Isolate-Dependent Inheritance of Resistance Against Pseudoperonospora cubensis in Cucumber

 ,
,
Abstract
1. Introduction
2. Methods and Materials
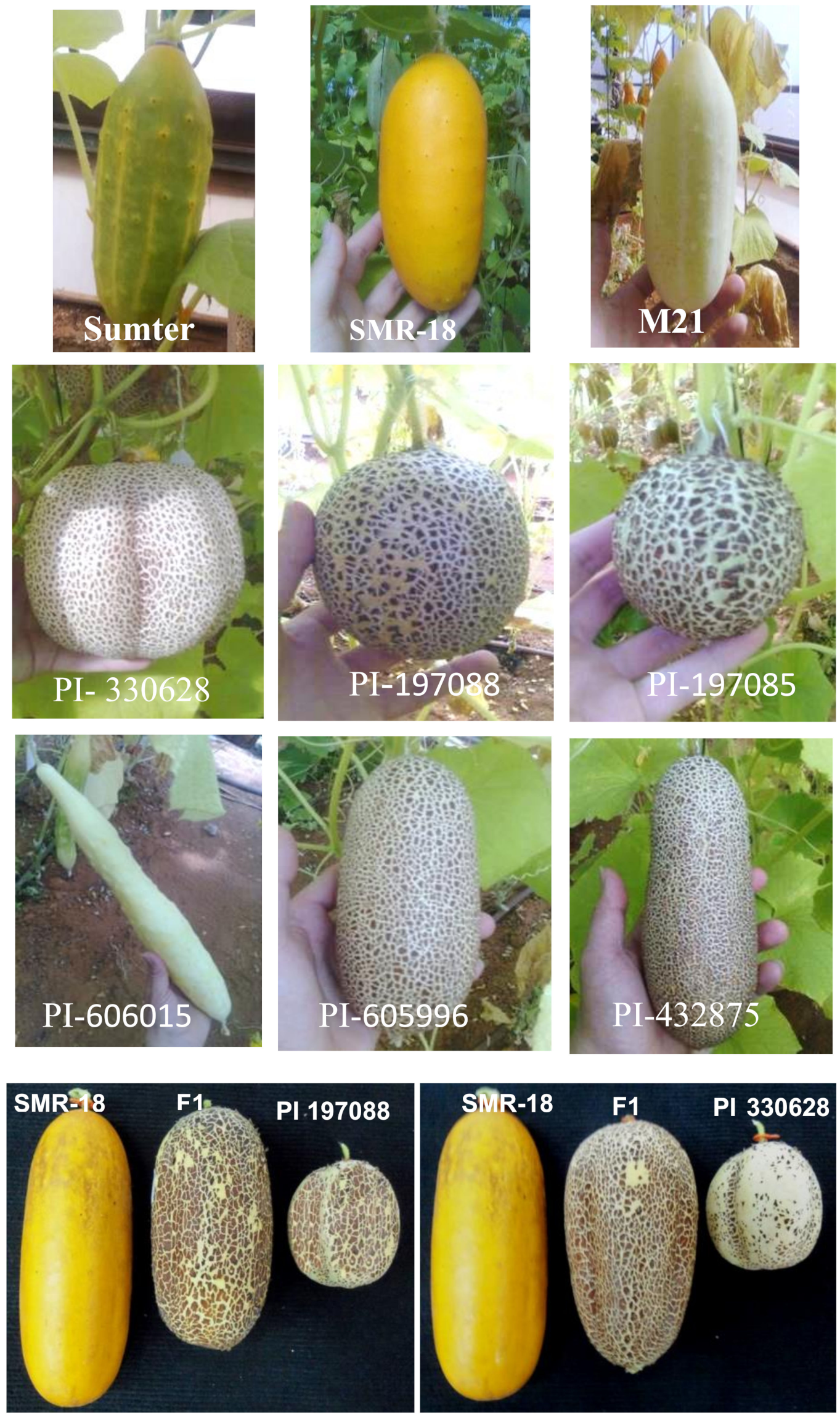
2.1. Cucumber Accessions
2.2. Pathogen
2.3. Crosses
2.4. Inoculation of Detached Leaves
2.5. Disease Assessment in Detached Leaves
2.6. Inheritance of Resistance in Detached Leaves Taken from Adult Plants
2.7. Inheritance of Resistance in Intact Field-Grown Plants
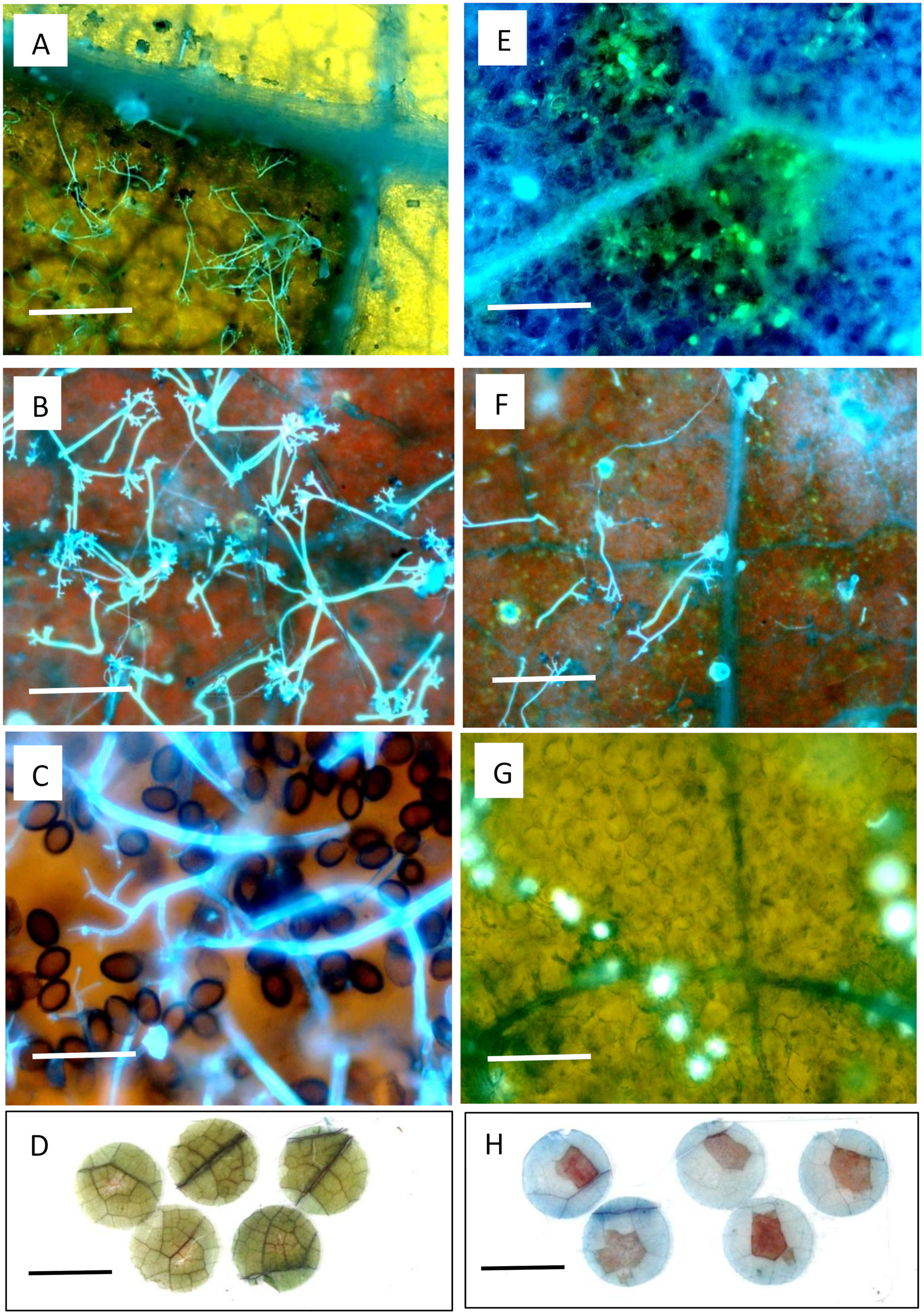
2.8. Microscopy
3. Results
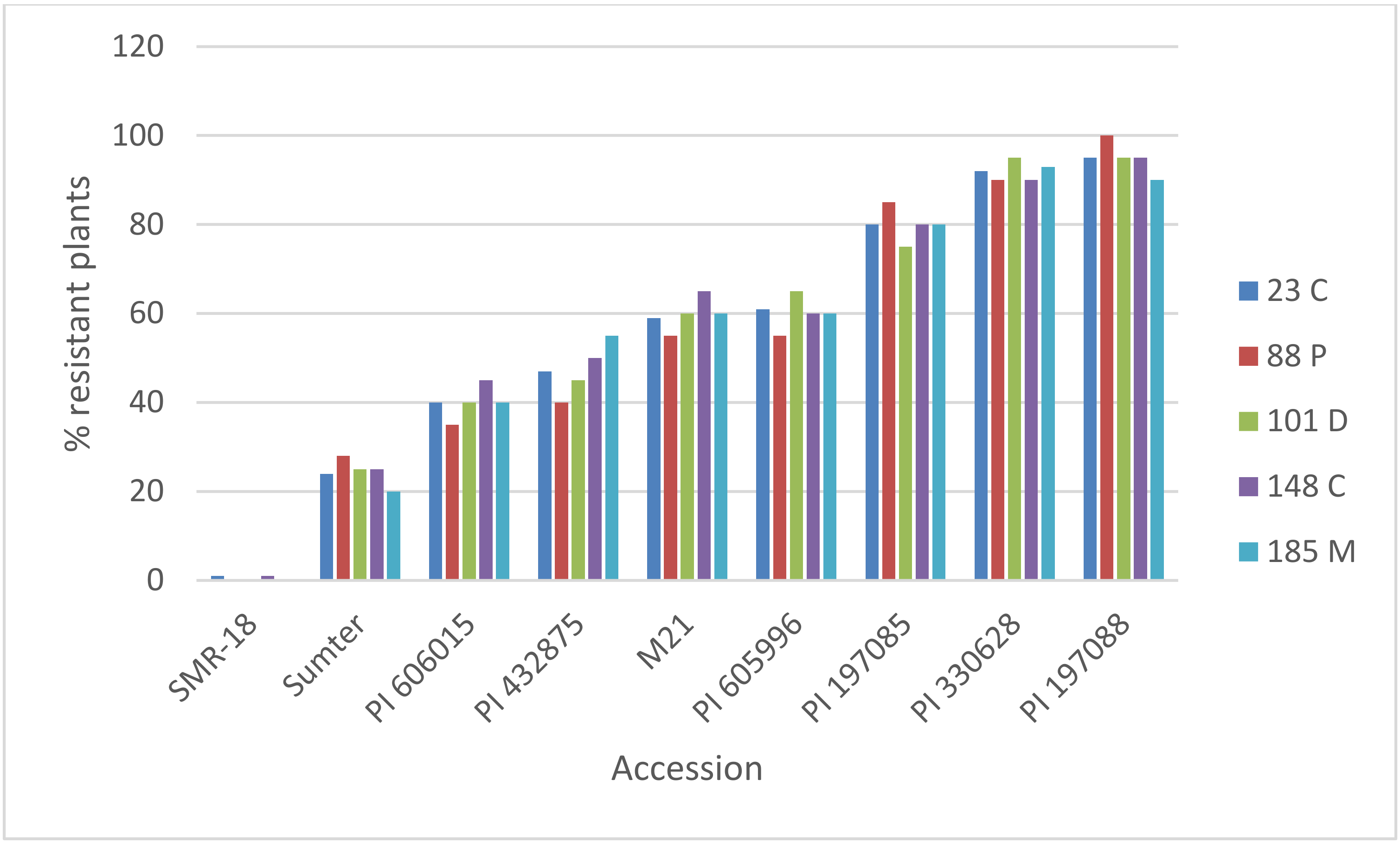
3.1. Resistance of C. sativus to Multiple Isolates of P. cubensis
3.2. Microscopy of Resistance in PI 197088 and PI 330628
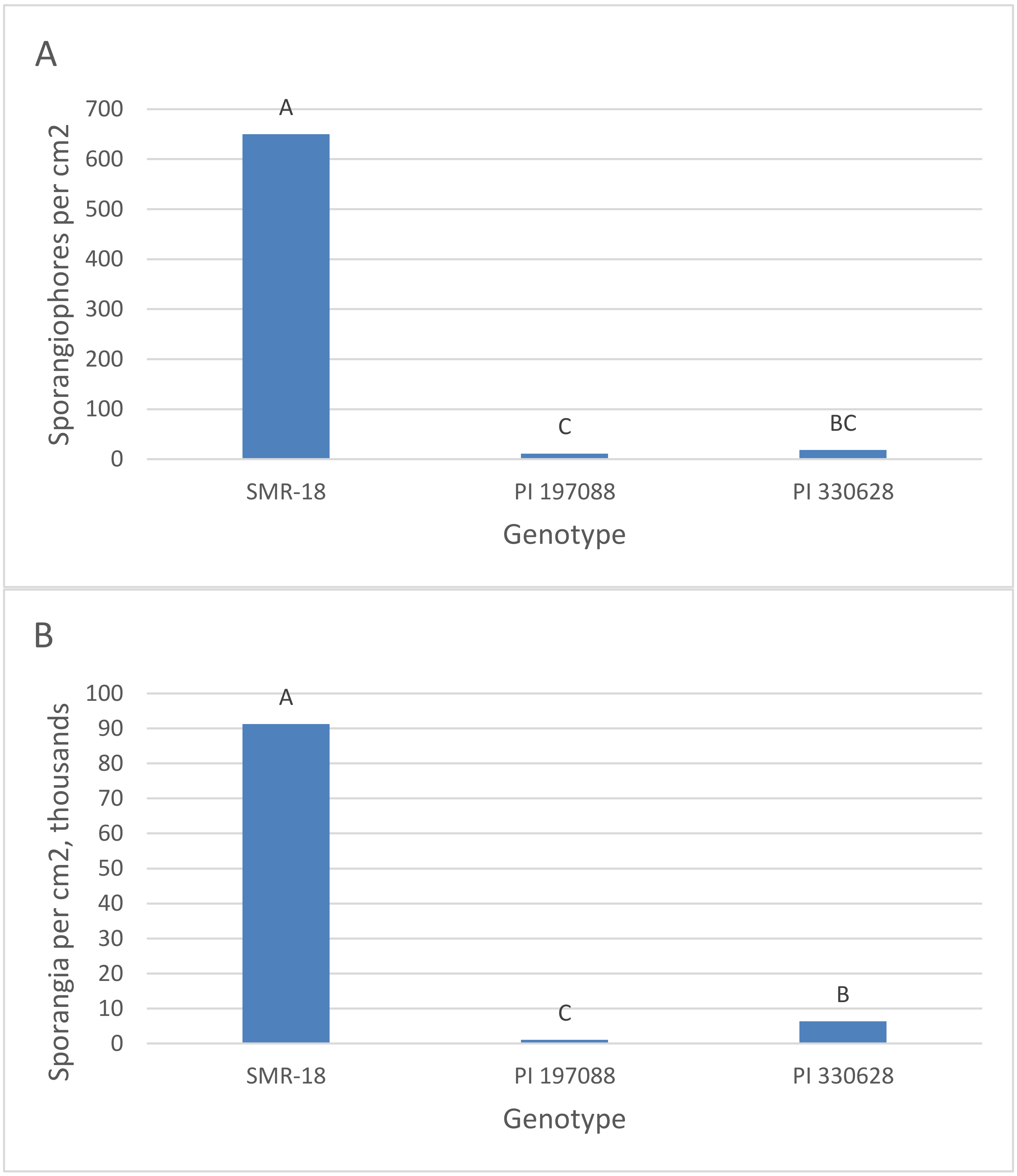
3.3. Quantification of Resistance in Adult Plants
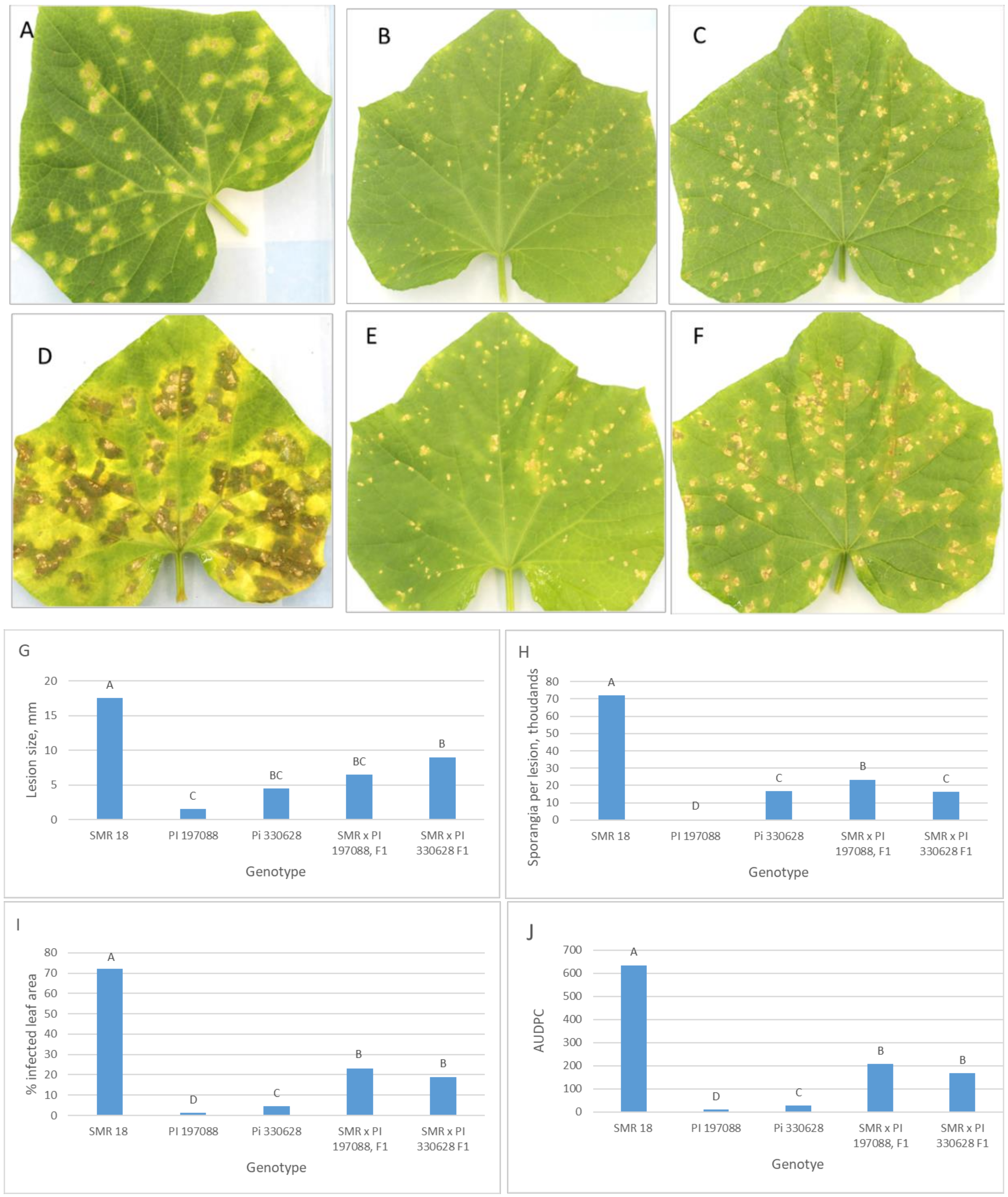
3.4. Inheritance of Resistance in Detached Leaves of Adult Plants
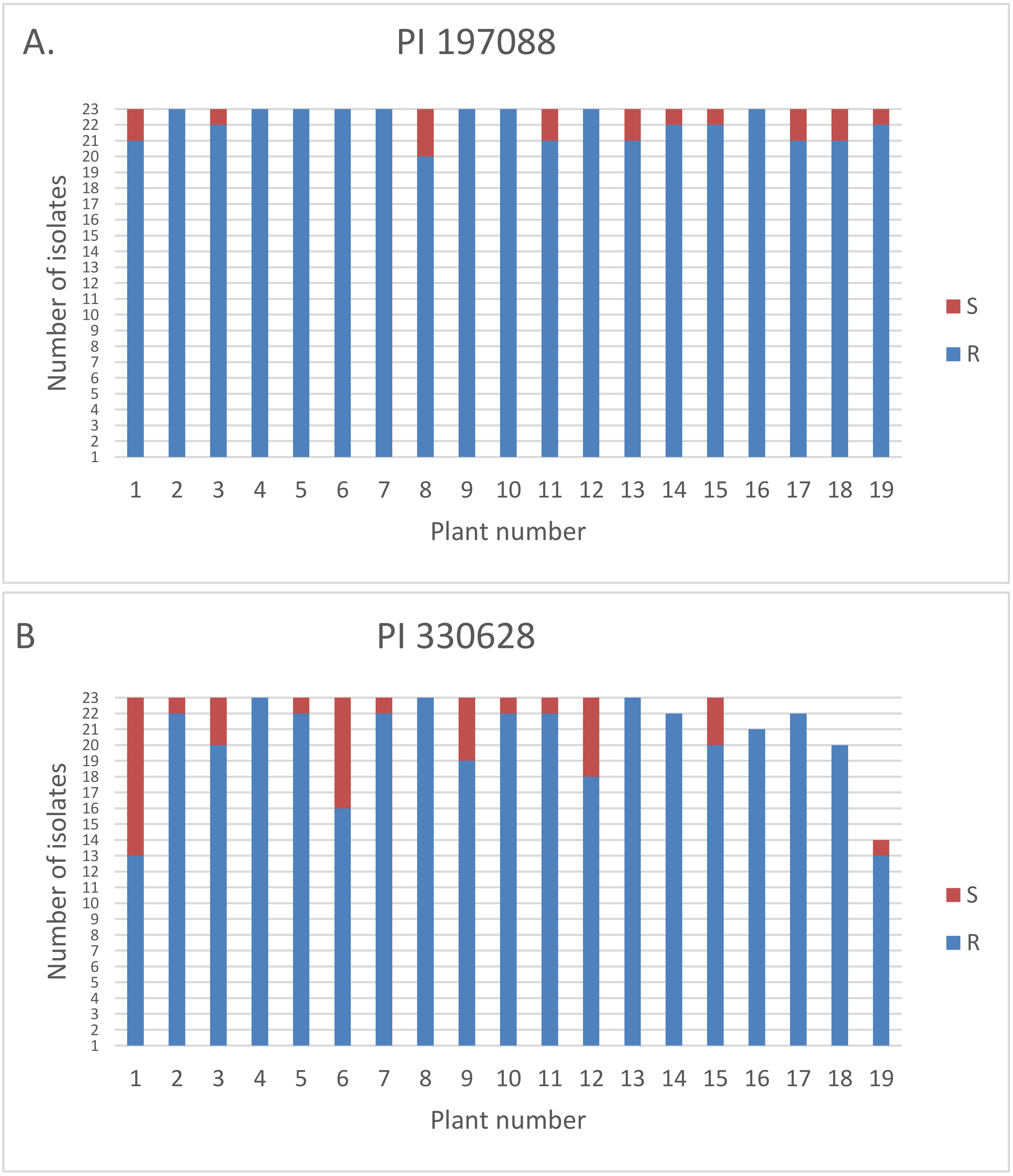
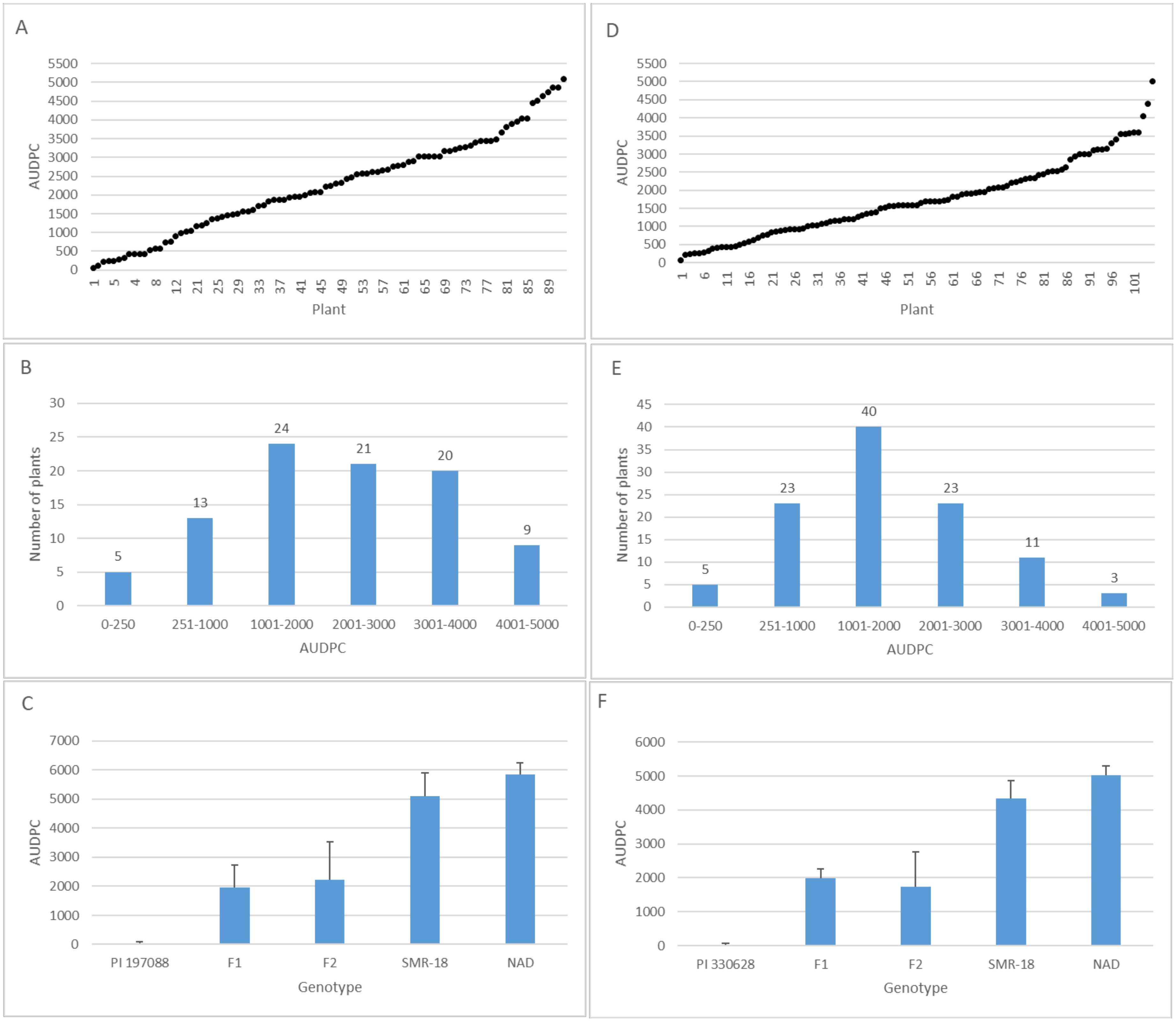
3.5. Inheritance of Resistance in Intact Field-Grown Plants
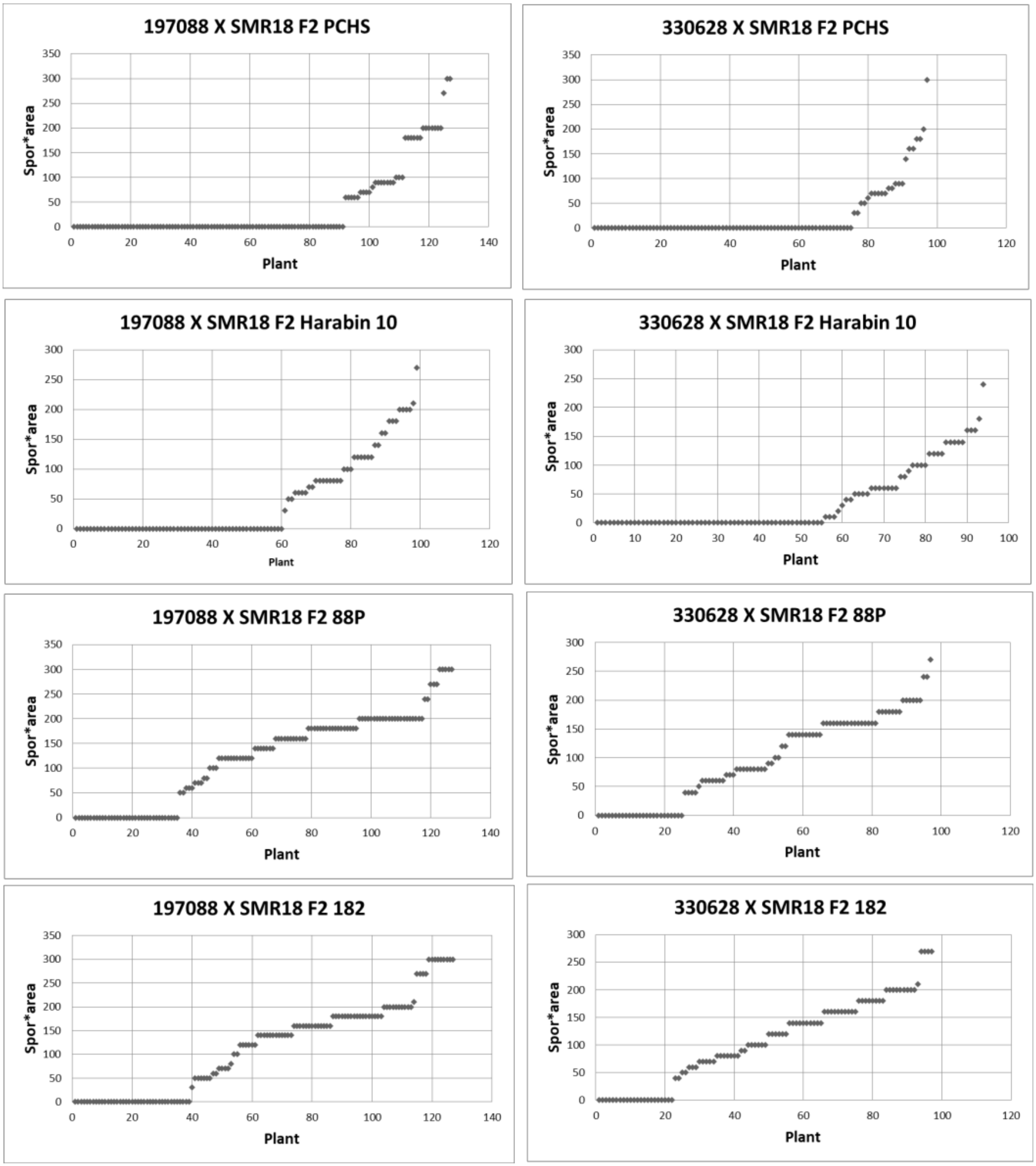
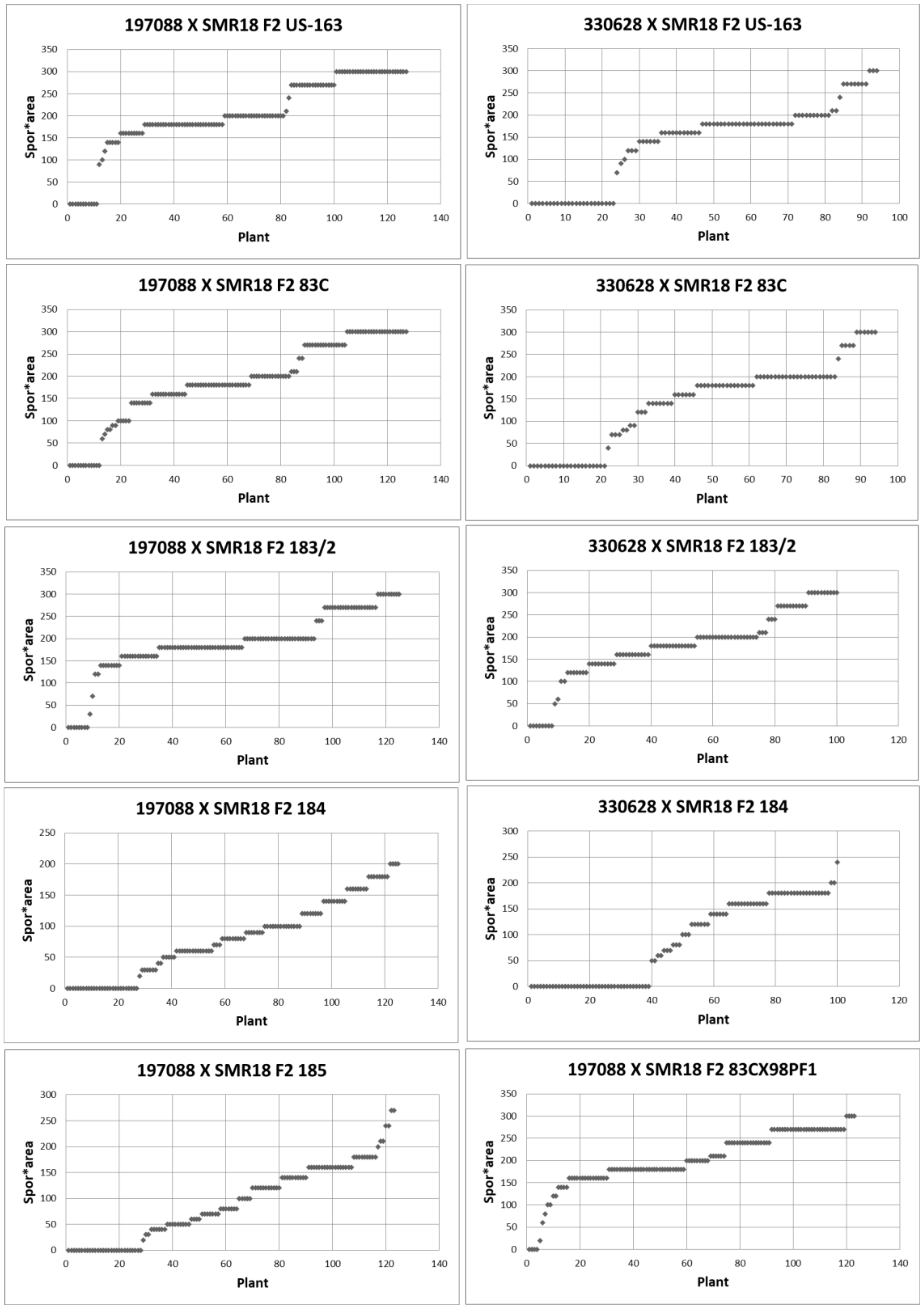
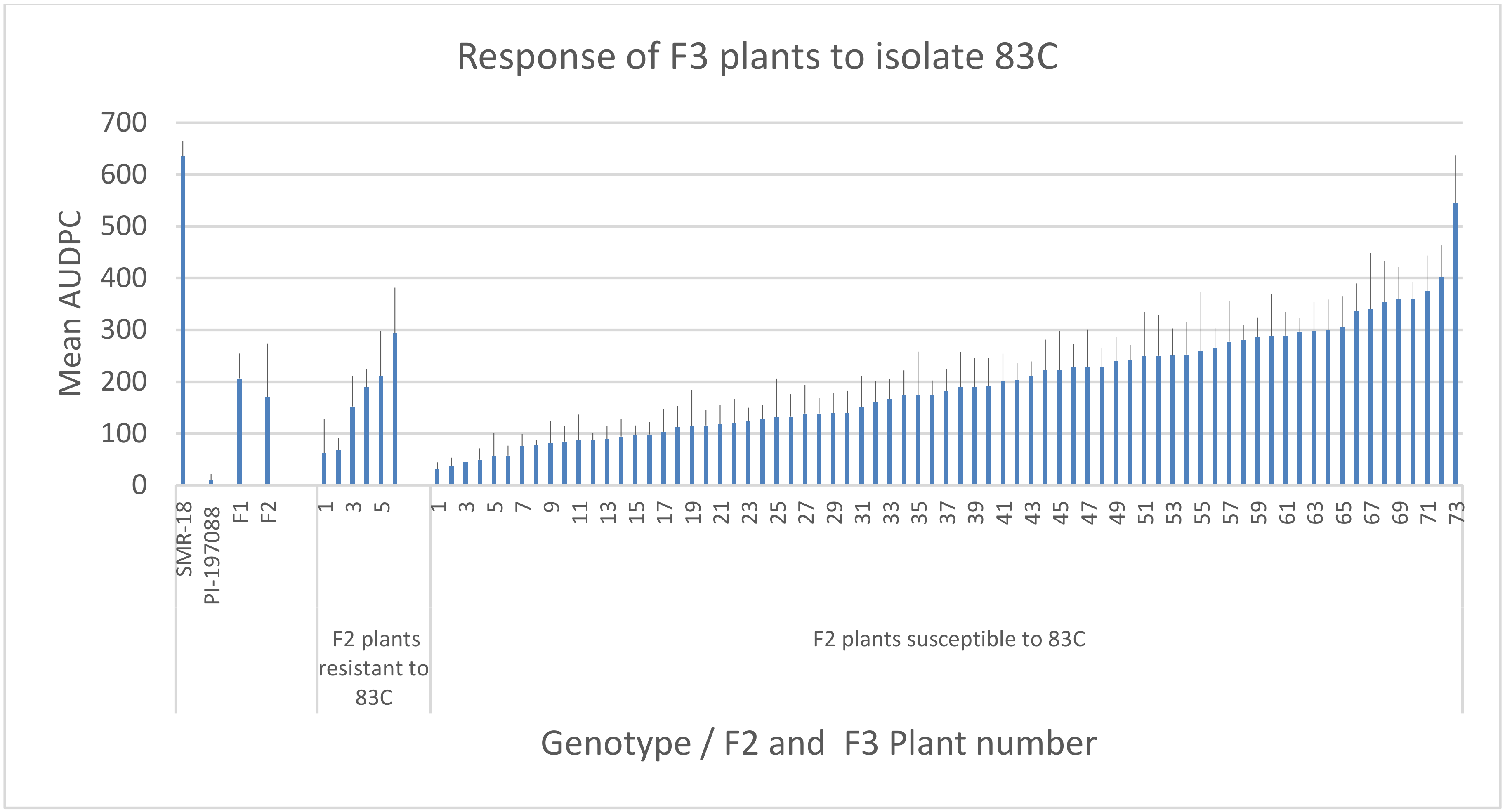
3.6. Resistance in F2: F3 Plants
3.7. Resistance of F3 Plants in the Field
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Palti, J.; Cohen, Y. Downy mildew of cucurbits (Pseudoperonospora cubensis)—The fungus and its hosts, distribution, epidemiology and control. Phytoparasitica 1980, 8, 109–147. [Google Scholar] [CrossRef]
- Lebeda, A.; Cohen, Y. Cucurbit downy mildew (Pseudoperonospora cubensis)—Biology, ecology, epidemiology, host-pathogen interaction and control. Eur. J. Plant Pathol. 2011, 129, 157–192. [Google Scholar] [CrossRef]
- Savory, E.; Granke, L.L.; Quesada-Ocampo, L.M.; Varbanova, M.; Hausbeck, M.K.; Day, B. The cucurbit downy mildew pathogen Pseudoperonospora cubensis. Mol. Plant Pathol. 2011, 12, 217–226. [Google Scholar] [CrossRef]
- Cohen, Y.; Langenberg, K.M.V.D.; Wehner, T.C.; Ojiambo, P.S.; Hausbeck, M.; Quesada-Ocampo, L.M.; Lebeda, A.; Sierotzki, H.; Gisi, U.; Whener, T.C. Resurgence of Pseudoperonospora cubensis—The causal agent of cucurbit downy mildew. Phytopathology 2015, 105, 1–15. [Google Scholar] [CrossRef]
- Ojiambo, P.; Gent, D.H.; Quesada-Ocampo, L.M.; Hausbeck, M.K.; Holmes, G.J. Epidemiology and population biology of Pseudoperonospora cubensis: A model system for management of downy mildews. Annu. Rev. Phytopathol. 2015, 53, 223–246. [Google Scholar] [CrossRef]
- Cohen, Y. The combined effects of temperature, leaf wetness and inoculum concentration on infection of cucumbers with Pseudoperonospora cubensis. Can. J. Bot. 1977, 55, 1478–1487. [Google Scholar] [CrossRef]
- Whisson, S.C.; Boevink, P.C.; Moleleki, L.N.; Avrova, A.O.; Morales, J.G.; Gilroy, E.M.; Armstrong, M.R.; Grouffaud, S.; Van West, P.; Chapman, S.; et al. A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 2007, 450, 115–118. [Google Scholar] [CrossRef]
- Perl, M.; Cohen, Y.; Rotem, J. The effect of humidity during darkness on the transfer of assimilates from cucumber leaves to sporangia of Pseudoperonospora cubensis. Physiol. Plant Pathol. 1972, 2, 113–122. [Google Scholar] [CrossRef]
