
Weed Diversity, Abundance, and Seedbank in Differently Tilled Faba Bean (Vicia faba L.) Cultivations

 ,
,  ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Treatments and Agronomic Practices
2.3. Methods
2.4. Statistical Analysis
3. Results
3.1. Meteorological Conditions
3.2. Topsoil Residue Coverage
3.3. Weed Species Composition and Density
3.4. Weed Biomass
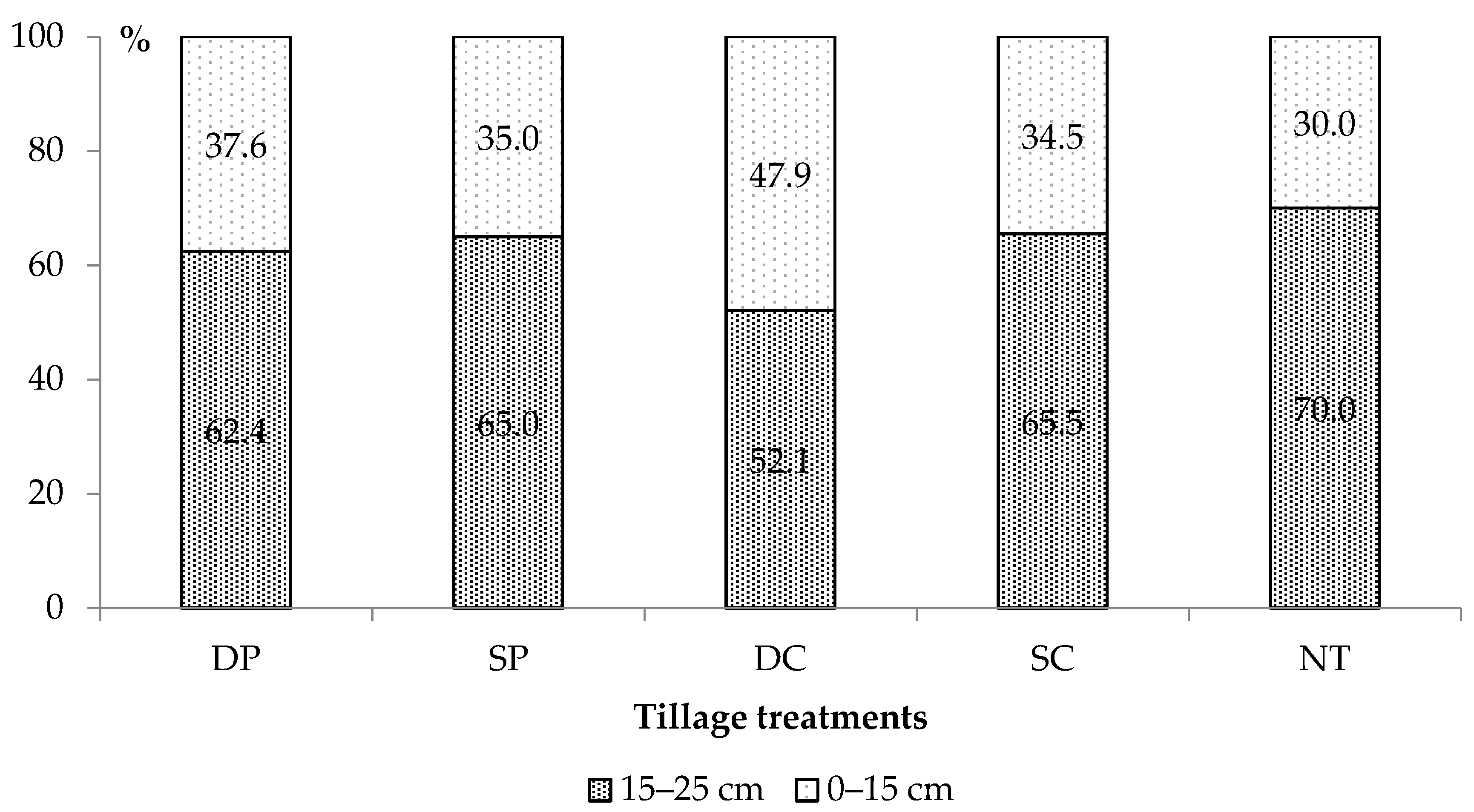
3.5. Weed Seedbank
4. Discussion
4.1. Topsoil Residue Coverage
4.2. Weeds Species Composition and Density
4.3. Weed Biomass
4.4. Weed Seedbank
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sosnoskie, L.M.; Herms, C.P.; Cardina, J.; Webster, T.M. Seedbank and emerged weed communities following adoption of glyphosate-resistant crops in a longterm tillage and rotation study. Weed Sci. 2009, 57, 261–270. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Melander, B.; Munier-Jolain, N.; Charles, R.; Wirth, J.; Schwarz, J.; Van der Weide, R.; Bonin, L.; Jensen, P.K.; Kudsk, P. European perspectives on the adoption of non-chemical weed management in reduced tillage systems for arable crops. Weed Technol. 2013, 27, 231–240. [Google Scholar] [CrossRef]
- Morris, N.L.; Miller, P.C.H.; Orson, J.H.; Froud-Williams, R.J. The adoption of non-inversion tillage systems in the United Kingdom and the agronomic impact on soil, crops and the environment. A review. Soil Till. Res. 2010, 108, 1–15. [Google Scholar] [CrossRef]
- Šarauskis, E.; Buragienė, S.; Masilionytė, L.; Romaneckas, K.; Avižienytė, D.; Sakalauskas, A. Energy balance, costs and CO2 analysis of tillage technologies in maize cultivation. Energy 2014, 69, 227–235. [Google Scholar] [CrossRef]
- Alarcón Víllora, R.; Hernández Plaza, E.; Navarrete, L.; Sánchez, M.J.; Sánchez, A.M. Climate and tillage system drive weed communities’ functional diversity in a Mediterranean cereal-legume rotation. Agric. Ecosyst. Environ. 2019, 283, 1–10. [Google Scholar] [CrossRef]
- Alarcón Víllora, R.; Hernández Plaza, E.; Navarrete, L.; Sánchez, M.J.; Escudero, A.; Hernanz, J.L.; Sánchez-Giron, V.; Sánchez, A.M. Effects of no-tillage and non-inversion tillage on weed community diversity and crop yield over nine years in a Mediterranean cereal-legume cropland. Soil Till. Res. 2018, 179, 54–62. [Google Scholar] [CrossRef]
- Colbach, N.; Busset, H.; Roger-Estrade, J.; Caneill, J. Predictive modelling of weed seed movement in response to superficial tillage tools. Soil Till. Res. 2014, 138, 1–8. [Google Scholar] [CrossRef]
