3.2.1. Plant Growth
The statistical analysis of the data indicated a significant difference between the
Salvia hispanica L. and
Salvia columbariae Benth. varieties concerning the obtained plant height, number of branches and inflorescences, and the length of the central axis main inflorescence at harvest time (
p = 0.0005,
p = 0.0002,
p = 0.0145,
p = 0.0020) (
Table 4).
The morphological differences shown in
Table 4 can be attributed to the different growth habitus of the cultivated varieties as shown in
Figure 1. To state only some observed differences, which are in accordance with literature,
Salvia columbariae Benth., for example, grows 10 to 50 cm tall. It develops a tap rooted, rosetted, and unbranched or sporadically branched stem. The stems emerge singly or in several branches at the base and form terminal inflorescences, subtended by a few leaves; one or more spherical clusters, each containing one dozen to several dozen flowers reaching 8 to 20 mm in diameter, also appear [
38,
39].
Salvia hispanica L., on the other hand, reaches growth heights of up to 1.75 m, has shallow roots, and its stem is sparsely branched and forms terminal pseudo whorl with blue or white flowers which can be more than 20 cm long [
1].
Golden Chia produced significantly shorter plants (37.48 cm) in comparison to the other two
Salvia hispanica L. varieties, G8 (74.23 cm) and SALV66 (78.60 cm), due to the difference in growth habitus as already indicated (
Table 4). With a mean of 2.08 branches per plant, Golden Chia produced significantly less branches compared to G8 (7.77) and SALV66 (8.27), which were in the range (7.9–9.3) of what Souza and Chavez [
40] observed under greenhouse conditions in Brazil (
Table 4). [
22]. In regard to the number of branches developed, Sosa Baldivia and Ibarra [
41] pointed to the fact that
Salvia hispanica L. is subject to high plasticity as it is directly influenced by row spacing and seeding density, which has to be matched to the specific, given environmental conditions.
G8 (7.98) produced a significantly lower number of inflorescences compared to Golden Chia (29.42) and SALV66 (15.97), decreasing from Golden Chia > SALV66 > G8. The number of inflorescences produced by
Salvia hispanica L. varieties in 2016 cultivated at Ihinger Hof amounted to 11.8–13.3 [
22]. As for the number of branches, the number of developed inflorescences is most probably also subject to the plasticity of chia [
41]. G8 and SALV66 formed pseudo whorl inflorescences whereas Golden Chia developed compact, round inflorescences as shown in
Figure 1.
Therefore, the length of the inflorescence of the central axis of SALV66 (12.26) and G8 (10.40) was measured whereas the width of the central axis inflorescence of Golden Chia (2.76) was recorded. The length of the central axis inflorescence of
Salvia hispanica L. was in the range (7.92–14.82 cm) of what Hüller Goergen et al. [
42] observed during a field trial conducted in Brazil but was considerably shorter compared to the results (16.8–19.2 cm) obtained at Ihinger Hof in 2016 by Grimes et al. [
22]. The difference in length is most probably due to environmental influences and weather conditions, crop management, or a combination of those influences and specific species responses [
43]. To our knowledge, the above-mentioned data of Golden Chia were presented for the first time; therefore, it would be of great interest to examine different environmental and crop management effects in regard to the cultivation of Golden Chia under the given conditions in southwestern Germany.