- Cohen, Y. The novel oomycide oxathiapiprolin inhibits all stages in the asexual life cycle of Pseudoperonospora cubensis—Causal agent of cucurbit downy mildew. PLoS ONE 2015, 10, e0140015. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E.; Galperin, M. Control of cucumber downy mildew with novel fungicidal mixtures of oxathiapiprolin. Phytoparasitica 2018, 46, 689–704. [Google Scholar] [CrossRef]
- Call, A.D.; Criswell, A.D.; Wehner, T.C.; Ando, K.; Grumet, R. Resistance of cucumber cultivars to a new strain of cucurbit downy mildew. HortScience 2012, 47, 171–178. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E. Mating type and sexual reproduction of Pseudoperonospora cubensis, the downy mildew agent of cucurbits. Eur. J. Plant Pathol. 2012, 132, 577–592. [Google Scholar] [CrossRef]
- Thomas, A.; Carbone, I.; Lebeda, A.; Ojiambo, P. Virulence structure within populations of Pseudoperonospora cubensis in the United States. Phytopathology 2017, 107, 777–785. [Google Scholar] [CrossRef]
- Polat, I.; Baysal, O.; Mercati, F.; Kitner, M.; Cohen, Y.; Lebeda, A.; Carimi, F. Characterization of Pseudoperonospora cubensis isolates from Europe and Asia using ISSR and SRAP molecular markers. Eur. J. Plant Pathol. 2014, 139, 641–653. [Google Scholar] [CrossRef]
- Wallace, E.C.; D’Arcangelo, K.N.; Quesada-Ocampo, L.M. Population analyses reveal two host-adapted clades of Pseudoperonospora cubensis, the causal agent of cucurbit downy mildew, on commercial and wild cucurbits. Phytopathology 2020. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E.; Galperin, M. Host preference of mating type in Pseudoperonospora cubensis, the downy mildew causal agent of cucurbits. Plant Dis. 2013, 97, 292. [Google Scholar] [CrossRef]
- Pang, X.; Zhou, X.; Wan, H.; Chen, J. QTL mapping of downy mildew resistance in an introgression line derived from interspecific hybridization between cucumber and Cucumis hystrix. J. Phytopathol. 2013, 161, 536–543. [Google Scholar] [CrossRef]
- Li, L.; He, H.; Zou, Z.; Li, Y. QTL analysis for downy mildew resistance in cucumber inbred line PI 197088. Plant Dis. 2018, 102, 1240–1245. [Google Scholar] [CrossRef]
- Szczechura, W.; Staniaszek, M.; Kłosińska, U.; Kozik, E.U. Molecular Analysis of New Sources of Resistance to Pseudoperonospora cubensis (Berk. et Curt.) Rostovzev in Cucumber. Russ. J. Genet. 2015, 51, 974–979. [Google Scholar] [CrossRef]
- Van Vliet, G.J.A.; Meysing, W.D. Inheritance of resistance to Pseudoperonospora cubensis Rost. in cucumber (Cucumis sativus L.). Euphytica 1974, 23, 251–255. [Google Scholar] [CrossRef]
- McFerson, J.R. A Screening Procedure for Determining Levels of Resistance in Cucumber. Master’s Thesis, Texas A&M University, College Station, TX, USA, 1978. [Google Scholar]
- Pershin, A.F.; Medvedeva, N.I.; Medvedev, A.V. Quantitative approach to studying the genetics of disease resistance. IV. Interaction of the genetic systems for resistance to powdery and downy mildews in cucumber. Genet. USSR. 1988, 24, 484–493. [Google Scholar]
