
Field Screening of Wheat Advanced Lines for Salinity Tolerance


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Experimental Site and Agronomic Practices
2.3. Measured Traits
2.4. Statistical Analysis
3. Results
3.1. ANOVA and Descriptive Evaluation of F6–F8
3.2. Genetic Variability in F6–F8
3.3. Genetic Gain and Selection Differential
3.4. Mean Performance of Promising Lines
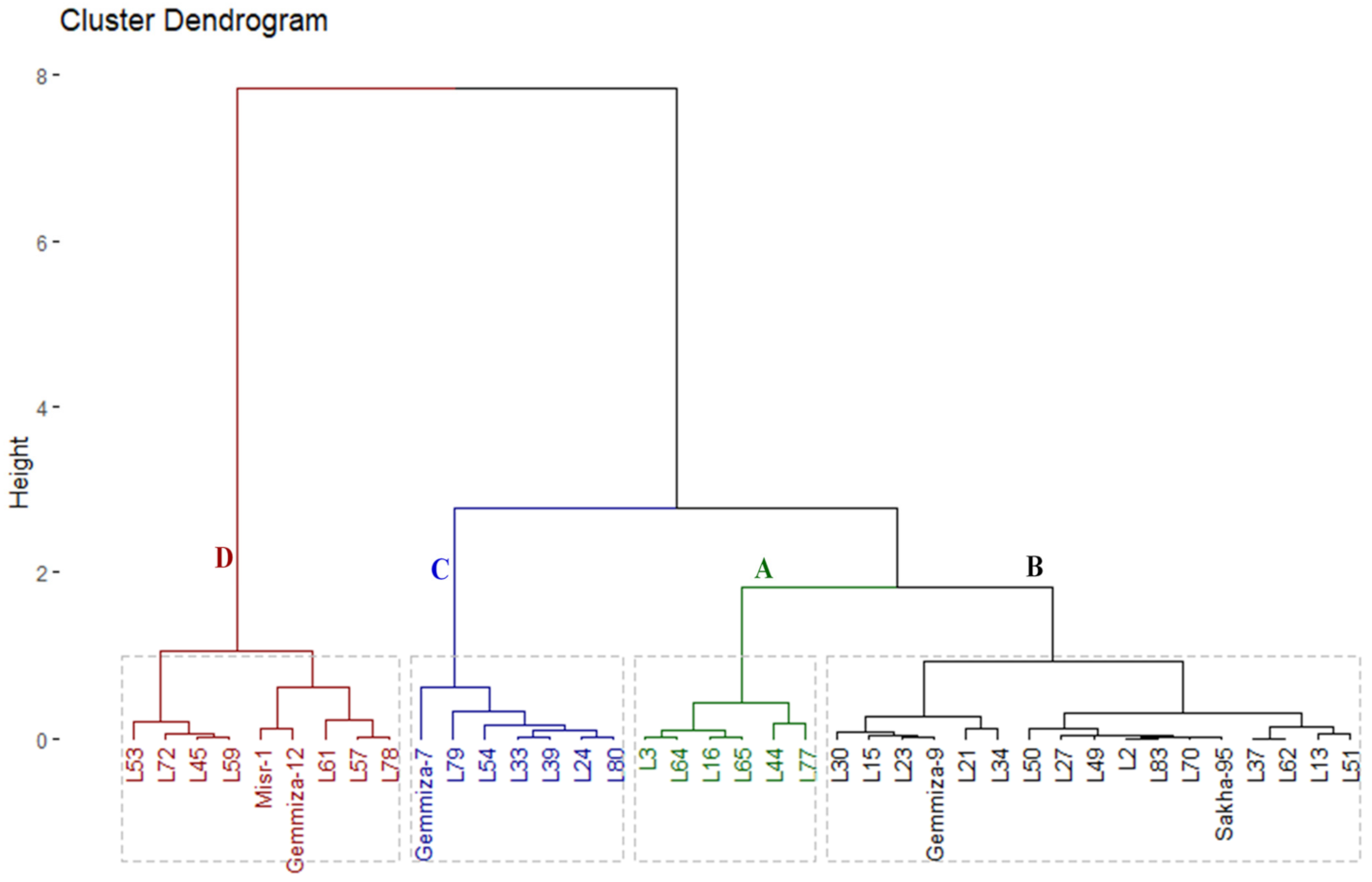
