3.1. Colour
The L*, a*, b*, C* parameters and the hue angle of the evaluated apples varied depending on the cultivar and treatment. The total colour difference factor (ΔE) calculated in relation to control sample 0 within a given cultivar was most often ΔE > 3, which means that the colour differences were noticeable to an average observer. The analysis of the values of individual colour parameters, the ΔE factor, and the significance of statistical differences between average values (results of Fisher’s least significant difference test) revealed that they did not follow any clear trend of variation and most probably resulted from the variability of the studied biological material rather than from vibration or storage. The analysed cultivars belong to class B (‘Gala’, Idared’, ‘Topaz’) and class C (‘Red Prince’) apples in terms of colour according to the OECD standards [
30], which means that at least 50% and 75%, respectively, of the skin area is covered with blush. Therefore, skin colour may vary considerably within a large batch. Such a high percentage of red colouring may mask possible changes in colour (bruising), which would be visible on base colour, i.e., yellow skin. It can be expected that the transport of apples with delicate texture and light creamy green skin, like e.g., ‘Papierowka’ [
26], would cause visible changes in skin colour. Among the studied cultivars, the smallest variations in skin colour parameters (statistically insignificant differences) were observed for ‘Idared’ apples. However, ‘Topaz’ and ‘Red Prince’ apples had darker and redder skin (higher a* values and lower L* and h* values) compared to ’Gala’ and ‘Idared’ apples. When assessing skin colour during the ripening of ‘Ligol’ and ‘Jonagored’ apples, Łysiak et al. [
40] noted similar L* and C* values, whereas the share of red (a*) was lower and the share of yellow (b*) was higher than those identified in this study, which is connected with the maturity stage. Apples analysed in the current work were fully ripe and were kept in cold storage after harvest till the start of the analyses. Łysiak et al. [
40] observed that changes in parameter a* in ripening apples were key for determining the harvest date. The colour parameter values obtained in this study showed the highest colour similarity between ‘Idared’ and ‘Ligol” apples [
40] (
Table 1).
Flesh colour parameters of the treated apples of each cultivar differed slightly compared to control apples in all treatments. However, the colour difference ΔE within one cultivar was small and ranged from 0.3 to 2.0, which means that there were no significant differences in flesh colour for an average observer. The analysis of the values of the colour parameters, the ΔE factor, and the significance of differences between average values (results of Fisher’s least significant difference test) did not reveal any patterns in the change of apple flesh colour. Although most of those changes were statistically significant, the low ΔE suggests that the flesh colour of treated samples was relative stable and similar to that of control samples within one cultivar. The flesh colour parameter values obtained in this study are comparable to those described in the literature for fresh (non-treated) and sulfite-pretreated apple slices [
35]. The presented flesh colour parameter values allow the conclusion that the flesh colour of ‘Idared’ apples was lighter and closer to the creamy-white colour than that of the other studied cultivars, which was darker and yellower, as can be inferred from the lower L* and h* values and higher a*, b* and C* values obtained for ‘Gala’, ‘Topaz’ and ‘Red Prince’ (
Table 2).
3.2. Firmness
The applied vibration and storage reduced firmness, but the changes were not always statistically significant. Initially, the highest firmness was observed for ‘Gala’ apples, but this cultivar also showed the biggest drop in firmness after vibration (by 9%), after storage (by 10%) and after vibration and storage (by 17%) relative to the control sample. Considerable changes in firmness were noted for ‘Idared’ apples—firmness decreased by up to 13% in the vibrated and stored sample. ‘Red Prince’ apples had the most stable firmness, which dropped by only 1–7%, depending on the treatment, but the changes were not significant (
Table 3).