- Doruchowski, R.W.E.; Lakowska-Ryk, E. Inheritance of Resistance to Downy Mildew (Pseudoperonospora cubensis Berk & Curt) in Cucumis sativus. In Proceedings of the 5th EUCARPIA Symposium, Warsaw, Poland, 27–31 July 1992; Doruchowski, R.W., Kozik, E., Niemirowicz Szczytt, K., Eds.; Research Institute of Vegetable Crops and Warsaw Agricultural University: Warsaw, Poland, 1992; pp. 132–138. [Google Scholar]
- Petrov, L.; Boogert, K.; Sheck, L.; Baider, A.; Rubin, E.; Cohen, Y. Resistance to downy mildew, Pseudoperonospora cubensis, in cucumbers. Acta Hortic. 2000, 510, 203–209. [Google Scholar] [CrossRef]
- Angelov, D. Inheritance of resistance to downy mildew, Pseudoperonospora cubensis (Berk. & Curt.) Rostow. In Proceedings of the Report of the 2nd National Symposium of Plant Immunity, Plovdiv, Bulgaria, 1994; Volume 3, pp. 99–105. [Google Scholar]
- Caldwell, D.; Chan, E.; de Vries, J.; Joobeur, T.; King, J.; Reina, A.; Shetty, N. Methods and Compositions for Identifying Downy Mildew Resistant Cucumber Plants. U.S. Patent Application No. 2011/0126309 A1, April 2011. [Google Scholar]
- Yoshioka, Y.; Sakata, Y.; Sugiyama, M.; Fukino, N. Identification of quantitative trait loci for downy mildew resistance in cucumber (Cucumis sativus L.). Euphytica 2014, 198, 265–276. [Google Scholar] [CrossRef]
- Wang, Y.; Vandenlangenberg, K.; Wen, C.; Wehner, T.C.; Weng, Y. QTL mapping of downy and powdery mildew resistances in PI 197088 cucumber with genotyping-by-sequencing in RIL population. Theor. Appl. Genet. 2018, 131, 597–611. [Google Scholar] [CrossRef]
- Katz, D.; Hammer, R.; Ben Naim, Y.; Cohen, Y. Resistance to downy mildew in cucumber breeding lines. Phytoparasitica 2018, 46, 320. [Google Scholar]
- Wang, Y.; Vandenlangenberg, K.; Wehner, T.C.; Kraan, P.A.G.; Suelmann, J.; Zheng, X.; Owens, K.; Weng, Y. QTL mapping for downy mildew resistance in cucumber inbred line WI7120 (PI 330628). Theor. Appl. Genet. 2016, 1298, 1493–1505. [Google Scholar] [CrossRef]
- Zhang, S.P.; Liu, M.M.; Miao, H.; Zhang, S.Q.; Yang, Y.H.; Xie, B.Y.; Wehner, T.C.; Gu, X.F. Chromosomal mapping and QTL analysis of resistance to downy mildew in Cucumis sativus. Plant Dis. 2013, 97, 245–251. [Google Scholar] [CrossRef]
- Win, K.T.; Vegas, J.; Zhang, C.; Song, K.; Lee, S. QTL mapping for downy mildew resistance in cucumber via bulked segregant analysis using next-generation sequencing and conventional methods. Theor. Appl. Genet. 2017, 130, 199–211. [Google Scholar] [CrossRef]
- Cohen, Y.; Eyal, H.; Hanania, J.; Malik, Z. Ultrastructure of Pseudoperonospora cubensis in muskmelon genotype susceptible and resistant to downy mildew. Physiol. Mol. Plant Pathol. 1989, 34, 27–40. [Google Scholar] [CrossRef]
- Balass, M.; Cohen, Y.; Bar-Joseph, M. Temperature-dependent resistance to downy mildew in muskmelon: Structural responses. Physiol. Mol. Plant Pathol. 1993, 43, 11–20. [Google Scholar] [CrossRef]
- Taler, D.; Galperin, M.; Benjamin, I.; Cohen, Y.; Kenigsbuch, D. Plant eR genes encoding for photorespiratory enzymes confer resistance against disease. Plant Cell 2004, 16, 172–184. [Google Scholar] [CrossRef]
