1. Introduction
Interest is increasing in including legumes in grass pastures as an alternative to synthetic nitrogen (N) fertilization and to improve nutrient cycling [
1]. Grassland fertilization with organic and synthetic N fertilizers can increase ammonia losses and nitrous oxide emissions, damaging the environment [
2]. In tropical areas, most beef and dairy cattle production is based on pastures, and N management is crucial to maintain the productivity and herbage protein levels required for satisfactory animal performance [
3].
Nitrogen is the key nutrient involved in pasture degradation [
4]. When the level of herbage N declines, herbage accumulation decreases, and consequently, performance per animal and per area declines [
5]. Nitrogen from N fertilization can increase herbage accumulation and leaf proportion, facilitating greater dry matter intake (DMI) by grazing animals. Nitrogen application can also increase the nutritive value of herbage by increasing N compounds and protein content and decreasing concentrations of structural components of herbage, such as pectin, cellulose, and hemicellulose [
6,
7]. However, N fertilization is expensive and can cause negative environmental impacts. As an alternative to N fertilizer for increasing the N supply to the system and, at the same time, reducing the potential for environmental impacts caused by using chemical fertilizers, the introduction of legumes in tropical grass pastures is promising [
8]. Pintoi peanut (
Arachis pintoi Krapov. and W.C. Gregory) is one option for inclusion in palisade grass pastures due to its persistence, nutritive value, and tolerance of grazing and moderate shading [
9,
10]. Nonetheless, more studies are required to confirm its viability in different climatic regions and under different grazing methods, such as intermittent stocking.
Protein and carbohydrates are the most important nutrients to determine animal performance [
11,
12]. Depending on herbage maturity, level of fertilization, and grass management, the concentration and availability of protein and carbohydrates vary. A system of protein and carbohydrate fractionation according to the solubility of each fraction in the rumen was proposed by Sniffen et al. [
13]. Fraction A includes the soluble N compounds that are totally degradable in the rumen, and the fraction B1 + B2 includes true proteins. As forage, legume plants differ from grasses in crude protein (CP) and fiber contents, the fractions of proteins and carbohydrates may vary in mixed pastures compared to monocultures. However, the effect of including forage legumes such as
Arachis pintoi on N compounds, protein, and carbohydrate fractions is not well defined.
Greater availability of technology for planting legumes, the necessity of reducing N environmental impacts, and production costs with fertilization can all stimulate the adoption of mixed pastures. Therefore, our study aimed to evaluate (1) the promising use of pintoi peanut in relation to palisade grass fertilized with N by evaluating the chemical composition and fractionation of N and carbohydrate in herbage, and (2) the herbage mass and accumulation rate, herbage disappearance rate, and stocking rate of pastures. We hypothesized that (1) including pintoi peanut in mixed pastures with palisade grass affects N and carbohydrate profile of herbage and increases its nutritive value compared to N-fertilized palisade grass or non-fertilized pastures of palisade grass, and (2) herbage mass and accumulation rate will not differ between the mixed pintoi peanut–palisade grass pasture and the palisade grass only fertilized with N.
2. Materials and Methods
The experiment was conducted at the Forage and Grasslands Sector of the Sao Paulo State University “Julio de Mesquita Filho” (UNESP), in Jaboticabal, SP, Brazil (21°15′22″ S latitude, 48°18′58″, 77 W longitude and 595 m elevation). According to the Köeppen system, the climate is Aw type (tropical, characterized by dry winters).
In Jaboticabal, SP, the average annual precipitation is 1210 mm, and the temperature oscillates between 17.1 and 30.1 °C, with a mean of 22.7 °C. In the experimental area, the soil is classified as a eutrophic dark red latosol, with a clayey texture, horizon A moderate, and smooth wavy relief [
14].
Experimental pastures were formed by palisade grass (
Brachiaria brizantha R. D. Webster cv. Marandu) in monoculture or mixed with pintoi peanut (
Arachis pintoi cv. Amarillo). This experiment was conducted concomitantly to Berça et al. [
15].
The study was realized during two growing seasons (2016/2017 and 2017/2018), in which chemical composition and herbage mass were evaluated monthly five times per year, and other variables were analyzed between November 2016 and March 2017, and December 2017 and April 2018.
2.1. Experimental Design
There were three treatments, and each was replicated four times in a completely randomized design. The treatments consisted of palisade grass pastures without N fertilizer (control); palisade grass pastures fertilized with urea (150 kg N ha/year; fertilized); and palisade grass pastures mixed with pintoi peanut (mixed).
We used 4.0 ha of total experimental area, where 1.44 was designated as paddocks and 2.56 ha as a reserve area. The 1.44 ha area was divided into 12 pastures (experimental units) that were each subdivided into 3 paddocks to facilitate intermittent stocking, with an approximate area per paddock of 400 m2. When animals were not grazing the experimental plots, we used the reserve pasture of palisade grass.
2.2. Management of Animals and Pastures
We used 21 non-lactating crossbred dairy heifers, 24 months old and 300 ± 5.9 kg average body weight (BW) in the first experimental year. The same genetic background of 26 heifers was used in the second year but 270 ± 3.9 kg BW.
Pastures were managed under intermittent and variable stocking rates, following the mob stocking technique [
16]. When pasture achieved 95% light interception (LI), the grazing was started, using the LI-COR
® canopy analyzer to measure it (LI-COR
®, Lincoln, NE, USA). In addition, animals were removed from the paddock when the average height of the herbage was 15 cm, the stocking density was adjusted based on the average BW of heifers, and the occupation period was only one day.
Maintenance fertilization of all treatments with P and K was determined according to the results of soil chemical analyses, carried out in July 2016 and July 2017, following the recommendation of Technical Bulletin 100 [
17], in order to maintain these nutrients at similar levels among all paddocks. In the N-fertilized pastures, 150 kg N ha/year were applied using urea (45% N) and split into 3 equal applications (10 November 2016, 10 January 2017, and 10 March 2017 in the first year; and 14 November 2017, 15 January 2018, and 25 March 2018 in the second year).
2.3. Chemical Composition of Pastures
In order to evaluate the chemical composition of herbage in each paddock and in all grazing cycles, samples of herbage were hand plucked at 40 locations within each experimental unit of 400 m
2, after observing animal grazing behavior [
18], which were analyzed after drying and grinding processes. The constituents of the herbage fiber fractions were determined by the ANKOM methodology (ANKOM Technology, Macedon, NY, USA), as neutral detergent fiber (NDF) [
19], acid detergent fiber (ADF) [
20], and lignin by the acid hydrolysis method [
21].
