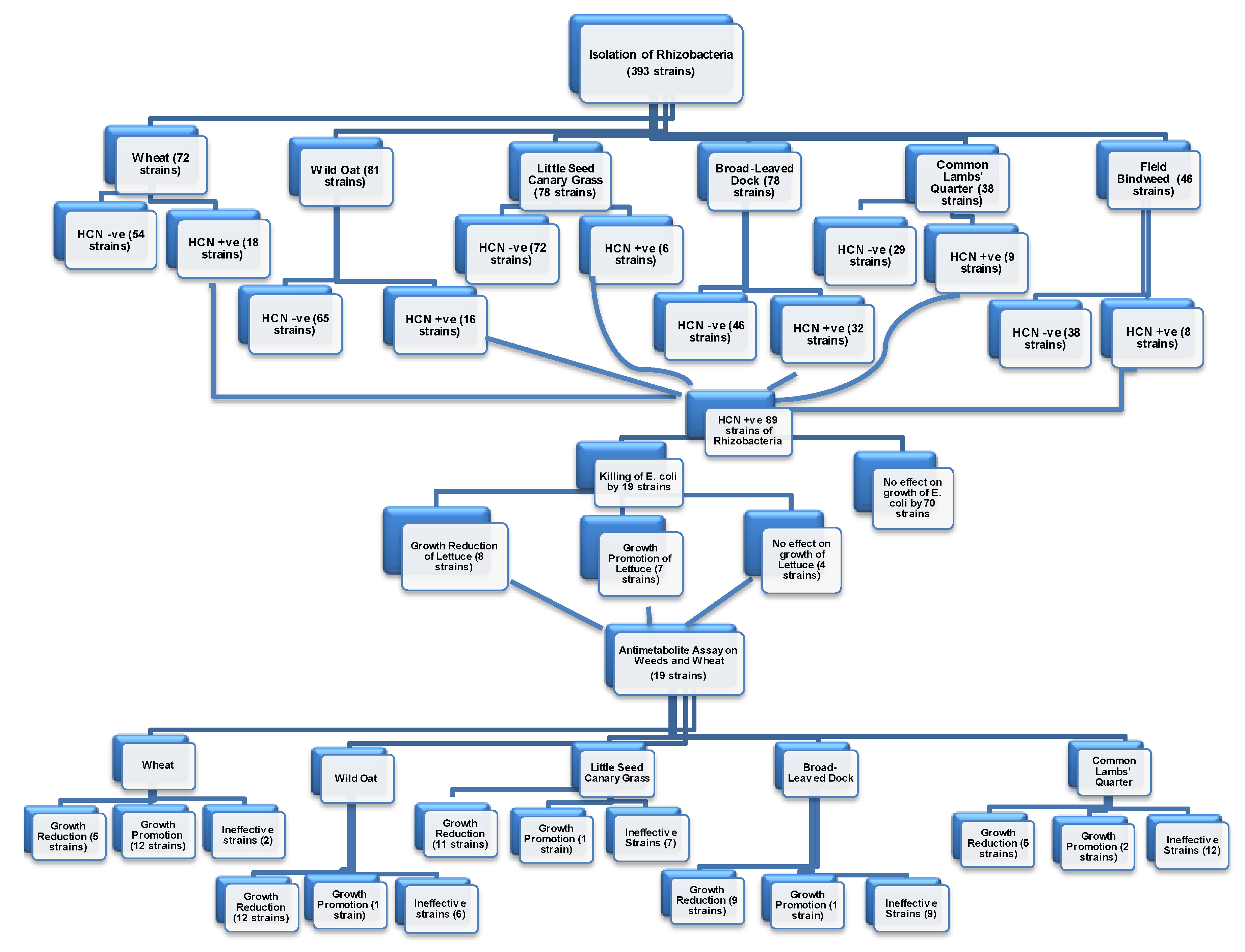
3.1. Isolation of Rhizobacteria
3.2. Production of HCN by Rhizobacteria
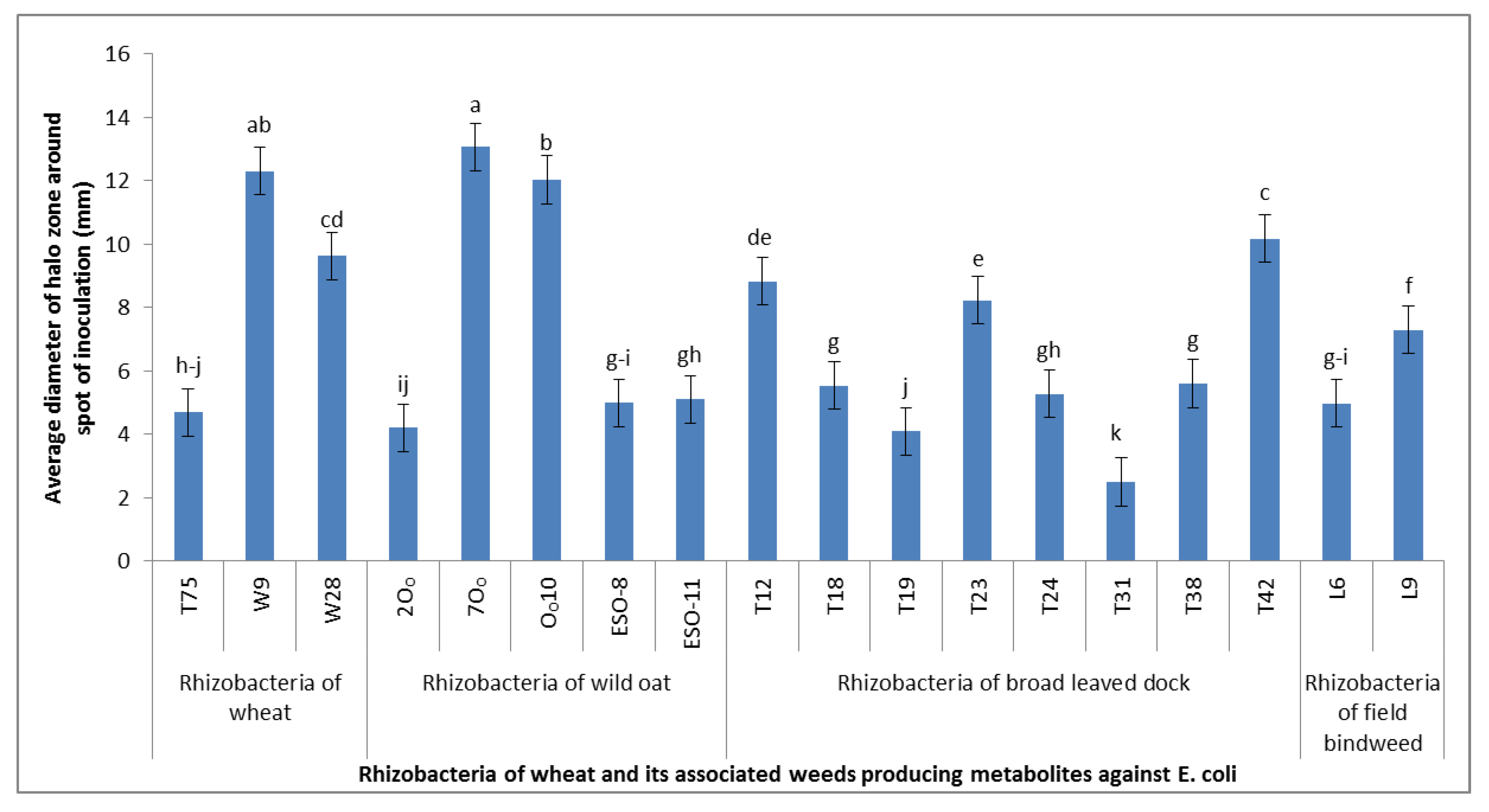
3.3. Antimetabolite Assay on E. coli
3.4. Antimetabolite Assay on Lettuce Seedlings
3.5. Antimetabolite Assay on Broad-Leaved Dock
3.6. Antimetabolite Assay on Wild Oat
3.7. Antimetabolite Assay on Little Seed Canary Grass
3.8. Antimetabolite Assay on Common Lambs’ Quarter