- Chen, T.; Cohen, Y. Isolate-dependent inheritance of resistance to downy mildew in cucumber. Phytoparasitica 2013, 41, 462. [Google Scholar]
- Lapin, D.; Meyer, R.C.; Takahashi, H.; Bechtold, U.; Ackerveken, G.V.D. Broad-spectrum resistance of Arabidopsis C24 to downy mildew is mediated by different combinations of isolate-specific loci. New Phytol. 2012, 196, 1171–1181. [Google Scholar] [CrossRef]
- Cohen, Y.; Eyal, H. Downy mildew-, powdery mildew- and fusarium wilt-resistant muskmelon breeding line PI-124111F. Phytoparasitica 1987, 15, 187–195. [Google Scholar] [CrossRef]
- Kenigsbuch, D.; Cohen, Y. Inheritance of resistance to downy mildew in Cucumis melo PI 124112 and commonalty of resistance genes with C. melo PI 124111F. Plant Dis. 1992, 76, 615–617. [Google Scholar] [CrossRef]
- Benjamin, I.; Kenigsbuch, D.; Galperin, M.; Abrameto, J.A.; Cohen, Y. Cisgenic melons over expressing glyoxylate-aminotransferase are resistant to downy mildew. Eur. J. Plant Pathol. 2009, 125, 355–365. [Google Scholar] [CrossRef]
- Tian, M.; Win, J.; Savory, E.; Burkhardt, A.K.; Held, M.A.; Brandizzi, F.; Day, B. 454 Genome sequencing of Pseudoperonospora cubensis reveals effector proteins with a QXLR translocation motif. Mol. Plant Microbe Interact. 2011, 24, 543–553. [Google Scholar] [CrossRef]
- Thomas, A.; Carbon, I.; Cohen, Y.; Ojiambo, P. Occurrence and distribution of mating types of Pseudoperonospora cubensis in the USA. Phytopathology 2017, 107, 313–321. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Country | Method | Genes/QTLs | Chromosome | Reference |
|---|---|---|---|---|---|
| PI 197085 | EU | F2, RAPD, SCAR | 3 | 5 | [19] |
| PI 197087 | EU | 1 recessive | [20] | ||
| PI 197087 | USA | 3 | [21] | ||
| PI 197087 | Russia | 3 partially dominant | [22] | ||
| PI 197087 | EU | 3 recessive | [23] | ||
| PI 197087 | Israel | 2 partially dominant | [24] | ||
| PI 197088 | EU | 2 recessive | [25] | ||
| PI 197088 | USA | SNP | 3 | 2, 4, 5 | [26] |
| PI 197088 | Japan | RIL | 14 | 1, 3, 5, 6, 7 | [27] |
| PI 197088 | USA | RIL/SNP | 11 | 1, 2, 3, 4, 5, 6 | [28] |
| PI 197088 | China | F2:F3/SSR | 5 | 1, 3, 4, 5 | [18] |
| PI 197088 | Israel | F2/SNP | 9 | 1, 2, 3, 4, 5, 6, 7 | [29] |
| PI 330628 | USA, EU | F2:F3/SNP | 5 | 2, 4, 5, 6 | [30] |
| Chinese Long | China | F2:F3/SSR | 5 | 1, 5, 6 | [31] |
| TH118FLM | Korea | F2:F3/SNP | 5 | 2, 4, 5, 6 | [32] |
| Isolate | Year | Country | Host | Mating Type | Pathotype | |
|---|---|---|---|---|---|---|
| 1 | PCHS | 1980 | Japan | Unknown | A1 | 3 |
| 2 | C1 | 1982 | South Carolina | C. melo | A1 | 3 |
| 3 | 62 | 1995 | Czech Republic | C. sativum | A1 | 3 |
| 4 | 66 | 2000 | France | C. sativum | A1 | 3 |
| 5 | 23 C | 2008 | Israel | C. sativum | A2 | 4 |
| 6 | US-299 | 2008 | Michigan | C. sativum | A1 | 3 |
| 7 | US-163 | 2008 | Florida | C. lanatus | A1 | 5 |
| 8 | 5 | 2010 | Bulgaria | C. sativum | A1 | 3 |
| 9 | 7 | 2010 | Bulgaria | C. sativum | A1 | 3 |