The other chemical components of herbage were analyzed following the methodologies of INCT-CA [
22], as total N (crude protein; CP) by the Kjeldahl method, and ash and ether extract (EE) by the goldfish method.
The organic matter (OM), hemicellulose, and lignin contents were calculated according to the following Equations (1)–(3).
The methodology of Sniffen et al. was used to determine fractions of herbage carbohydrate [
13], including total carbohydrate (TC), non-fibrous carbohydrate (NFC), and total digestible nutrients (TDNs).
Feces samples were collected directly in the paddocks, at the time of animal defecation, with the aid of gloves and plastic bags. Then, samples were identified, weighed, and dried in an oven with forced air circulation at 55 °C for 72 h and ground in a Willey mill (1 mm).
The indigestible neutral detergent fiber (iNDF) was determined by in situ incubation. Feces and hand-plucked herbage samples were conditioned in the rumen of fistulated animals for 240 h, inside ANKOM F-57 filter bags [
23]. Afterwards, bags were removed from the rumen, totally washed, and then dried in an oven at 55 °C for 72 h. In sequence, neutral detergent extraction of bags in an ANKOM fiber analyzer was performed, and the whole drying methodology was repeated for iNDF quantification [
12].
Fractionation of N was determined following the Cornell system [
13]. Fraction A was obtained by analyzing the hand-plucked samples in trichloroacetic acid (TCA) for the extraction of soluble N [
24], and later calculated by the difference between the total N content and the non-protein nitrogen (NPN) [
13]. The B3 fraction was calculated by the difference between the neutral detergent-insoluble nitrogen (NDIN) and the acid detergent-insoluble nitrogen (ADIN) contents. Fraction C, in turn, was considered as ADIN, described by Sniffen et al. [
13] and further discussed by Tedeschi and Fox [
11]. Finally, fraction B1 + B2 (true protein) was calculated as the difference between the total N and the other fractions (B1 + B2 = total N − (A + B3 + C)).
2.4. Herbage Accumulation Rate
To quantify the herbage accumulation rate (HAR), before and after grazing, 16 points of the height of palisade grass were randomly measured with a graduated ruler [
25]. From the average height of the paddock, two representative grass samples were collected by cutting 5 cm from the soil all herbage present in a 0.25 m
2 metal frame.
The agronomic method of the difference was used to determine HAR, from the pre- and post-grazing masses, in an equation described by Campbell [
26] (Equation (4)):
where HARi corresponds to the accumulation rate of daily dry matter in period i; Gj is the pre-grazing dry mass of the grazing cycle j; F(j-1) is the post-grazing dry mass of the grazing cycle j; and
n is the rest period.
2.5. Herbage Disappearance Rate
The same methodology of cutting the pre- and post-grazing mass, as described in the previous section [
25], was used to determine the herbage disappearance rate (HDR), which was calculated by the difference between pre- and post-grazing mass of the same cycle.
Herbage losses were determined using 0.25 m
2 metal frames. The area was cleaned before grazing and after the exit of the animals, removing all dead plants present on the soil surface, as well as the senescent herbage tissue portions in the structure of the plants (i.e., the litter) [
27].
By the difference between pre-grazing and post-grazing mass of the same grazing cycle and discounting the herbage losses, the intake by animals was estimated, following Equation (5):
where FI corresponds to herbage intake (kg DM/day); Q1 to the amount of herbage before grazing; Q2 to the amount of herbage after removal of the animals; and L to the herbage losses.
2.6. Herbage Mass, Morphological and Botanical Composition
From the pre- and post-grazing cuts, each sample was separated into two subsamples, one for determination of DM content and the other for morphological fractionation in leaf, stem + sheath, and dead material. In the material related to the mixed treatment, the samples were also separated into botanical components (grass and legumes). In order to estimate the total herbage mass (HM), samples were dried for 72 h in a 55 °C oven to obtain the total DM and percentage of morphological components [
28]. The herbage allowance (kg DM/100 kg BW) was determined by dividing HM by animal liveweight, and, also, in order to calculate leaf allowance, we multiplied herbage allowance by the percentage of leaves.
2.7. Statistical Analyses
To check for the homoscedasticity and normality of the residues, data were analyzed using the Nortest package of R, version 3.5.2, through Box–Cox and Cramér–von Mises tests, respectively [
29]. A completely randomized design in a factorial arrangement was considered to conduct the analysis of variance, with time-repeated measures using the lme package of R. The best covariance structure was selected using the corrected Akaike criterion. The treatments, cycles, and interactions of treatments x cycles were considered fixed effects and replications (pastures) were random effects. When the analysis of variance was significant for treatments, the means of the three treatments were compared by the Tukey’s honestly significant difference (Tukey HSD) test at 5% probability, and we used orthogonal polynomial contrasts to identify the effect of the cycles when differences occurred between the grazing cycles (
p < 0.05).
3. Results
3.1. Fractionation of Herbage Carbohydrate and Protein
Total carbohydrates were affected by treatments but not by the grazing cycles. The TC content was lower (
p < 0.05) in fertilized pastures in relation to the control, and mixed pastures were similar to both (
Table 1). There was no effect of legume inclusion or N fertilization on NFC and TDN content of herbage (
p > 0.05) (
Table 1). A quadratic effect of grazing cycle on TDN content was found in the fertilized treatment. Additionally, there was no influence of fertilization or legume introduction on herbage iNDF and NFC contents (
p > 0.05) (
Table 1). Cubic and linear effects of the grazing cycle on the iNDF content of the mixed and control treatments were detected, respectively (
p = 0.0014).
The fraction A pool size was greater (
p < 0.05) in the mixed and control treatments when compared to the fertilized pastures (
Table 1). There was a quadratic grazing cycle effect in the control treatment (
p = 0.0017). The fraction B1 + B2 pool size was greater in the fertilized treatment (
p < 0.05) compared with the mixed treatment, but both were similar to the control (
Table 1). There was no grazing cycle effect on this variable. Nitrogen fertilization promoted greater concentrations of fraction B3 (
p < 0.05), compared to the mixed and control treatments (
Table 1). Linear (
p = 0.0072) and quadratic (
p = 0.0135) effects of grazing were found for mixed and control treatments, respectively.
Nitrogen fertilization caused lower contents of fraction C (
p < 0.05), compared to the mixed and control treatments (
Table 1). There was no grazing cycle effect on fraction C.
