Non-Inversion Tillage as a Chance to Increase the Biodiversity of Ground-Dwelling Spiders in Agroecosystems: Preliminary Results

 , , , and
, , , and
Abstract
:1. Introduction
- Ploughing causes a decrease in the abundance, species richness, and biodiversity of spiders in the growing season;
- In fields under the non-inversion tillage system, which are less disturbed, there are more active epigeic hunters (cursorials), whereas web spiders are more abundant in conventional fields.
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
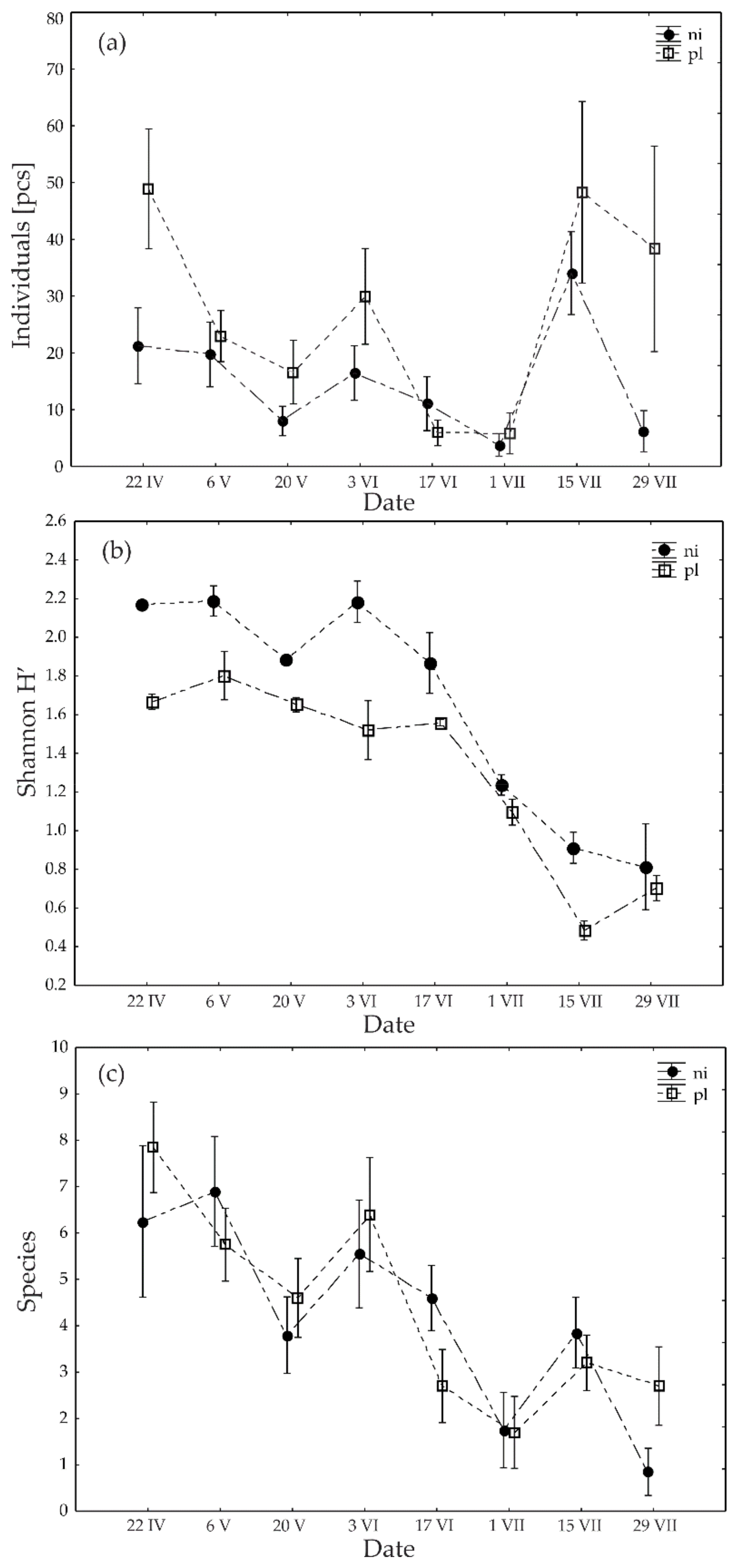
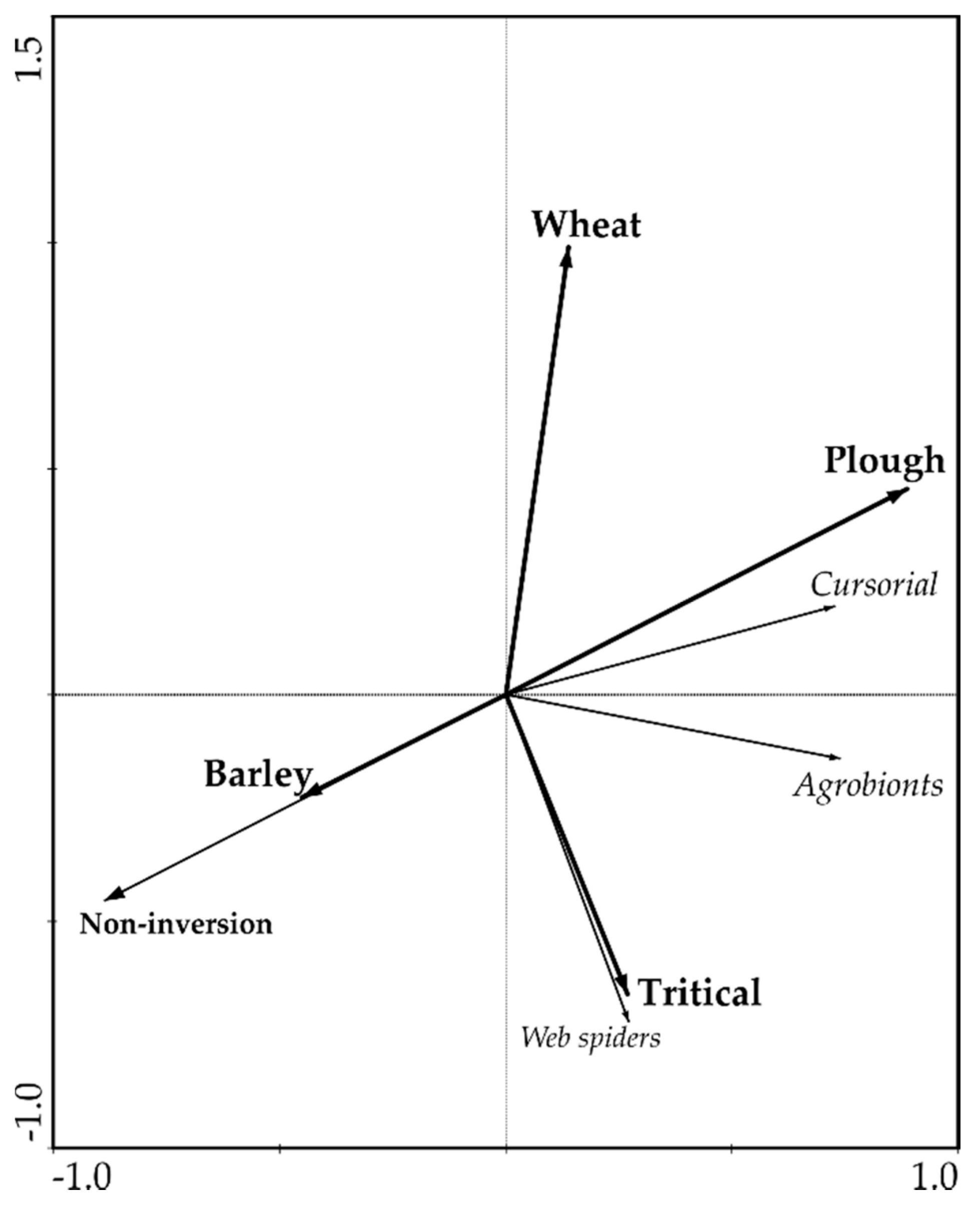
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duru, M.; Therond, O.; Martin, G.; Martin-Clouaire, R.; Magne, M.A.; Justes, E.; Journet, E.P.; Aubertot, J.N.; Savary, S.; Bergez, J.E.; et al. How to implement biodiversity-based agriculture to enhance ecosystem services: A review. Agron. Sustain. Dev. 2015, 35, 1259–1281. [Google Scholar] [CrossRef]
- Downie, I.S.; Ribera, I.; McCracken, D.I.; Wilson, W.L.; Foster, G.N.; Waterhouse, A.; Abernethy, V.J.; Murphy, K.J. Modelling populations of Erigone atra and E. dentipalpis (Araneae: Linyphiidae) across an agricultural gradient in Scotland. Agric. Ecosyst. Environ. 2000, 80, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.M.; Reynolds, C.J.M. The impact of soil cultivation on arthropod (Coleoptera and Araneae) emergence on arable land. Pedobiologia 2003, 47, 181–191. [Google Scholar] [CrossRef]
- Holland, J.M. The environmental consequences of adopting conservation tillage in Europe: Reviewing the evidence. Agr. Ecosyst. Environ. 2004, 103, 1–25. [Google Scholar] [CrossRef]
- Thorbek, P.; Bilde, T. Reduced numbers of generalist arthropod predators after crop management. J. Appl. Ecol. 2004, 41, 526–538. [Google Scholar] [CrossRef]
- Prieto-Benítez, S.; Méndez, M. Effects of land management on the abundance and richness of spiders (Araneae): A meta-analysis. Biol. Conserv. 2011, 144, 683–691. [Google Scholar] [CrossRef]
- Kosewska, A.; Skalski, T.; Nietupski, M. Effect of conventional and non-inversion tillage systems on the abundance and some life history traits of carabid beetles (Coleoptera: Carabidae) in winter triticale fields. Eur. J. Entomol. 2014, 111, 669–676. [Google Scholar] [CrossRef]