The range of firmness values obtained for the four cultivars analysed in this study is lower than that mentioned in the literature for ‘Jonagold’ apples (65–70 N) [
23]. In both studies, firmness decreased during storage. The above-mentioned study reported a decline in the firmness of ‘Jonagold’ apples by 13–15% after 6 months of ULO storage at 0.5 °C, whereas in this study (standard cold storage at 6 °C) firmness decreased by 3–10% in the control and by 5–9% in the vibrated samples. The decrease in firmness during storage is a natural process commonly described by scholars [
41,
42,
43]. It is related to the disintegration of complex hydrocolloid compounds, which are responsible for the plant tissue texture, through, among other things, enzymatic degradation of pectin compounds leading to cell wall disintegration and atrophy in the middle lamella [
44]. No significant impact of mechanical vibration on the firmness of the treated samples compared to the control was observed, except ‘Gala’ apples. Probably, the 14-day period set for the assessment of shelf life was too short for any significant firmness differences between control and vibrated samples to occur. From among the tested samples, ‘Gala’ apples proved to be the most vulnerable to changes in firmness caused by storage and vibration. Other authors also confirm that vibration reduces the firmness of apples [
45] (
Table 3).
3.3. Total Soluble Solids, Dry Matter, Titratable Acidity and pH Value
The content of total soluble solids depended on cultivar and treatment (
Table 4). The lowest level of total soluble solids was noted in ‘Idared’. Apples of this cultivar are characterized by a relatively low TSS content, which has also been shown in other studies [
46]. The highest TSS level was measured in ‘Topaz’ apples from the control sample and from the sample subjected to vibration and storage. Vibration caused a TSS increase in ‘Gala’ apples and a TSS decrease in ‘Topaz’ apples, both from samples not subject to storage. In turn, among the samples kept in cold storage for 14 days only ‘Gala’ apples showed a decrease in TSS. No statistically significant changes in TSS were observed in the remaining cultivars. The TSS values were consistent with the literature on the subject [
22]. The level of total soluble solids is a good indicator of sugar concentration in the fruit, which is strongly affected by storage [
46]. Hydrolysis of polysaccharides into monosaccharides or disintegration of cell wall components usually result in an increase in TSS during the storage of apples. In this study, vibration mostly had no influence on the TSS content in apples [
47]. Nevertheless, this parameter may drop [
48] or rise [
44] as a result of vibration or storage. Changes in TSS depend on the duration and conditions of transport and storage and on the cultivar’s genetic properties. There were slight, insignificant variations in TSS in the analysed cultivars, including ‘Idared’ and ‘Red Prince’. Probably, metabolic processes and carbohydrate hydrolysis occurred only to a small extent during the relatively short period of storage. ‘Idared’ and ‘Red Prince’ apples are characterized by good storability. Changes in TSS highly correspond to changes in firmness because the increase in the content of monosaccharides is related to the disintegration of complex carbohydrates, which are responsible for tissue texture [
44]. This is confirmed by the results obtained for ‘Red Prince’, for which the smallest, insignificant changes in firmness and TSS content were observed (
Table 4).
Dry matter in the studied samples varied depending on cultivar and treatment (
Table 4). Just like with TSS, the lowest level of dry matter was found in ‘Idared’ apples, whereas the highest level was observed in ‘Topaz’ apples. The dry matter values matched those reported in the literature [
22,
49]. Neither vibration nor storage caused any significant changes in the dry matter content, irrespective of the cultivar, which was probably due to the relatively short storage period or low physiological activity [
44].
The pH value varied depending on cultivar and treatment (
Table 4). Compared with control samples, there was a decrease in the pH of the vibrated or stored apples and an increase in the pH of the vibrated and stored apples. The exception was ‘Red Prince’, where the changes in the pH value showed a reverse direction. The increase in pH in apples (by 4–14% depending on the cultivar) after vibration followed by 14-day storage can be explained by the fact that vibration accelerates metabolic processes in the fruit by increasing the respiration rate [
50].
The measured pH values were in agreement with the literature [
22] (
Table 4). The pH value of the fruit depends mainly on the content of organic acids. Their breakdown during respiration results in lower acidity and pH increase in during storage. However, in the study by Jan et al., pH changed little and insignificantly during the first 30 days of storage [
44]. In this study, the same was observed for the vibrated sample of ‘Red Prince’.