3.5. Genotypic Classification according to Salinity Tolerance
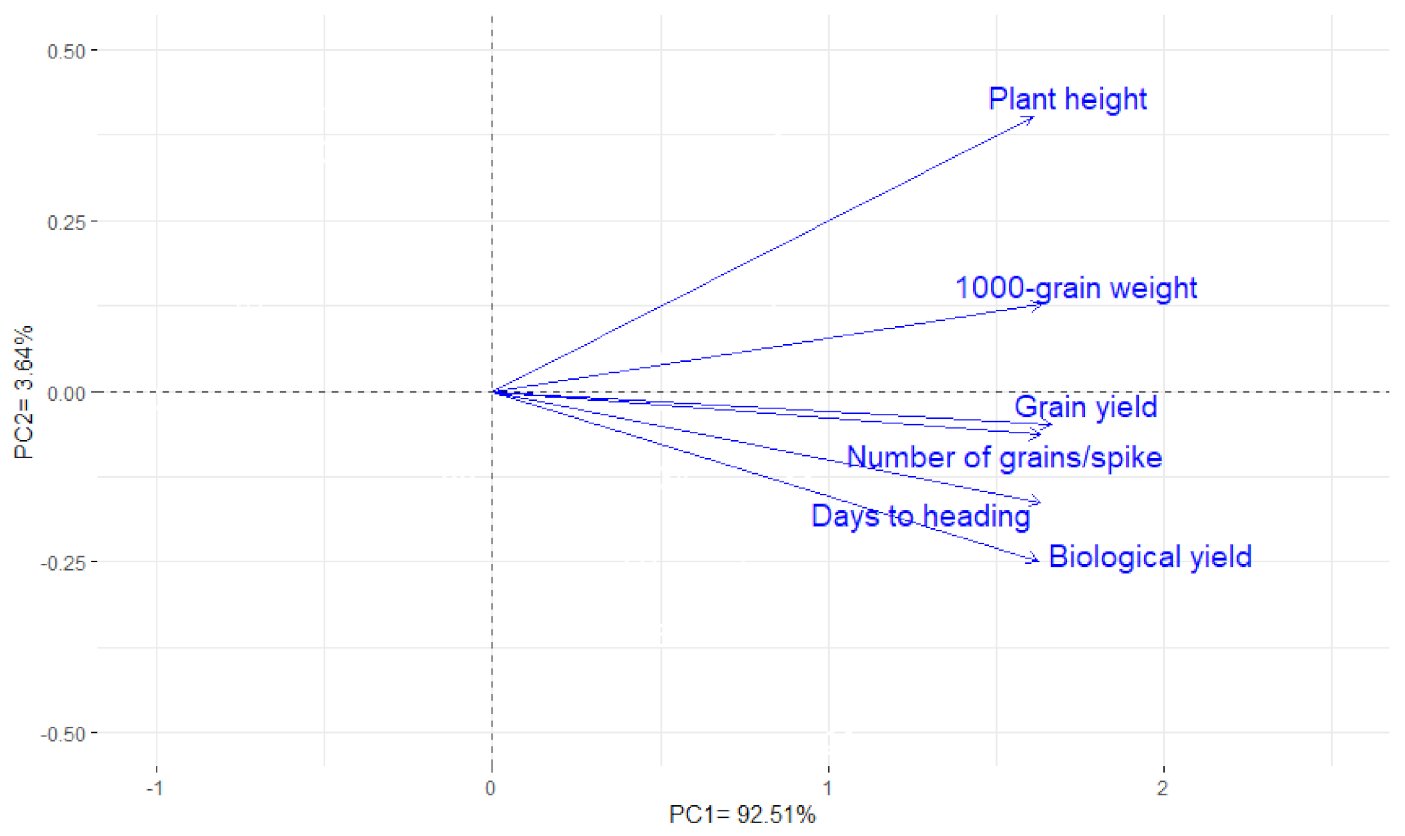
3.6. Interrelationship among Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Joye, I.J. Dietary fibre from whole grains and their benefits on metabolic health. Nutrients 2020, 12, 3045. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Statistical Database. Available online: http://www.fao.org/faostat/en/#data (accessed on 21 December 2020).