- Thies, C.; Haenke, C.; Scherber, S.; Bengtsson, J.; Bommarco, R.; Clement, L.W.; Ceryngier, P.; Dennis, C.; Emmerson, M.; Gagic, V.; et al. The relationship between agricultural intensification and biological control: Experimental tests across Europe. Ecol. Appl. 2011, 21, 2187–2196. [Google Scholar] [CrossRef]
- Samu, F.; Szinetar, C.; Szita, E.; Fetyko, K.; Neidert, D. Regional variations in agrobiont composition and agrobiont life history of spiders (Araneae) within Hungary. Arachnol. Mitt. 2011, 40, 105–109. [Google Scholar] [CrossRef]
- Birkhofer, K.; Entling, M.H.; Lubin, Y. Agroecology: Trait composition, spatial relationships, trophic interactions. In Spider Research in the 21st Century: Trends & Perspectives; Penney, D., Ed.; SIRI Scientific Press: Rochdale, UK, 2013; pp. 220–228. [Google Scholar]
- Rusch, A.; Birkhofer, K.; Bommmarco, R.; Smith, H.G.; Ekbom, B. Management intensity at field and landscape levels affects the structure of generalist predator communities. Oecologia 2014, 175, 971–983. [Google Scholar] [CrossRef]
- Hänggi, A.; Stöeckli, E.; Nentwig, W. Habitats of Central European Spiders; Miscellanea Faunistica Helvetiae; Musée d’Histoire Naturelle de Neuchatel: Neuchâtel, Switzerland, 1995; Volume 4, p. 460. ISBN 978-288-414-00-89. [Google Scholar]
- Nyffeler, M.; Sunderland, K.D. Composition, abundance and pest control potential of spider communities in agroecosystems: A comparison of European and US studies. Agric. Ecosyst. Environ. 2003, 95, 579–612. [Google Scholar] [CrossRef]
- Michalko, R.; Pekar, S. Different hunting strategies of generalist predators result in functional differences. Oecologia 2016, 181, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Łuczak, J. Spiders in agrocoenoses. Pol. Ecol. Stud. 1979, 5, 151–200. [Google Scholar]
- Samu, F.; Szinetar, C. On the nature of agrobiont spiders. J. Arachnol. 2002, 30, 389–402. [Google Scholar] [CrossRef]
- Lang, A. Intraguild interference and biocontrol effects of generalist predators in a winter wheat field. Oecologia 2003, 134, 144–153. [Google Scholar] [CrossRef]