Titratable acidity assumed the lowest values in the ‘Gala’ samples and the highest in the ‘Topaz’ samples, which corresponds to the pH values (the highest for ‘Gala’ and lower for ‘Topaz’). In the above cultivars, TA was lower in the samples subject to vibration than in the control samples before storage and in the vibrated samples after storage. It decreased (by 6–13%) in the samples subjected to vibration and storage, which went hand in hand with the rise in their pH values. The TA decline is associated with the respiration rate [
44]. However, it should be stressed that changes in TA varied in individual cultivars depending on treatment and did not follow the above rule in each cultivar. The increase and decrease in TA observed during storage of fruit in this study was also described by other scholars [
51]. Napolitano et al. noted both an increase and decrease in TA in four apple cutivars after four months of cold storage at 2 °C [
52]. Other studies did not find any statistically significant changes in TA of four apple cultivars, including ‘Gala’, during 30-day cold storage at 5 ± 1 °C [
44].
The analysis of the above-described fruit parameters after harvest, during transport and storage should also take into account the indirect influence of ethylene, which is synthesized in the fruit during respiration and ripening, especially in stress conditions such as mechanical vibration induced by a moving vehicle. Ethylene initiates various physiological and biochemical processes, mainly respiration, and thus indirectly affects transpiration and disintegration of spare substances. It may influence those processes still on the tree, but this influence is most pronounced during harvest and storage. In climacteric fruit, such as apples, ethylene production increases considerably, even several dozen times, a few days preceding the optimum harvest date and a few days after harvest [
53]. To reduce ethylene production, ripening processes, and fruit decay after harvest, the ethylene perception inhibitor 1-methylcyclopropene (1-MCP) is applied at the beginning of storage [
54].
3.4. Polyphenol Content and Antioxidant Capacity
TPC varied widely, depending on cultivar and treatment (
Table 5). The lowest TPC was noted in ‘Gala’, whereas the highest was found in ‘Topaz’ (this refers to the average value from all treatments and the results of individual treatments). Apples that were subject to vibration had a higher TPC than control apples. The same was found after 14-day storage. In most cases, those differences were statistically significant. A two-way ANOVA showed that TPC significantly depended on the storage time, and in the case of ‘Topaz’—on vibration.
Antioxidant capacity varied depending on whether apples were vibrated and stored (
Table 6) and varied strongly depending on the cultivar. Similarly as in the case of TPC, the lowest antioxidant capacity was observed in the ‘Gala’ control sample on day 0, and the highest in the ‘Topaz’ sample after vibration and storage. Vibrated samples showed higher antioxidant capacity than control samples—both on day 0 and after 14 days of storage. All samples had higher antioxidant capacity after than before storage. Differences between the average values were significant in most of the samples. A two-way ANOVA showed that the changes in antioxidant capacity were significantly affected by both vibration and storage, except ‘Topaz’ apples, in which case this parameter significantly depended on vibration only.
Mechanical vibration generated during transport makes fresh fruit more likely to undergo physical, chemical, and biological changes during trade and storage. Transport shocks and vibration result directly in visual damage such as abrasion and bruising, which facilitates microbial invasion and multiplication, thus accelerating fruit decay [
27,
55]. Biochemical changes induced by stress conditions during transport may also include an increase in the content of polyphenols, which are secondary metabolites playing defensive functions in plants, e.g., preventing microbial access and development [
56]. The content of polyphenolic compounds in apples also depends on the agricultural and climatic conditions during growth as well as on storage conditions [
57].
The TPC obtained in this work is comparable to the data provided by previous studies: 221–476 mg/100 g d.m. in ‘Jonagold’ [
23], 564–748 mg/100 g d.m. in ‘Golden Delicious’ [
58] and 790–1330 mg/100 g d.m. in ‘Papierowka’ and ‘Gold Milenium’ [
26]. The literature reports both an increase and decrease in TPC in fruit after harvest. During long-term storage in a controlled atmosphere of ‘Golden Delicious’ apples, polyphenol content was found to have increased in between the 1st and 3rd month and to have decreased afterwards [
58]. Polyphenol content was observed to have increased in ‘Auksis’ apples cold-stored in normal and controlled atmosphere (ultra-low oxygen), including after applying 1-MCP [
57]. The same was noted during the storage of e.g., ‘Golden Delicious’, ‘Pinova’ and ‘Topaz’ apples and the proposed explanation was that the increase may have been due to, among other things, the activity of ethylene, which stimulates L-phenylalanine ammonia lyase (PAL), one of key enzymes responsible for the synthesis of polyphenols [
56]. Cold storage triggers in apples a synthesis of ACC-oxidase (1-aminocyclopropane-1-carboxylate
oxidase), an enzyme crucial to the synthesis of ethylene. Ethylene, in turn, induces the synthesis of PAL enzyme, which regulates the biosynthesis of flavonoids [
59]. An increase in the polyphenol content may also result from the depolymerization of phenolic compounds to more water-soluble free phenols [
52]. During ripening and storage, polyphenols bound to the cell wall evolve into a free form, which can be more efficiently extracted for analytical purposes [
59].