3.2.2. Yield Traits
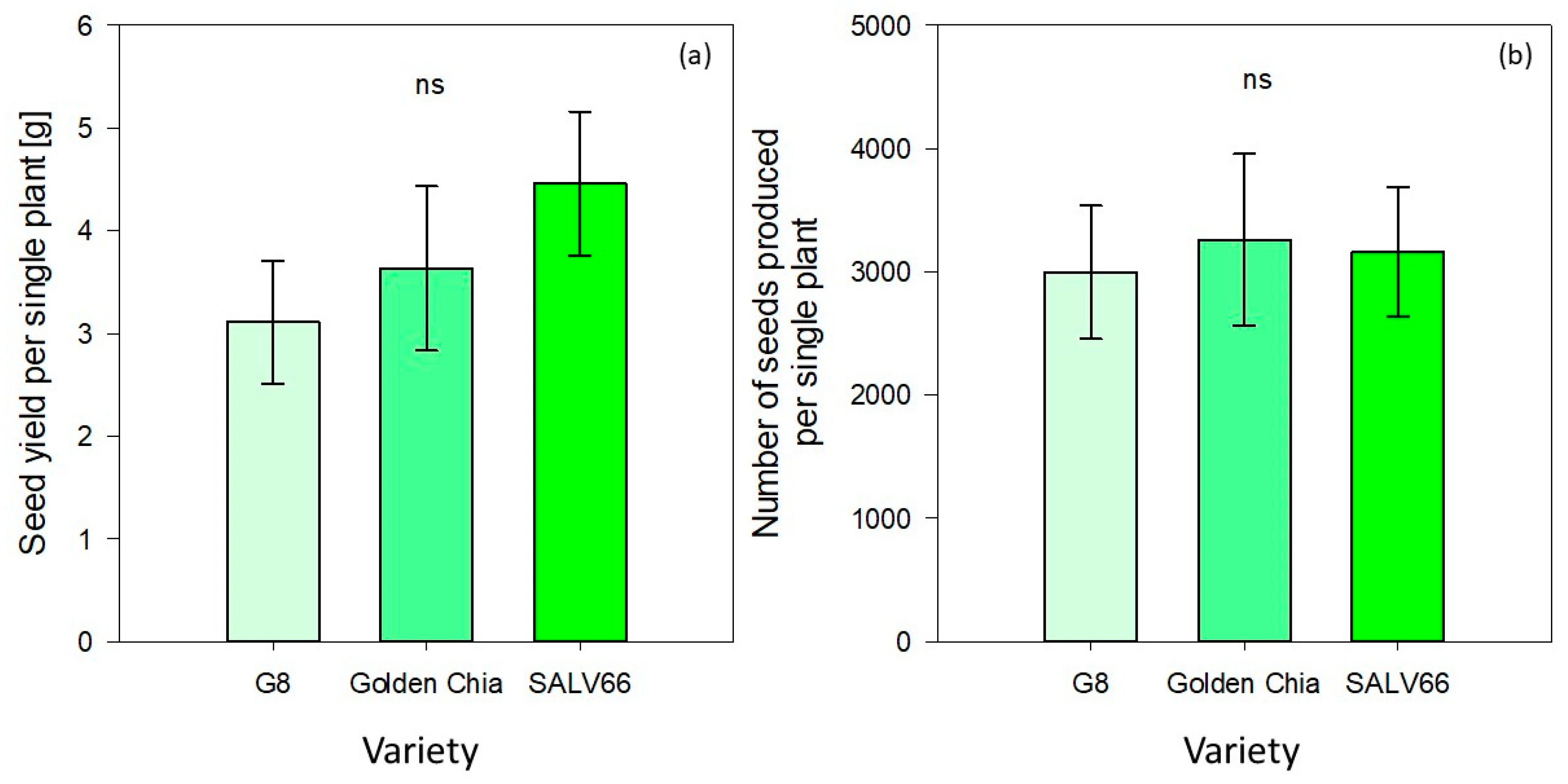
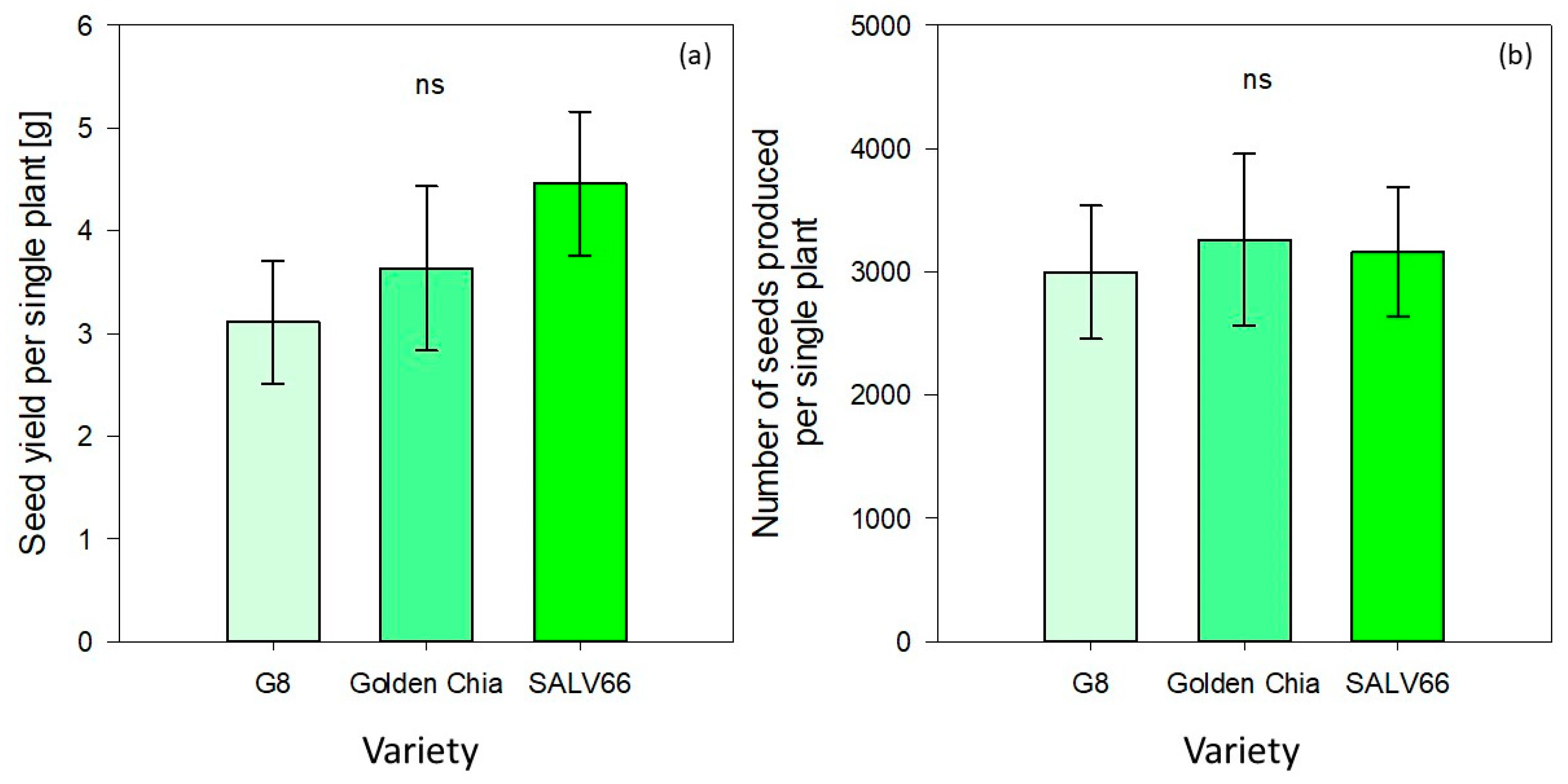
Seed weight per single plant was not significantly influenced by variety (
p > 0.05) (
Table 5). It ranged from 3.11 ± 0.6 (G8) to 3.63 ± 0.8 (Golden Chia) and 4.46 ± 0.7 (SALV66) respectively (
Figure 3). The same applied for the number of seeds produced per single plant, which varied from 2996 ± 540 (G8) to 3260 ± 695 (Golden Chia) and 3162 ± 525 (SALV66) (
Figure 3).
Reported manually obtained seed yields and amounts of seeds produced per single plant were in the range of 6.3 to 6.4 g and 4051 to 4289 for
Salvia hispanica L., respectively, being distinctly higher compared to the values obtained by the present study [
22]. It is, in general, stated that obtained seed yields per single plant and seeds produced per single plant in different plant species could be influenced by various factors, such as plant density, genotypic differences, climatic conditions, and mineral nitrogen content [
44,
45,
46,
47]. For both SALV66 and Golden Chia, it can be assumed that single plant yield and seeds produced were most likely predominantly influenced by the environmental conditions of the respective growing season. The assumption which was made in regard to the high plasticity of
Salvia hispanica L. enabling the compensation of lower plant densities with higher seed numbers per single plant could not be verified by the present study [
22,
41]. To our knowledge, this is the first time these specific yield traits of
Salvia columbariae Benth. were published. Further field trials would have to be carried out in order to make well-founded statements about the possible specific effects influencing seed yields and numbers of seeds produced per single plant.