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khadka, K.; Raizada, M.N.; Navabi, A. Recent progress in germplasm evaluation and gene mapping to enable breeding of drought-tolerant wheat. Front. Plant Sci. 2020, 11, 1149. [Google Scholar] [CrossRef]
- Mansour, E.; Merwad, A.; Yasin, M.; Abdul-Hamid, M.; El-Sobky, E.; Oraby, H. Nitrogen use efficiency in spring wheat: Genotypic variation and grain yield response under sandy soil conditions. J. Agric. Sci. 2017, 155, 1407–1423. [Google Scholar] [CrossRef]
- Dadshani, S.; Sharma, R.C.; Baum, M.; Ogbonnaya, F.C.; Léon, J.; Ballvora, A. Multi-dimensional evaluation of response to salt stress in wheat. PLoS ONE 2019, 14, e0222659. [Google Scholar] [CrossRef] [PubMed]
- Mansour, E.; Moustafa, E.S.; Desoky, E.-S.M.; Ali, M.; Yasin, M.A.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S. Multidimensional evaluation for detecting salt tolerance of bread wheat genotypes under actual saline field growing conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef]
- Luo, J.-Y.; Zhang, S.; Peng, J.; Zhu, X.-Z.; Lv, L.-M.; Wang, C.-Y.; Li, C.-H.; Zhou, Z.-G.; Cui, J.-J. Effects of soil salinity on the expression of Bt toxin (Cry1Ac) and the control efficiency of Helicoverpa armigera in field-grown transgenic Bt cotton. PLoS ONE 2017, 12, e0170379. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Hassan, W.M.; Al-Suhaibani, N.A.; Refay, Y.; Abdella, K.A. Comparative performance of multivariable agro-physiological parameters for detecting salt tolerance of wheat cultivars under simulated saline field growing conditions. Front. Plant Sci. 2017, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Al-Ashkar, I.; Alderfasi, A.; El-Hendawy, S.; Al-Suhaibani, N.; El-Kafafi, S.; Seleiman, M.F. Detecting salt tolerance in doubled haploid wheat lines. Agronomy 2019, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Oyiga, B.C.; Sharma, R.; Shen, J.; Baum, M.; Ogbonnaya, F.; Léon, J.; Ballvora, A. Identification and characterization of salt tolerance of wheat germplasm using a multivariable screening approach. J. Agron. Crop Sci. 2016, 202, 472–485. [Google Scholar] [CrossRef]
- St. Martin, S.; McBlain, B. Procedure to estimate genetic gain by stages in multistage testing programs. Crop Sci. 1991, 31, 1367–1369. [Google Scholar] [CrossRef]
- Gavuzzi, P.; Rizza, F.; Palumbo, M.; Campanile, R.; Ricciardi, G.; Borghi, B. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci. 1997, 77, 523–531. [Google Scholar] [CrossRef]