- Öberg, S.; Ekbom, B.; Bommarco, R. Influence of habitat type and surrounding landscape on spider diversity in Swedish agroecosystems. Agric. Ecosyst. Environ. 2007, 122, 211–219. [Google Scholar] [CrossRef]
- Harwood, J.D.; Sunderland, K.D.; Symondson, W.O.C. Prey selection by linyphiid spiders: Molecular tracking of the effects of alternative prey on rates of aphid consumption in the field. Mol. Ecol. 2004, 13, 3549–3560. [Google Scholar] [CrossRef]
- Harwood, J.D.; Sunderland, K.D.; Symondson, W.O.C. Monoclonal antibodies reveal the potential of the tetragnathids spider Pachygnatha degeeri (Araneae: Tetragnathidae) as an aphid predator. Bull. Entomol. Res. 2005, 95, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.H.; Roschewitz, I.; Thies, C.; Tscharntke, T. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. J. Appl. Ecol. 2005, 42, 281–287. [Google Scholar] [CrossRef]
- Chapman, E.G.; Schmidt, J.M.; Welch, K.D.; Harwood, J.D. Molecular evidence for dietary selectivity and pest suppression potential in an epigeal spider community in winter wheat. Biol. Contr. 2013, 65, 72–86. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, H.M. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Entling, M.H.; Stämpfli, K.; Ovaskainen, O. Increased propensity for aerial dispersal in disturbed habitats due to intraspecific variation and species turnover. Oikos 2011, 120, 1099–1109. [Google Scholar] [CrossRef]
- Gallé, R.; Császár, P.; Makra, T.; Gallé-Szpisjak, N.; Ladányi, Z.; Torma, A.; Ingle, K.; Szilassi, P. Small-scale agricultural landscapes promote spider and ground beetle densities by offering suitable overwintering sites. Landsc. Ecol. 2018, 33, 1435–1446. [Google Scholar] [CrossRef]
- Topa, E. The impact of ploughing and zero tillage soil cultivation on species diversity of ground-dwelling spiders in the cultivation of winter rape. Prog. Plant Prot. 2020, 60, 247–253. (In Polish) [Google Scholar] [CrossRef]
- Duffey, E. Ecological strategies in spiders including some characteristics of species in pioneer and mature habitats. Symp. Zool. Soc. Lond. 1978, 42, 109–123. [Google Scholar]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 5. 2021. Available online: https://www.araneae.nmbe.ch (accessed on 1 May 2021).