- Batlla, D.; Benech-Arnold, R.L. Weed seed germination and the light environment: Implications for weed management. Weed Biol. Manag. 2014, 14, 77–87. [Google Scholar] [CrossRef]
- Milberg, P.; Andersson, L.; Thompson, K. Large-seeds species are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef]
- Mohler, C.L. A model of the effects of tillage on emergence of weed seedlings. Ecol. Appl. 1993, 3, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Forcella, F.; Benech Arnold, R.; Sánchez, R.; Ghersa, C.M. Modelling seedling emergence. Field Crop. Res. 2000, 67, 123–139. [Google Scholar] [CrossRef]
- Scherner, A.; Melander, B.; Kudsk, P. Vertical distribution and composition of weed seeds within the plough layer after eleven years of contrasting crop rotation and tillage schemes. Soil Till. Res. 2016, 161, 135–142. [Google Scholar] [CrossRef]
- Almoussawi, A.; Lenoir, J.; Spicher, F.; Dupont, F.; Chabrerie, O.; Closset-Kopp, D.; Brasseur, B.; Kobaissi, A.; Dubois, F.; Decocq, G. Direct seeding associated with a mixture of winter cover crops decreases weed abundance while increasing cash-crop yields. Soil Till. Res. 2020, 200, 1–11. [Google Scholar] [CrossRef]
- Barnes, E.R.; Werle, R.; Sandell, L.D.; Lindquist, J.L.; Knezevic, S.Z.; Sikkema, P.H.; Jhala, A.J. Influence of tillage on common ragweed (Ambrosia artemisiifolia) emergence pattern in Nebraska. Weed Technol. 2017, 31, 623–631. [Google Scholar] [CrossRef]
- Ganie, Z.A.; Lindquist, J.L.; Jugulam, M.; Kruger, G.R.; Marx, D.B.; Jhala, A.J. An integrated approach to control glyphosate-resistant Ambrosia trifida with tillage and herbicides in glyphosate-resistant maize. Weed Res. 2017, 57, 112–122. [Google Scholar] [CrossRef]
- Santín-Montanyá, M.I.; Zambrana-Quesada, E.; Tenorio-Pasamón, J.L. Weed abundance and soil seedbank responses to tillage systems in continuous maize crops. Arch. Agron. Soil Sci. 2018, 64, 1705–1713. [Google Scholar] [CrossRef]
- Travlos, I.S.; Cheimona, N.; Roussis, I.; Bilalis, D.J. Weed-species abundance and diversity indices in relation to tillage systems and fertilization. Front. Environ. Sci. 2018, 6, 11. [Google Scholar] [CrossRef]
- Soane, B.D.; Ball, B.C.; Arvidsson, J.; Basch, G.; Moreno, F.; Roger-Estrade, J. No-till in northern, western and south-western Europe. A review of problems and opportunities for crop production and the environment. Soil Till. Res. 2012, 118, 66–87. [Google Scholar] [CrossRef]