| 10 | 17 | 2010 | Turkey | C. sativum | A1 | 3 |
| 11 | 21 | 2010 | Turkey | C. sativum | A1 | 3 |
| 12 | 56 | 2010 | Turkey | C. sativum | A1 | 3 |
| 13 | 81 C | 2011 | Spain | C. sativum | A0 | 2 |
| 14 | 83 C | 2011 | Spain | C. sativum | A1 | 3 |
| 15 | 84 C | 2011 | Spain | C. sativum | A1 | 3 |
| 16 | 88 P | 2011 | Israel | C. maxima | A2 | 6 |
| 17 | 90 p | 2012 | Israel | C. maxima | A2 | 7 |
| 18 | 98 P | 2011 | Israel | C. maxima | A1 | 6 |
| 19 | 101 D | 2011 | Israel | C. maxima | A2 | 6 |
| 20 | 109 | 2011 | Ukraine | C. sativum | A1 | 3 |
| 21 | 148 | 2011 | Israel | C. sativum | A1 | 3 |
| 22 | US-504 | 2011 | New York | C. sativum | ||
| 23 | US-506 | 2011 | Ohio | C. sativum | A1 | 3 |
| 24 | Noam 19P | 2011 | Kenya | C. maxima | A2 | 6 |
| 25 | Noam C | 2011 | Israel | C. sativum | A1 | 3 |
| 26 | TW-01 | 2011 | Taiwan | C. sativum | A1 | 3 |
| 27 | 151/17 | 2012 | Israel | C. sativum | A1 | 3 |
| 28 | 185 | 2012 | Israel | C. melo | A1 | 3 |
| 29 | 171 | 2012 | Israel | C. sativum | A1 | 3 |
| 30 | 182 D | 2012 | Israel | C. sativum | A1 | 3 |
| 31 | 183/2 | 2012 | Israel | C. sativum | A1 | 3 |
| 32 | 184 | 2012 | Israel | C. moschata | A2 | 6 |
| 33 | C-29 | 2012 | China | C. sativum | A1 | 3 |
| 34 | Harbin 10 | 2012 | China | C. sativum | A1 | 3 |
| 35 | SG-11 | 2012 | China | C. sativum | A1 | 3 |
| 36 | Petiole 1 | 2012 | Israel | C. moschata | A2 | 6 |
| 37 | 172 B × 183 C, F1 | 2012 | Israel | C. melo | Hybrid | |
| 38 | 83 C x 98 P, F1 | 2012 | Israel | C. melo | Hybrid | |
| 39 | 197 C | 2013 | Israel | C. sativum | A1 | 3 |
| 40 | 198 B | 2013 | Israel | C. pepo | A2 | 6 |
| 41 | 245 | 2015 | Israel | C. sativum | A1 | 3 |
| 42 | Pol 1 | 2016 | Poland | C. sativum | A1 | 3 |
| 43 | Pol 4 | 2016 | Poland | C. sativum | A1 | 3 |
| 44 | 260 | 2018 | Israel | C. sativum | A1 | 3 |
| Isolate | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Number | 88 | 98 | 101D | 148 | 151/17 | 171 | Noam C | 17 | 21 | 56 | 66 | 84C | 7 | 5 | 109 | 62 | C1 | US-163 | US-504 | US-506 | US-299 | TW-01 | PCHS |
| PI 197088 (1) | S | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R | R |
| PI 197088 (2) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (3) | S | R | R | R | R | R | R | R | R | R | S | R | R | R | R | S | S | R | R | R | R | R | R |
| PI 197088 (4) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (5) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (6) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (7) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (8) | R | R | R | R | S | S | R | R | R | S | R | R | R | R | R | R | R | S | R | R | R | R | R |
| PI 197088 (9) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (10) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (11) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | S |
| PI 197088 (12) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (13) | R | R | R | R | S | S | R | S | R | S | S | R | R | R | R | R | S | R | R | R | R | R | R |
| PI 197088 (14) | R | R | R | R | S | S | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (15) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | S |