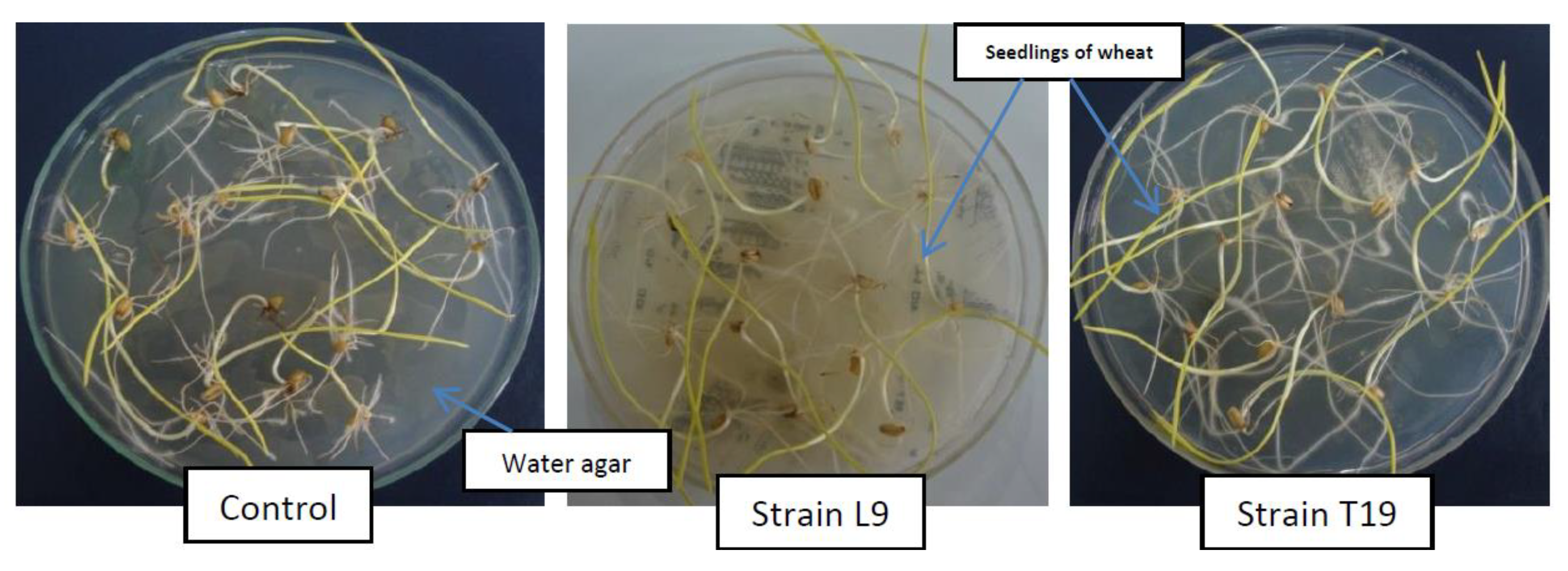
3.9. Antimetabolite Assay on Wheat
3.10. Cluster Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anonymous. Pakistan Economic Survey; Economic Adviser, Government of Pakistan, Ministry of Finance: Rawalpindi, Pakistan, 1966. Available online: http://www.irispunjab.gov.pk/StatisticalReport/Pakistan%20Economic%20Surveys/Economic%20Survey%201996-97.pdf (accessed on 24 September 2020).
