
Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought Tolerance


 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Interspecific Congruity-Backcross Lines
2.2. Multi-Locality Field Trials
2.3. Experimental Design
2.4. Phenotyping
2.5. Statistical Analyses
3. Results
3.1. Phenotypic and Physiological Variability of Interspecific Lines Per Locality
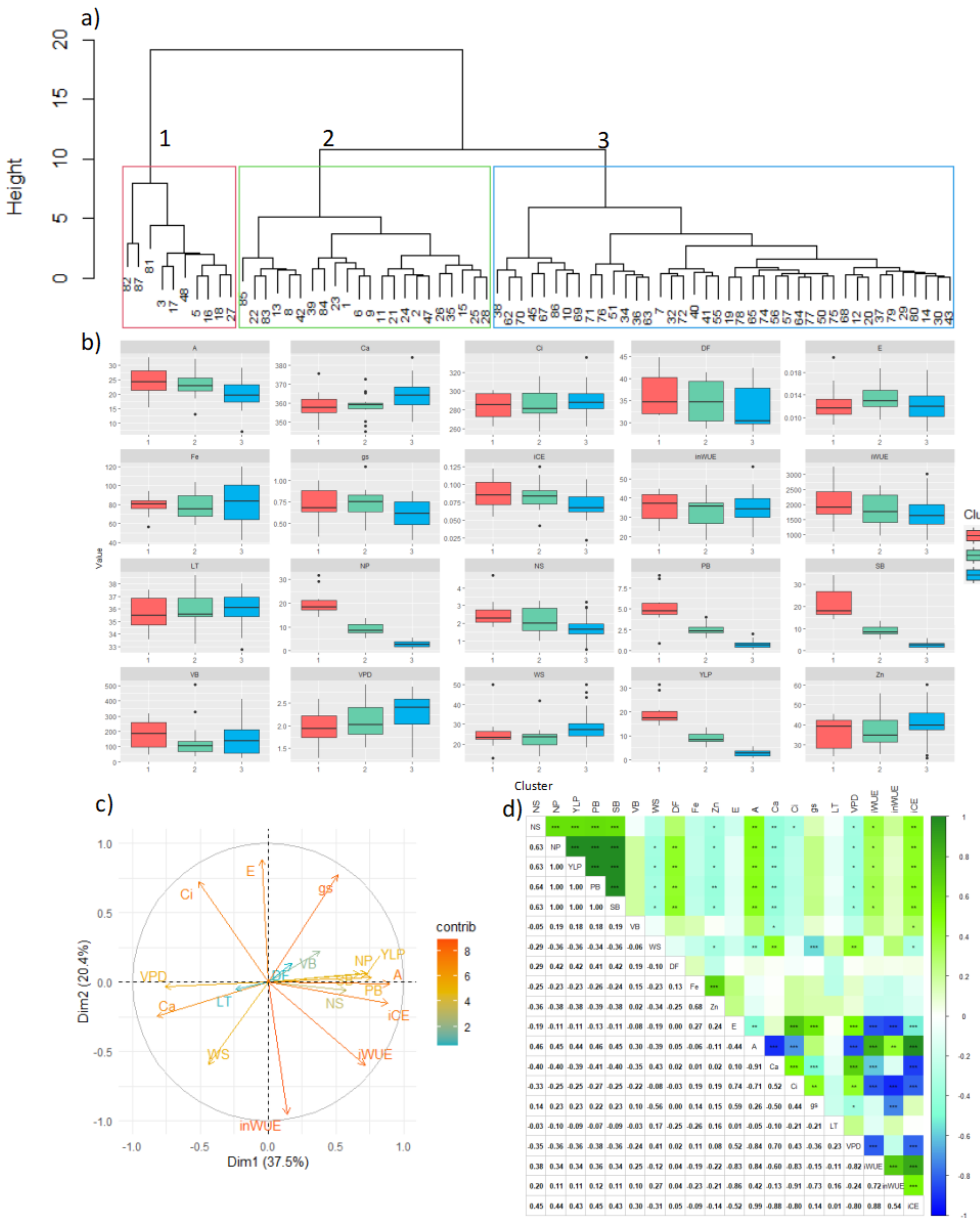
3.2. Motilonia (Warmest Dry Caribbean Savanna Sub-Region)
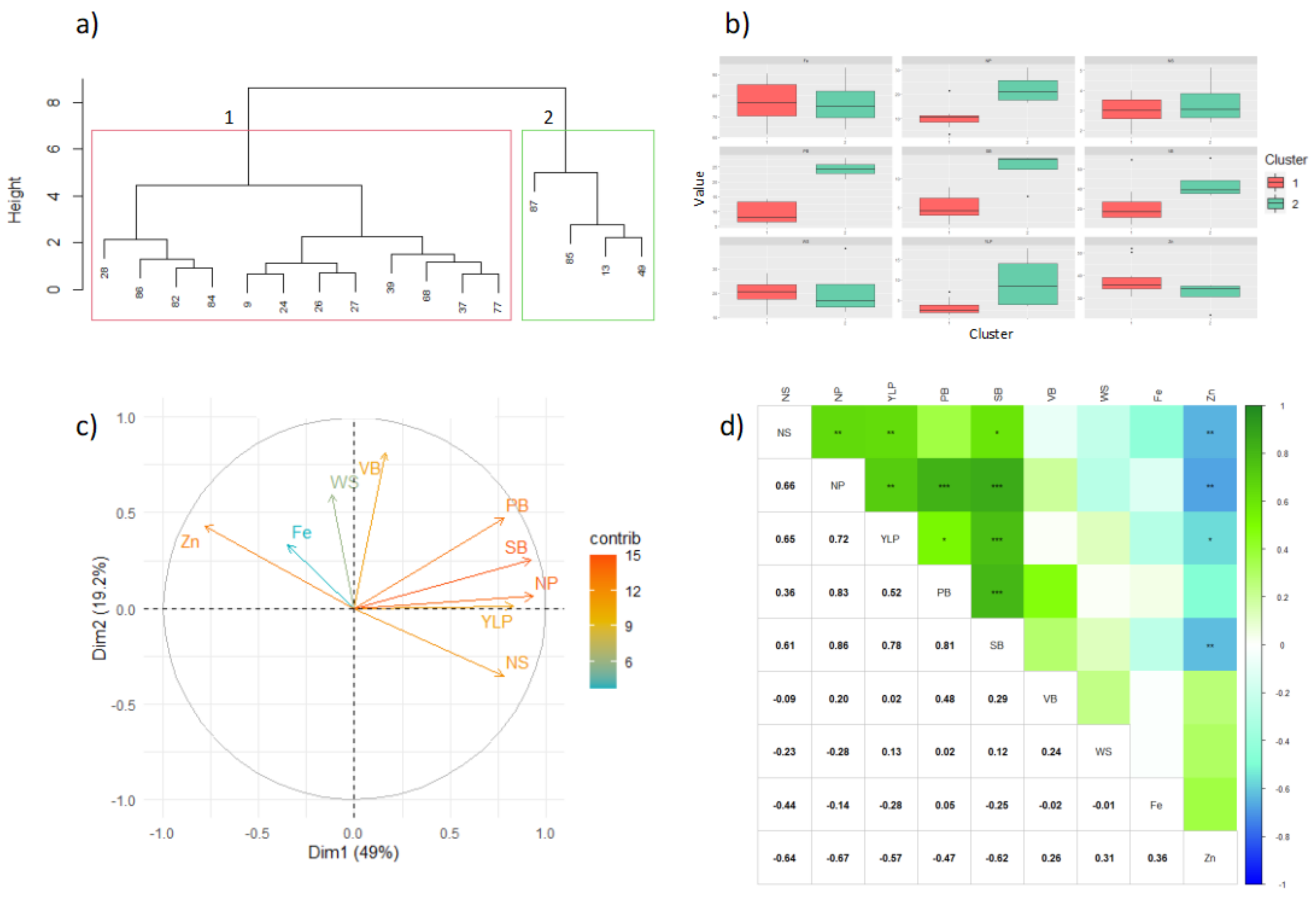
3.3. Caribia (Driest Caribbean Sub-Region)
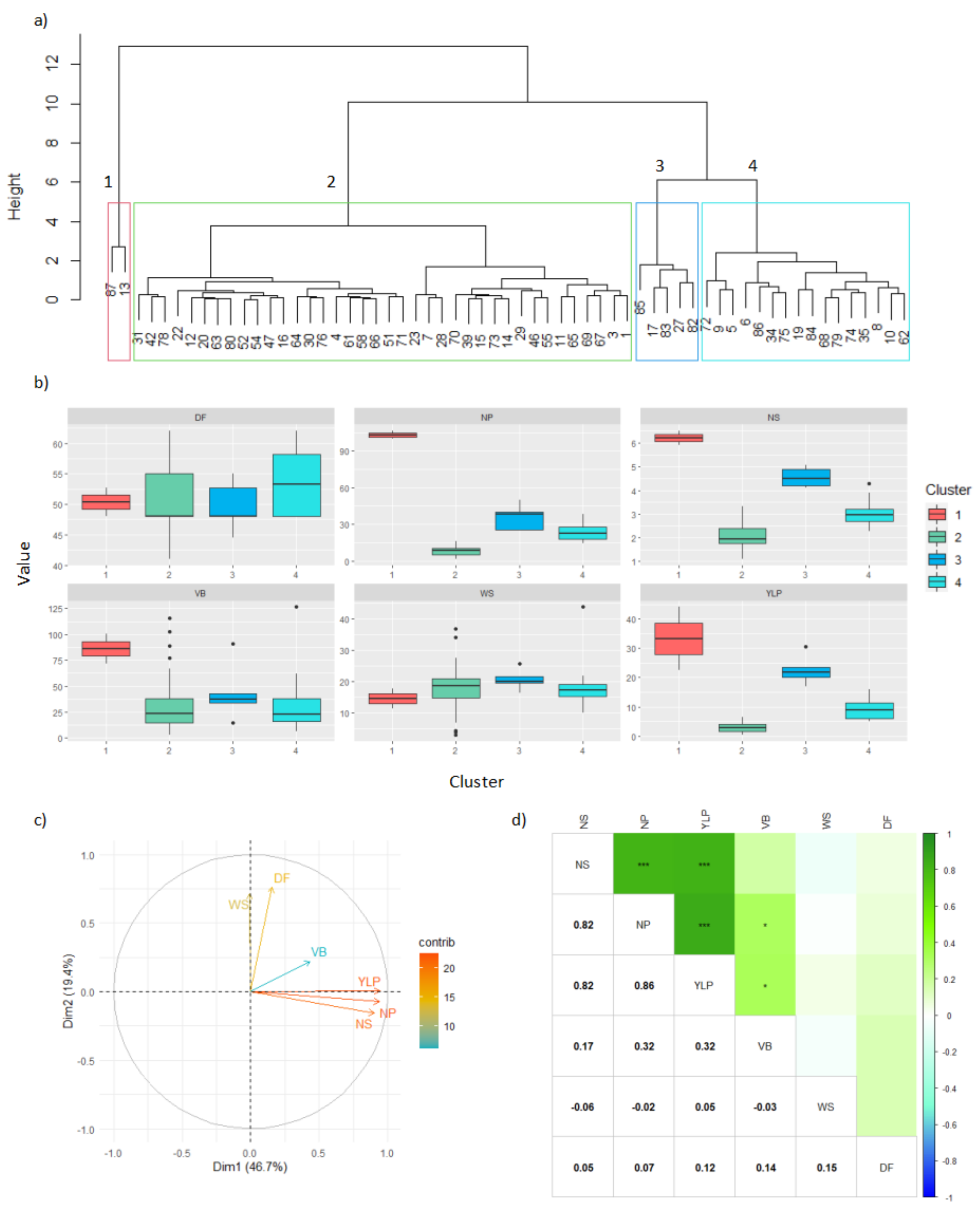
3.4. Turipaná (Humid Caribbean Sub-Region)
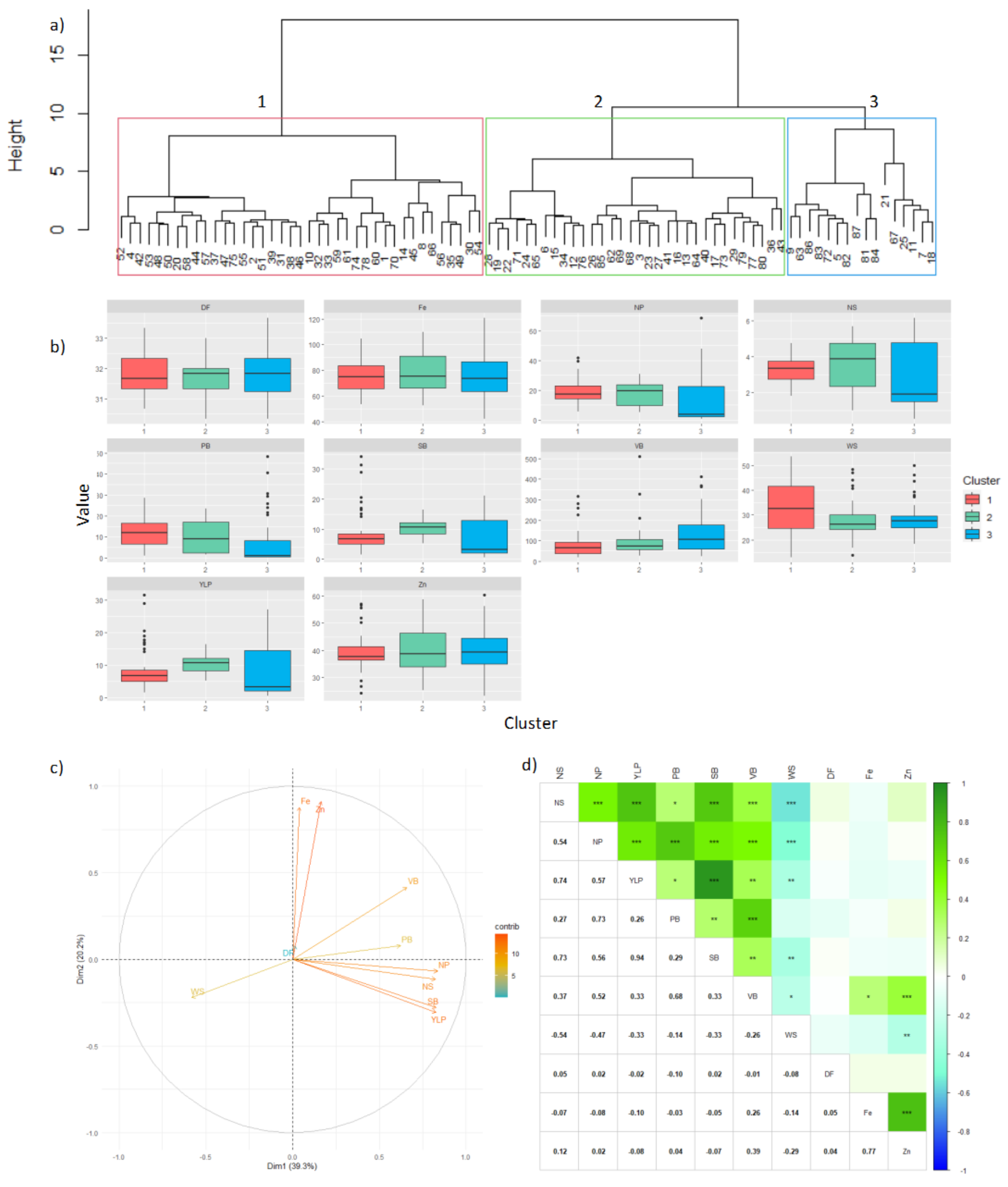
3.5. Carmen de Bolívar (Humid Caribbean Sub-Region)
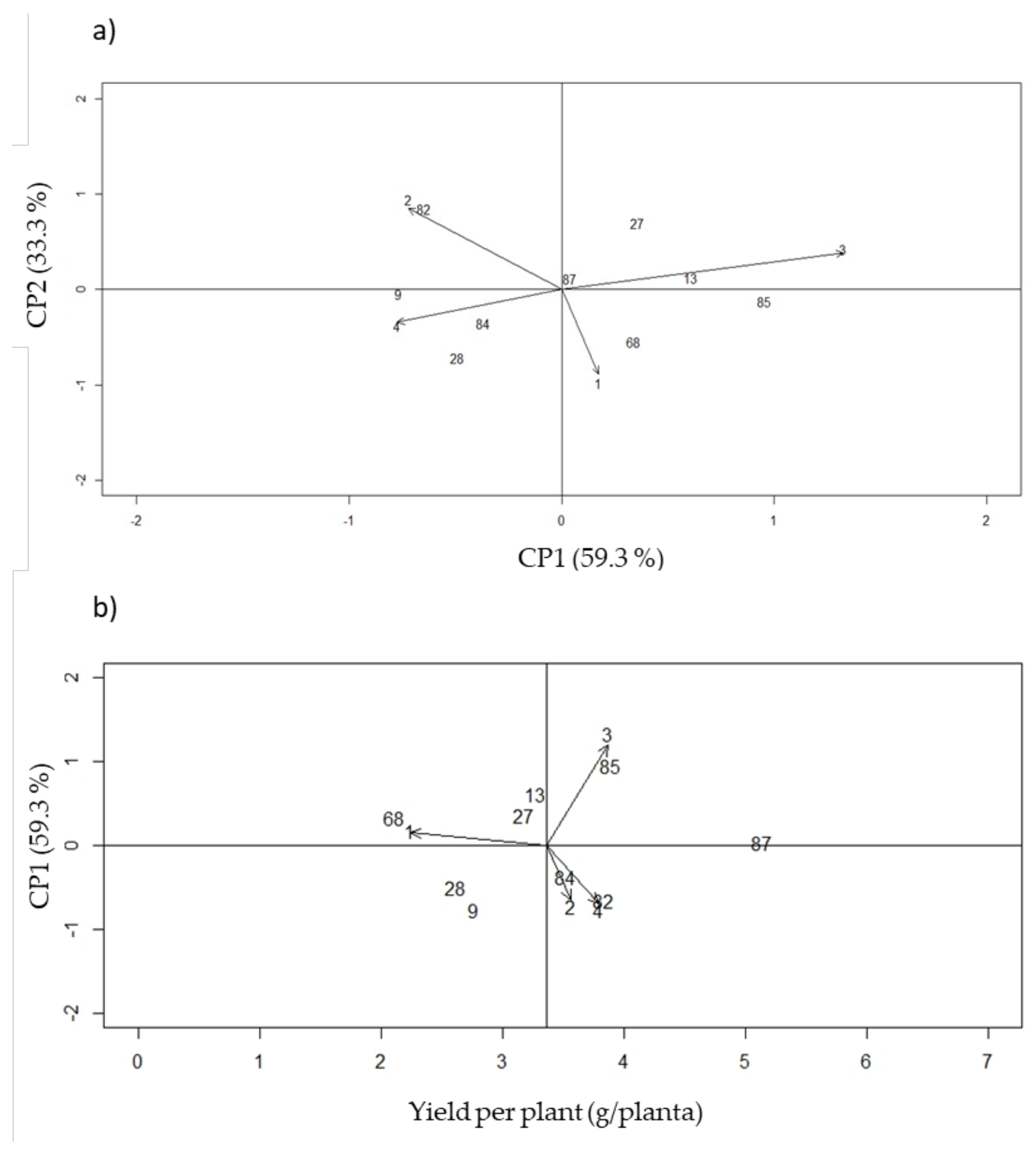
3.6. Phenotypic Stability
4. Discussion
5. Perspectives
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beebe, S. Common Bean Breeding in the Tropics. Plant Breed. Rev. 2012, 36, 357–426. [Google Scholar]
- Caproni, L.; Raggi, L.; Talsma, E.F.; Wenzl, P.; Negri, V. European landrace diversity for common bean biofortification: A genome-wide association study. Sci. Rep. 2020, 10, 19775. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Veneklaas, E.; Polania, J.; Rao, I.M.; Beebe, S.E.; Merchant, A. Field drought conditions impact yield but not nutritional quality of the seed in common bean (Phaseolus vulgaris L.). PLoS ONE 2019, 14, e0217099. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Porcel, M.; Velásquez, D.F.A.; Orduz-Rodríguez, J.O. Extreme climate variability weakens a major tropical agricultural hub. Ecol. Indic. 2020, 111, 106015. [Google Scholar] [CrossRef]
- Mukherjee, S.; Ashfaq, M.; Mishra, A.K. Compound drought and heatwaves at a global scale: The role of natural climate variability-associated synoptic patterns and land-surface energy budget anomalies. J. Geophys. Res. Atmos. 2020, 125, e2019JD031943. [Google Scholar] [CrossRef]
- Dosio, A.; Mentaschi, L.; Fischer, E.M.; Wyser, K. Extreme heat waves under 1.5 °C and 2 °C global warming. Environ. Res. Lett. 2018, 13, 054006. [Google Scholar] [CrossRef]
- Beebe, S.; Rao, I.; Blair, M.; Acosta, J. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef]
- Buitrago-Bitar, M.A.; Cortés, A.J.; López-Hernández, F.; Londoño-Caicedo, J.M.; Muñoz-Florez, J.E.; Muñoz, L.C.; Blair, M.W. Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes 2021, 12, 556. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Mashilo, J.; Shimelis, H.; Assefa, A.B.; Modi, A.T. Progress in genetic analysis and breeding of tepary bean (Phaseolus acutifolius A. Gray): A review. Sci. Hortic. 2018, 237, 119. [Google Scholar] [CrossRef]
- Jiri, O.; Mafongoya, P.L.; Chivenge, P. Climate Smart Crops for food and nutritional security for semi-arid zones of Zimbabwe. Afr. J. Food Agric. Nutr. Dev. 2017, 17, 12280–12294. [Google Scholar] [CrossRef]
- Moghaddam, S.M.; Oladzad, A.; Koh, C.; Ramsay, L.; Hart, J.P.; Mamidi, S.; McClean, P.E. The tepary bean genome provides insight into evolution and domestication under heat stress. Nat. Commun. 2021, 12, 14. [Google Scholar] [CrossRef]