- Hernández Plaza, E.H.; Navarrete, L.; González-Andújar, J.L. Intensity of soil disturbance shapes response trait diversity of weed communities: The long-term effects of different tillage systems. Agric. Ecosyst. Environ. 2015, 207, 101–108. [Google Scholar] [CrossRef]
- Armengot, L.; Blanco-Moreno, J.M.; Bàrberi, P.; Bocci, G.; Carlesi, S.; Aendekerk, R.; Berner, A.; Celette, F.; Grosse, M.; Huiting, H.; et al. Tillage as a driver of change in weed communities: A functional perspective. Agric. Ecosyst. Environ. 2016, 222, 276–285. [Google Scholar] [CrossRef]
- Fried, G.; Kazakou, E.; Gaba, S. Trajectories of weed communities explained by traits associated with species’ response to management practices. Agric. Ecosyst. Environ. 2012, 158, 147–155. [Google Scholar] [CrossRef]
- Romaneckienė, R.; Pilipavičius, V.; Romaneckas, K. Weed emergence and death in the crop of spring barley of different competitive ability. Vagos 2008, 15, 17–24, (In Lithuanian with English summary). [Google Scholar]
- Barberi, P.; Cascio Lo, B. Long-term tillage and crop rotation effects and weed seed bank size and composition. Weed Res. 2001, 41, 325–340. [Google Scholar] [CrossRef]
- Cardina, J.; Herms, C.P.; Doohan, D.J. Crop rotation and tillage system effects on weed seedbanks. Weed Sci. 2002, 50, 448–460. [Google Scholar] [CrossRef]
- Trichard, A.; Alignier, A.; Chauvel, B.; Petit, S. Identification of weed community traits response to conservation agriculture. Agric. Ecosyst. Environ. 2013, 179, 179–186. [Google Scholar] [CrossRef]
- Santín-Montanyá, M.I.; Martín-Lammerding, D.; Zambrana, E.; Tenorio, J.L. Management of weed emergence and weed seed bank in response to different tillage, cropping systems and selected soil properties. Soil Till. Res. 2016, 161, 38–46. [Google Scholar] [CrossRef]
- Karkanis, A.; Ntatsi, G.; Lepse, L.; Fernández, J.A.; Vågen, I.M.; Rewald, B.; Alsiņa, I.; Kronberga, A.; Balliu, A.; Olle, M.; et al. Faba bean cultivation—Revealing novel managing practices for a more sustainable and competitiveness European cropping systems. Front. Plant Sci. 2018, 9, 1115. [Google Scholar] [CrossRef]
- Statistic Lithuania 2019. Available online: https://www.vic.lt/ppis/statistine-informacija (accessed on 22 May 2020). (In Lithuanian).