Antioxidant capacity of plant materials is significantly correlated with the content of bioactive compounds with antioxidant properties, such as polyphenols or vitamins [
56,
60,
61,
62]. This is also confirmed by this study, in which the correlation coefficient between TPC and antioxidant capacity was between 0.94 and 0.97 depending on the cultivar, thus demonstrating a high positive correlation. The antioxidant capacity values observed in this study are comparable with those presented in the literature [
22,
23]. Studies also show that an increase in polyphenol content during storage is accompanied by an increase in antioxidant capacity [
23,
56,
63]. At the same time, it has been noted that keeping apples in room temperature, i.e., in conditions similar to consumers’ homes, leads to a drop in their polyphenol content and thus also antioxidant capacity [
56]. Generally, there has been a lot of controversy over the relation between the content of phenolic compounds and the antioxidant capacity of food products. Some studies identified no correlation at all, whereas other studies found a strong correlation between antioxidant capacity and the content of phenolic compounds in apple extracts. This discrepancy is probably due to the differences in the methods used to measure antioxidant capacity as well as different extraction procedures [
52].
Biochemical changes occur not only when the fruit ripens on the plant but also after harvest, when such changes are triggered by physiological processes which depend on external conditions. During the transport and storage of apples, as a result of metabolic processes, respiration and transpiration, changes occur in the content and profile of bioactive compounds, including polyphenols. Those compounds are crucial to the plant’s resistance to pathogens, so their concentration rises in response to mechanical damage and microbial infections in fruit after harvest [
56]. Harvest and transport trigger a physiological reaction of fruit to stress conditions, which is expressed by, among other things, an increase in polyphenol content and antioxidant activity during distribution and storage. The high polyphenol content and antioxidant capacity of the studied samples allow the conclusion that apples remain a valuable and healthy food component even after being transported and kept in cold storage or on a shop shelf.