- Mwale, S.E.; Shimelis, H.; Mafongoya, P.; Mashilo, J. Breeding tepary bean (Phaseolus acutifolius) for drought adaptation: A review. Plant Breed. 2020, 139, 821–833. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Duque, M.C.; Debouck, D.G.; Blair, M.W. Taxonomy of Tepary Bean and Wild Relatives as Determined by Amplified Fragment Length Polymorphism (AFLP) Markers. Crop. Sci. 2006, 46, 1744–1754. [Google Scholar] [CrossRef]
- Mejía-Jiménez, A.; Muñoz, C.; Jacobsen, H.J.; Roca, W.M.; Singh, S.P. Interspecific hybridization between common and tepary beans: Increased hybrid embryo growth, fertility, and efficiency of hybridization through recurrent and congruity backcrossing. Appl. Genet. 1994, 88, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Belivanis, T.; Doré, C. Interspecific hybridization of Phaseolus vulgaris L. and Phaseolus angustissimus A. Gray using in vitro embryo culture. Plant Cell Rep. 1986, 5, 329–331. [Google Scholar] [CrossRef]
- Souter, J.R.; Gurusamy, V.; Porch, T.G.; Bett, K.E. Successful Introgression of Abiotic Stress Tolerance from Wild Tepary Bean to Common Bean. Crop. Sci. 2017, 57, 1160–1171. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Blair, M.W.; Duque, M.C.; Tohme, J.; Roca, W. Introgression in Common Bean × Tepary Bean Interspecific Congruity-Backcross Lines as Measured by AFLP Markers. Crop. Sci. 2004, 44, 637–645. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortes, A.J.; Farmer, A.D.; Huang, W.; Ambachew, D.; Penmetsa, R.V.; Carrasquilla-Garcia, N.; Assefa, T.; Cannon, S.B. Uneven recombination rate and linkage disequilibrium across a reference SNP map for common bean (Phaseolus vulgaris L.). PLoS One 2018, 13, e0189597. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortés, A.J.; Soler, A. Diversification and Population Structure in Common Beans (Phaseolus vulgaris L.). PLoS ONE 2012, 7, e49488. [Google Scholar] [CrossRef]
- Borba, M.E.A.; Maciel, G.M.; Júnior, E.F.; Júnior, C.M.; Marquez, G.R.; Silva, I.G.; Almeida, R.S. Gas exchanges and water use efficiency in the selection of tomato genotypes tolerant to water stress. Genet. Mol. Res. 2017, 16, gmr16029685. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A.; Mundt, F. Package ‘factoextra’. Extract and visualize the results of multivariate data analyses. RCRAN 2017, 76, 1–74. [Google Scholar]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R Package for Determining the Relevant Number of Clusters in a Data Set. J. Stat. Softw. 2014, 61, 36. [Google Scholar] [CrossRef]
- Kassambara, A. Practical guide to principal component methods in R: PCA, M (CA), FAMD, MFA, HCPC, factoextra. STHDA 2016, 2, 152. [Google Scholar]
- Van Buuren, S.; Groothuis-Oudshoorn, K. Mice: Multivariate imputation by chained equations in R. J. Stat. Softw. 2011, 45, 67. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. RCRAN 2011, a1748, 1–155. [Google Scholar]