- Lake, L.; Godoy-Kutchartt, D.E.; Calderini, D.F.; Verrell, A.; Sadras, V.O. Yield determination and the critical period of faba bean (Vicia faba L.). Field Crop. Res. 2019, 241, 107575. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources, 3rd ed.; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2014; Available online: http://www.fao.org/3/i3794en/I3794en.pdf (accessed on 3 April 2020).
- Celik, I.; Turgut, M.M.; Acir, N. Crop rotation and tillage effects on selected soil physical properties of a Typic Haploxerert in an irrigated semi-arid Mediterranean region. Int. J. Plant Prod. 2012, 6, 457–480. [Google Scholar]
- Kumar, A.; Chen, Y.; Sadek, A.; Rahman, S. Soil cone index in relation to soil texture, moisture content, and bulk density for no-tillage and conventional tillage. Agric. Eng. Int. CIGR J. 2012, 14, 26–37. [Google Scholar]
- Li, Y.; Li, Z.; Cui, S.; Jagadamma, S.; Zhang, Q. Residue retention and minimum tillage improve physical environment of the soil in croplands: A global meta-analysis. Soil Tillage Res. 2019, 194, 104292. [Google Scholar] [CrossRef]
- Romaneckas, K.; Kimbirauskienė, R.; Adamavičienė, A.; Buragienė, S.; Sinkevičienė, A.; Šarauskis, E.; Jasinskas, A.; Minajeva, A. Impact of sustainable tillage on biophysical properties of Planosol and on faba bean yield. Agric. Food Sci. 2019, 28, 101–111. [Google Scholar] [CrossRef][Green Version]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants. BBCH Monograph; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018. [Google Scholar]
- Stancevicius, A. Weed Accounting and Weed Field Mapping; Mokslas: Vilnius, Lithuania, 1979. (In Lithuanian) [Google Scholar]
- Stancevicius, A. New Rational Method for Determining Weed Seed Content in the Soil. Agronomy, Land Reclamation and Hydro-Engineering; Lithuanian University of Agriculture: Kaunas-Akademija, Lithuania, 1980. (In Lithuanian) [Google Scholar]
- Buchanan, A.L.; Kolb, L.N.; Hook, C.R.R. Can winter cover crops influence weed density and diversity in a reduced tillage vegetable system? Crop Prot. 2016, 90, 9–16. [Google Scholar] [CrossRef]
- Büchi, L.; Wendling, M.; Amossé, C.; Jeangros, B.; Charles, R. Cover crops to secure weed control strategies in a maize crop with reduced tillage. Field Crop. Res. 2020, 247, 107583. [Google Scholar] [CrossRef]
- Romaneckas, K.; Adamavičienė, A.; Pilipavičius, V.; Šarauskis, E.; Avižienytė, D.; Buraguenė, S. Interaction of maize and living mulch. Crop weediness and productivity. Zemdirb. Agric. 2012, 99, 23–30. [Google Scholar]
- Karkanis, A.; Ntatsi, G.; Kontopoulou, C.K.; Pristeri, A.; Bilalis, D.; Savvas, D. Field pea in European cropping systems: Adaptability, biological nitrogen fixation and cultivation practices. Not. Bot. Horti. Agrobot. Cluj Napoca 2016, 442, 325–336. [Google Scholar] [CrossRef]
- Karkanis, A.; Travlos, I.S.; Bilalis, D.; Tabaxi, E.I. Integrated weed management in winter cereals in Southern Europe. In Weed and Pest Control: Molecular Biology, Practices and Environmental Impact; Travlos, I.S., Bilalis, D.J., Chachalis, D., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2016; pp. 1–15. [Google Scholar]