| PI 197088 (16) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 197088 (17) | S | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | S |
| PI 197088 (18) | R | R | R | R | R | S | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R |
| PI 197088 (19) | R | R | R | R | R | S | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (1) | S | R | R | R | R | S | R | R | S | S | S | R | R | S | S | S | S | R | R | R | R | R | S |
| PI 330628 (2) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R |
| PI 330628 (3) | R | R | R | R | R | R | R | R | R | S | S | R | R | R | R | R | S | R | R | R | R | R | R |
| PI 330628 (4) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (5) | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (6) | R | R | R | R | S | S | R | S | R | S | S | R | S | R | S | R | R | R | R | R | R | R | R |
| PI 330628 (7) | R | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R | R |
| PI 330628 (8) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (9) | S | R | R | R | R | R | R | R | S | R | S | R | R | R | R | R | R | R | R | R | R | R | S |
| PI 330628 (10) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | S |
| PI 330628 (11) | R | R | R | R | R | R | R | R | R | R | R | R | R | S | R | R | R | R | R | R | R | R | R |
| PI 330628 (12) | S | R | R | R | R | R | R | S | R | S | S | R | R | R | R | S | R | R | R | R | R | R | R |
| PI 330628 (13) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (14) | R | R | R | R | R | R | R | R | R | R | R | * | R | R | R | R | R | R | R | R | R | R | R |
| PI 330628 (15) | R | R | R | R | R | S | R | R | R | S | R | R | R | R | R | R | R | R | R | R | R | R | S |
| PI 330628 (16) | R | R | R | R | R | R | R | R | R | R | R | * | R | R | R | R | * | R | R | R | R | R | R |
| PI 330628 (17) | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | * | R | R | R | R | R | R |
| PI 330628 (18) | R | R | R | * | R | R | R | R | R | R | R | R | R | R | R | R | * | R | R | R | R | * | R |
| PI 330628 (19) | R | R | R | * | R | S | R | * | R | * | * | * | * | R | * | R | * | R | R | R | R | * | R |
| A. 197088 × SMR-18, F2 | |||||||||||||
| Two categories | Three categories | ||||||||||||
| Number of plants | Number of plants | ||||||||||||
| Isolate | Total | R | S | Ratio | p | X2 | Total | R | MR | S | Ratio | p | X2 |
| PCHS | 127 | 91 | 36 | 3:1 | 0.384 | 0.759 | 127 | 91 | 33 | 3 | 9:6:1 | * | * |
| Harbin 10 | 100 | 61 | 39 | 9:7 | 0.338 | 0.917 | 100 | 61 | 33 | 6 | 9:6:1 | 0.622 | 0.951 |
| 88P | 127 | 35 | 92 | 1:3 | 0.505 | 0.444 | 127 | 35 | 84 | 8 | 1:2:1 | * | * |
| 182D | 127 | 39 | 88 | 1:3 | 0.137 | 2.207 | 127 | 39 | 75 | 13 | 1:2:1 | * | * |
| US-163 | 127 | 11 | 116 | 1:15 | 0.262 | 1.260 | 127 | 11 | 72 | 44 | ** | ** | ** |
| 83C | 127 | 12 | 115 | 1:15 | 0.136 | 2.218 | 127 | 12 | 76 | 39 | ** | ** | ** |
| 183/2 | 125 | 8 | 117 | 1:15 | 0.945 | 0.005 | 125 | 8 | 88 | 29 | ** | ** | ** |
| 184 | 123 | 25 | 98 | 1:3 | 0.231 | 1.434 | 123 | 25 | 98 | 0 | ** | ** | ** |