- Gauch, H.G., Jr.; Piepho, H.-P.; Annicchiarico, P. Statistical Analysis of Yield Trials by AMMI and GGE: Further Considerations. Crop Sci. 2008, 48, 866–889. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar Evaluation and Mega-Environment Investigation Based on the GGE Biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Gonçalves, J.G.R.; Chiorato, A.F.; Perina, E.F.; Carbonell, S.A.M. Estabilidade fenotípica em feijoeiro estimada por análise ammi com genótipo suplementar. Bragantia 2009, 68, 871. [Google Scholar] [CrossRef]
- Barros, M.A.; Rocha, M.D.M.; Gomes, R.L.F.; Silva, K.J.D.; Neves, A.C.D. Adaptabilidade e estabilidade produtiva de feijão-caupi de porte semiprostrado. Pesqui. Agropecuária Bras. 2013, 48, 410. [Google Scholar] [CrossRef]
- De Oliveira, E.J.; de Freitas, J.P.X.; de Jesus, O.N. AMMI analysis of the adaptability and yield stability of yellow passion fruit varieties. Sci. Agric. 2014, 71, 145. [Google Scholar] [CrossRef]
- Escobar, E.A.V.; Sánchez, J.E.V.; García, D.B. Analysis of stability and adaptability of QPM hybrids of maize growing in different Colombian agroecological zones. Acta Agronómica 2016, 65, 79. [Google Scholar]
- Polania, J.; Rao, I.M.; Cajiao, C.; Grajales, M.; Rivera, M.; Velasquez, F.; Raatz, B.; Beebe, S.E. Shoot and Root Traits Contribute to Drought Resistance in Recombinant Inbred Lines of MD 23–24 × SEA 5 of Common Bean. Front. Plant Sci. 2017, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.C.; Polanía, J.A.; Contreras, A.T.; Rodríguez, L.; Machado, L.; Ordoñez, C.; Beebe, E.; Rao, I.M. Adaptation of common bean lines to high temperature conditions: Genotypic differences in phenological and agronomic performance. Euphytica 2020, 216, 22. [Google Scholar] [CrossRef]
- Guevara-Escudero, M.; Osorio, A.N.; Cortés, A.J. Integrative Pre-Breeding for Biotic Resistance in Forest Trees. Plants 2021, 10, 2022. [Google Scholar] [CrossRef]
- Camilo, S.; Odindo, A.O.; Kondwakwenda, A.; Sibiya, J. Root Traits Related with Drought and Phosphorus Tolerance in Common Bean (Phaseolus vulgaris L.). Agronomy 2021, 11, 552. [Google Scholar] [CrossRef]
- Da Silva Sá, F.V.; Ferreira Neto, M.; de Lima, Y.B.; de Paiva, E.P.; Prata, R.C.; Lacerda, C.F.; Brito, M.E.B. Growth, gas exchange and photochemical efficiency of the cowpea bean under salt stress and phosphorus fertilization. Comun. Sci. 2018, 9, 668–679. [Google Scholar]
- Parsons, L.R.; Howe, T.K. Effects of water stress on the water relations of Phaseolus vulgaris and the drought resistant Phaseolus acutifolius. Physiol. Plant. 1984, 60, 197–202. [Google Scholar] [CrossRef]
- Jimenez-Galindo, J.C.; Alvarez-Iglesias, L.; Revilla-Temino, P.; Jacinto-Soto, R.; Garcia-Dominguez, L.E.; de La Fuente-Martinez, M.; Osorno, J.M. Screening for Drought Tolerance in Tepary and Common Bean Based on Osmotic Potential Assays. Plant 2018, 6, 24–32. [Google Scholar] [CrossRef]
- Gujaria-Verma, N.; Ramsay, L.; Sharpe, A.G.; Sanderson, L.A.; Debouck, D.G.; Tar’an, B.; Bett, K.E. Gene-based SNP discovery in tepary bean (Phaseolus acutifolius) and common bean (P. vulgaris) for diversity analysis and comparative mapping. BMC Genom. 2016, 17, 239. [Google Scholar] [CrossRef]