- Uetz, G.W.; Halaj, J.; Cady, A.B. Guild structure of spiders in major crops. J. Arachnol. 1999, 27, 270–280. [Google Scholar]
- Górny, M.; Grüm, L. Metody Stosowane w Zoologii Gleby; PWN: Warszawa, Poland, 1981; p. 482. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Roberts, D.W. Package “Labdsv”. 2012. Available online: http://cran.rproject.org/web/packages/labdsv/labdsv.pdf (accessed on 1 September 2021).
- Marc, P.; Canard, A.; Ysnel, F. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 1999, 74, 229–273. [Google Scholar] [CrossRef]
- Schmidt, J.M.; Harwood, J.D.; Rypstra, A.L. Foraging activity of a dominant epigeal predator: Molecular evidence for the effect of prey density on consumption. Oikos 2012, 121, 1715–1724. [Google Scholar] [CrossRef]
- Michalko, R.; Pekar, S.; Entling, M.H. An updated perspective on spiders as generalist predators in biological control. Oecologia 2019, 189, 21–36. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Tscharntke, T. The role of perennial habitats for Central European farmland spiders. Agric. Ecosyst. Environ. 2005, 105, 235–242. [Google Scholar] [CrossRef]
- Glück, E.; Ingrisch, S. The effect of bio-dynamic and conventional agriculture management on Erigoninae and Lycosidae spiders. J. Appl. Entomol. 1990, 110, 136–148. [Google Scholar] [CrossRef]
- Wolak, M. The significance of unmanaged “island” habitats for epigeic spiders in a uniform agricultural landscape. In European Arachnology 2002, Proceedings of the 20th European Colloquium of Arachnology, Szombathely, Budapest, Hungary, 22–26 July 2002; Samu, F., Szinetár, C., Eds.; Plant Protection Institute: Budapest, Hungary, 2004; pp. 327–336. [Google Scholar]
- Pfiffner, L.; Luka, H. Effects of low-input farming systems on carabids and epigeal spiders—A paired farm approach. Basic. Appl. Ecol. 2003, 4, 117–127. [Google Scholar] [CrossRef]
- Clough, Y.; Kruess, A.; Kleijn, D.; Tscharntke, T. Spider diversity in cereal fields: Comparing factors at local, landscape and regional scales. J. Biogeogr. 2005, 32, 2007–2014. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Tscharntke, T. Landscape context of sheetweb spider (Araneae: Linyphiidae) abundance in cereal fields. J. Biogeogr. 2005, 32, 467–473. [Google Scholar] [CrossRef]
- Lemke, A.; Poehling, H.M. Sown weed strips in cereal fields: Overwintering site and “source” habitat for Oedothorax apicatus (Blackwall) and Erigone atra (Blackwall) (Araneae: Erigonidae). Agric. Ecosyst. Environ. 2002, 90, 67–80. [Google Scholar] [CrossRef]
- Kajak, A.; Oleszczuk, M. Effect of shelterbelts on adjoining cultivated fields: Patrolling intensity of Carabid beetles (Carabidae) and spiders (Araneae). Pol. J. Ecol. 2004, 52, 155–172. [Google Scholar]