- Shahzad, M.; Farooq, M.; Jabran, K.; Hussain, M. Impact of different crop rotations and tillage systems on weed infestation and productivity of bread wheat. Crop Prot. 2016, 89, 161–169. [Google Scholar] [CrossRef]
- Légère, A.; Vanasse, A.; Craig Stevenson, F. Low-input management and mature conservation tillage: Agronomic potential in a cool humid climate. Agron. J. 2013, 105, 745–754. [Google Scholar] [CrossRef]
- Hossain, K.; Timsina, J.; Johnson, D.E.; Gathala, M.K.; Krupnik, T.J. Multi-year weed community dynamics and rice yields as influenced by tillage, crop establishment, and weed control: Implications for rice-maize rotations in the eastern Gangetic plains. Crop Prot. 2020, 138, 105334. [Google Scholar] [CrossRef]
- Peigné, J.; Ball, B.C.; Roger-Estrade, J.; David, C. Is conservation tillage suitable for organic farming? Soil Use Manag. 2007, 23, 129–144. [Google Scholar] [CrossRef]
- Carter, M.R.; Ivany, J.A. Weed seed bank composition under three long-term tillage regimes on a fine sandy loam in Atlantic Canada. Soil Till. Res. 2006, 90, 29–38. [Google Scholar] [CrossRef]
- Hernández Plaza, E.; Kozak, M.; Navarrete, L.; González-Andújar, J.L. Tillage system did not affect weed diversity in a 23-year experiment in Mediterranean dryland. Agric. Ecosyst. Environ. 2011, 140, 102–105. [Google Scholar] [CrossRef]
- Thomas, A.G.; Derksen, D.A.; Blachshaw, R.E.; Van Acker, R.C.; Lègére, A.; Watson, P.R.; Turnbull, G.C. A multistudy approach to understanding weed population shifts in medium- to long-term tillage systems. Weed Sci. 2004, 52, 874–880. [Google Scholar] [CrossRef]
- Bilalis, D.; Efthimiadis, P.; Sidiras, N. Effect of three tillage systems on weed flora in a 3-year rotation with four crops. J. Agron. Crop Sci. 2001, 186, 135–141. [Google Scholar] [CrossRef]
- Stancevičius, A.; Špokienė, N.; Jodaugienė, D.; Trečiokas, K.; Raudonius. Impact of reduced soil tillage on crop weedinesss. Proc. Lith. Acad. Agric. 2002, 55, 50–58, (In Lithuanian with English summary). [Google Scholar]
- Armengot, L.; Berner, A.; Blanco-Moreno, J.M. Xavier Sans, F. Long-term feasibility of reduced tillage in organic farming. Agron. Sustain. Dev. 2015, 35, 339–346. [Google Scholar] [CrossRef]
- Santín-Montanyá, M.I.; Martín-Lammerding, D.; Walterb, I.; Zambrana, E.; Tenorio, J.L. Effects of tillage, crop systems and fertilization on weed abundance and diversity in 4-year dry land winter wheat. Eur. J. Agron. 2013, 48, 43–49. [Google Scholar] [CrossRef]
- Alba, O.S.; Syrovy, L.D.; Duddu, H.S.N.; Shirtliffe, S.J. Increased seeding rate and multiple methods of mechanical weed control reduce weed biomass in a poorly competitive organic crop. Field Crop. Res. 2020, 245, 1–9. [Google Scholar] [CrossRef]