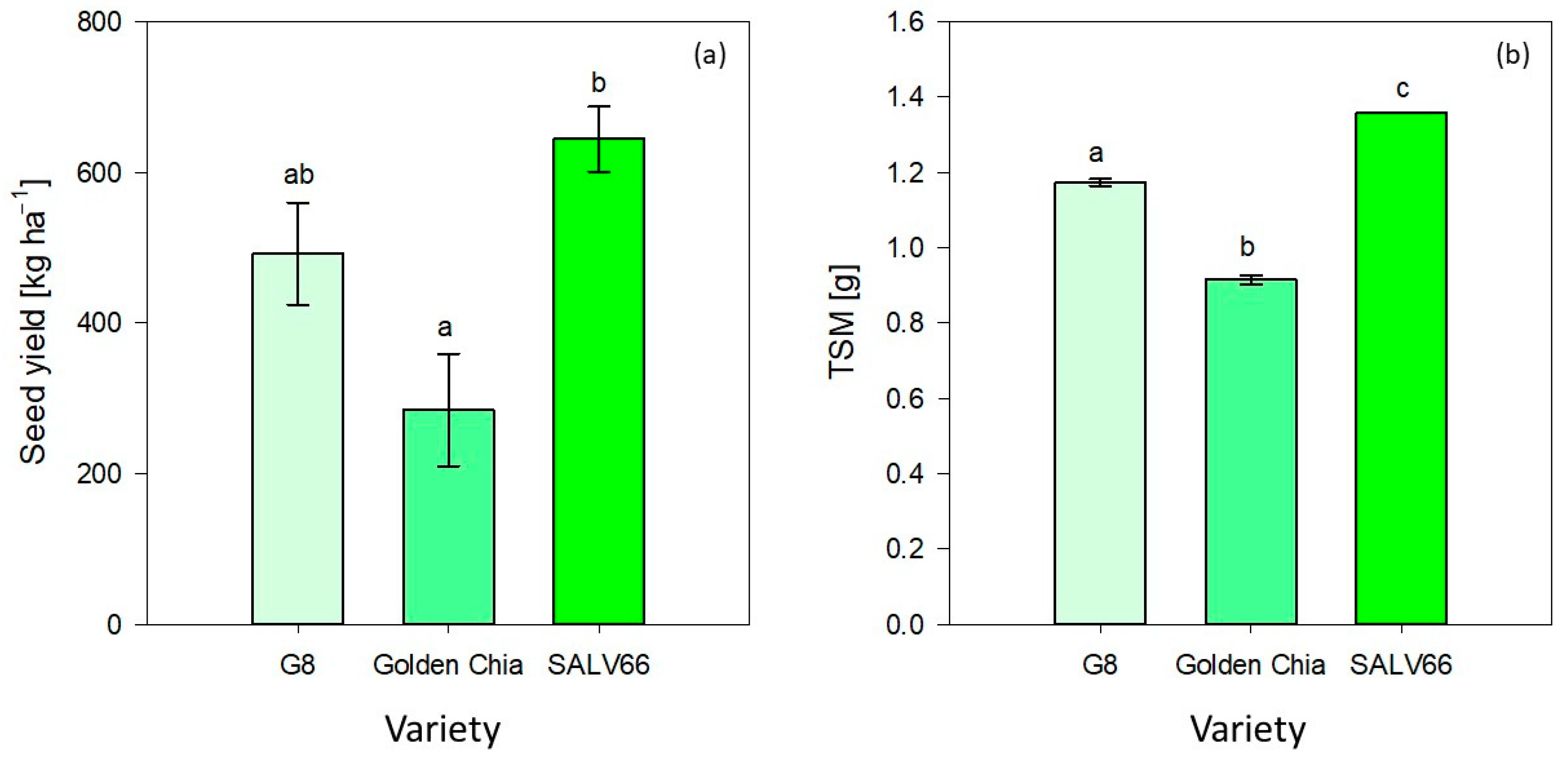
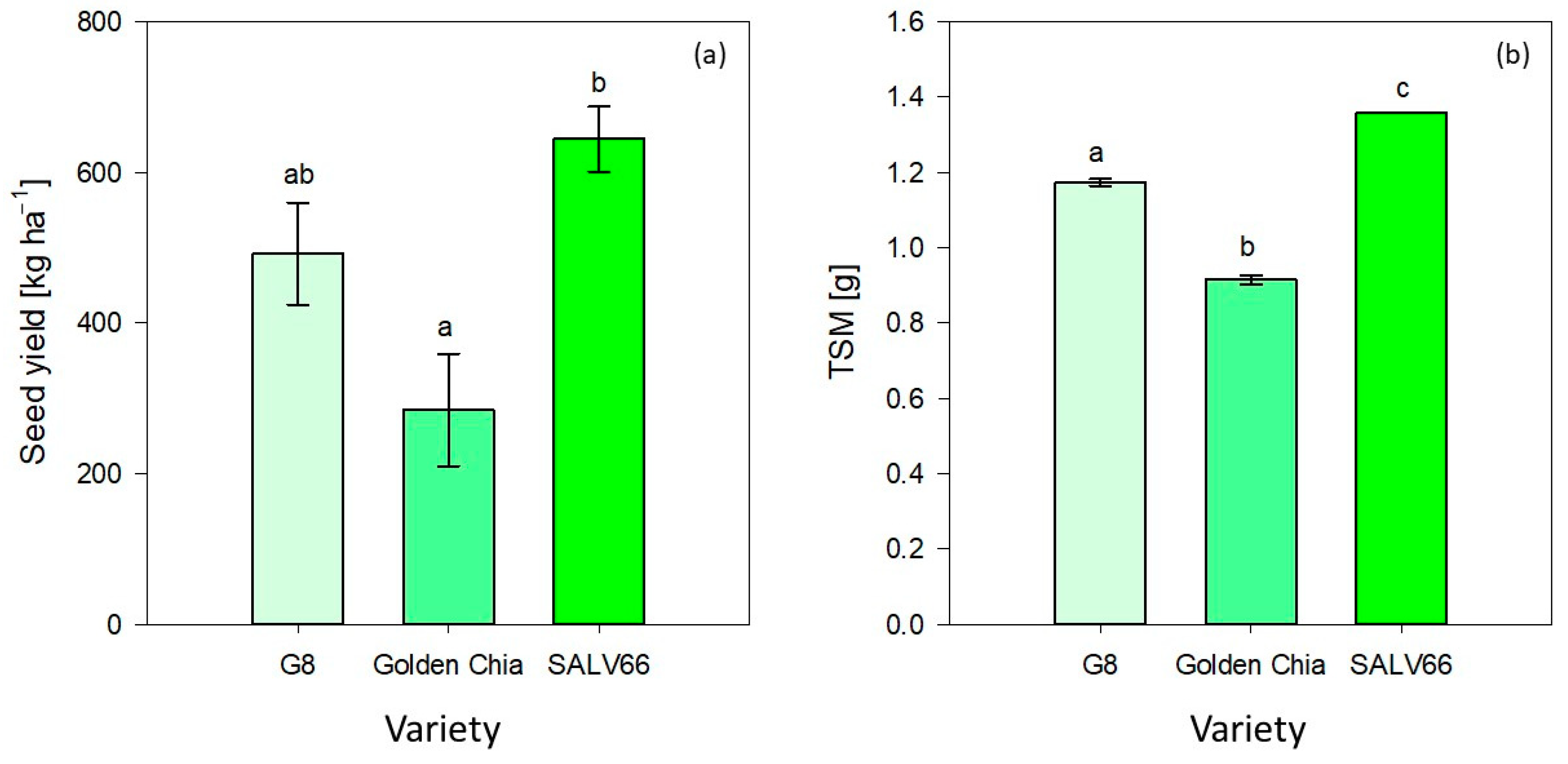
Golden Chia obtained a significantly lower mean yield (284.13 ± 74.44 kg ha
−1) compared to SALV66 (643.99 ± 42.92 kg ha
−1), whereas the obtained yield of G8 (491.91 ± 67.45 kg ha
−1) did not differ significantly from Golden Chia and SALV66, as shown in
Figure 4. As presented in
Table 5, the statistical analysis showed that the number of plants per m