- Rosas, J.C.; Beaver, J.S.; Escoto, D.; Pérez, C.A.; Llano, A.; Hernández, J.C.; Araya, R. Registration of ‘Amadeus 77’ Small Red Common Bean. Crop Sci. 2004, 44, 1867–1868. [Google Scholar] [CrossRef]
- Cortés, A.J.; Blair, M.W. Genotyping by Sequencing and Genome–Environment Associations in Wild Common Bean Predict Widespread Divergent Adaptation to Drought. Front. Plant Sci. 2018, 9, 128. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortés, A.J.; This, D. Identification of an ERECTA gene and its drought adaptation associations with wild and cultivated common bean. Plant Sci. 2016, 242, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Galeano, C.H.; Cortés, A.J.; Fernández, A.C.; Soler, Á.; Franco-Herrera, N.; Makunde, G.; Vanderleyden, J.; Blair, M.W. Gene-Based Single Nucleotide Polymorphism Markers for Genetic and Association Mapping in Common Bean. BMC Genet. 2012, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Chavarro, M.C.; Madriñán, S.; This, D.; Blair, M.W. Molecular ecology and selection in the drought-related Asr gene polymorphisms in wild and cultivated common bean (Phaseolus vulgaris L.). BMC Genet. 2012, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- López-Hernández, F.; Cortés, A.J. Last-Generation Genome–Environment Associations Reveal the Genetic Basis of Heat Tolerance in Common Bean (Phaseolus vulgaris L.). Front. Genet. 2019, 10, 954. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Amelework, B.; Shimelis, H.; Modi, A.T.; Mashilo, J. Genetic differentiation among selected tepary bean collections revealed by morphological traits and simple sequence repeat markers. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 608–618. [Google Scholar] [CrossRef]
- Blair, M.W.; Pantoja, W.; Carmenza Munoz, L. First use of microsatellite markers in a large collection of cultivated and wild accessions of tepary bean (Phaseolus acutifolius A. Gray). Appl. Genet. 2012, 125, 1137–1147. [Google Scholar] [CrossRef]
- Crespo-Muñoz, S.; Rivera-Peña, M.; Rosero-Alpala, D.A.; Muñoz-Florez, J.E.; Rao, I.M.; Muñoz-Florez, L.C. Pollen viability of Tepary bean (Phaseolus acutifolius A. Gray) mutant lines under water stress conditions and inoculation with rhizobia. Acta Agron. 2018, 67, 319–325. [Google Scholar] [CrossRef]
- Rao, I.; Polania, J.; Beebe, S.; Ricaurte, J.; Cajiao, C.; Garcia, R.; Rivera, M. Can tepary bean be a model for improvement of drought resistance in common bean. Afr. Crop. Sci. J. 2013, 21, 256–281. [Google Scholar]
- Barghi, N.; Hermisson, J.; Schlötterer, C. Polygenic adaptation: A unifying framework to understand positive selection. Nat. Rev. Genet. 2020, 21, 769–781. [Google Scholar] [CrossRef]
- Rivera, A.P.T.; Murgas, R.E.C.; Ríos, A.E.M.; Merini, L.J. Efecto del glifosato sobre la microbiota, calidad del suelo y cultivo de frijol biofortificado en el departamento del Cesar, Colombia. Rev. Argent. Microbiol. 2020, 52, 61–71. [Google Scholar]
- Isik, F.; Holland, J.; Maltecca, C. Genetic Data Analysis for Plant and Animal Breeding; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Medina, C.A.; Hawkins, C.; Liu, X.P.; Peel, M.; Yu, L.X. Genome-Wide Association and Prediction of Traits Related to Salt Tolerance in Autotetraploid Alfalfa (Medicago sativa L.). Int. J. Mol. Sci. 2020, 21, 3361. [Google Scholar] [CrossRef]