- Matloob, A.; Khaliq, A.; Tanveer, A.; Hussain, S.; Aslam, F.; Chauhan, B.S. Weed dynamics as influenced by tillage system, sowing time and weed competition duration in dry-seeded rice. Crop Prot. 2015, 71, 25–38. [Google Scholar] [CrossRef]
- Samarajeewa, K.B.D.P.; Horiuchi, T.; Oba, S. Finger millet (Eleucine corocana L. Gaertn.) as a cover crop on weed control, growth and yield of soybean under different tillage systems. Soil Till. Res. 2006, 90, 93–99. [Google Scholar] [CrossRef]
- Susha, V.S.; Das, T.K.; Nath, C.P.; Pandey, R.; Paul, S.; Ghosh, S. Impacts of tillage and herbicide mixture on weed interference, agronomic productivity and profitability of a maize—Wheat system in the North-western Indo-Gangetic Plains. Field Crop. Res. 2018, 219, 180–191. [Google Scholar] [CrossRef]
- Wei, S.H.; Qiang, S.; Ma, B.; Wei, J.G. Soil weed seedbank and integrated weed management. Soils 2005, 37, 121–128, (In Chinese with English abstract). [Google Scholar]
- Buhler, D.D.; Stoltenberg, D.E.; Becker, R.L.; Gunsolus, J.L. Perennial weed populations after 14 years of variable tillage and cropping practices. Weed Sci. 1994, 43, 205–209. [Google Scholar] [CrossRef]
- Gardarin, A.; Durr, C.; Colbach, N. Which model species for weed seedbank and emergence studies? A review. Weed Res. 2009, 49, 117–130. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Peco, B.; Traba, J.; Levassor, C.; Sánchez, A.M.; Azcárate, F.M. Seed size, shape and persistence in dry Mediterranean grass and scrublands. Seed Sci. Res. 2003, 13, 87–93. [Google Scholar] [CrossRef]
- Koning, L.A.; De Mol, F.; Gerowitt, B. Effects of management by glyphosate or tillage on the weed vegetation in a field experiment. Soil Till. Res. 2019, 186, 79–86. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Weed Groups | Tillage Systems | ||||
|---|---|---|---|---|---|
| DP | SP | DC | SC | NT | |
| 2016 | |||||
| BBCH 25–27 | |||||
| Annual | 59.5ab | 74.1a | 71.2a | 72.0a | 50.8b |
| Perennial | 4.5b | 6.6b | 14.1b | 19.5ab | 34.5a ** |
| Total | 64.1a | 80.7a | 85.4a | 91.6a | 85.4a |
| BBCH 75–79 | |||||
| Annual | 35.8a | 36.6a | 24.1a | 38.7a | 25.4a |
| Perennial | 0.8b | 3.3b | 12.5ab | 16.2ab | 30.4a * |
| Total | 36.6a | 39.9a | 36.6a | 54.9a | 55.8a |
| 2017 | |||||
| BBCH 25–27 | |||||
| Annual | 104.6a | 97.5a | 103.7a | 119.1a | 20.9b ** |
| Perennial | 2.9a | 5.4a | 3.3a | 4.5a | 4.1a |
| Total | 107.5a | 102.9a | 107.0a | 123.6a | 25.0b ** |
| BBCH 75–79 | |||||
| Annual | 27.9a | 17.0a | 18.7a | 17.6a | 17.9a |
| Perennial | 2.5a | 4.6a | 2.9a | 1.2a | 5.4a |
| Total | 30.4a | 21.6a | 21.6a | 18.8a | 23.3a |
| 2018 | |||||
| BBCH 25–27 | |||||
| Annual | 103.7b | 130.4b | 132.9b | 117.3b | 226.6a * |
| Perennial | 2.9b | 14.2ab | 9.5 ab | 25.4a * | 19.6 ab |
| Total | 106.6b | 144.6b | 142.4b | 142.7b | 246.2a * |
| BBCH 75–79 | |||||
| Annual | 45.8b | 40.0b | 42.5b | 39.1b | 62.3 *a |
| Perennial | 0.8b | 12.9a * | 10.0ab | 8.4ab | 19.5a * |
| Total | 46.6b | 52.9b | 52.5b | 47.5b | 80.8a * |
| Weed Groups | Tillage Systems | ||||
|---|---|---|---|---|---|
| DP | SP | DC | SC | NT | |
| 2016 | |||||