| 185 | 123 | 29 | 94 | 1:3 | 0.716 | 0.133 | 123 | 29 | 92 | 2 | ** | ** | ** |
| 83CX98P, F1 | 123 | 4 | 119 | 1:15 | 0.170 | 1.887 | 123 | 4 | 87 | 32 | ** | ** | ** |
| 172BX183C, F1 | 118 | 24 | 94 | 1:3 | 0.242 | 1.367 | 118 | 24 | 91 | 3 | ** | ** | ** |
| US-504 | 123 | 70 | 53 | 9:7 | 0.883 | 0.022 | 123 | 70 | 47 | 6 | 9:6:1 | 0.820 | 0.397 |
| SG-11 | 104 | 7 | 97 | 1:15 | 0.839 | 0.041 | 104 | 7 | 32 | 65 | ** | ** | ** |
| Noam 19P | 118 | 41 | 77 | 1:3 | * | * | 118 | 41 | 75 | 2 | ** | ** | ** |
| Petiole 1 | 104 | 5 | 99 | 1:15 | 0.543 | 0.369 | 104 | 5 | 38 | 61 | ** | ** | ** |
| 23 C | 155 | 36 | 119 | 1:3 | 0.610 | 0.260 | 155 | 36 | 111 | 8 | ** | ** | ** |
| Pol 1 | 155 | 47 | 108 | 1:3 | 0.126 | 2.342 | 155 | 47 | 89 | 19 | 1:2:1 | * | * |
| Pol 4 | 158 | 134 | 24 | 13:3 | 0.252 | 1.315 | 158 | 134 | 24 | 0 | ** | ** | ** |
| US-506 | 155 | 114 | 41 | 3:1 | 0.676 | 0.174 | 155 | 114 | 41 | 0 | ** | ** | ** |
| 81 C | 151 | 30 | 121 | 1:3 | 0.145 | 2.121 | 151 | 30 | 121 | 0 | ** | ** | ** |
| 88 C | 148 | 35 | 113 | 1:3 | 0.704 | 0.144 | 148 | 35 | 113 | 0 | ** | ** | ** |
| 90 P | 156 | 46 | 110 | 1:3 | 0.196 | 1.675 | 156 | 46 | 110 | 0 | ** | ** | ** |
| B. 330628 × SMR-18, F2 | |||||||||||||
| Number of plants | Number of plants | ||||||||||||
| Isolate | Total | R | S | Ratio | p | X2 | Total | R | MR | S | Ratio | P | X2 |
| PCHS | 97 | 75 | 22 | 3:1 | 0.598 | 0.278 | 97 | 75 | 21 | 1 | 12:3:1 | * | * |
| Harbin 10 | 94 | 55 | 39 | 9:7 | 0.659 | 0.195 | 94 | 55 | 39 | 0 | 9:7 | 0.659 | 0.195 |
| 88P | 97 | 25 | 72 | 1:3 | 0.860 | 0.031 | 97 | 25 | 69 | 3 | ** | ** | ** |
| 182D | 97 | 22 | 75 | 1:3 | 0.598 | 0.278 | 97 | 22 | 71 | 4 | ** | ** | ** |
| US-163 | 94 | 23 | 71 | 1:3 | 0.905 | 0.014 | 94 | 23 | 58 | 13 | 1:2:1 | * | * |
| 83C | 93 | 20 | 73 | 1:3 | 0.436 | 0.606 | 93 | 20 | 62 | 11 | ** | ** | ** |
| 183/2 | 99 | 7 | 92 | 1:15 | 0.736 | 0.114 | 99 | 7 | 72 | 20 | ** | ** | ** |
| 184 | 99 | 38 | 61 | 1:3 | * | * | 99 | 38 | 60 | 1 | ** | ** | ** |
| 185 | 85 | 28 | 57 | 1:3 | 0.091 | 2.859 | 85 | 28 | 43 | 14 | 1:2:1 | * | * |
| 83CX98P, F1 | 86 | 7 | 79 | 1:15 | 0.469 | 0.524 | 86 | 7 | 54 | 25 | ** | ** | ** |
| 172BX183C, F1 | 68 | 12 | 56 | 1:3 | * | * | 68 | 12 | 45 | 11 | 1:2:1 | * | * |
| US-504 | 129 | 50 | 79 | 7:9 | 0.253 | 1.305 | 129 | 50 | 58 | 21 | ** | ** | ** |
| 101D | 125 | 55 | 70 | 7:9 | 0.955 | 0.003 | 125 | 55 | 65 | 5 | ** | ** | ** |
| Noam 19P | 67 | 16 | 51 | 1:3 | 0.832 | 0.045 | 67 | 16 | 44 | 7 | 1:2:1 | * | * |
| A. 197088 × SMR-18, F2 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of plants | Number of plants | |||||||||||||
| Year | Isolate | Total | R | S | Ratio | p | X2 | Total | R | MR | S | Ratio | p | X2 |
| 2013 | natural | 75 | 5 | 70 | 1:15 | 0.881 | 0.022 | 75 | 5 | 68 | 2 | 1:14:1 | 0.439 | 1.648 |
| 2014 | artificial, 83C | 180 | 11 | 169 | 1:15 | 0.939 | 0.006 | 180 | 11 | 158 | 11 | 1:14:1 | 0.994 | 0.013 |
| 2016 | artificial, 245C | 95 | 3 | 86 | 1:15 | 0.212 | 1.559 | 95 | 3 | 89 | 3 | 1:14:1 | 0.190 | 3.322 |