3.2. Chemical Composition of Herbage
Crude protein was 20% greater in the fertilized treatment (
p < 0.05), compared to the control, but it was similar in the mixed and control treatments. There was no effect of the grazing cycle on the CP contents (
Table 2).
Nitrogen fertilization affected the NDF content, of which the mean was lower than the control (
p < 0.05) (
Table 2). There was no grazing cycle effect on this variable. There was no difference in hemicellulose content of herbage (
p > 0.05) (
Table 2). Furthermore, fertilization or legume introduction did not alter ADF and lignin contents (
p > 0.05) (
Table 2). There was a quadratic effect of the grazing cycle on the ADF content in fertilized treatment (
p = 0.0002). Cellulose content was not affected by treatments or grazing cycles (
p > 0.05).
3.3. Herbage Accumulation Rate, Herbage Disappearance Rate, and Stocking Rate
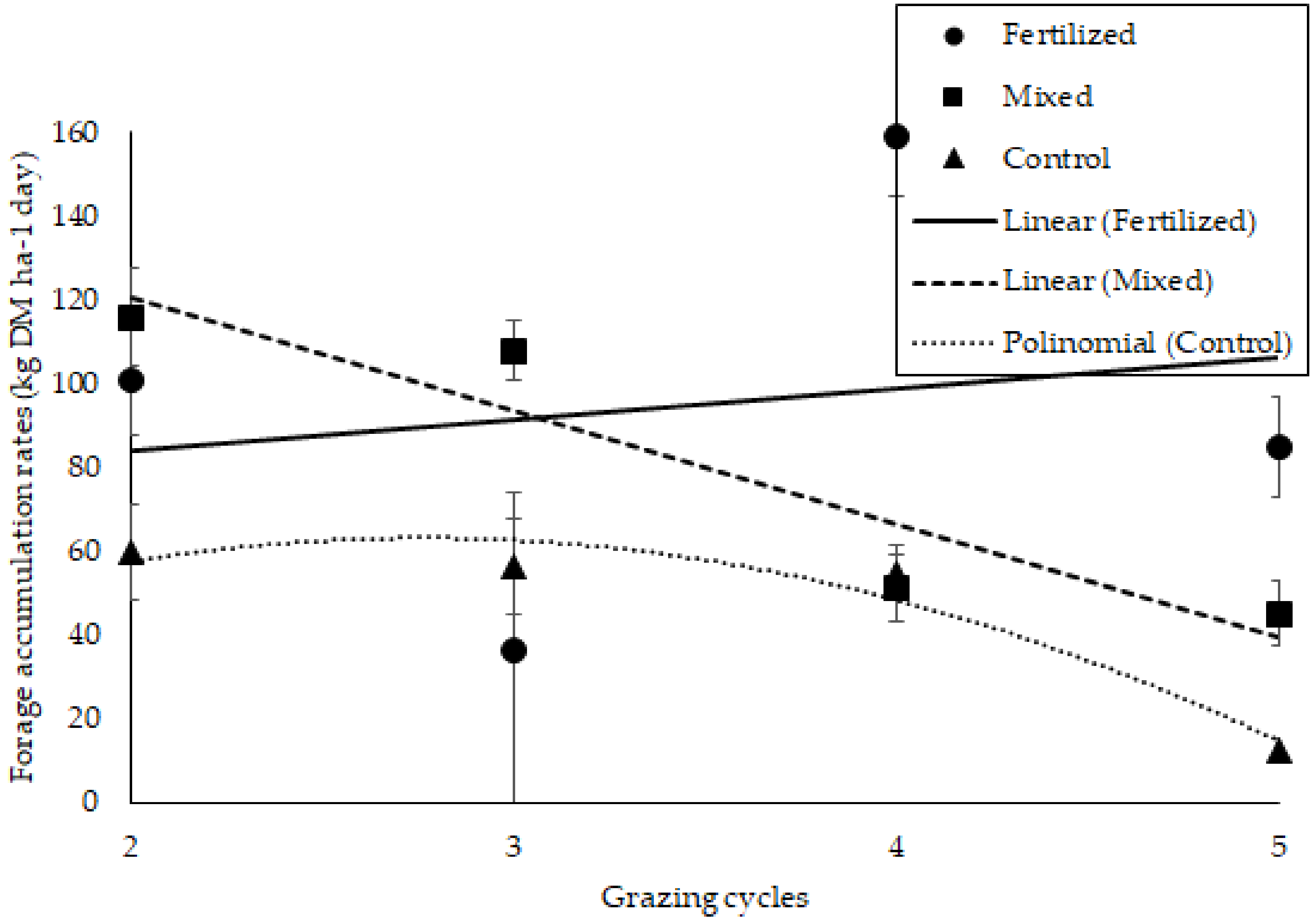
There was an influence of treatments and grazing cycles on HAR, which was greater in the fertilized treatment compared to the control (
p < 0.05), and both were similar to the mixed treatment (
Table 3). The grazing cycle effect was linear for fertilized and mixed treatments and quadratic for the control (
p = 0.0198) (
Figure 1).
Treatments and grazing cycles did not influence HDR (
p > 0.05) (
Table 3). The stocking rate was greater (
p < 0.05) in fertilized pastures in relation to mixed pastures, and both were similar to the control (
Table 3). The grazing cycle effect was linear for treatments (
p < 0.001).
3.4. Herbage Mass and Allowance
The pre-grazing HM was greater in the fertilized treatment compared to the mixed treatment (
p < 0.05), both of which did not differ from the control (
Table 4). Post-grazing HM was similar in all treatments (
p > 0.05). There was no grazing cycle effect on HM (
p > 0.05). Based on the proportion of pintoi peanut in the botanical composition of the mixed treatment (
Figure 2), there was 360 kg DM/ha of pintoi peanut in pre-grazing conditions and 191 kg DM/ha in post-grazing conditions. Additionally, herbage allowance was greater in fertilized and mixed pastures in relation to the control (
p < 0.05) (
Table 4). No cycle effect was observed for herbage allowance. The leaf allowance was greater in the fertilized treatment compared to the control (
p < 0.05), both of which did not differ from the mixed treatment (
Table 4). There was also no effect of the grazing cycle on the leaf allowance.
3.5. Botanical Composition of Mixed Pastures
On average, mixed pastures were composed of 91.2% grass and 8.8% legumes under pre-grazing conditions, and 94.1% and 5.9%, respectively, under post-grazing conditions (
Figure 2). There was no effect among cycles.