- Murillo, D.A.; Gezan, S.A.; Heilman, A.M.; Walk, T.C.; Aparicio, J.S.; Horsley, R.D. FielDHub: A Shiny App for Design of Experiments in Life Sciences. J. Open Src. Softw. 2021, 6, 3122. [Google Scholar] [CrossRef]
- Aparicio, J.; Ariza-Suarez, D.; Raatz, B. Web Application for Spatial Modelling of Field Trials. In Proceedings of the XXIX Simposio Internacional de Estadística, Barranquilla, Colombia, 15–19 July 2019. [Google Scholar]
- Cortés, A.J.; Skeen, P.; Blair, M.W.; Chacón-Sánchez, M.I. Does the Genomic Landscape of Species Divergence in Phaseolus Beans Coerce Parallel Signatures of Adaptation and Domestication? Front. Plant Sci. 2018, 9, 1816. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.B.W.; Ellegren, H. Making sense of genomic islands of differentiation in light of speciation. Nat. Rev. Genet. 2017, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H.; Galtier, N. Determinants of genetic diversity. Nat. Rev. Genet. 2016, 17, 433. [Google Scholar] [CrossRef]
- Cortés, A.J.; Restrepo-Montoya, M.; Bedoya-Canas, L.E. Modern Strategies to Assess and Breed Forest Tree Adaptation to Changing Climate. Front. Plant Sci. 2020, 11, 1606. [Google Scholar] [CrossRef]
- Cortés, A.J.; López-Hernández, F. Harnessing Crop Wild Diversity for Climate Change Adaptation. Genes 2021, 12, 783. [Google Scholar] [CrossRef]
- Pandit, E.; Pawar, S.; Barik, S.R.; Mohanty, S.P.; Meher, J.; Pradhan, S.K. Marker-Assisted Backcross Breeding for Improvement of Submergence Tolerance and Grain Yield in the Popular Rice Variety ‘Maudamani’. Agronomy 2021, 11, 1263. [Google Scholar] [CrossRef]
- Leitão, S.T.; Bicho, M.C.; Pereira, P.; Paulo, M.J.; Malosetti, M.; de Sousa Araújo, S.; van Eeuwijk, F.; Patto, M.C.V. Common bean SNP alleles and candidate genes affecting photosynthesis under contrasting water regimes. Hortic. Res. 2021, 8, 4. [Google Scholar] [CrossRef]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Md Hatta, M.A.; Hinchliffe, A.; Steed, A. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2017, 4, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; López-Hernández, F.; Osorio-Rodriguez, D. Predicting Thermal Adaptation by Looking Into Populations’ Genomic Past. Front. Genet. 2020, 11, 1093. [Google Scholar] [CrossRef] [PubMed]
- Resende, R.T.; Piepho, H.P.; Rosa, G.J.; Silva-Junior, O.B.; e Silva, F.F.; de Resende, M.D.V.; Grattapaglia, D. Enviromics in breeding: Applications and perspectives on envirotypic-assisted selection. Theor. Appl. Genet. 2021, 134, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Arenas, S.; Cortés, A.J.; Mastretta-Yanes, A.; Jaramillo-Correa, J.P. Evaluating the accuracy of genomic prediction for the management and conservation of relictual natural tree populations. Tree Genet. Genomes 2021, 17, 12. [Google Scholar] [CrossRef]