- Harwood, J.D.; Sunderland, K.D.; Symondson, W.O.C. Living where the food is: Web-location by linyphiid spiders in relation to prey availability in winter wheat. J. Appl. Entomol. 2001, 38, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, A.M.; Bohan, D.A.; Bell, J.R. Ballooning dispersal in arthropod taxa with convergent behaviours: Dynamic properties of ballooning silk in turbulent flows. Biol. Lett. 2006, 2, 371–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schellhorn, N.A.; Bianchi, F.J.; Hsu, C.L. Movement of entomophagous arthropods in agricultural landscapes: Links to pest suppression. Annu. Rev. Entomol. 2014, 59, 559–581. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Walters, B.J.; Cunningham, S.A.; Hemerik, L.; Schellborn, N.A. Landscape-scale mass-action of spiders explains early-season immigration rates in crops. Landsc. Ecol. 2017, 32, 1257–1267. [Google Scholar] [CrossRef] [Green Version]
- Wolak, M. The spider fauna of balks. In Proceedings of the European Arachnology 2000—19th European Colloquium of Arachnology, Århus, Denmark, 17–22 July 2000; pp. 229–326. [Google Scholar]
- Woolley, C.; Thomas, C.F.G.; Blackshaw, R.P.; Goodacre, S.L. Aerial dispersal activity of spiders sampled from farmland in southern England. J. Arachnol. 2016, 44, 347–358. [Google Scholar] [CrossRef]
- Plath, E.; Rischen, T.; Mohr, T.; Fischer, K. Biodiversity in agricultural landscapes: Grassy field margins and semi-natural fragments both foster spider diversity and body size. Agric. Ecosyst. Environ. 2021, 316, 107457. [Google Scholar] [CrossRef]
- Thorbek, P.; Sunderland, K.D.; Topping, C. Reproductive biology of agrobiont linyphiid spiders in relation to habitat, season and biocontrol potential. Biol. Control. 2004, 30, 193–202. [Google Scholar] [CrossRef]
- Rusch, A.; Birkhofer, K.; Bommarco, R.; Smith, H.G.; Ekbom, B. Predator body sizes and habitat preferences predict predation rates in an agroecosystem. Basic. Appl. Ecol. 2015, 16, 250–259. [Google Scholar] [CrossRef]
- Pedley, S.M.; Dolman, P.M. Multi-taxa trait and functional responses to physical disturbance. J. Anim. Ecol. 2014, 83, 1542–1552. [Google Scholar] [CrossRef] [Green Version]
- Birkhofer, K.; Gavish-Regev, E.; Endlweber, K.; Lubin, Y.D.; von Berg, K.; Wise, D.H.; Scheu, S. Cursorial spiders retard initial aphid population growth at low densities in winter wheat. Bull. Entomol. 2008, 98, 249–255. [Google Scholar] [CrossRef]
- Michałko, R.; Birkhofer, K. Habitat niches suggest that non-crop habitat types differ in quality as source habitats for Central European agrobiont spiders. Agric. Ecosyst. Environ. 2021, 308, 107248. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination | |||||
|---|---|---|---|---|---|
| T pl | W pl | T ni | B ni | ||
| Cereal | Triticale | Wheat | Triticale | Barley | |
| Cultivar | Baltico | Bamberka | Elpaso | Titus | |
| DS 1 | 7 September 2015 | 15 September 2015 | 10 September 2015 | 17 September 2015 | |
| Area | 38 ha | 21.5 ha | 60 ha | 22 ha | |
| Herbicide | AS 2 | metribuzin + flufenacet + chlorsulfuron | metribuzin + flufenacet + chlorsulfuron carfentrazone -ethyl | chlorsulfuron + metribuzin + isoproturon | chlorsulfuron |
| DA 3 | 14 September 2015 | 21 September 2015 | 5 October 2015 | 24 September 2015 | |
| AS | pinoxaden | carfentrazone-ethyl | methyl sodium iodosulfuron + methyl mesosulfuron | pinoxaden | |
| DA | 26 April 2016 | 30 March 2016 | 25 April 2016 | 25 April 2016 | |
| Fungicide | AS | propiconazole + fenpropidin | propiconazole + fenpropidin | prothioconazole + spiroxamine | prothioconazole + spiroxamine |