- Burton, G.W.; Devane, D.E. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material 1. Agron. J. 1953, 45, 478–481. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Gharib, M.; Qabil, N.; Salem, A.; Ali, M.; Awaad, H.; Mansour, E. Characterization of wheat landraces and commercial cultivars based on morpho-phenological and agronomic traits. Cereal Res. Commun. 2020, 1–11. [Google Scholar] [CrossRef]
- Akbarpour, O.A.; Dehghani, H.; Rousta, M.J. Evaluation of salt stress of Iranian wheat germplasm under field conditions. Crop Pasture Sci. 2015, 66, 770–781. [Google Scholar] [CrossRef]
- Genc, Y.; Taylor, J.; Lyons, G.H.; Li, Y.; Cheong, J.; Appelbee, M.; Oldach, K.; Sutton, T. Bread wheat with high salinity and sodicity tolerance. Front. Plant Sci. 2019, 10, 1280. [Google Scholar] [CrossRef] [Green Version]
- Abaza, G.M.S.M.; Awaad, H.A.; Attia, Z.M.; Abdel-lateif, K.S.; Gomaa, M.A.; Abaza, S.M.S.M.; Mansour, E. Inducing potential mutants in bread wheat using different doses of certain physical and chemical mutagens. Plant Breed. Biotech. 2020, 8, 252–264. [Google Scholar] [CrossRef]
- Mansour, E.; Desoky, E.M.; Ali, M.M.A.; Abdul-Hamid, M.I.; Ullah, H.; Attia, A.; Datta, A. Identifying drought-tolerant genotypes of faba bean and their agro-physiological responses to different water regimes in an arid Mediterranean environment. Agric. Water Manag. 2021, 247, 106754. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Shamaya, N.; Baho, M.; Edwards, J.; Ramsey, C.; Nevo, E.; Langridge, P.; Tester, M. Salinity tolerance and Na+ exclusion in wheat: Variability, genetics, mapping populations and QTL analysis. Czech J. Genet. Plant Breed. 2011, 47, S85–S93. [Google Scholar] [CrossRef] [Green Version]
- Gadimaliyeva, G.; Akparov, Z.; Aminov, N.; Aliyeva, A.; Ojaghi, J.; Salayeva, S.; Serpoush, M.; Mammadov, A.; Morgounov, A. Assessment of synthetic wheat lines for soil salinity tolerance. Zemdirbyste 2020, 107, 55–62. [Google Scholar] [CrossRef]
- Lethin, J.; Shakil, S.S.; Hassan, S.; Sirijovski, N.; Töpel, M.; Olsson, O.; Aronsson, H. Development and characterization of an EMS-mutagenized wheat population and identification of salt-tolerant wheat lines. BMC Plant Biol. 2020, 20, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gracia, M.; Mansour, E.; Casas, A.; Lasa, J.; Medina, B.; Cano, J.L.M.; Moralejo, M.; López, A.; Fuster, P.L.; Escribano, J. Progress in the Spanish National Barley Breeding Program. Span. J. Agric. Res. 2012, 741–751. [Google Scholar] [CrossRef] [Green Version]
- Venuprasad, R.; Lafitte, H.R.; Atlin, G.N. Response to direct selection for grain yield under drought stress in rice. Crop Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- Bhutta, W.M.; Hanif, M. Genetic variability of salinity tolerance in spring wheat (Triticum aestivum L.). Acta Agric. Scand. B Soil Plant Sci. 2010, 60, 256–261. [Google Scholar]