- Cortés, A.J.; Monserrate, F.A.; Ramírez-Villegas, J.; Madriñán, S.; Blair, M.W. Drought Tolerance in Wild Plant Populations: The Case of Common Beans (Phaseolus vulgaris L.). PLoS ONE 2013, 8, e62898. [Google Scholar] [CrossRef]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortés, A.J.; Penmetsa, R.V.; Farmer, A.; Carrasquilla-Garcia, N.; Cook, D.R. A high-throughput SNP marker system for parental polymorphism screening, and diversity analysis in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2013, 126, 535–548. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield Trait | Motilonia (72 Genotypes) | Caribia (16 Genotypes) | Turipaná (61 Genotypes) | Carmen (87 Genotypes) | ||||
|---|---|---|---|---|---|---|---|---|
| Rep | Gen | Rep | Gen | Rep | Gen | Rep | Gen | |
| NS | 0.4801 | <0.0001 | 0.1 | 0.52 | 0.28 | 0.0177 | <0.0001 | <0.0001 |
| NP | < 0.0001 | <0.0001 | 0.14 | 0.38 | 0.4 | 0.001 | <0.0001 | 0.005 |
| YLP | < 0.0001 | <0.0001 | 0.04 | 0.46 | 0.07 | 0.04 | <0.0001 | <0.0001 |
| PB | < 0.0001 | <0.0001 | 0.09 | 0.38 | NA | NA | 0.052 | 0.242 |
| SB | < 0.0001 | <0.0001 | 0.46 | NA | NA | NA | <0.0001 | <0.0001 |
| VB | 0.8546 | <0.0001 | 0.13 | 0.16 | 0.03 | 0.06 | <0.0001 | <0.0001 |
| WS | NA | NA | 0.08 | 0.4 | 0.2398 | 0.42 | 0.2497 | 0.001 |
| DF | 0.7666 | <0.0001 | NA | NA | 0.08 | 0.262 | 0.442 | 0.122 |
| Parameter | Units | Repetitions Effect | Genotype Effect |
|---|---|---|---|
| A | µmol CO2 m−2 s−1 | 0.8126 | <0.0001 |
| Ca | µmol mol−1 | 0.3461 | <0.0001 |
| Ci | µmol CO2 mol−1 | 0.4298 | <0.0001 |
| gs | mol H2O m−2 s−1 | 0.4597 | <0.0001 |
| VPD | kPa | 0.2497 | <0.0001 |
| E | mmol H2O m−2 s−1 | 0.9013 | <0.0001 |
| Tleaf | °C | 0.2881 | <0.0001 |
| iWUE | NA | 0.6131 | <0.001 |
| inWUE | NA | 0.184 | <0.001 |
| iCE | NA | 0.4277 | <0.001 |
| Trait | Unit | Cluster 1 | Cluster 2 | Cluster 3 |
|---|---|---|---|---|
| NS | Seeds | 2.57 a | 2.16 a | 1.73 b |
| NP | Pods | 20.27 a | 9.21 b | 2.78 c |
| YLP | g/plant | 19.72 a | 9.05 b | 2.73 c |
| PB | G | 5.18 a | 2.49 b | 0.76 c |
| SB | G | 21.36 a | 9.05 b | 2.68 c |
| VB | G | 183.49 | 154.63 | 130.13 |
| WS | g/100 seeds | 25.46 a,b | 23.03 b | 28.89 a |
| DF | Days | 36.3 | 34.96 | 32.92 |
| E | mmol H2O m−2 s −1 | 0.012 | 0.013 | 0.012 |
| A | µmol CO2 m−2 s−1 | 24.29 a | 23.52 a | 20.15 b |
| Ca | µmol mol−1 | 358.51 b | 358.41 b | 364.04 a |
| Ci | µmol CO2 mol−1 | 284.55 | 286.45 | 289.25 |
| gs | mol H2O m−2 s−1 | 0.73 a | 0.72 a,b | 0.60 b |
| LT | °C | 35.66 | 35.99 | 36.11 |
| VPD | kPa | 1.98 b | 2.10 a,b | 2.29 a |
| iWUE | NA | 2035.99 | 1819.60 | 1698.78 |
| inWUE | NA | 35.28 | 33.76 | 35.33 |
| iCE | NA | 0.08 a | 0.08 a | 0.07 b |
| Selection Criteria | Cluster | Genotype | YLP | WS | Fe | Zn |
|---|---|---|---|---|---|---|
| 1 | G40001 | 31.43 | 13 | 66.8 | 24.3 | |
| 1 | 82 | 28.96 | 28.6 | 56.2 | 26.7 | |
| 1 | 48 | 20.55 | 50 | - | - | |
| 1 | 17 | 19.03 | 23.2 | 94.35 | 28.8 | |
| 1 | 81 | 17.7 | 27 | - | - | |
| 1 | 5 | 17.04 | 23.3 | 82.1 | 44.9 | |
| 1 | 16 | 16.78 | 23.3 | 83.1 | 41.5 | |
| 1 | 3 | 16.47 | 22.34 | 87.8 | 37.4 | |
| Yield | 1 | 18 | 15.07 | 24.7 | 79.2 | 45.4 |
| 1 | 27 | 14.18 | 19.2 | 79.5 | 41.2 | |
| 2 | 1 | 13.34 | 25.2 | 80.0 | 30.4 | |