- Anonymous. Economic Survey of Pakistan 2015–16; Economic Advisor’s Wing, Finance Division, Government of Pakistan: Islamabad, Pakistan, 2016. Available online: http://www.finance.gov.pk/survey_1516.html (accessed on 24 September 2020).
- Oerke, C.E. Centenary review on crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Shad, R.A. Weeds and weed control. In Crop Production; Nazir, S., Bashir, E., Bantel, R., Eds.; National Book Foundation: Islamabad, Pakistan, 2015; pp. 175–204. [Google Scholar]
- Ghorbani, R.; Leifert, C.; Seel, W. Biological control of weeds with antagonistic plant pathogens. Adv. Agron. 2005, 86, 191–225. [Google Scholar]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Kupper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef] [PubMed]
- Quimby, P.C.; King, L.R.; Grey, W.E. Biological control as a means of enhancing the sustainability of crop/land management systems. Agric. Ecosyst. Environ. 2002, 88, 147–152. [Google Scholar] [CrossRef]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tschantke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Pimentel, D. Environmental and economic costs of the application of pesticides primarily in the United States. Environ. Dev. Sustain. 2005, 7, 229–252. [Google Scholar] [CrossRef]
- Alavanja, M.C.R.; Hoppin, J.A.; Kamel, F. Health effects of chronic pesticide exposure: Cancer and neurotoxicity. Annu. Rev. Public Health 2004, 25, 155–197. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, Z.A.; Naveed, M.; Kremer, R.J. Limitations of existing weed control practices necessitate the development of alternative techniques based on biological approaches. Adv. Agron. 2017, 147, 239–280. [Google Scholar]
- Quimby, P.C.; Birdsall, J.L. Fungal agents for biological control of weeds: Classical and augmentative approaches. In Novel Approaches to Integrated Pest Management; Reuveni, R., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 293–308. [Google Scholar]
- Boyette, C.D.; Hoagland, R.E. Bioherbicidal potential of a strain of Xamthomonas spp. for control of common cocklebur (Xanthium strumarium). Biocontrol. Sci. Technol. 2013, 23, 183–196. [Google Scholar] [CrossRef]
- Coombs, E.M.; Clark, J.K.; Piper, G.L.; Cofrancesco, A.F., Jr. Biological Control of Invasive Plants in the United States; Oregon State University Press: Corvallis, OR, USA, 2004. [Google Scholar]
- Denslow, J.S.; D’Antonio, C.M. After biocontrol: Assessing indirect effects of insect releases. Biol. Control 2005, 35, 307–318. [Google Scholar] [CrossRef]
- Farooq, M.; Bajwa, A.A.; Cheema, S.A.; Cheema, Z.A. Application of allelopathy in crop production. Int. J. Agric. Biol. 2013, 15, 1367–1378. [Google Scholar]
- Kobayashi, K. Factors affecting phytotoxic activity of allelochemicals in soil. Weed Biol. Manag. 2004, 4, 1–7. [Google Scholar] [CrossRef]
- Kremer, R.J. The role of allelopathic bacteria in weed management. In Allelochemicals: Biological Control of Plant Pathogens and Diseases; Inderjit, M.K.G., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 143–156. [Google Scholar]