- Green, A.J.; Berger, G.; Griffey, C.; Pitman, R.; Thomason, W.; Balota, M.; Ahmed, A. Genetic yield improvement in soft red winter wheat in the Eastern United States from 1919 to 2009. Crop Sci. 2012, 52, 2097–2108. [Google Scholar] [CrossRef]
- Okechukwu, E.C.; Agbo, C.U.; Uguru, M.I.; Ogbonnaya, F.C. Germplasm evaluation of heat tolerance in bread wheat in Tel Hadya, Syria. Chil. J. Agric. Res. 2016, 76, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Lozada, D.N.; Ward, B.P.; Carter, A.H. Gains through selection for grain yield in a winter wheat breeding program. PLoS ONE 2020, 15, e0221603. [Google Scholar] [CrossRef]
- Padi, F. Response to selection for grain yield and correlated response for grain size and earliness in cowpea based on early generation testing. Ann. Appl. Biol. 2008, 152, 361–368. [Google Scholar] [CrossRef]
- Mansour, E.; Moustafa, E.S.; El-Naggar, N.Z.; Abdelsalam, A.; Igartua, E. Grain yield stability of high-yielding barley genotypes under Egyptian conditions for enhancing resilience to climate change. Crop Pasture Sci. 2018, 69, 681–690. [Google Scholar] [CrossRef]
- El-Hendawy, S.; Ruan, Y.; Hu, Y.; Schmidhalter, U. A comparison of screening criteria for salt tolerance in wheat under field and controlled environmental conditions. Agron. Crop Sci. 2009, 195, 356–367. [Google Scholar] [CrossRef]
- Saade, S.; Maurer, A.; Shahid, M.; Oakey, H.; Schmöckel, S.M.; Negrão, S.; Pillen, K.; Tester, M. Yield-related salinity tolerance traits identified in a nested association mapping (NAM) population of wild barley. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Generation | Trait | Genotypes | Error | Total |
|---|---|---|---|---|
| F6 | Df † | 98 | 196 | 296 |
| Days to heading | 69.24 ** | 2.86 | 24.83 | |
| Plant height | 145.44 ** | 7.58 | 53.23 | |
| Number of grains/spike | 57.64 ** | 5.55 | 22.76 | |
| 1000-grain weight | 31.59 ** | 3.13 | 12.53 | |
| Grain yield | 2,076,531 ** | 344,898 | 935,619 | |
| Biological yield | 3,948,980 ** | 560,894 | 1,679,154 | |
| F7 | Df † | 41 | 82 | 125 |
| Days to heading | 46.22 ** | 1.61 | 16.23 | |
| Plant height | 86.66 ** | 4.92 | 31.74 | |
| Number of grains/spike | 15.15 ** | 1.30 | 5.82 | |
| 1000-grain weight | 9.78 ** | 0.85 | 3.78 | |
| Grain yield | 90,488 ** | 13,672 | 38,665 | |
| Biological yield | 1,234,146 ** | 167,345 | 514,633 | |
| F8 | Df † | 38 | 76 | 116 |
| Days to heading | 40.77 ** | 1.86 | 14.57 | |
| Plant height | 64.82 ** | 3.22 | 23.47 | |