| 2 | 84 | 13.26 | 24.5 | - | - | |
| 2 | 85 | 12.14 | 27 | 61.7 | 31.6 | |
| 2 | 9 | 11.69 | 19.8 | 103.75 | 50.1 | |
| 2 | 6 | 11.69 | 27 | 61.9 | 25.4 | |
| 2 | 39 | 10.9 | 38 | - | - | |
| 2 | 23 | 10.73 | 23.8 | 66.1 | 34.1 | |
| 2 | 13 | 10.02 | 25.6 | 82.5 | 31.6 | |
| 2 | 9 | 11.69 | 19.8 | 103.75 | 50.1 | |
| 3 | 12 | 3.14 | 27.5 | 111.25 | 40.45 | |
| 2 | 26 | 6.29 | 19.9 | 103.5 | 44.25 | |
| Fe content | 3 | 64 | 1.96 | 27 | 102.6 | 60.25 |
| 3 | 76 | 1.32 | 25.2 | 118.3 | 56.15 | |
| 3 | 77 | 2.08 | 29.2 | 120.5 | 53.6 | |
| 3 | 80 | 3.79 | 23 | 107.7 | 52.25 |
| Parameter | Unit | Cluster 1 | Cluster 2 |
|---|---|---|---|
| NP | Pods | 10.29 b | 22.37 a |
| YLP | g/plant | 3.17 b | 9.53 a |
| PB | G | 9.37 b | 24.32 a |
| SB | G | 4.98 b | 11.82 a |
| VB | G | 21.78 b | 45.28 a |
| Cluster | Units | Cluster 1 | Cluster 3 | Cluster 4 | Cluster 2 |
|---|---|---|---|---|---|
| NS | Seeds | 6.20 a | 4.55 b | 3.07 c | 2.09 d |
| NP | Pods | 103.17 a | 35.73 b | 3.07 c | 2.09 d |
| YLP | g/plant | 33.31 a | 22.59 b | 9.22 c | 2.81 d |
| Cluster | Genotype | NS | NP | YLP | VB | WS | DF |
|---|---|---|---|---|---|---|---|
| 1 | G40001 | 5.90 | 106.35 | 44.06 | 100.27 | 11.32 | 52.67 |
| 3 | 85 | 4.50 | 25.50 | 30.65 | 91.00 | 20.10 | 48.00 |
| 3 | 17 | 4.20 | 50.00 | 23.50 | 33.67 | 16.50 | 52.67 |
| 1 | 13 | 6.50 | 100.00 | 22.57 | 72.00 | 17.77 | 48.00 |
| 3 | 83 | 4.10 | 38.50 | 21.80 | 43.00 | 25.76 | 55.00 |
| 3 | 27 | 4.90 | 40.00 | 20.00 | 15.00 | 19.50 | 44.50 |
| 3 | 82 | 5.07 | 24.67 | 17.00 | 37.00 | 21.56 | 48.00 |
| 4 | 86 | 2.78 | 32.33 | 16.04 | 37.67 | 21.49 | 60.67 |
| 4 | 19 | 3.11 | 16.33 | 13.67 | 17.67 | 18.39 | 62.00 |
| 4 | 34 | 3.07 | 31.67 | 12.70 | 22.33 | 17.09 | 57.33 |
| 4 | 84 | 2.57 | 16.00 | 12.50 | 11.00 | 15.00 | 48.00 |
| 4 | 72 | 4.30 | 25.00 | 11.00 | 33.00 | 18.61 | 55.00 |
| 4 | 75 | 2.90 | 27.00 | 10.67 | 126.33 | 17.52 | 48.00 |
| 4 | 6 | 3.47 | 38.33 | 10.30 | 42.00 | 15.39 | 48.00 |
| Cluster | NS | NP | YLP | SB | VB | WS | DM |
|---|---|---|---|---|---|---|---|
| 1 | 3.46 c | 18.84 c | 5.98 c | 5.99 c | 57.60 b | 36.18 a | 76.11 a |
| 2 | 4.51 b | 23.34 b | 11.61 b | 11.61 b | 67.13 b | 30.35 b | 72.70 b |
| 3 | 5.25 a | 34.39 a | 17.89 a | 16.91 a | 87.86 a | 28.11 b | 72.37 b |
| Criteria | Cluster | Genotype | YLP | WS | Fe | Zn |
|---|---|---|---|---|---|---|
| 3 | G40001 | 27.12 | 29.47 | |||
| 3 | 21 | 21.12 | 26 | 71.5 | 36.9 | |
| 3 | 86 | 20.96 | 27.33 | 64.4 | 36.8 | |
| 3 | 82 | 20.15 | 28.33 | 53.1 | 34.2 | |
| 3 | 84 | 20.07 | 28.33 | 58 | 30.9 | |
| 3 | 81 | 19.77 | 30.33 | 70.3 | 44.4 | |
| 3 | 5 | 19.11 | 27.67 | 62 | 38 | |
| 3 | 72 | 18.7 | 28.67 | 83.6 | 41 | |
| 3 | 83 | 17.43 | 26 | 67.9 | 32.9 | |
| 3 | 9 | 16 | 26 | 79.2 | 45.5 | |
| 3 | 63 | 15.79 | 32 | 92.25 | 48.4 | |
| 3 | 67 | 15.76 | 33 | 81.8 | 51.2 | |
| 3 | 7 | 15 | 29 | 77.5 | 42 | |
| 3 | 18 | 14.9 | 24.67 | 66.4 | 33.6 | |
| 3 | 25 | 13.37 | 24 | 56.3 | 31.6 | |
| 3 | 11 | 11 | 29 | 73.6 | 43.9 | |
| 2 | 43 | 16.48 | 35.67 | 70.2 | 28.4 | |
| 2 | 62 | 15.13 | 31 | 76.6 | 42.4 | |
| 2 | 80 | 14.78 | 32.67 | 90.55 | 41.7 | |