- Zeller, S.L.; Brandl, H.; Schmid, B. Host-plant selectivity of rhizobacteria in a crop/weed model system. PLoS ONE 2007, 2, 846–858. [Google Scholar] [CrossRef]
- Kennedy, A.C.; Johnson, B.N.; Stubbs, T.L. Host range of a deleterious rhizobacterium for biological control of downy brome. Weed Sci. 2001, 49, 792–797. [Google Scholar] [CrossRef]
- King, E.; Ward, M.; Raney, D. Two simple media for the demonstration of pycyanin and Xuorescein. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Bakker, A.W.; Schipper, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp. mediated plant growth stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Gasson, M.J. Indicator technique for antimetabolic toxin production by phytopathogenic species of Pseudomonas. Appl. Environ. Microbiol. 1980, 39, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, R.J. Interactions between the plants and microorganisms. Allelopath. J. 2013, 31, 51–70. [Google Scholar]
- Abd-Alla, M.H.; Morsy, F.M.; El-Enany, A.E.; Ohyama, T. Isolation and characterization of a heavy metal resistant isolate of Rhizobium leguminosarum bv. viciae potentially applicable for biosorption of Cd+2 and Co+2. Int. Biodeterior. Biodegrad. 2012, 67, 48–55. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dicky, D.A. Principles and Procedures of Statistics—A Biometrical Approach, 3rd ed.; McGraw Hill Book International Co.: Singapore, 1997. [Google Scholar]
- Matloob, A.; Safdar, M.E.; Abbas, T.; Aslam, F.; Khaliq, A.; Tanveer, A.; Rehman, A.; Chadhar, A.R. Challenges and prospects for weed management in Pakistan: A review. Crop Prot. 2020, 134, 104724. [Google Scholar] [CrossRef]
- Schippers, B.; Bakker, A.W.; Bakker, P.A.H.M. Interactions of deleterious and beneficial rhizosphere microorganisms and the effect of cropping practices. Annu. Rev. Phytopathol. 1987, 25, 339–358. [Google Scholar] [CrossRef]
- Kremer, R.J.; Souissi, T. Cyanide production of rhizobacteria and potential for suppression of weed seedling growth. Curr. Microbiol. 2001, 43, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Kremer, R.J.; Begonia, M.F.T.; Stanley, L.; Lanham, E.T. Characterization of rhizobacteria associated with weed seedlings. Appl. Environ. Microbiol. 1990, 56, 1649–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, R.J.; Kennedy, A.C. Rhizobacteria as biocontrol agents of weeds. Weed Technol. 1996, 10, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Zermane, N.; Souissi, T.; Kroschel, J.; Sikora, R. Biocontrol of broomrape (Orobanche crenata Forsk. and Orobanche foetida Poir.) by Pseudomonas fluorescens isolate Bf7-9 from the faba bean rhizosphere. Biocontrol Sci. Technol. 2007, 17, 483–497. [Google Scholar] [CrossRef]
- Weiland, G.W.; Neumann, R.; Backhaus, H. Variation of microbial communities in soil, rhizosphere and rhizoplane in response to crop species, soil type and crop development. Appl. Environ. Microbiol. 2001, 67, 5849–5854. [Google Scholar] [CrossRef] [Green Version]
- Souissi, T.; Kremer, R.J. A rapid microplate callus bioassay for assessment of rhizobacteria for biocontrol of leafy spurge (Euphorbia esula L.). Biocontrol Sci. Technol. 1998, 8, 83–92. [Google Scholar] [CrossRef]