| Number of grains/spike | 9.17 ** | 1.09 | 3.75 | |
| 1000-grain weight | 5.75 ** | 0.85 | 2.46 | |
| Grain yield | 39,737 ** | 6120 | 17,095 | |
| Biological yield | 694,176 ** | 89,947 | 286,397 |
| Generation | Trait | Advanced Lines | Check Cultivars | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | SD | CV (%) | Min | Max | Mean | SD | CV (%) | ||
| F6 | Days to heading | 66.67 | 90.00 | 74.09 | 4.78 | 6.45 | 66.33 | 75.67 | 70.20 | 3.44 | 4.90 |
| Plant height (cm) | 57.10 | 90.90 | 76.34 | 7.03 | 9.21 | 81.67 | 89.33 | 85.87 | 2.97 | 3.46 | |
| Number of grains/spike | 29.97 | 47.29 | 41.49 | 4.61 | 11.11 | 36.94 | 46.06 | 39.98 | 3.58 | 8.95 | |
| 1000-grain weight (g) | 26.30 | 38.70 | 34.91 | 3.27 | 9.36 | 36.79 | 48.93 | 42.64 | 4.47 | 10.48 | |
| Grain yield (kg/ha) | 1385 | 4492 | 3241 | 1015 | 31.30 | 3337 | 4314 | 3906 | 508.0 | 13.01 | |
| Biological yield (kg/ha) | 4400 | 12,736 | 8932 | 1364 | 15.27 | 8872 | 12,368 | 10,843 | 1258 | 11.60 | |
| F7 | Days to heading | 67.33 | 83.00 | 73.03 | 3.94 | 5.39 | 65.67 | 73.67 | 69.40 | 2.90 | 4.18 |
| Plant height (cm) | 65.33 | 90.83 | 79.56 | 5.49 | 6.90 | 75.67 | 87.33 | 81.93 | 4.65 | 5.68 | |
| Number of grains/spike | 38.61 | 46.36 | 42.78 | 2.19 | 5.12 | 37.13 | 45.01 | 41.27 | 3.84 | 9.30 | |
| 1000-grain weight (g) | 29.96 | 39.27 | 36.02 | 1.86 | 5.16 | 38.27 | 45.47 | 41.77 | 3.41 | 8.16 | |
| Grain yield (kg/ha) | 3518 | 4492 | 4254 | 200.9 | 4.72 | 3409 | 4335 | 3951 | 415.0 | 10.50 | |
| Biological yield (kg/ha) | 8950 | 12,916 | 11,224 | 753 | 6.71 | 9693 | 12,453 | 11,204 | 1068 | 9.53 | |
| F8 | Days to heading | 67.33 | 84.00 | 73.63 | 3.72 | 5.05 | 66.3 | 74.97 | 70.04 | 3.21 | 4.58 |
| Plant height (cm) | 74.67 | 91.00 | 80.73 | 4.63 | 5.74 | 82.00 | 90.17 | 87.73 | 3.38 | 3.85 | |
| Number of grains/spike | 39.79 | 48.43 | 43.50 | 2.05 | 4.72 | 36.71 | 45.83 | 42.09 | 3.77 | 8.96 | |
| 1000-grain weight (g) | 32.15 | 39.81 | 36.80 | 1.57 | 4.25 | 37.29 | 49.80 | 41.94 | 4.82 | 11.49 | |
| Grain yield (kg/ha) | 3942 | 4560 | 4327 | 133.1 | 3.08 | 3147 | 4275 | 3824 | 594.2 | 15.54 | |
| Biological yield (kg/ha) | 10,709 | 12,928 | 11,717 | 562.5 | 4.80 | 8635 | 12,374 | 10,578 | 1164 | 11.00 | |
| Generation | Parameter † | DH * | PH | NG/S | TGW | GY | BY |
|---|---|---|---|---|---|---|---|
| F6 | σ2g | 20.49 | 43.01 | 16.77 | 8.62 | 688,408 | 1,307,226 |
| σ2p | 22.82 | 49.40 | 21.25 | 10.68 | 1,029,216 | 1,860,496 | |
| GCV | 6.11 | 8.59 | 9.87 | 8.41 | 25.60 | 12.80 | |
| PCV | 6.45 | 9.21 | 11.11 | 9.36 | 31.30 | 15.27 | |
| h2b | 0.90 | 0.87 | 0.79 | 0.81 | 0.67 | 0.70 | |