| Annual | 104.5a | 77.3ab | 71.8ab | 64.2ab | 47.9b * |
| Perennial | 1.0a | 4.3a | 28.1ab | 19.9ab | 93.6b * |
| Total | 105.5a | 81.6a | 100.0a | 84.1a | 141.5a |
| 2017 | |||||
| Annual | 34.4a | 34.0a | 34.7a | 36.7a | 21.2a |
| Perennial | 1.3a | 8.2a | 6.4a | 2.5a | 9.6a |
| Total | 35.7a | 42.2a | 41.1a | 39.2a | 30.8a |
| 2018 | |||||
| Annual | 203.2a | 154.6a | 155.5a | 159.7a | 185.9a |
| Perennial | 0.6b | 39.4ab | 63.7a * | 45.8ab | 18.9ab |
| Total | 203.8a | 194.0a | 219.2a | 205.5a | 204.8a |
| Weed Groups | Tillage Systems | ||||
|---|---|---|---|---|---|
| DP | SP | DC | SC | NT | |
| 2016 | |||||
| 0–15 cm | |||||
| Annual | 64.5b | 65.1b | 77.7ab | 88.4a ** | 63.0b |
| Perennial | 6.3a | 6.3a | 12.0a | 4.2a | 6.8a |
| Total | 70.8b | 71.4b | 89.7a * | 92.6a * | 69.8b |
| 15–25 cm | |||||
| Annual | 133.8a | 85.7b * | 37.6c *** | 28.0c *** | 42.0bc *** |
| Perennial | 9.7b | 31.5b | 44.6b | 115.5a *** | 112.8a *** |
| Total | 143.5a | 117.2b * | 82.2c * | 143.5a | 154.8a |
| 2017 | |||||
| 0–15 cm | |||||
| Annual | 55.1a | 44.6ab | 45.6ab | 64.5a | 30.9b * |
| Perennial | 1.5a | 3.1a | 7.8a | 6.8a | 8.4a |
| Total | 56.6b | 47.7bc | 53.4b | 71.3a * | 39.3c * |
| 15–25 cm | |||||
| Annual | 55.2b | 94.5a * | 47.2b | 48.1b | 29.7b |
| Perennial | 0.8b | 17.5b | 26.5b | 174.1a ** | 114.6ab |
| Total | 56.0d | 112.0bc | 73.7cd | 222.1a * | 144.3b * |
| 2018 | |||||
| 0–15 cm | |||||
| Annual | 77.7a | 92.4a | 112.8a | 107.6a | 91.3a |
| Perennial | 1.5a | 0.0a | 4.2a | 0.0a | 0.0a |
| Total | 79.2c | 92.4bc | 117.0a * | 107.6ab | 91.3bc |
| 15–25 cm | |||||
| Annual | 96.2a | 151.3a | 89.2a | 149.6a | 167.1a |
| Perennial | 47.2a | 12.2a | 37.6a | 0.0a | 1.7a |
| Total | 143.4a | 163.6a | 126.8a | 149.6a | 168.8a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romaneckas, K.; Kimbirauskienė, R.; Sinkevičienė, A.; Jaskulska, I.; Buragienė, S.; Adamavičienė, A.; Šarauskis, E. Weed Diversity, Abundance, and Seedbank in Differently Tilled Faba Bean (Vicia faba L.) Cultivations. Agronomy 2021, 11, 529. https://doi.org/10.3390/agronomy11030529
Romaneckas K, Kimbirauskienė R, Sinkevičienė A, Jaskulska I, Buragienė S, Adamavičienė A, Šarauskis E. Weed Diversity, Abundance, and Seedbank in Differently Tilled Faba Bean (Vicia faba L.) Cultivations. Agronomy. 2021; 11(3):529. https://doi.org/10.3390/agronomy11030529
Chicago/Turabian StyleRomaneckas, Kęstutis, Rasa Kimbirauskienė, Aušra Sinkevičienė, Iwona Jaskulska, Sidona Buragienė, Aida Adamavičienė, and Egidijus Šarauskis. 2021. "Weed Diversity, Abundance, and Seedbank in Differently Tilled Faba Bean (Vicia faba L.) Cultivations" Agronomy 11, no. 3: 529. https://doi.org/10.3390/agronomy11030529
APA StyleRomaneckas, K., Kimbirauskienė, R., Sinkevičienė, A., Jaskulska, I., Buragienė, S., Adamavičienė, A., & Šarauskis, E. (2021). Weed Diversity, Abundance, and Seedbank in Differently Tilled Faba Bean (Vicia faba L.) Cultivations. Agronomy, 11(3), 529. https://doi.org/10.3390/agronomy11030529