| DA | 30 April 2016 | 30 April 2016 | 16 May 2016 | 16 May 2016 | |
| AS | tebuconazole | epoxiconazole | fluoxastrobin + prothioconazole | fluoxastrobin + prothioconazole | |
| DA | 2 June 2016 | 27 May 2016 | 15 June 2016 | 15 June 2016 | |
| AS | tebuconazole | ||||
| DA | 26 June 2016 | ||||
| Insecticide | AS | deltamethrin | deltamethrin | deltamethrin | deltamethrin |
| DA | 15 June 2016 | 15 June 2016 | 15 June 2016 | 15 June 2016 | |
| NPK fertilisation | DA | 3 March, 12 April, 18 May and 2 June 2016 | 29 March, 12 April and 27 May 2016 | 17 September 2015, 15 April, 28 April 2016 | 9 October 2015, 15 April, 28 April 2016 |
| Family | Species | Ecological Description 1 | Cereal Soil Tillage | |||
|---|---|---|---|---|---|---|
| T ni | B ni | T pl | W pl | |||
| Linyphiidae | Oedothorax apicatus (Blackwall, 1850) | A, C | 553 | 511 | 1711 | 975 |
| Linyphidae | Erigone atra (Blackwall, 1833) | A, W | 178 | 130 | 293 | 38 |
| Linyphidae | Erigone dentipalpis (Wider, 1834) | A, W | 121 | 182 | 216 | 15 |
| Lycosidae | Pardosa prativaga (L. Koch, 1870) | A, C | 98 | 89 | 90 | 220 |
| Lycosidae | Pardosa paludicola (Clerck, 1758) | C | 8 | 47 | 33 | 188 |
| Lycosidae | Pardosa palustris (Linnaeus, 1758) | A, C | 32 | 108 | 44 | 89 |
| Lycosidae | Trochosa ruricola (De Geer, 1778) | A, C | 21 | 24 | 9 | 51 |
| Thomisidae | Xysticus kochi (Thorell, 1872) | A, C | 6 | 30 | 3 | 42 |
| Linyphiidae | Bathyphantes gracilis (Blackwall, 1841) | A, W | 36 | 26 | 15 | 3 |
| Tetragnathidae | Pachygnatha degeeri (Sundevall, 1830) | A, C | 11 | 26 | 27 | 15 |
| Lycosidae | Pardosa agrestis (Westring, 1861) | A, C | 7 | 16 | 13 | 21 |
| Lycosidae | Pardosa pullata (Clerck, 1758) | A, C | 9 | 17 | 17 | 12 |
| Linyphiidae | Oedothorax retusus (Westring, 1851) | A, C | 10 | 0 | 17 | 27 |
| Tetragnathidae | Pachygnatha clercki (Sundevall, 1823) | A, C | 2 | 1 | 14 | 9 |
| Tetragnathidae | Tetragnatha extensa (Linnaeus, 1758) | A, W | 5 | 3 | 18 | 0 |
| Linyphiidae | Tenuiphantes tenebricola (Wider, 1834) | W | 0 | 11 | 0 | 6 |
| Linyphiidae | Araeoncus humilis (Blackwall, 1841) | A, W | 3 | 4 | 4 | 2 |
| Linyphiidae | Dicymbium nigrum brevisetosum (Locket, 1962) | W | 1 | 0 | 12 | 0 |
| Lycosidae | Pardosa amentata (Clerck, 1757) | A, C | 0 | 7 | 0 | 6 |
| Lycosidae | Pardosa lugubris (Walckenaer, 1802) | C | 1 | 6 | 1 | 4 |
| Thomisidae | Xysticus cristatus (Clerck, 1758) | A, C | 1 | 5 | 2 | 2 |
| Lycosidae | Pardosa riparia (C.L. Koch, 1833) | C | 2 | 4 | 2 | 1 |
| Pisauridae | Pisaura mirabilis (Clerck, 1758) | C | 1 | 1 | 2 | 5 |
| Lycosidae | Alopecosa cuneata (Clerck, 1758) | C | 0 | 5 | 1 | 2 |
| Clubionidae | Clubiona reclusa (O.P.-Cambridge, 1863) | C | 0 | 0 | 1 | 7 |
| Thomisidae | Ozyptila trux (Blackwall, 1846) | C | 1 | 3 | 3 | 1 |
| Lycosidae | Trochosa terricola (Thorell, 1856) | C | 1 | 0 | 1 | 6 |
| Linyphiidae | Neriene clathrata (Sundevall, 1830) | W | 0 | 4 | 0 | 3 |
| Linyphiidae | Stemonyphantes lineatus (Linnaeus, 1758) | W | 1 | 0 | 0 | 6 |
| Linyphidae | Diplostyla concolor (Wider, 1834) | A, W | 1 | 1 | 2 | 2 |
| Linyphiidae | Savignia frontata (Blackwall, 1833) | W | 1 | 1 | 3 | 0 |