3.6. Morphological Composition
In the pre-grazing conditions, the proportion of leaves was greater in the fertilized and control treatments, compared to the mixed treatment (
p < 0.05) (
Table 5). On the other hand, in the post-grazing conditions, the leaf proportion was greater in the fertilized treatment (
p < 0.05) than the control, and both were similar to the mixed treatment (
Table 5).
The proportion of stem + sheath in the pre-grazing conditions was lower (
p < 0.05) in the mixed treatment compared to the fertilized treatment, and both were similar to the control (
Table 5). In post-grazing conditions, however, the proportion of stem + sheath of the control was similar to the mixed treatment, and both were lower than the fertilized treatment (
Table 5).
Regarding the proportion of dead material, in pre- and post-grazing conditions, the means were higher (
p < 0.05) in the mixed and control pastures, compared to fertilized pastures (
Table 5).
4. Discussion
4.1. Fractionation of Herbage Carbohydrate and Protein
Variations in TC and CP contents are related. Nitrogen fertilization increases protein synthesis from carbohydrates and the inclusion of legumes in tropical grass pastures has been shown to reduce TC content, mainly fibrous carbohydrate, i.e., pectin, cellulose, and hemicellulose [
30]. This was observed in our experiment, evidencing the potential efficiency of N arising from both chemical fertilization and biological fixation in providing similar CP and TC contents of herbage.
The TDN concentration results of our study differ from those of Benett et al. [
31], who reported a considerable increase in TDN with greater N fertilization, with a mean of 567 g/kg DM. Similar results to our study were observed in pastures of palisade grass fertilized with urea and managed at three different heights (15, 25, and 35 cm), which varied from 410 to 570 g/kg DM [
32].
However, different values of iNDF were found by Fonseca et al. [
33]. They reported 163 and 174 g/kg DM of iNDF in pastures managed at 30 and 45 cm pre-grazing, respectively, and 151 and 166 g/kg DM of iNDF in the non-fertilized pasture of mulatto grass II, also managed at these heights. Our study, in turn, showed greater values of iNDF in all treatments, which is probably related to the increase in the leaf proportion of herbage provided by the 95% LI management. Additionally, these authors found that a lower content of cell wall components, such as NDF, ADF, and iNDF, decreased the extent of fermentation in the rumen and, consequently, caused higher digestibility of the plant.
Regardless of the N source for fertilization, it was expected that fertilization with 150 kg N/ha/year via urea would present the greatest values of fraction A [
34]. This is due to the nitrate absorbed by the plant and reduced to ammonia, which is a substrate for amino acid and protein synthesis, increasing plant NPN contents [
9].
True protein tends to be extensively degraded in the rumen, contributing to the N supply of the ruminal microorganisms and incorporation into carbon skeletons [
13]. However, rapid rumen proteolysis of this fraction can lead to peptide construction and escape into the intestine, as the use of these components is limiting to protein degradation.
Fraction B1 + B2, which represents true proteins, increased with N fertilization (
Table 5), since it increased the N compounds of tissues and the protein synthesis of plants, being important in the formation and development of tillers and, consequently, in DM production [
35]. In this scenario, our study showed that the more N, the higher the production of new tillers and, consequently, the higher the content of fraction B.
4.2. Chemical Composition of Herbage
Several authors report that N from chemical fertilization raises the content of CP in DM by increasing soluble N compounds such as amino acids, nitrates, and peptides in their tissues and, consequently, enhancing the synthesis of soluble protein in the plant [
12,
36,
37]. The increment of CP promoted by N fertilization was also recorded by Ruggieri et al. and Delevatti et al. [
38,
39]. In the latter, the authors obtained averages of 136 and 151 g/kg in palisade grass fertilized with 90 and 180 kg N ha/year, results higher than those of our study.
Nonetheless, our study reported similar CP content of herbage in fertilized and mixed treatments, showing that the inclusion of pintoi peanut and N fertilization presents the same potential efficiency to increase herbage CP, mainly when compared to non-fertilized tropical pastures.
The NDF content of our study confirms the previous knowledge that the increase in N fertilizer applied promotes a linear reduction of NDF content [
31,
35,
40,
41]. Additionally, the NDF content was lower than the 600 g/kg DM that was suggested to restrict the herbage intake by grazing animals [
42], and this response to N can be considered a positive aspect of N fertilization.
The concentration of NDF in the plant can be reduced by N fertilization due to the stimulation of new tissue growth, which consequently reduces structural carbohydrate concentrations in DM [
43]. The greater production of new tissues provided by the N fertilization increased the leaf proportion and, consequently, decreased NDF content, which is mostly located in the stem, although it is found in a small proportion in the leaves.
The results of ADF and lignin contents are consistent with other studies that the application of N promotes minimal effects on the contents of ADF and lignin in herbage plants [
44,
45]. In our study, the pastures were managed in order to avoid stem elongation, which could control the increase in herbage lignin content.
The absence of differences in cellulose content may be associated with a variation in the hemicellulose components of fiber because NDF was affected by the treatments. Cellulose is most important for plant sustenance, and to increase CP content due to N fertilization, the amount of hemicellulose varied instead of cellulose.
4.3. Herbage Accumulation Rate, Forage Disappearance Rate, and Stocking Rate
The higher HAR in fertilized pastures is a consequence of the greater production of leaf blades and also of stems [
46]. Therefore, HAR is maximal at the 95% LI conditions [
47] and when canopies are managed under a higher herbage allowance, which maintains a higher leaf area index during the regrowth period.
The HDR is an important variable that assists in interpreting others, such as the estimation of DMI, especially when using rotational grazing, being directly related to the herbage allowance [
48]. Usually, the relationship between HDR and herbage allowance is linear; that is, the HDR increases with the increase in herbage allowance [
49]. In our study, however, the HDR was influenced neither by the herbage allowance (
Table 4) nor by treatments.
In this experiment, the management of animals followed the mob stocking technique [
16]. To adjust the 1-day occupation period and the 15 cm of residue height, we adjusted the stocking rate according to the pre-grazing conditions of each paddock. The average grazing height was 28 cm, which provided 50% grazing efficiency.
The fertilized treatment had the greatest pre-grazing herbage mass and, consequently, the greatest stocking rate (
Table 3). On the other hand, the control treatment showed the lowest pre-grazing herbage mass and stocking rate. Therefore, the similarity of HDR among treatments of our study may show that the stocking rate adjustment based on the pre-grazing herbage mass was correctly handled since there was no difference in the post-grazing mass among treatments.