| Rs | 4.22 | 6.02 | 3.58 | 2.59 | 667.7 | 943.0 | |
| F7 | σ2g | 14.31 | 26.83 | 3.89 | 2.89 | 27,802 | 403,559 |
| σ2p | 15.50 | 30.11 | 4.79 | 3.45 | 40,356 | 567,270 | |
| GCV | 5.18 | 6.51 | 4.61 | 4.72 | 3.92 | 5.66 | |
| PCV | 5.39 | 6.90 | 5.12 | 5.16 | 4.72 | 6.71 | |
| h2b | 0.92 | 0.89 | 0.81 | 0.84 | 0.69 | 0.71 | |
| Rs | 0.71 | 0.95 | 0.35 | 0.30 | 26.99 | 104.5 | |
| F8 | σ2g | 12.65 | 19.41 | 3.45 | 2.06 | 12,254 | 228,527 |
| σ2p | 13.81 | 21.46 | 4.21 | 2.45 | 17,728 | 316,424 | |
| GCV | 4.83 | 5.46 | 4.27 | 3.90 | 2.56 | 4.08 | |
| PCV | 5.05 | 5.74 | 4.72 | 4.25 | 3.08 | 4.80 | |
| h2b | 0.92 | 0.90 | 0.82 | 0.84 | 0.69 | 0.72 | |
| Rs | 0.66 | 0.82 | 0.33 | 0.26 | 17.95 | 79.22 |
| Trait | Generations | First Generation | Evaluated in Second Generation | S | GG | |
|---|---|---|---|---|---|---|
| All Lines | Selected Lines | |||||
| Days to heading | F6–F7 | 103.5 | 104.3 | 104.2 | 0.73 | 0.69 |
| F7–F8 | 104.2 | 104.5 | 104.4 | 0.27 | 0.14 | |
| Plant height | F6–F7 | 88.91 | 93.88 | 92.96 | 4.97 | 4.05 |
| F7–F8 | 92.96 | 94.24 | 93.80 | 1.28 | 0.84 | |
| Number of grains/spike | F6–F7 | 102.0 | 105.7 | 104.8 | 3.69 | 2.87 |
| F7–F8 | 104.8 | 106.3 | 105.2 | 1.46 | 0.33 | |
| 1000-grain weight | F6–F7 | 101.2 | 108.2 | 107.5 | 6.92 | 6.28 |
| F7–F8 | 107.5 | 108.9 | 107.9 | 1.38 | 0.40 | |
| Grain yield | F6–F7 | 83.02 | 111.2 | 108.0 | 28.17 | 24.96 |
| F7–F8 | 108.0 | 109.1 | 108.7 | 1.13 | 0.71 | |
| Biological yield | F6–F7 | 80.59 | 103.6 | 100.2 | 23.05 | 19.60 |
| F7–F8 | 100.2 | 102.4 | 101.9 | 2.23 | 1.70 | |
| Genotype | Parents | DH * | PH | NG/S | TGW | GY | BY | GY Ranking |
|---|---|---|---|---|---|---|---|---|
| L2 | P1 × P2 | 72.22 | 86.06 | 42.71 | 36.45 | 4314 | 11,201 | 20 |
| L3 | P1 × P2 | 74.67 | 82.68 | 41.29 | 37.53 | 4419 | 11,799 | 4 |
| L13 | P1 × P2 | 76.67 | 88.76 | 41.63 | 36.38 | 4323 | 11,674 | 16 |
| L15 | P5 × P2 | 77.89 | 78.88 | 41.76 | 37.63 | 4367 | 11,811 | 9 |
| L16 | P5 × P2 | 79.33 | 80.41 | 41.28 | 34.66 | 4408 | 11,820 | 6 |
| L21 | P5 × P2 | 78.11 | 77.22 | 45.09 | 37.37 | 4376 | 11,944 | 8 |
| L23 | P5 × P2 | 77.89 | 83.87 | 43.47 | 36.95 | 4363 | 11,320 | 10 |
| L24 | P5 × P2 | 68.67 | 84.06 | 41.76 | 38.05 | 4238 | 10,949 | 29 |
| L27 | P5 × P2 | 77.22 | 82.97 | 43.72 | 35.12 | 4311 | 11,209 | 22 |
| L30 | P9 × P10 | 72.33 | 79.89 | 44.09 | 37.95 | 4355 | 11,479 | 12 |
| L33 | P1 × P3 | 68.89 | 77.23 | 43.60 | 37.09 | 4249 | 11,182 | 26 |
| L34 | P1 × P3 | 75.67 | 72.31 | 44.15 | 37.69 | 4393 | 11,809 | 7 |
| L37 | P1 × P3 | 77.56 | 79.26 | 43.52 | 37.33 | 4340 | 11,967 | 13 |