2 (
p = 0.0372) and variety (
p = 0.0354) significantly influenced obtained yields (kg ha
−1). Mean number of plants per m
2 of the present study varied from 34 (G8) to 20 (Golden Chia) and 26 (SALV66), being roughly in the range of the plants per m
2 obtained in 2016 (17–30) and considerably lower compared to the obtained number of plants per m
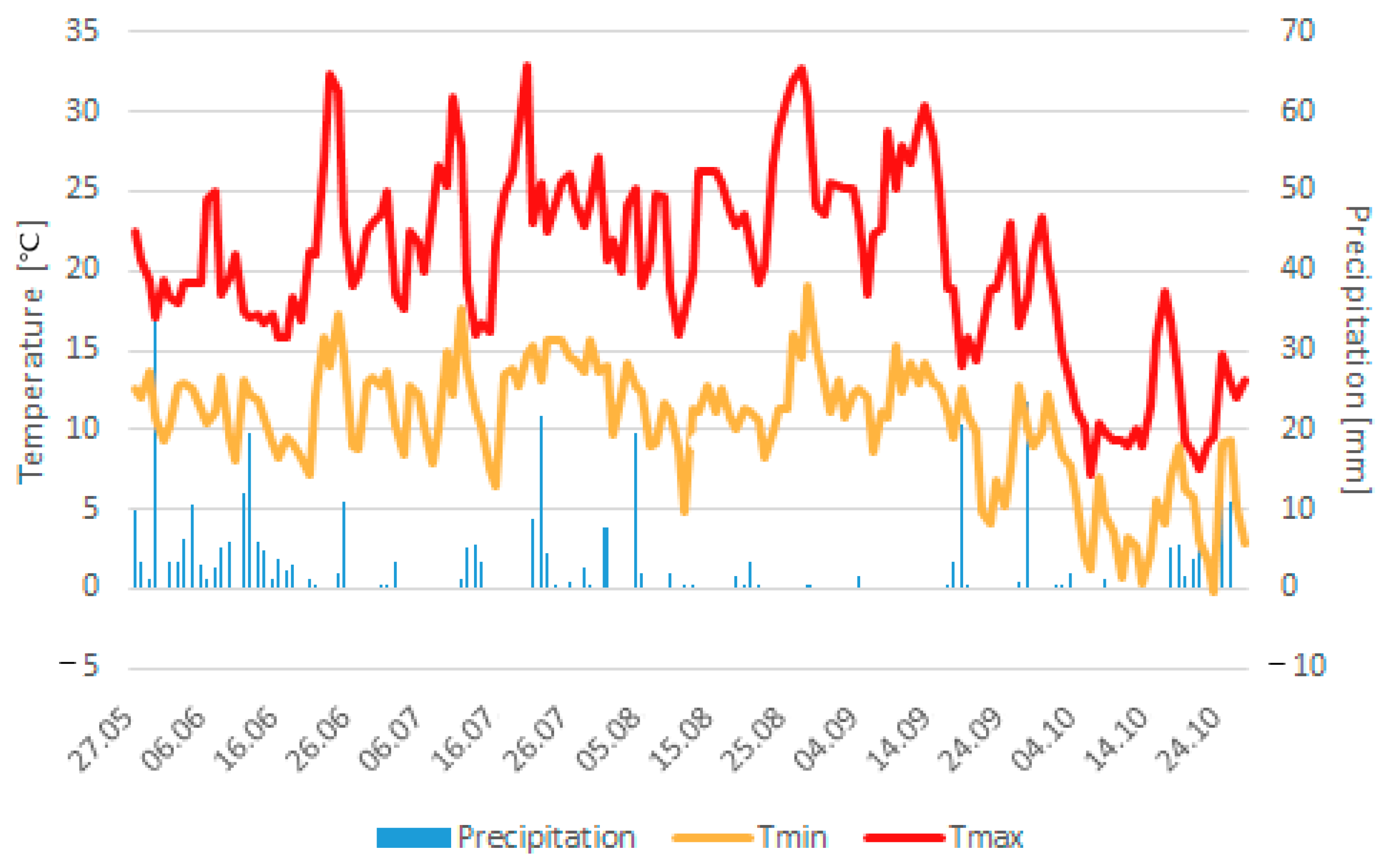