4.4. Herbage Mass and Allowance
Nitrogen is the nutrient that most limits pasture productivity and also increases the production of new leaves and facilitates plant growth, which consequently enhances DM production [
6,
50]. Additionally, N stimulates tiller development since it is an important component of proteins and nucleic acids, which actively participate in the synthesis of organic compounds that form the plant structure [
51].
In our study, N fertilization increased the HM compared to the other treatments (
Table 1). The HM of the treatment that received 150 kg N/ha/year was close to 6350 kg/ha and 6440 kg/ha, under applications of 90 and 180 kg N/ha/year, respectively, measured in the same experimental area [
39].
In contrast to Cardoso et al. [
52], the contribution of N arising from biological nitrogen fixation (BNF) of legumes was insufficient to promote increases in pre-grazing HM, the mean of which was similar to the control. The average HM of mixed pastures was lower than the 9373 kg DM/ha found in a Cerrado region (Brazilian savanna) in palisade grass pastures mixed with
A. pintoi cv Mandobi managed at 95% LI, and the mass of legumes was 1924 kg DM/ha [
47], higher than measured in our research. In the conditions of the experiment of Pereira et al. [
47], pasture swards presented approximately 80% grass and 20% legumes, while only 8.8% was registered in our study (
Figure 2).
Herbage allowance followed the behavior of the HM of treatments, showing that pastures with higher pre-grazing HM provided a greater supply of herbage to the animals and vice versa. In all treatments studied, herbage allowance ranged from 5.9 to 8.2% BW, values lower than those found by Machado et al. [
53], between 8 and 12% BW. Usually, an animal consumes 2% of its BW in DM; therefore, herbage allowance was about four times the intake of the animals. This herbage allowance can promote a maximum intake of green leaves of the pasture and, consequently, maximum individual performance of grazing animals [
54]. Additionally, herbage allowance was above the minimum limit of 5% BW when it was insufficient to maintain a constant herbage availability to grazing animals [
55].
Nitrogen from the chemical fertilization provided a greater contribution and nutritional conditions favorable to the appearance and development of tillers. Additionally, N is the nutrient responsible for the production of new leaves, which explains the higher proportion of leaves obtained in the fertilized treatment [
56].
There was a similarity of leaf allowance in the fertilized and mixed treatments. This type of response suggests that, although the mixed treatment had lower pre- and post-grazing HM and a lower herbage allowance than the fertilized treatment, the N contribution promoted by the BNF of legumes was enough to provide greater leaf production by recycling or transfer of N to the grasses. The quality of available herbage for animals is directly influenced by higher leaf allowance [
57] due to the higher nutritional value and acceptability for grazing animals of this morphological component than stem tissue.
4.5. Botanical Composition of Mixed Pastures
In the first two cycles of grazing, there was a greater proportion of pintoi peanut before the grazing of the animals compared to the subsequent cycles and after their exit, which indicates a greater intake of the legume in these cycles (
Figure 2). Different botanical composition results were found by Pereira et al. [
47] in pastures of palisade grass mixed with
A. pintoi cv. Mandobi and managed at 95% LI, which were composed of 79.5% grass and 20.5% legumes in pre-grazing conditions, and 89.1% and 10.9%, respectively, in post-grazing conditions. In addition, mixed pastures were formed seven years prior to the evaluation in the experiment of Diehl et al. [
48], compared to 4 years in our research, which is a possible reason for the greater participation of pintoi peanut in the canopy.
The compatibility of legumes is determined by the growth habit and grazing management (defoliation) of pasture [
58]. The strategies of management of mixed pastures should aim at the regrowth capacity and the elasticity of the canopy structure in order for the recovery of both species to be vigorous and persistent after grazing. Due to the stoloniferous growth habit of pintoi peanut, the intake by animals can be tricky since the stolons in horizontal growth are close to the ground.
In our research, seeding of palisade grass and pintoi peanut was concomitantly done in November 2014 and, in order to enhance the legume stand in the mixed pastures, we planted the legume seedlings in the two subsequent years. Nonetheless, the low intake of pintoi peanut by grazing animals can be explained by its low proportion in sward (8%) and slow establishment [
5,
59,
60].
4.6. Morphological Composition
In pre-grazing conditions, all treatments presented a higher proportion of leaves compared to the stem + sheath and dead material in the produced herbage, components that are difficult to be grazed by animals; therefore, it can restrict the animal intake [
47]. A higher proportion of leaves in the herbage consumed by the animals leads to greater degradability and shorter retention time in the rumen, which, in the long term, allows greater intake [
61].
A possible explanation for the greater amount of stem + sheath in the fertilized treatment is that the higher sward height provided by the N fertilization, which created a canopy unfavorable to the maximum use of light and, therefore, it stimulated stem elongation as a plant strategy [
62]. Additionally, the lower percentage of stem + sheath in non-fertilized pastures is due to the shorter stem length and less tillering [
63].
In the mixed treatment, the averages of morphological composition were similar to Pereira et al. [
47], which, in pre-grazing conditions, found 36.2% leaves, 26.6% stem + sheath, and 37.2% dead material in pastures of palisade grass mixed with pintoi peanut, and managed at 95% LI. In the post-grazing conditions, the averages were 13.5% leaves, 30.3% stem + sheath, and 56.2% dead material.
5. Conclusions
Inclusion of pintoi peanut in tropical grass pastures increases soluble nitrogen compounds and total carbohydrate contents of plants, as well as leaf allowance and herbage accumulation rate, and decreases their slowly degraded protein content. On the other hand, N fertilization of palisade grass pastures increases the true protein content of herbage, herbage mass, and allowance for grazing animals, besides decreases indigestible protein and fiber contents of grass. Palisade grass pastures fertilized with 150 kg N/ha/year, mixed with pintoi peanut or without N fertilization managed at the height equivalent to 95% of LI, with 15 cm of residue height and at a grazing efficiency of 50%, can prevent plants from accumulating structural carbohydrates such as cellulose and hemicellulose, as it reduces the production of fibrous tissues, and this management can avoid the degradation of the pastures.
Author Contributions
Conceptualization, A.S.B., A.d.S.C. and A.C.R.; methodology, A.S.B. and V.Z.L.; software, validation, and formal analysis, A.S.B.; investigation, A.S.B.; resources, A.S.B.; data curation, A.S.B. and A.d.S.C.; writing—original draft preparation, A.S.B.; writing—review and editing, A.d.S.C., L.O.T. and R.A.R.; visualization, R.A.R. and R.M.B.; supervision, A.C.R.; project administration, A.C.R. and R.M.B.; funding acquisition, A.C.R. and R.M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the São Paulo Research Foundation (FAPESP, grant n° 2015/16631-5; 2016/11086-1; 2017/11274-5); the Coordination of Improvement of Higher Education Personnel (CAPES); and the National Council for Scientific and Technological Development (CNPq, grant nº 404169/2013-9).