| L39 | P1 × P3 | 69.56 | 78.11 | 43.88 | 37.04 | 4248 | 11,129 | 27 |
| L44 | P1 × P3 | 72.56 | 78.72 | 43.79 | 38.46 | 4440 | 11,625 | 2 |
| L45 | P1 × P3 | 71.85 | 76.41 | 39.55 | 33.82 | 3987 | 9950 | 37 |
| L49 | P8 × P10 | 68.78 | 74.30 | 43.79 | 37.02 | 4318 | 11,394 | 17 |
| L50 | P8 × P10 | 74.44 | 79.23 | 43.60 | 37.47 | 4302 | 11,209 | 23 |
| L51 | P8 × P10 | 72.11 | 79.44 | 42.39 | 37.16 | 4330 | 11,606 | 15 |
| L53 | P4 × P6 | 69.11 | 70.98 | 39.07 | 34.67 | 3962 | 9709 | 39 |
| L54 | P4 × P6 | 68.89 | 75.97 | 40.12 | 37.58 | 4224 | 10,042 | 30 |
| L57 | P4 × P6 | 72.00 | 71.49 | 45.19 | 36.86 | 4041 | 10,093 | 33 |
| L59 | P4 × P6 | 68.33 | 76.31 | 40.89 | 37.54 | 3989 | 9814 | 36 |
| L61 | P4 × P6 | 69.22 | 74.37 | 40.90 | 37.46 | 4011 | 10,222 | 35 |
| L62 | P4 × P6 | 75.44 | 79.39 | 44.43 | 37.11 | 4340 | 10,978 | 13 |
| L64 | P4 × P6 | 83.89 | 78.57 | 43.81 | 38.21 | 4422 | 11,965 | 3 |
| L65 | P4 × P6 | 78.00 | 76.42 | 42.69 | 37.23 | 4411 | 12,860 | 5 |
| L70 | P1 × P4 | 73.56 | 77.23 | 44.60 | 36.86 | 4316 | 11,586 | 18 |
| L72 | P1 × P4 | 67.22 | 71.76 | 40.68 | 36.00 | 3981 | 10,260 | 38 |
| L77 | P1 × P4 | 73.44 | 78.48 | 45.89 | 38.71 | 4467 | 11,464 | 1 |
| L78 | P1 × P4 | 67.67 | 75.92 | 40.50 | 37.87 | 4038 | 10,175 | 34 |
| L79 | P1 × P4 | 73.67 | 76.99 | 47.19 | 36.32 | 4276 | 11,333 | 25 |
| L80 | P1 × P4 | 69.89 | 78.83 | 45.78 | 37.05 | 4240 | 11,375 | 28 |
| L83 | P10 × P7 | 76.44 | 79.00 | 44.70 | 36.68 | 4314 | 11,015 | 20 |
| Sakha-94 | Check | 73.57 | 82.89 | 43.59 | 38.63 | 4315 | 12,852 | 19 |
| Misr-1 | Check | 74.77 | 81.44 | 41.30 | 38.37 | 4079 | 10,067 | 32 |
| Gemmiza-7 | Check | 72.10 | 86.44 | 40.71 | 37.37 | 4277 | 11,210 | 24 |
| Gemmiza-9 | Check | 73.57 | 88.50 | 44.76 | 37.98 | 4360 | 11,848 | 11 |
| Gemmiza-12 | Check | 70.40 | 86.61 | 45.54 | 36.68 | 4097 | 11,398 | 31 |
| LSD0.05% | 1.48 | 2.07 | 1.65 | 1.78 | 102 | 254.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moustafa, E.S.A.; Ali, M.M.A.; Kamara, M.M.; Awad, M.F.; Hassanin, A.A.; Mansour, E. Field Screening of Wheat Advanced Lines for Salinity Tolerance. Agronomy 2021, 11, 281. https://doi.org/10.3390/agronomy11020281
Moustafa ESA, Ali MMA, Kamara MM, Awad MF, Hassanin AA, Mansour E. Field Screening of Wheat Advanced Lines for Salinity Tolerance. Agronomy. 2021; 11(2):281. https://doi.org/10.3390/agronomy11020281
Chicago/Turabian StyleMoustafa, Ehab S. A., Mohamed M. A. Ali, Mohamed M. Kamara, Mohamed F. Awad, Abdallah A. Hassanin, and Elsayed Mansour. 2021. "Field Screening of Wheat Advanced Lines for Salinity Tolerance" Agronomy 11, no. 2: 281. https://doi.org/10.3390/agronomy11020281