- Owen, A.; Zdor, R. Effect of cyanogenic rhizobacteria on the growth of velvetleaf (Abutilon theophrasti) and corn (Zea mays L.) in autoclaved soil and the influence of supplemental glycine. Soil Biol. Biochem. 2001, 33, 801–809. [Google Scholar] [CrossRef]
- Abbas, T.; Zahir, Z.A.; Naveed, M. Bioherbicidal activity of allelopathic bacteria against weeds associated with wheat and their effects on growth of wheat under axenic conditions. BioControl 2017, 62, 719–730. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Rhizosphere of | Total Strains | |||||
|---|---|---|---|---|---|---|---|
| Wheat | Broad-Leaved Dock | Wild Oat | Little Seed Canary Grass | Field Bindweed | Common Lambs’ Quarter | ||
| Non-cyanogenic strains | 54 | 46 | 65 | 72 | 38 | 29 | 304 |
| Low cyanide activity strains | 8 | 3 | 8 | 5 | 3 | 6 | 33 |
| Medium cyanide activity strains | 6 | 12 | 3 | 0 | 2 | 2 | 25 |
| High cyanide activity strains | 2 | 14 | 0 | 1 | 2 | 1 | 20 |
| Very high cyanide activity strains | 2 | 3 | 5 | 0 | 1 | 0 | 11 |
| Total strains | 72 | 78 | 81 | 78 | 46 | 38 | 393 |
| Treatments | Root Length (cm) | Shoot Length (cm) | Dry Matter (mg) |
|---|---|---|---|
| Control | 5.08 ± 0.14 d,e | 4.01 ± 0.13 f,g | 51.49 ± 0.006 f,g |
| T12 | 4.07 ± 0.16 g,h | 3.19 ± 0.10 i | 38.94 ± 0.006 i,j |
| T18 | 3.79 ± 0.16 h,i | 3.32 ± 0.09 i | 41.81 ± 0.006 h,i,j |
| T19 | 7.1 ± 0.13 b | 5.39 ± 0.03 b,c | 67.13 ± 0.006 a,b |
| T23 | 6.08 ± 0.12 c | 5.22 ± 0.13 c | 61.7 ± 0.010 b,c,d |
| T24 | 5.92 ± 0.27 c | 5.07 ± 0.15 c | 59.57 ± 0.007 c,d,e |
| T31 | 4.36 ± 0.20 f,g | 3.68 ± 0.12 g,h | 44.2 ± 0.007 h,i |
| T38 | 4.06 ± 0.16 g,h | 3.37 ± 0.08 h,i | 42.07 ± 0.007 hij |
| T42 | 6.35 ± 0.22 c | 4.63 ± 0.15 d | 58.42 ± 0.007 d,e |
| T75 | 4.8 ± 0.26 e,f | 4.02 ± 0.15 f | 50.7 ± 0.003 f,g |
| 2O0 | 6.19 ± 0.20 c | 5.16 ± 0.14 c | 65.26 ± 0.007 b,c |
| 7O0 | 8.19 ± 0.17 a | 5.76 ± 0.11 a | 72.19 ± 0.006 a |
| O010 | 3.4 ± 0.14 i,j | 3.18 ± 0.09 i | 39.78 ± 0.006 hij |
| ESO-8 | 5.37 ± 0.20 d | 4.43 ± 0.17 d | 55.0 ± 0.003 e,f |
| ESO-11 | 5.03 ± 0.25 d,e | 4.06 ± 0.11 e,f | 52.0 ± 0.007 f,g |
| L6 | 4.48 ± 0.13 f,g | 3.79 ± 0.11 f,g | 45.79 ± 0.003 g,h |
| L9 | 8.01 ± 0.19 a | 5.73 ± 0.05 a,b | 72.88 ± 0.007 a |
| B11 | 5.26 ± 0.11 d,e | 4.38 ± 0.18 d,e | 54.53 ± 0.006 e,f |
| W9 | 3.34 ± 0.12 i,j | 3.11 ± 0.08 i | 35.99 ± 0.003 j |
| W28 | 3.231 ± 0.19 j | 3.04 ± 0.14 i | 38.08 ± 0.003 i,j |
| LSD | 0.517 | 0.345 | 6.42 |
| Treatment | Broad-Leaved Dock | Wild Oat | ||||
|---|---|---|---|---|---|---|
| Germination Rate (%) | Root Length (cm) | Dry Matter (g) | Germination Rate (%) | Root Length (cm) | Dry Matter (g) | |
| Control | 75.0 ± 0.58 b,c | 3.52 ± 0.13 b | 0.307 ± 0.014 a,b,c | 73.3 ± 0.33 b,c | 6.0 ± 0.16 b,c,d | 0.32 ± 0.03 b,c,d |
| T12 | 73.4 ± 0.88 b,c | 3.48 ± 0.12 b | 0.29 ± 0.035 a,b,c,d | 62.7 ± 0.88 d,e,f | 5.6 ± 0.17 c,d,e | 0.29 ± 0.02 b,c,d |
| T18 | 80.0 ± 1.00 a,b | 3.5 ± 0.10 b | 0.303 ± 0.026 a,b,c | 58.7 ± 0.67 f,g | 5.52 ± 0.24 d,e | 0.28 ± 0.02 c,d |
| T19 | 63.4 ± 0.67 d,e | 2.98 ± 0.18 c,d | 0.247 ± 0.013 c,d,e | 84.0 ± 0.58 a | 6.79 ± 0.13 a | 0.41 ± 0.03 a |