Institutional Review Board Statement
The study was approved by the Ethics Committee of UNESP (protocol code 10356/14 and 7/7/2014).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are not publicly available due to privacy or ethical reasons.
Acknowledgments
The authors thank the members of Unesp Jaboticabal Forage Team (UnespFOR) for the contributions during the field trial setup.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Allemu, A.W.; Kröbel, R.; McConkey, B.G.; Iwaasa, A.D. Effect of Increasing Species Diversity and Grazing Management on Pasture Productivity, Animal Performance, and Soil Carbon Sequestration of Re-Established Pasture in Canadian Prairie. Animals 2019, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, A.S.; Berndt, A.; Leytem, A.; Alves, B.J.; de Carvalho, I.D.N.; de Barros Soares, L.H.; Urquiaga, S.; Boddey, R.M. Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use. Agric. Syst. 2016, 143, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, H.L.B.; Pedreira, B.C.; Sollenberger, L.E.; Pereira, D.H.; Magalhães, C.A.S.; Chizzotti, F.H.M. Herbage accumulation, canopy structure and tiller morphology of marandu palisadegrass growing in open pasture and in silvopasture. Agrofor. Syst. 2021, 95, 1–14. [Google Scholar] [CrossRef]
- Boddey, R.M.; Macedo, R.; Tarré, R.M.; Ferreira, E.; de Oliveira, O.C.; Rezende, C.P.; Cantarutti, R.B.; Pereira, J.M.; Alves, B.J.R.; Urquiaga, S. Nitrogen cycling in Brachiaria pastures: The key to understanding the process of pasture decline. Agr. Ecosyst. Environ. 2004, 103, 389–403. [Google Scholar] [CrossRef]
- Alencar, N.M.; Vendramini, J.M.B.; dos Santos, A.C.; Silveira, M.L.; Dubeux, J.C.B., Jr.; Sousa, L.F.; Neiva, J.N.M. Herbage Characteristics of forage peanut and Palisadegrass Established as Monoculture or Mixed Swards. Crop Sci. 2018, 58, 2131–2137. [Google Scholar] [CrossRef]
- Machado, V.D.; da Fonseca, D.M.; Lima, M.A.; Martuscello, J.A.; Paciullo, D.S.; Chizzotti, F.H. Grazing management strategies for Urochloa decumbens (Stapf) R. Webster in a silvopastoral system under rotational stocking. Grass Forage Sci. 2020, 75, 266–278. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Molle, A.; Menendez, H.M.; Cannas, A.; Fonseca, M.A. The assessment of supplementation requirements of grazing ruminants using nutrition models. Transl. Anim. Sci. 2019, 3, 1–18. [Google Scholar]
- Muir, J.P.; Tedeschi, L.O.; Dubeux, J.C.B.; Peters, M.; Burkart, S. Enhancing food security in Latin America with forage legumes. Arch. Latinoam. Prod. Anim. 2017, 25, 113–130. [Google Scholar]
- Gomes, F.K.; Oliveira, M.D.; Homem, B.G.; Boddey, R.M.; Bernardes, T.F.; Gionbelli, M.P.; Casagrande, D.R. Effects of grazing management in brachiaria grass-forage peanut pastures on canopy structure and forage intake. J. Anim. Sci. 2018, 96, 3837–3849. [Google Scholar] [CrossRef]
- Pereira, J.M.; Rezende, C.D.P.; Ferreira Borges, A.M.; Homem, B.G.C.; Casagrande, D.R.; Macedo, T.M.; Boddey, R.M. Production of beef cattle grazing on Brachiaria brizantha (Marandu grass)—Arachis pintoi (forage peanut cv. Belomonte) mixtures exceeded that on grass monocultures fertilized with 120 kg N/ha. Grass Forage Sci. 2020, 75, 28–36. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Fox, D.G. The Ruminant Nutrition System: Volume I-An Applied Model for Predicting Nutrient Requirements and Feed Utilization in Ruminants, 3rd ed.; XanEdu: Ann Arbor, MI, USA, 2020. [Google Scholar]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Sniffen, C.J.; O’Connor, J.D.; Van Soest, P.J.; Fox, D.G.; Russell, J.B. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 1992, 70, 3562–3577. [Google Scholar] [CrossRef]
- Embrapa-Empresa Brasileira De Pesquisa Agropecuária. Sistema Brasileiro de Classificação de Solos, 3rd ed.; EMBRAPA: Brasília, Brazil, 2013. [Google Scholar]
- Berça, A.S.; Cardoso, A.S.; Longhini, V.Z.; Tedeschi, L.O.; Boddey, R.M.; Berndt, A.; Reis, R.A.; Ruggieri, A.C. Methane production and nitrogen balance of dairy heifers grazing palisade grass cv. Marandu alone or with forage peanut. J. Anim. Sci. 2019, 97, 4625–4634. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; Mclvor, J.; Morris, C.; Peeters, A.; Sanderson, M. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Van Raij, B. Recommendations of Fertilization and Liming in São Paulo State; IAC: Campinas, Brazil, 1997. [Google Scholar]
- Halls, L.K. The Approximation of Cattle Diet through Herbage Sampling. Rangel. Ecol. Manag. 1954, 7, 7269–7270. [Google Scholar] [CrossRef] [Green Version]
- ANKOM. Neutral Detergent Fiber in Feeds. Filter Bag Technique (for A200, A200I). Ankom Technology Method 9; Ankom Technology Corp: Macedon, NY, USA, 2006. [Google Scholar]
- ANKOM. Acid Detergent Fiber in Feeds. Filter Bag Technique (for A200, A200I). Ankom Technology Method 8; Ankom Technology Corp: Macedon, NY, USA, 2006. [Google Scholar]
- ANKOM. Method for Determining Acid Detergent Lignin in Beakers; Ankom Technology Corp: Macedon, NY, USA, 2006. [Google Scholar]
- AOAC. AOAC official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Van Soest, P.J.; Van Amburgh, M.E.; Tedeschi, L.O. Rumen balance and rates of fiber digestion. In Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, Rochester, NY, USA, 19–21 October 2010; New York State College of Agriculture and Life Sciences, Cornell University: Ithaca, NY, USA, 2000; pp. 150–166. [Google Scholar]
- Licitra, G.; Hernandez, T.M.; Van Soest, P.J. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. Tech. 1996, 57, 347–358. [Google Scholar] [CrossRef]
- Barthram, G.T. Experimental Techniques: The HFRO Sward Stick; Biennial Report; Hill Farming Research Organization (HFRO): Midlothian, Scotland, 1985; pp. 29–30. [Google Scholar]
- Campbell, A.G. Grazed pasture parameters. I. Pasture dry-matter production and availability in a stocking rate and grazing management experiment with dairy cows. J. Agric. Sci. 1966, 67, 199–210. [Google Scholar] [CrossRef]
- Cecato, U.; Castro, C.D.C.; Canto, M.D.; Peternelli, M.; Almeida Júnior, J.; Jobim, C.C.; Cano, C.C.P. Forage losses in tanzania grass (Panicum maximum Jacq cv. Tanzania-1) managed in different sward height under grazing. Rev. Bras. Zootec. 2001, 30, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Gimenes, F.M.A.; Silva, S.C.; Fialho, C.A.; Gomes, M.B.; Berndt, A.; Gerdes, L.; Colozza, M.T. Weight gain and animal productivity on Marandu palisade grass under rotational stocking and nitrogen fertilization. Pesqui. Agropecu. Bras. 2011, 46, 751–759. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2018. Available online: https://www.R-project.org/ (accessed on 8 January 2019).