| 2016 | natural | 156 | 3 | 153 | 1:63 | 0.717 | 0.132 | 156 | 3 | 95 | 58 | 1:9:6 | * | * |
| 2017 | natural | 103 | 5 | 98 | 1:15 | 0.558 | 0.342 | 103 | 5 | 57 | 41 | 1:9:6 | * | * |
| 2019 | artificial, 260 | 92 | 5 | 87 | 1:15 | 0.747 | 0.104 | 92 | 5 | 78 | 9 | 1:14:1 | 0.366 | 2.012 |
| B. 330628 × SMR-18, F2 | ||||||||||||||
| 2013 | natural | 88 | 6 | 82 | 1:15 | 0.826 | 0.048 | 88 | 6 | 75 | 7 | 1:14:1 | 0.776 | 0.506 |
| 2019 | artificial, 260 | 105 | 5 | 100 | 1:15 | 0.529 | 0.397 | 105 | 5 | 97 | 3 | 1:14:1 | 0.274 | 2.592 |
| A. 197088 × SMR-18, F2 | |||||||
| Isolate | Plant 107 | Plant 111 | |||||
| PCHS | R | R | |||||
| Harbin 10 | R | R | |||||
| 88P | R | R | |||||
| 182D | R | R | |||||
| US-163 | R | R | |||||
| 83C | R | R | |||||
| 183/2 | R | R | |||||
| 184 | R | R | |||||
| 185 | R | R | |||||
| 83C × 98P, F1 | S | R | |||||
| 172B × 183C, F1 | R | S | |||||
| US-504 | R | R | |||||
| SG-11 | R | R | |||||
| Petiole 1 | R | R | |||||
| B. 197088 × SMR-18, F3 | |||||||
| Number of plants | |||||||
| Plant/Isolate | Total | R | S | Ratio | X2 | p | |
| Plant 107 | |||||||
| 182 D | 40 | 30 | 10 | 3:1 | 1 | 0 | |
| 83C × 98P, F1 | 42 | 14 | 28 | 1:3 | 0.212 | 1.556 | |
| 83C | 41 | 5 | 36 | 1:15 | 0.116 | 2.473 | |
| US-504 | 40 | 11 | 29 | 1:3 | 0.715 | 0.133 | |
| Plant 111 | |||||||
| 182 D | 34 | 11 | 23 | 1:3 | 0.322 | 0.980 | |
| 83C × 98P, F1 | 36 | 8 | 28 | 1:3 | 0.700 | 0.148 | |
| 83C | 35 | 7 | 28 | 1:3 | 0.495 | 0.467 | |
| US-504 | 35 | 13 | 22 | 7:9 | 0.097 | 2.752 | |
| Number of plants | |||||||
| Plant/Isolate | Total | R | MR | S | Ratio | X2 | p |
| Plant 107 | |||||||
| 182 D | 40 | 30 | 10 | 0 | 3:1:0 | ** | ** |
| 83C × 98P, F1 | 42 | 14 | 28 | 0 | 1:3:0 | ** | ** |
| 83 C | 41 | 5 | 36 | 0 | 1:3:0 | ** | ** |
| US-504 | 40 | 11 | 29 | 0 | 1:3:0 | ** | ** |
| Plant 111 | |||||||
| 182 D | 34 | 11 | 23 | 0 | 1:2:0 | ** | ** |
| 83C × 98P, F1 | 36 | 8 | 28 | 0 | 1:3:0 | ** | ** |
| 83 C | 35 | 7 | 28 | 0 | 1:3:0 | ** | ** |
| US-504 | 35 | 13 | 21 | 1 | * | ** | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Katz, D.; Ben Naim, Y.; Hammer, R.; Ben Daniel, B.H.; Rubin, A.E.; Cohen, Y. Isolate-Dependent Inheritance of Resistance Against Pseudoperonospora cubensis in Cucumber. Agronomy 2020, 10, 1086. https://doi.org/10.3390/agronomy10081086
Chen T, Katz D, Ben Naim Y, Hammer R, Ben Daniel BH, Rubin AE, Cohen Y. Isolate-Dependent Inheritance of Resistance Against Pseudoperonospora cubensis in Cucumber. Agronomy. 2020; 10(8):1086. https://doi.org/10.3390/agronomy10081086
Chicago/Turabian StyleChen, Tomer, Daniel Katz, Yariv Ben Naim, Rivka Hammer, Bat Hen Ben Daniel, Avia E. Rubin, and Yigal Cohen. 2020. "Isolate-Dependent Inheritance of Resistance Against Pseudoperonospora cubensis in Cucumber" Agronomy 10, no. 8: 1086. https://doi.org/10.3390/agronomy10081086
APA StyleChen, T., Katz, D., Ben Naim, Y., Hammer, R., Ben Daniel, B. H., Rubin, A. E., & Cohen, Y. (2020). Isolate-Dependent Inheritance of Resistance Against Pseudoperonospora cubensis in Cucumber. Agronomy, 10(8), 1086. https://doi.org/10.3390/agronomy10081086