| T23 | 68.3 ± 0.33 c,d | 3.33 ± 0.11 b,c | 0.277 ± 0.018 b,c,d | 66.7 ± 0.33 c,d,e | 6.16 ± 0.16 b | 0.30 ± 0.02 b,c,d |
| T24 | 71.6 ± 0.33 b,c,d | 3.46 ± 0.11 b | 0.297 ± 0.023 a,b,cd | 68.0 ± 0.58 c,d | 5.28 ± 0.14 e,f | 0.29 ± 0.02 b,c,d |
| T31 | 86.6 ± 0.33 a | 4.1 ± 0.23 a | 0.353 ± 0.014 a | 73.3 ± 0.33 b,c | 5.75 ± 0.15 b,c,d,e | 0.32 ± 0.02 b,c,d |
| T38 | 55.0 ± 0.58 e,f | 2.68 ± 0.28 d,e | 0.233 ± 0.017 d,e,f | 77.3 ± 0.33 a,b | 5.96 ± 0.08 b,c,d | 0.33 ± 0.02 b,c |
| T42 | 33.3 ± 0.67 i,j | 1.73 ± 0.29 h,i | 0.13 ± 0.020 h,i | 32.0 ± 0.58 k | 2.74 ± 0.18 j | 0.11 ± 0.01 h,i |
| T75 | 70.0 ± 1.00 c,d | 3.33 ± 0.06 b,c | 0.28 ± 0.036 b,c,d | 60.0 ± 1.00 e,f,g | 5.88 ± 0.33 b,c,d | 0.28 ± 0.02 b,c,d |
| 2O0 | 73.4 ± 0.33 b,c | 3.61 ± 0.14 b | 0.303 ± 0.022 a,b,c | 54.7 ± 0.88 g,h | 4.55 ± 0.18 g | 0.20 ± 0.01 e,f |
| 7O0 | 41.7 ± 0.33 h,i | 1.96 ± 0.06 g,h | 0.17 ± 0.023 f,g,h | 49.3 ± 0.67 h,i | 3.77 ± 0.22 h,i | 0.16 ± 0.003 f,g,h |
| O010 | 51.6 ± 0.33 f,g | 2.17 ± 0.10 f,g | 0.203 ± 0.027 e,f,g | 45.3 ± 0.67 i,j | 3.6 ± 0.19 i | 0.15 ± 0.03 f,g,h |
| ESO-8 | 73.4 ± 0.67 b,c | 3.41 ± 0.21 b | 0.29 ± 0.026 a,c,d | 60.0 ± 0.58 e,f,g | 4.83 ± 0.18 f,g | 0.20 ± 0.01 f,g |
| ESO-11 | 48.4 ± 1.20 f,g,h | 2.53 ± 0.06 e,f | 0.203 ± 0.018 e,f,g | 54.7 ± 0.88 g,h | 5.56 ± 0.29 d,e | 0.26 ± 0.02 d,e |
| L6 | 56.6 ± 0.67 e,f | 2.77 ± 0.08 d,e | 0.24 ± 0.020 c,d,e | 82.7 ± 0.33 a | 6.13 ± 0.17 b,c | 0.35 0.03 a,b |
| L9 | 41.6 ± 0.67 h,i | 1.88 ± 0.10 g,h | 0.157 ± 0.022 g,h,i | 21.3 ± 0.33 l | 2.39 ± 0.23 j | 0.63 ± 0.01 i |
| B11 | 76.6 ± 0.67 b,c | 3.65 ± 0.11 b | 0.317 ± 0.033 a,b | 70.7 ± 0.33 b,c | 6.0 ± 0.19 b–d | 0.32 ± 0.02 b,c,d |
| W9 | 26.6 ± 0.67 j | 1.34 ± 0.12 i | 0.097 ± 0.018 i | 41.3 ± 0.33 j | 3.42 ± 0.13 i | 0.13 ± 0.03 g,h |
| W28 | 43.4 ± 0.88 g,h | 1.41 ± 0.08 i | 0.14 ± 0.029 g,h,i | 44.0 ± 1.16 i,j | 4.3 ± 0.13 g,h | 0.17 ± 0.02 f,g,h |
| LSD | 9.8205 | 0.429 | 0.068 | 7.32 | 0.545 | 0.0642 |
| Treatments | Little Seed Canary Grass | Common Lambs’ Quarter | ||||
|---|---|---|---|---|---|---|
| Germination Rate (%) | Root Length (cm) | Dry Matter (g) | Germination Rate (%) | Root Length (cm) | Dry Matter (g) | |
| Control | 73.3 ± 1.20 a,b | 4.59 ± 0.22 b,c | 0.283 ± 0.026 b,c | 63.3 ± 1.00 c,d,e | 2.87 ± 0.19 b,c,d,e | 0.27 ± 0.01 a,b,c |
| T12 | 54.7 ± 1.45 d,e,f,g | 4.11 ± 0.06 d,e | 0.287 ± 0.024 b,c | 61.0 ± 0.67 c,d,e | 2.57 ± 0.12 d,e,f,g | 0.25 ± 0.02 b,c,d |
| T18 | 60.0 ± 1.53 c,d,e,f | 3.68 ± 0.08 e,f | 0.267 ± 0.026 b,c,d | 61.5 ± 0.88 c,d,e | 2.29 ± 0.11 f,g,h | 0.24 ± 0.01 b,c,d |
| T19 | 81.3 ± 0.67 a | 5.07 ± 0.11 a | 0.35 ± 0.021 a | 63.2 ± 1.00 c,d,e | 2.50 ± 0.17 e,f,g,h | 0.27 ± 0.01 a,b,c |
| T23 | 66.7 ± 0.33 b,c | 4.64 ± 0.15 a,b | 0.26 ± 0.015 b,c,d | 58.7 ± 1.45 c,d,e | 2.73 ± 0.11 c,d,e,f | 0.25 ± 0.02 b,c,d |
| T24 | 68.0 ± 0 b,c | 4.28 ± 0.16 b,c,d | 0.237 ± 0.013 c,d,e | 66.7 ± 1.00 b,c | 3.14 ± 0.22 a,b,c | 0.29 ± 0.01 a,b |
| T31 | 74.7 ± 0.33 a,b | 4.32 ± 0.08 b,c,d | 0.287 ± 0.014 b,c | 57.7 ± 0.33 d,e,f | 2.9 ± 0.22 b,c,d,e | 0.28 ± 0.06 a,b |
| T38 | 74.7 ± 0.67 a,b | 4.58 ± 0.11 b,c | 0.277 ± 0.024 b,c | 57.2 ± 0.33 d,e,f | 2.61 ± 0.28 d,e,f | 0.24 ± 0.03 b,c,d |