- De Oliveira, J.K.; Corrêa, D.C.C.; Cunha, A.M.; Rêgo, A.C.D.; Faturi, C.; da Silva, W.L.; Domingues, F.N. Effect of Nitrogen Fertilization on Production, Chemical Composition and Morphogenesis of Guinea Grass in the Humid Tropics. Agron. J. 2020, 10, 1840. [Google Scholar]
- Benett, C.G.S.; Buzetti, S.; Silva, K.S.; Bergamaschine, A.F.; Fabricio, J.A. Productivity and quality composition of Marandu grass to sources and doses of nitrogen. Ciênc. Agrotec. 2008, 32, 1629–1636. [Google Scholar] [CrossRef]
- Delevatti, L.M.; Romanzini, E.P.; Koscheck, J.F.W.; de Araujo, T.L.D.R.; Renesto, D.M.; Ferrari, A.C.; Reis, R.A. Forage management intensification and supplementation strategy: Intake and metabolic parameters on beef cattle production. Anim. Feed Sci. Technol. 2018, 247, 74–82. [Google Scholar] [CrossRef]
- Fonseca, A.S.R.; Silva, L.C.R.P.; Borges, K.M.; da Silva, S.M.V.; Vargas, L.E.D.P.; de Lima, L.R.; Galati, R.L. Digestion kinetics of mulato ii and marandu grasses at different canopy heights. In Proceedings of the 55a Reunião Anual da Sociedade Brasileira de Zootecnia, 28° Congresso Brasileiro de Zootecnia, Goiânia, Brasil, 27–30 August 2018. [Google Scholar]
- Henriques, L.T.; da Silva, J.C.; Detmann, E.; Vásquez, H.M.; Pereira, O.G. Fractions of nitrogenous compounds of four tropical grasses in the different cutting ages and nitrogen fertilizer levels. Arq. Bras. Med. Vet. Zootec. 2007, 59, 740–748. [Google Scholar] [CrossRef]
- Johnson, C.R.; Reiling, B.A.; Mislevy, P.; Hall, M.B. Effects of N fertilization and harvest date on yield, digestibility, fiber, and protein fractions of tropical grasses. J. Anim. Sci. 2001, 79, 2439–2448. [Google Scholar] [CrossRef]
- Loaiza, P.A.; Balocchi, O.; Bertrand, A. Carbohydrate and crude protein fractions in perennial ryegrass as affected by defoliation frequency and nitrogen application rate. Grass Forage Sci. 2017, 72, 556–567. [Google Scholar] [CrossRef]
- Solati, Z.; Jørgensen, U.; Eriksen, J.; Søegaard, K. Dry matter yield, chemical composition and estimated extractable protein of legume and grass species during the spring growth. J. Sci. Food Agric. 2017, 97, 3958–3966. [Google Scholar] [CrossRef] [Green Version]
- Ruggieri, A.C.; Cardoso, A.S.; Ongaratto, F.; Casagrande, D.R.; Barbero, R.P.; Brito, L.F.; Azenha, M.V.; Oliveira, A.A.; Koscheck, J.F.W.; Reis, R.A. Grazing Intensity Impacts on Herbage Mass, Sward Structure, Greenhouse Gas Emissions, and Animal Performance: Analysis of Brachiaria Pastureland. Agron. J. 2020, 10, 1750. [Google Scholar] [CrossRef]
- Delevatti, L.M.; Cardoso, A.S.; Barbero, R.P.; Leite, R.G.; Romanzini, E.P.; Ruggieri, A.C.; Reis, R.A. Effect of nitrogen application rate on yield, forage quality, and animal performance in a tropical pasture. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Primavesi, A.C.; Primavesi, O.; Corrêa, L.D.A.; Cantarella, H.; da Silva, A.G.; Freitas, A.R.; Vivaldi, L.J. Nitrogen fertilization on coastcross grass: Effects on nutrient extraction and apparent nitrogen recovery. Rev. Bras. Zootec. 2004, 33, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Dupas, E.; Buzetti, S.; Sarto, A.L.; Hernandez, F.B.T.; Bergamaschine, A.F. Dry matter yield and nutritional value of Marandu grass under nitrogen fertilization and irrigation in cerrado in São Paulo. Rev. Bras. Zootec. 2010, 39, 2598–2603. [Google Scholar] [CrossRef] [Green Version]
- Poppi, D.P.; Vega, A. Extent of digestion and rumen condition as factors affecting passage of liquid and digesta particles in sheep. J. Agric. Sci. 1997, 128, 207–215. [Google Scholar]
- Santini, J.M.K.; Perin, A.; Coaguila, D.N.; Valderrama, M.; Dupas, E.; dos Santos, C.G.; Buzetti, S. Nitrogen fertilization in implementation of Urochloa brizantha cv. Xaraés in Brazilian Savanna: Nutritional characteristics-Part 2. Braz. J. Chem. Eng. 2016, 10, 140–153. [Google Scholar]
- Cuomo, G.J.; Anderson, B.E. N fertilization and burning effects on rumen protein degradation and nutritive value of native grasses. Agron. J. 1996, 88, 439–442. [Google Scholar] [CrossRef]
- Gándara, L.; Borrajo, C.I.; Fernández, J.A.; Pereira, M.M. Effect of nitrogen fertilization and the age of regrowth on the nutritional value of Brachiaria brizantha cv. “Marandu”. Rev. Fac. Cienc. Agrar. 2017, 49, 69–77. [Google Scholar]