| Miturgidae | Zora spinimana (Sundevall, 1833) | C | 1 | 2 | 0 | 1 |
| Hahniidae | Hahnia pusilla (C.L. Koch, 1841) | W | 0 | 1 | 0 | 2 |
| Linyphiidae | Porrhomma pygmaeum (Blackwall, 1834) | W | 1 | 1 | 1 | 0 |
| Thomisidae | Xysticus ulmi (Hahn, 1831) | A, C | 1 | 1 | 0 | 1 |
| Gnaphosidae | Zelotes subterraneus (C.L. Koch, 1833) | C | 1 | 2 | 0 | 0 |
| Linyphiidae | Agyneta affinis (Kulczyński, 1898) | A, W | 2 | 0 | 0 | 0 |
| Lycosidae | Alopecosa pulverulenta (Clerck, 1758) | A, C | 1 | 1 | 0 | 0 |
| Linyphiidae | Bathyphantes parvulus (Westring, 1851) | W | 1 | 0 | 0 | 1 |
| Linyphiidae | Centromerus sylvaticus (Blackwall, 1841) | W | 1 | 0 | 1 | 0 |
| Clubionidae | Clubiona subtilis (L. Koch, 1867) | C | 2 | 0 | 0 | 0 |
| Linyphidae | Erigonella hiemalis (Blackwall, 1841) | W | 0 | 2 | 0 | 0 |
| Araneidae | Larinioides patagiatus (Clerck, 1758) | W | 0 | 2 | 0 | 0 |
| Linyphiidae | Pocadicnemis juncea (Locket and Millidge, 1953) | W | 1 | 1 | 0 | 0 |
| Theridiidae | Robertus lividus (Blackwall, 1836) | W | 1 | 0 | 1 | 0 |
| Lycosidae | Trochosa spinipalpis (F.P.-Cambridge, 1895) | C | 0 | 0 | 1 | 1 |
| Linyphiidae | Walckenaeria antica (Wider, 1834) | W | 0 | 0 | 2 | 0 |
| Linyphiidae | Walckenaeria nudipalpis (Westring, 1851) | W | 0 | 0 | 2 | 0 |
| Linyphiidae | Agyneta rurestris (C. L. Koch, 1836) | A, W | 0 | 1 | 0 | 0 |
| Araneidae | Argiope bruennichi (Scopoli, 1772) | W | 0 | 0 | 0 | 1 |
| Linyphiidae | Ceratinella brevis (Wider, 1834) | W | 0 | 1 | 0 | 0 |
| Clubionidae | Clubiona neglecta (O.P.-Cambridge, 1862) | C | 0 | 1 | 0 | 0 |
| Gnaphosidae | Drassyllus lutetianus (L. Koch, 1866) | C | 0 | 0 | 0 | 1 |
| Mimetidae | Ero furcata (Villers, 1789) | W | 0 | 1 | 0 | 0 |
| Linyphiidae | Kaestneria pullata (O.P.-Cambridge, 1863) | W | 1 | 0 | 0 | 0 |
| Araneidae | Mangora acalypha (Walckenaer, 1802) | W | 0 | 1 | 0 | 0 |
| Araneidae | Neoscona adianta (Walckenaer, 1802) | W | 0 | 1 | 0 | 0 |
| Linyphiidae | Oedothorax fuscus (Blackwall, 1841) | A, C | 0 | 1 | 0 | 0 |
| Linyphiidae | Oedothorax gibbosus (Blackwall, 1841) | C | 0 | 0 | 0 | 1 |
| Thomisidae | Ozyptila praticola (C.L. Koch, 1837) | C | 0 | 1 | 0 | 0 |
| Lycosidae | Piratula uliginosa (Thorell, 1856) | C | 0 | 0 | 0 | 1 |
| Linyphiidae | Tallusia experta (O.P.-Cambridge, 1871) | W | 1 | 0 | 0 | 0 |
| Linyphiidae | Tenuiphantes cristatus (Menge, 1866) | W | 1 | 0 | 0 | 0 |
| Philodromidae | Thanatus striatus (C.L. Koch, 1845) | C | 0 | 0 | 1 | 0 |
| Philodromidae | Tibellus maritimus (Menge, 1874) | C | 0 | 0 | 1 | 0 |
| Philodromidae | Tibellus oblongus (Walckenaer, 1802) | C | 0 | 0 | 1 | 0 |
| Gnaphosidae | Zelotes petrensis (C.L. Koch, 1839) | C | 0 | 0 | 0 | 1 |
| Total Individuals | 1127 | 1283 | 2565 | 1769 | ||
| 2410 | 4334 | |||||
| Total Species | 40 | 43 | 36 | 38 | ||
| 55 | 49 | |||||
| Shannon H’ Log Base 2.718 | 1.814 | 2.177 | 1.35 | 1.744 | ||
| 2.052 | 1.613 | |||||
| Shannon J’ | 0.492 | 0.579 | 0.377 | 0.479 | ||
| 0.512 | 0.415 | |||||
| df | Wald Stat. | p | |
|---|---|---|---|
| Abundance | |||
| Date | 7 | 1728.20 | 0.00 |
| Tillage | 1 | 266.73 | 0.00 |
| Date × Tillage | 7 | 357.00 | 0.00 |