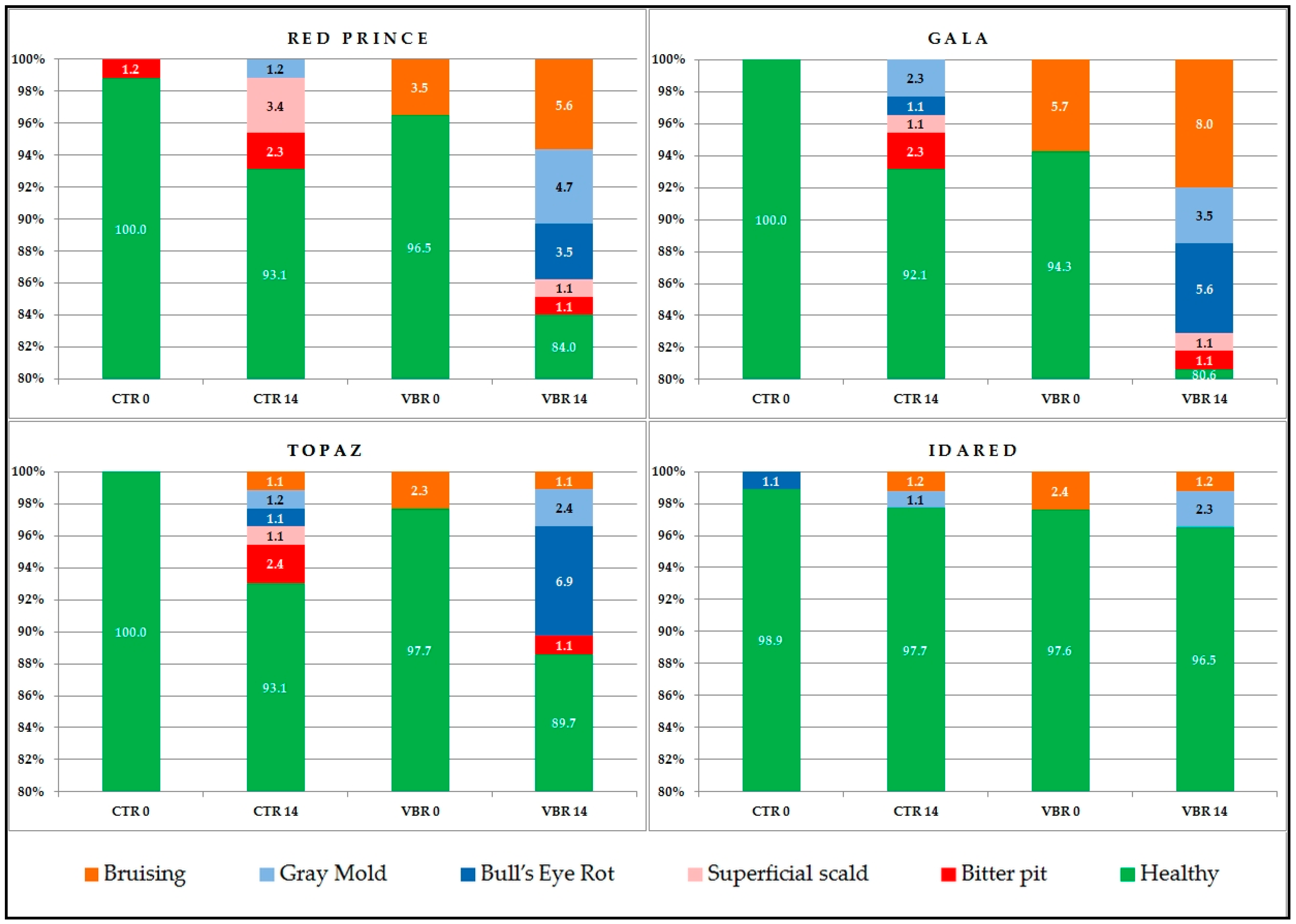
3.5. Changes Caused by Diseases and Disorders
Postharvest diseases of pome fruit result in substantial economic losses during storage worldwide every year [
64]. In the study, the occurrence of fungal diseases, physiological disorders, and visible physical damage of fruit flesh depended on the cultivar and treatment (
Figure 3). The apples were thoroughly selected for the experiment and of top quality, therefore virtually all apples were healthy during a preliminary assessment, and any diseases, disorders, or damage were identified only in isolated cases (bitter pit in ‘Red Prince’ and bull’s eye rot in ‘Idared’) and were most probably due to grading errors. Apples vibrated and assessed on the same day did not have any fungal diseases or physiological disorders; only bruising was identified, probably as an effect of treatment. After 14-day storage, both vibrated and control apples had fungal diseases and physiological disorders, which surely resulted from ripening. Many fungal diseases and physiological disorders appear on the fruit as soon as it becomes mature for consumption [
4].
The lowest percentage of loss was observed for ‘Idared’, a cultivar considered as having good storability even in a normal atmosphere cold room [
65], i.e., in conditions similar to those applied in this study. The highest percentage of losses caused by diseases and disorders (11.5%) and bruising (8.0%) was identified in ‘Gala’ apples after 14 days following vibration, which is quite understandable given the fact that ‘Gala’ also suffered the biggest drop in firmness in the same period (
Table 3). ‘Topaz’ is sensitive to
Pezicula sp. infection leading to bull’s eye rot [
66]—this was also confirmed by our study because ‘Topaz’ had the highest bull’s eye rot incidence rate after 14 days of storage. ‘Jonagold’ and its mutants are characterized by excellent storability in a controlled atmosphere [
67], but are far less storable in a normal atmosphere. The same will apply if apples suffer physical damage and are then stored at a relatively high temperature. The diagram shows that the treatment-induced losses in apples amounted to about 16% and were over 10% higher than the losses identified in fruit not subject to simulated transport vibration.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}