- Carvalho, P.C.D.F.; Kozloski, G.V.; Nabinger, C.; Ribeiro Filho, H.M.N.; Reffatti, M.V.; Genro, T.C.M.; Euclides, V.P.B. Methodological advances in the determination of the consumption of ruminants under grazing. Rev. Bras. Zootec. 2006, 36, 151–170. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.C.; Gomes, F.K.; Oliveira, M.D.; Lara, M.A.; Bernardes, T.F.; Casagrande, D.R. Defoliation management affects morphogenetic and structural characteristics of mixed pastures of Brachiaria grass and forage peanut. Afr. J. Range Forage Sci. 2017, 34, 13–19. [Google Scholar] [CrossRef]
- Diehl, M.S.; Olivo, C.J.; Agnolin, C.A.; Bratz, V.F.; Bem, C.M.; Agüirre, P.F.; Serafim, G. Productivity of pastures-based systems mixed to forage peanut or red clover. Arq. Bras. Med. Vet. Zootec. 2013, 65, 1527–1536. [Google Scholar] [CrossRef]
- Braga, G.J.; Pedreira, C.G.S.; Herling, V.R.; de Cerqueira Luz, P.H. Grazing efficiency on Marandu palisadegrass as affected by herbage allowance. Pesqui. Agropec. Bras. 2007, 42, 1641–1649. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.A.; Almeida, E.X.; Raupp, G.T.; Miguel, M.F.; de Liz, D.M.; Carvalho, P.C.F.; Ribeiro-Filho, H.M.N. Herbage intake, methane emissions and animal performance of steers grazing dwarf elephant grass v. dwarf elephant grass and peanut pastures. Animals 2016, 10, 1684–1688. [Google Scholar] [CrossRef] [PubMed]
- Malavolta, E. Manual de Nutrição Mineral de Plantas; Ceres: São Paulo, Brazil, 2006; 638p. [Google Scholar]
- Cardoso, A.S.; Barbero, R.P.; Romanzini, E.P.; Teobaldo, R.W.; Ongaratto, F.; Fernandes, M.H.M.R.; Ruggieri, A.C.; Reis, R.A. Intensification: A key strategy to achieve great animal and environmental beef cattle production sustainability in Brachiaria grasslands. Sustainability 2020, 12, 6656. [Google Scholar] [CrossRef]
- Machado, L.A.Z.; Fabrício, A.C.; Gomes, A.; De Assis, P.G.G.; Lempp, B.; Maraschin, G.E. Performance of animals fed with green leaf in Marandu grass pasture. Pesqui. Agropecu. Bras. 2008, 43, 1609–1616. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J. Sward conditions, herbage allowance and animal production: An evaluation of research results. Proc. N. Zeal. Soc. Anim. Prod. 1984, 44, 99–104. [Google Scholar]
- Schio, A.R.; Mattos Veloso, C.; Ferreira Silva, F.; Vinhas Ítavo, L.C.; Gonçalves Mateus, R.; Rodrigues Silva, R. Forage on offer for Nellore heifers supplemented during the drought and dry/rainy transition seasons. Acta Sci. 2011, 33, 9–18. [Google Scholar]
- Martuscello, J.A.; Ribeiro, Y.N.; Braz, T.G.S.; Ferreira, M.R.; Assis, J.A.; Jank, L.; Reis, G.A. Forage production, morphogenesis and agronomic efficiency in Panicum maximum BRS Quênia under nitrogen levels. Bol. Ind. Anim. 2018, 75, 12. [Google Scholar]
- Casagrande, D.R.; Ruggieri, A.C.; Moretti, M.H.; Berchielli, T.T.; Vieira, B.R.; de Toledo Piza Roth, A.P.; Reis, R.A. Sward canopy structure and performance of beef heifers under supplementation in Brachiaria brizantha cv. Marandu pastures maintained with three grazing intensities in a continuous stocking system. Rev. Bras. Zootec. 2011, 40, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Casagrande, D.R.; Lara, M.A.S.; Vieira, B.R. Legumes of tropical and subtropical climate. In Forragicultura: Ciência, Tecnologia e Gestão dos Recursos Forrageiros, 2nd ed.; Funep: Jaboticabal, Brazil, 2014; pp. 137–154. [Google Scholar]
- Cook, B.G.; Jones, R.M.; Williams, R.J. Regional Experience with Forage Arachis in Australia. Biology and Agronomy of Forage Arachis; CIAT: Cali, Colombia, 1994; Volume 1, pp. 158–168. [Google Scholar]
- Sanchez, J.M.; Vendramini, J.M.; Silveira, M.L.; Sollenberger, L.E.; Dubeux, J.C., Jr.; Moriel, P.; Sellers, B.; Yarborough, J.K.; Leite de Oliveira, F.C. Seeding strategies of bahiagrass and forage peanut affect pasture establishment under weed competition. Grass Forage Sci. 2018, 74, 381–388. [Google Scholar] [CrossRef]
- Poppi, D.P.; Hughes, T.P.; L’huillier, P.J. Intake of pasture by grazing ruminants. In Livestock Feeding on Pasture; Nicol, A.M., Ed.; New Zealand Society of Animal Production: Hamilton, NY, USA, 1997; Volume 1, pp. 55–64. [Google Scholar]
- Galzerano, L.; Malheiros, E.B.; Raposo, E.; Morgado, E.S.; Ruggieri, A.C. Accumulation and disappearance of forage and changes in the pasture structure of Xaraes palisade grass subjected to grazing intensities and intermittent stocking. Semin. Cienc. Agrar. 2013, 34, 2485–2496. [Google Scholar] [CrossRef] [Green Version]
- Alexandrino, E.; do Nascimento Júnior, D.; Regazzi, A.J.; Mosquim, P.R.; Cipriano Rocha, F.; de Paula Souza, D. Morphogenic and structural traits of Brachiaria brizantha cv. Marandu submitted to different levels of Nitrogen and frequency harvest. Acta Sci. Agron. 2005, 27, 17–24. [Google Scholar]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


 ,
, 


{kind=link}
{kind=link}