| Richness | |||
| Date | 7 | 240.19 | 0.00 |
| Tillage | 1 | 2.35 | 0.13 |
| Date × Tillage | 7 | 36.22 | 0.00 |
| Shannon H’ | |||
| Date | 7 | 49.12 | 0.00 |
| Tillage | 1 | 6.36 | 0.01 |
| Date × Tillage | 7 | 1.80 | 0.97 |
| Agrobionts | |||
| Date | 7 | 1642.65 | 0.00 |
| Tillage | 1 | 244.14 | 0.00 |
| Date × Tillage | 7 | 327.54 | 0.00 |
| Web spiders | |||
| Date | 7 | 379.26 | 0.00 |
| Tillage | 1 | 0.60 | 0.44 |
| Date × Tillage | 7 | 103.30 | 0.00 |
| Cursorial | |||
| Date | 7 | 1402.70 | 0.00 |
| Tillage | 1 | 388.50 | 0.00 |
| Date × Tillage | 7 | 242.51 | 0.00 |
| Dominance Class | |
|---|---|
| Non-Inversion Tillage | Ploughing Tillage |
| Superdominant species (>30%) | |
| Oedothorax apicatus (44.15%) | Oedothorax apicatus (61.98%) |
| Eudominant species (10–30%) | |
| Erigone atra (12.78%), Erigone dentipalpis (12.57%) | |
| Dominant species (5–10%) | |
| Pardosa prativaga (7.76%), Pardosa palustris (5.81%) | Erigone atra (7.64%), Pardosa prativaga (7.15%), Erigone dentipalpis (5.33%), Pardosa paludicola (5.1%) |
| Subdominant species (2–5%) | |
| Bathyphantes gracilis (2.57%), Pardosa paludicola (2.28%) | Pardosa palustris (3.07%) |
| Recedent species (1–2%) | |
| Trochosa ruricola (1.87%), Pachygnatha degeeri (1.54%), Xysticus kochi (1.49%), Pardosa pullata (1.08%) | Trochosa ruricola (1.38%), Xysticus kochi (1.04%), Oedothorax retusus (1.02%) |
| Subrecedent species (<1%) | |
| The remaining 45 species (5.15%) | The remaining 40 species (6.3%) |
| IndVal | p | Frequency | |
|---|---|---|---|
| Non-Inversion | |||
| Erigone dentipalpis | 32.3 | 0.012 | 150 |
| Bathyphantes gracilis | 18.4 | 0.001 | 54 |
| Plough | |||
| Oedothorax apicatus | 60.9 | 0.001 | 258 |
| Pardosa prativaga | 30.4 | 0.026 | 137 |
| Pardosa paludicola | 21.0 | 0.001 | 62 |
| Pachygnatha clercki | 9.4 | 0.001 | 20 |
| Dicymbium nigrum brevisetosum | 5.2 | 0.007 | 10 |
| Clubiona reclusa | 4.4 | 0.018 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topa, E.; Kosewska, A.; Nietupski, M.; Trębicki, Ł.; Nicewicz, Ł.; Hajdamowicz, I. Non-Inversion Tillage as a Chance to Increase the Biodiversity of Ground-Dwelling Spiders in Agroecosystems: Preliminary Results. Agronomy 2021, 11, 2150. https://doi.org/10.3390/agronomy11112150
Topa E, Kosewska A, Nietupski M, Trębicki Ł, Nicewicz Ł, Hajdamowicz I. Non-Inversion Tillage as a Chance to Increase the Biodiversity of Ground-Dwelling Spiders in Agroecosystems: Preliminary Results. Agronomy. 2021; 11(11):2150. https://doi.org/10.3390/agronomy11112150
Chicago/Turabian StyleTopa, Elżbieta, Agnieszka Kosewska, Mariusz Nietupski, Łukasz Trębicki, Łukasz Nicewicz, and Izabela Hajdamowicz. 2021. "Non-Inversion Tillage as a Chance to Increase the Biodiversity of Ground-Dwelling Spiders in Agroecosystems: Preliminary Results" Agronomy 11, no. 11: 2150. https://doi.org/10.3390/agronomy11112150
APA StyleTopa, E., Kosewska, A., Nietupski, M., Trębicki, Ł., Nicewicz, Ł., & Hajdamowicz, I. (2021). Non-Inversion Tillage as a Chance to Increase the Biodiversity of Ground-Dwelling Spiders in Agroecosystems: Preliminary Results. Agronomy, 11(11), 2150. https://doi.org/10.3390/agronomy11112150