| T42 | 54.7 ± 0.88 d,e,f,g | 2.89 ± 0.10 g,h | 0.153 ± 0.017 g,h,i | 65.7 ± 0.67 b,c,d | 3.04 ± 0.19 a,b,c,d | 0.24 ± 0.04 b,c,d |
| T75 | 53.3 ± 1.67 e,f,g,h | 2.34 ± 0.09 i | 0.175 ± 0.005 f,g,h | 47.7 ± 0.67 g | 2.04 ± 0.17 h,i | 0.19 ± 0.02 d,e,f |
| 2O0 | 64.0 ± 1.00 b,c,d,e | 4.1 ± 0.24 d,e | 0.223 ± 0.007 d,e,f | 75.6 ± 0.33 a | 3.51 ± 0.09 a | 0.32 ± 0.01 a |
| 7O0 | 42.7 ± 0.67 h,i | 2.37 ± 0.18 i | 0.100 ± 0.006 j | 55.7 ± 0.33 e,f,g | 2.90 ± 0.17 b,c,d,e | 0.24 ± 0.01 b,c,d,e |
| O010 | 34.7 ± 0.67 i | 1.8 ± 0.19 j | 0.087 ± 0.019 j | 63.3 ± 0.58 c,d,e | 2.93 ± 0.11 b,c,d,e | 0.24 ± 0.02 b,c,d |
| ESO-8 | 65.3 ± 0.88 b,c,d | 4.16 ± 0.21 c,d | 0.237 ± 0.012 c,d,e | 64.3 ± 0.67 b,c,d | 2.93 ± 0.27 b,c,d,e | 0.28 ± 0.02 a,b,c |
| ESO-11 | 48.0 ± 1.53 g,h | 3.48 ± 0.08 f | 0.237 ± 0.022 c,d,e | 49.0 ± 0.67 g | 1.69 ± 0.14 i,j | 0.17 ± 0.02 e,f |
| L6 | 80.0 ± 0.58 a | 4.57 ± 0.11 b,c | 0.293 ± 0.013 b | 72.3 ± 0.88 a,b | 3.32 ± 0.25 a,b | 0.30 ± 0.01 a,b |
| L9 | 49.3 ± 0.67 f,g,h | 2.54 ± 0.20 h,i | 0.123 ± 0.007 i,j | 63.3 ± 1.53 c,d,e | 2.69 ± 0.12 c,d,e,f | 0.27 ± 0.04 a,b,c |
| B11 | 74.7 ± 0.67 a,b | 4.52 ± 0.11 b,c,d | 0.257 ± 0.012 b,c,d | 49.0 ± 0.88 g | 2.09 ± 0.11 g,h,i | 0.21 ± 0.01 c,d,e |
| W9 | 58.7 ± 0.33 c,d,e,f,g | 3.26 ± 0.27 f,g | 0.197 ± 0.032 e,f,g | 59.0 ± 0.67 c,d,e | 2.98 ± 0.29 b,c,d,e | 0.26 ± 0.03 a,b,c,d |
| W28 | 49.3 ± 1.77 f,g,h | 1.67 ± 0.09 j | 0.137 ± 0.003 h,i,j | 50.0 ± 1.16 f,g | 1.29 ± 0.13 j | 0.13 ± 0.02 f |
| LSD | 11.432 | 0.445 | 0.0507 | 8.127 | 0.524 | 0.073 |
| Treatments | Germination Rate (%) | Root Length (cm) | Shoot Length (cm) | Dry Matter (g) |
|---|---|---|---|---|
| Control | 80.0 ± 0.58 d,e | 6.60 ± 0.50 c,d,e | 8.58 ± 0.22 c,d | 0.313 ± 0.018 f,g |
| T12 | 75.0 ± 0.58 e,f | 4.97 ± 0.08 f | 6.96 ± 0.27 e | 0.233 ± 0.003 h |
| T18 | 73.35 ± 0.33 f | 4.81 ± 0.10 f | 6.78 ± 0.16 e | 0.240 ± 0.015 h |
| T19 | 93.4 ± 0.33 a | 8.22 ± 0.15 b | 9.83 ± 0.37 b | 0.407 ± 0.012 c |
| T23 | 90.0 ± 0.58 a,b | 8.13 ± 0.14 b | 10.13 ± 0.34 b | 0.390 ± 0.006 c,d |
| T24 | 90.0 ± 0 a,b | 7.92 ± 0.13 b | 10.05 ± 0.21 b | 0.400 ± 0.015 c |
| T31 | 80.0 ± 0.58 d,e | 6.47 ± 0.23 d,e | 10.19 ± 0.20 b | 0.300 ± 0.020 g |
| T38 | 80.0 ± 0.58 d,e | 6.67 ± 0.27 c,d,e | 10.21 ± 0.17 b | 0.310 ± 0.015 f,g |
| T42 | 86.5 ± 0.33 b,c | 7.15 ± 0.25 c | 9.097 ± 0.38 c | 0.343 ± 0.012 e,f |
| T75 | 75.0 ± 0 e,f | 3.3 ± 0.17 g | 5.53 ± 0.10 f | 0.230 ± 0.012 h |
| 2O0 | 93.4 ± 0.33 a | 7.89 ± 0.26 b | 10.03 ± 0.18 b | 0.413 ± 0.003 b,c |
| 7O0 | 93.4 ± 0.33 a | 9.09 ± 0.21 a | 11.13 ± 0.25 a | 0.450 ± 0.010 a,b |
| O010 | 85 ± 0 b–d | 7.17 ± 0.15 c | 8.69 ± 0.19 c,d | 0.353 ± 0.003 d,e |
| ESO-8 | 81.5 ± 0.67 c,d | 6.53 ± 0.17 c–e | 8.52 ± 0.16 c,d | 0.347 ± 0.014 e,f |
| ESO-11 | 73.4 ± 0.67 f | 3.38 ± 0.33 g | 5.56 ± 0.15 f | 0.220 ± 0.006 h |
| L6 | 83.4 ± 0.33 c,d | 6.41 ± 0.29 e | 8.6 ± 0.14 c,d | 0.357 ± 0.019 d,e |
| L9 | 95 ± 0 a | 9.09 ± 0.12 a | 11.13 ± 0.20 a | 0.463 ± 0.020 a |
| B11 | 83.4 ± 0.67 c,d | 6.32 ± 0.23 e | 8.42 ± 0.17 d | 0.400 ± 0.006 c |
| W9 | 81.7 ± 0.67 c,d | 7.14 ± 0.31 c,d | 8.62 ± 0.38 c,d | 0.337 ± 0.014 e,f,g |