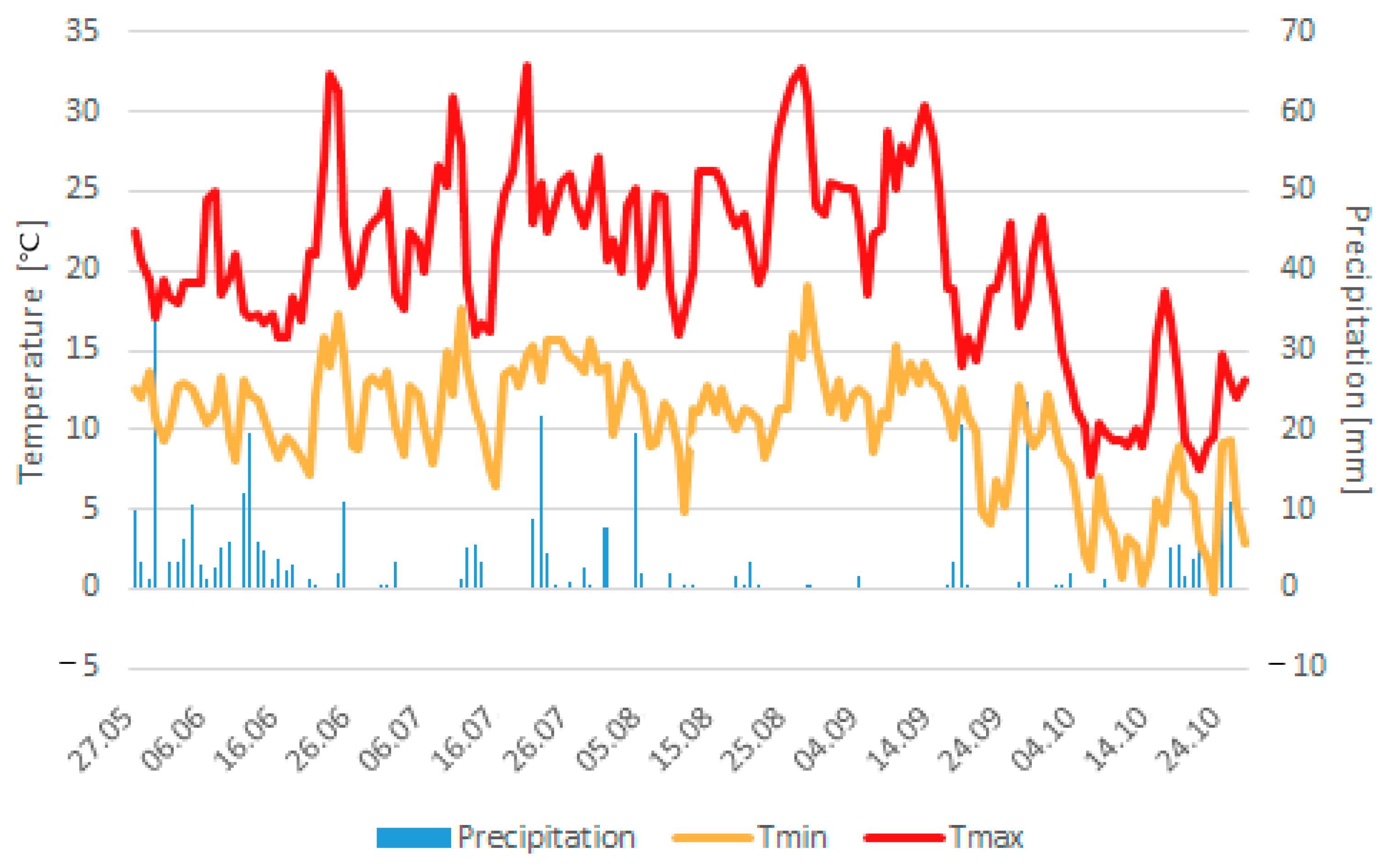
2 in 2017 (41–47) at Ihinger Hof. In this regard, it must be stated that three days after sowing, a storm with downpour (35 mm) took place, followed by a dry period (
Figure 2), leading to soil crusting and a partially irregular germination, which probably significantly influenced the obtained number of plants per m
2 and yields of the cultivated varieties [
48]. The myxocarpic characteristic of the
Salvia genus generally constitutes an advantage to seeds coping with restricted and irregular water availability during germination and early seedling development, as its mucilage may aid in supplying water to the seed [
49,
50]. However, the potential advantage of the mucilaginous chia seeds might have been overruled by the susceptibility of the seedlings, which could not penetrate the soil crust [
51].