| 2 | 77 | 14.75 | 29.42 | 109.7 | 58.4 | |
| 2 | 40 | 14.61 | 44 | 66.6 | 27 | |
| Yield | 2 | 79 | 13.96 | 26.25 | 99.9 | 56.3 |
| 2 | 29 | 13.91 | 40.67 | 63.1 | 29.7 | |
| 2 | 17 | 13.52 | 24.89 | 70.3 | 35.2 | |
| 2 | 85 | 12.72 | 28 | 53.5 | 35.9 | |
| 2 | 73 | 12.57 | 30.33 | 82.3 | 46.3 | |
| 2 | 26 | 12.23 | 25.33 | 74.9 | 35 | |
| 2 | 69 | 12.03 | 33.67 | 92 | 46.8 | |
| 2 | 41 | 11.99 | 48.33 | 64.4 | 34.1 | |
| 2 | 16 | 11.95 | 28.67 | 64.6 | 35.8 | |
| 2 | 13 | 11.9 | 26 | 75.1 | 45 | |
| 2 | 36 | 11.81 | 47.02 | 73.5 | 33.5 | |
| 2 | 28 | 11.7 | 26.03 | 56.4 | 35.4 | |
| 2 | 24 | 11.35 | 25 | 61.1 | 32.1 | |
| 2 | 65 | 11.02 | 29.33 | 93.15 | 58.7 | |
| 2 | 23 | 10.92 | 25.62 | 62.6 | 35.5 | |
| 2 | 19 | 10.72 | 25 | 72.3 | 40.4 | |
| 2 | 22 | 10.62 | 24.33 | 82.2 | 39 | |
| 2 | 64 | 10.34 | 30 | 101.1 | 50.4 | |
| 2 | 3 | 10.14 | 27.7 | 71.4 | 38.1 | |
| 2 | 71 | 10.09 | 30.67 | 92.65 | 46.4 | |
| 1 | 45 | 9.42 | 41 | 65.4 | 39.2 | |
| 1 | 56 | 9.03 | 47.87 | 74 | 38.4 | |
| 1 | 35 | 8.24 | 44 | 74.3 | 39.5 | |
| 1 | 47 | 8.15 | 46.75 | 74.3 | 37.4 | |
| WS | 1 | 37 | 7.36 | 40.04 | 61.7 | 41.9 |
| 1 | 49 | 7.09 | 39.78 | 74.7 | 33.1 | |
| 1 | 50 | 7.02 | 41.42 | 96.5 | 33.9 | |
| 1 | 48 | 6.76 | 40.89 | 65.8 | 37.9 | |
| 1 | 53 | 6.58 | 39.91 | 57.8 | 37.5 | |
| 1 | 14 | 7.68 | 23.49 | 101.4 | 50.2 | |
| Fe content | 1 | 78 | 7.52 | 25.33 | 104.4 | 57 |
| 2 | 6 | 8.4 | 28.66 | 99.45 | 41.6 | |
| 2 | 76 | 7.44 | 27 | 106.05 | 57.9 |
| Genotypes | ASV | YSI | rASV | rYSI | Square Root | Yield (g/plant) |
|---|---|---|---|---|---|---|
| 13 | 1.08 | 10 | 5 | 5 | 3.26 | 12.14 |
| 27 | 0.94 | 10 | 4 | 6 | 3.17 | 11.54 |
| 28 | 1.1 | 14 | 6 | 8 | 2.61 | 7.43 |
| 68 | 0.80 | 12 | 3 | 9 | 2.10 | 5.62 |
| 82 | 1.43 | 11 | 8 | 3 | 3.83 | 17.32 |
| 84 | 0.74 | 6 | 2 | 4 | 3.51 | 13.22 |
| 85 | 1.69 | 11 | 9 | 2 | 3.88 | 17.06 |
| G40001 | 0.12 | 2 | 1 | 1 | 5.13 | 28.16 |
| 9 | 1.36 | 14 | 7 | 7 | 2.76 | 8.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burbano-Erazo, E.; León-Pacheco, R.I.; Cordero-Cordero, C.C.; López-Hernández, F.; Cortés, A.J.; Tofiño-Rivera, A.P. Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought Tolerance. Agronomy 2021, 11, 1978. https://doi.org/10.3390/agronomy11101978
Burbano-Erazo E, León-Pacheco RI, Cordero-Cordero CC, López-Hernández F, Cortés AJ, Tofiño-Rivera AP. Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought Tolerance. Agronomy. 2021; 11(10):1978. https://doi.org/10.3390/agronomy11101978
Chicago/Turabian StyleBurbano-Erazo, Esteban, Rommel Igor León-Pacheco, Carina Cecilia Cordero-Cordero, Felipe López-Hernández, Andrés J. Cortés, and Adriana Patricia Tofiño-Rivera. 2021. "Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought Tolerance" Agronomy 11, no. 10: 1978. https://doi.org/10.3390/agronomy11101978
APA StyleBurbano-Erazo, E., León-Pacheco, R. I., Cordero-Cordero, C. C., López-Hernández, F., Cortés, A. J., & Tofiño-Rivera, A. P. (2021). Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought Tolerance. Agronomy, 11(10), 1978. https://doi.org/10.3390/agronomy11101978