| W28 | 71.6 ± 0.67 f | 3.12 ± 0.26 g | 5.28 ± 0.17 f | 0.207 ± 0.014 h |
| LSD | 6.565 | 0.682 | 0.673 | 0.0376 |
| Category of Strains | Strain | Effects on Weeds and Wheat | ||
|---|---|---|---|---|
| Inhibition | Promotion | No Effect | ||
| Non-selective | ESO-11 | All the tested weeds and wheat | -- | -- |
| W28 | ||||
| Selective and inhibitory to wheat | T12 | Wheat, wild oat, and little seed canary grass | -- | Broad-leaved dock and common lambs’ quarter |
| T18 | Wheat, wild oat, little seed canary grass, and common lambs’ quarter | -- | Broad-leaved dock | |
| T75 | ||||
| Selective and non-inhibitory to wheat | T38 | Broad-leaved dock | -- | Wheat, wild oat, little seed canary grass, and common lambs’ quarter |
| ESO-8 | Wild oat | -- | Wheat, little seed canary grass, broad-leaved dock, and common lambs’ quarter | |
| W9 | Wild oat, little seed canary grass, and broad-leaved dock | -- | Wheat and common lambs’ quarter | |
| Selective and promotory to wheat | T19 | Broad-leaved dock | Wheat, wild oat, and little seed canary grass | Common lambs’ quarter |
| T24 | Wild oat | Wheat | Little seed canary grass, broad-leaved dock, and common lambs’ quarter | |
| T42 | Wild oat, little seed canary grass, and broad-leaved dock | Wheat | Common lambs’ quarter | |
| 7O0 | ||||
| O010 | ||||
| L9 | ||||
| 2O0 | Wild oat and little seed canary grass | Wheat and common lambs’ quarter | Broad-leaved dock | |
| L6 | Broad-leaved dock | Wheat, wild oat, and common lambs’ quarter | Little seed canary grass | |
| B11 | Common lambs’ quarter | Wheat | Wild oat, little seed canary grass, and broad-leaved dock | |
| -- | T23 | -- | Wheat | Wild oat, little seed canary grass, broad-leaved dock, and common lambs’ quarter |
| T31 | -- | Broad-leaved dock | Wheat, wild oat, little seed canary grass, and common lambs’ quarter | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, T.; Zahir, Z.A.; Naveed, M.; Abbas, S.; Alwahibi, M.S.; Elshikh, M.S.; Mustafa, A. Large Scale Screening of Rhizospheric Allelopathic Bacteria and Their Potential for the Biocontrol of Wheat-Associated Weeds. Agronomy 2020, 10, 1469. https://doi.org/10.3390/agronomy10101469
Abbas T, Zahir ZA, Naveed M, Abbas S, Alwahibi MS, Elshikh MS, Mustafa A. Large Scale Screening of Rhizospheric Allelopathic Bacteria and Their Potential for the Biocontrol of Wheat-Associated Weeds. Agronomy. 2020; 10(10):1469. https://doi.org/10.3390/agronomy10101469
Chicago/Turabian StyleAbbas, Tasawar, Zahir Ahmad Zahir, Muhammad Naveed, Sana Abbas, Mona S. Alwahibi, Mohamed Soliman Elshikh, and Adnan Mustafa. 2020. "Large Scale Screening of Rhizospheric Allelopathic Bacteria and Their Potential for the Biocontrol of Wheat-Associated Weeds" Agronomy 10, no. 10: 1469. https://doi.org/10.3390/agronomy10101469
APA StyleAbbas, T., Zahir, Z. A., Naveed, M., Abbas, S., Alwahibi, M. S., Elshikh, M. S., & Mustafa, A. (2020). Large Scale Screening of Rhizospheric Allelopathic Bacteria and Their Potential for the Biocontrol of Wheat-Associated Weeds. Agronomy, 10(10), 1469. https://doi.org/10.3390/agronomy10101469