In this present study, G8 obtained a noticeably lower seed yield compared to previously obtained seed yields of G8 at Ihinger Hof in 2016 and 2017 (752–1170 kg ha
−1) cultivated at the same sowing rate, row spacing, and nitrogen fertilization rate. The relatively low seed yields obtained by both of the
Salvia hispanica L. varieties in this study could be explained first and foremost due to unfavorable field conditions and the accelerated plant development resulting from them, as addressed in
Section 3.1 and shown in
Table 3. In this regard, Mathan et al. stated that flower induction, which marks the transition from vegetative to generative plant growth, is, inter alia, decisive for optimal yield in crop plants [
52]. The average mean temperature until flower induction was slightly higher (0.1–1.3 °C) compared to the previous studies conducted at Ihinger Hof in 2015 to 2017; furthermore, the average mean temperature from flower induction to harvest maturity was considerably higher (3–5.5 °C). Those high temperatures during the grain filling phase probably led to an increase in the photosynthesis rate above its optimum, leading to reduced metabolic activity during grain filling and maturation and, thus, to yield losses [
6,
16,
22]. Alongside the relatively high temperatures, precipitation was partly substantially lower (between 38–193 mm) compared to the studies conducted at Ihinger Hof between 2015 and 2017, which certainly contributed to the comparably low seed yields obtained. Even though it is stated that
Salvia hispanica L. is able to grow in dry conditions and is semi-tolerant to drought, this only applies to already established plant stocks [
35,
53,
54,
55]. It is indicated that
Salvia hispanica L. profits from precipitation and general water availability ranging from 300 to 1000 mm during the entire vegetation period, having a higher water demand during its vegetative phase and while establishing the reproductive organs, whereas drier conditions are required during the generative phase and seed maturity [
56,
57,
58]. In this regard, Steduto et al. stated two possible negative effects on yield in general: (1) the inhibition of water stress on pollination and successful formation of the embryo, which could have led to a reduced number of set fruits (or grains) and, therefore, limited harvest index and seed yield, respectively, and (2) the under-filling and abortion of younger fruits resulting from a lack of photosynthetic assimilates available to be used for fruit developing [
58].
In contrast to that, the seed yield of Golden Chia could, in comparison to the obtained seed yield reported by Ayerza and Coats (133 kg ha
−1), be considered as quite high. The disparity between the seed yield (g) produced per single plant and the seed yield per kg ha
−1 of Golden Chia and the significantly lower seed yield (kg ha
−1) compared to the other
Salvia hispanica L. varieties can be explained by the degree of domestication. Domestication of
Salvia hispanica L. began, according to Ayerza and Coats, more than 2000 years ago, whereas Golden Chia remains highly undomesticated to date [
1]. The high seed shattering level is a distinct attribute of the wild stage (lack of domestication), leading to enormous seed yield losses due to threshing [
34,
59]. Further breeding approaches towards reduced seed shattering should, therefore, be developed in order to be able to cultivate Golden Chia on an economically profitable basis. Based on our results, it can be stated that Golden Chia holds great potential, especially with regard to its photosensitive reaction, as flower induction and harvest maturity were reached significantly earlier, reducing the risk of frost damage due to late harvests. One general factor possibly leading to lower obtained seed yields is the choice of harvesting time and harvest intensity (manually/mechanically). It has been reported that sub-optimal combine operations and harsh weather conditions could lead to yield losses of up to 37% as chia is highly susceptible to seed shattering [
60,
61]. In this context, the non-uniform (top-down) maturation of
Salvia hispanica L. should be mentioned, posing an issue in regard to optimal harvest time as the central inflorescence matures and ripens while inflorescences on side branches are still immature [
23,
62]. The same applies to
Salvia columbariae Benth., as it could be observed that the flowers generally ripen from the inside out in addition to a delayed ripening of the side branches. In addition, the short vegetative period could, in this regard, negatively influence the potential seed yield of
Salvia columbariae Benth.
Thousand seed mass as another yield determining trait seemed to be influenced exclusively by variety (
p < 0.0001). As illustrated in
Figure 4, the TSM of G8 was 1.17 ± 0.01 g, while the TSM of Golden Chia was 0.92 ± 0.01 g, and SALV66 obtained a TSM of 1.36 ± 0.01 g. TSM differed significantly between each of the varieties. The TSMs for G8 and SALV66 were in the range of current literature (1.1–1.4 g) [
19,
22,
63]. In regard to obtained
Salvia hispanica L. TSMs, it could be shown that a high number of plants per m
2 leads to interplant competition and therefore to a decrease in TSM [
23]. Other studies related to wheat (
Triticum aestivum L.) showed that plant density and nitrogen fertilization influenced TSM significantly [
64,
65,
66]. Identifying factors which influence the TSM of the cultivated
Salvia varieties could be advantageous in order to further improve agronomic traits of the cultivated varieties in general and especially under the given climatic conditions in southwestern Germany.
With regard to Golden Chia, it should be mentioned that there might be a need to develop a specific cultivation system which differs in row distance, seeding density, and nitrogen fertilizer rates, as well as a specific harvest process from that established for
Salvia hispanica L. in order to exploit its full potential, leading to higher seed yields and TSM [
23].




{kind=link}
{kind=link}
{kind=link}
{kind=link}