
Effect of Bacillus spp. on Lettuce Growth and Root Associated Bacterial Community in a Small-Scale Aquaponics System



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Location
2.3. System Set Up and Operation
2.4. Biofilter Establishment
2.5. Fish Rearing Conditions
2.6. Lettuce Conditions
2.7. Data Collection
2.8. Water and Leaf Mineral Analysis
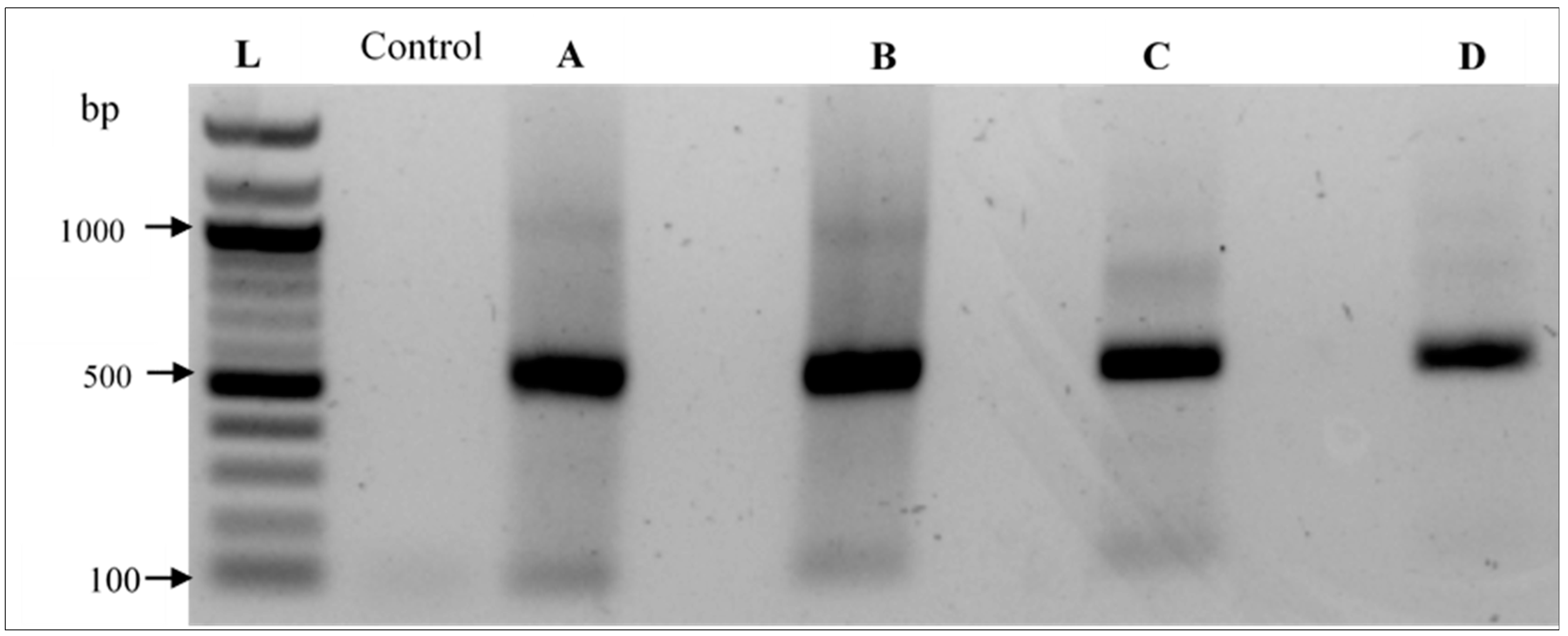
2.9. DNA Extraction and PCR Amplification
2.10. Statistical Analysis and Bioinformatics Processing
3. Results
3.1. Water Quality Parameters in Fish Rearing Tanks
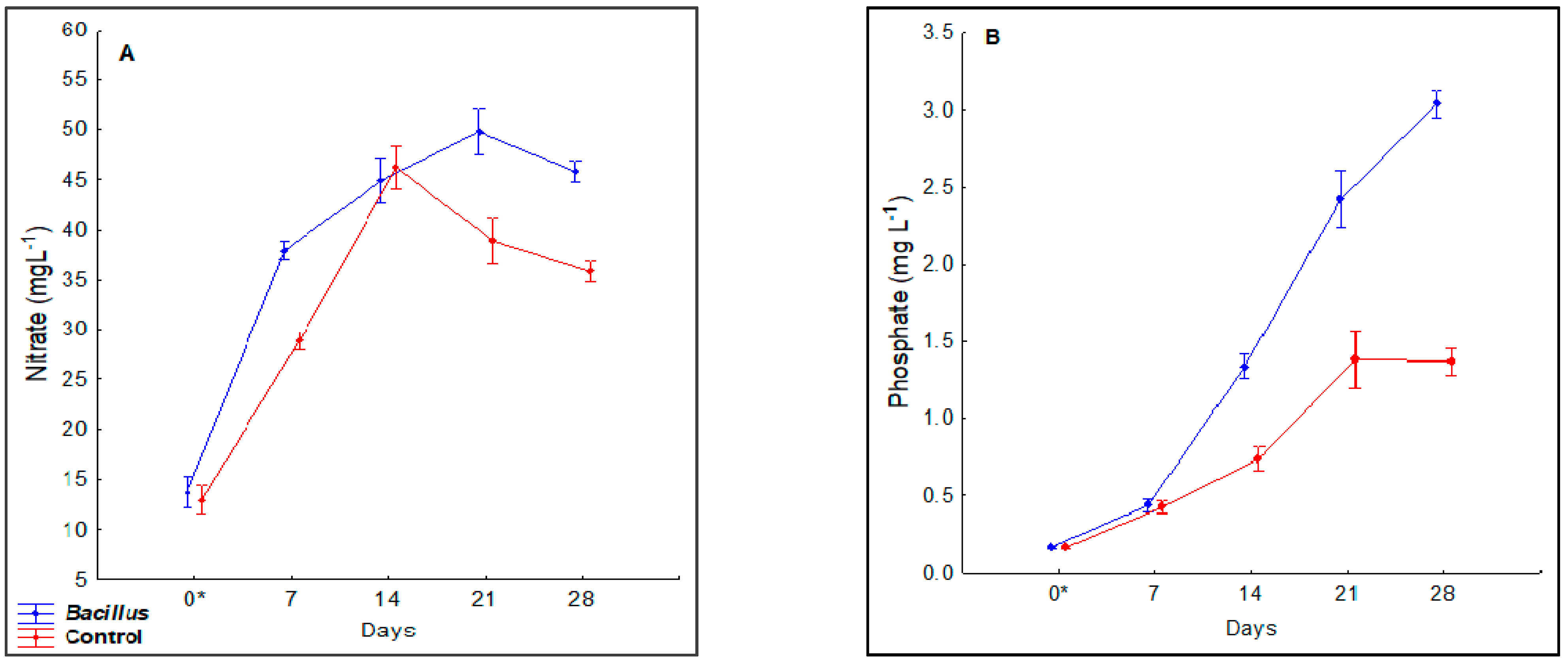
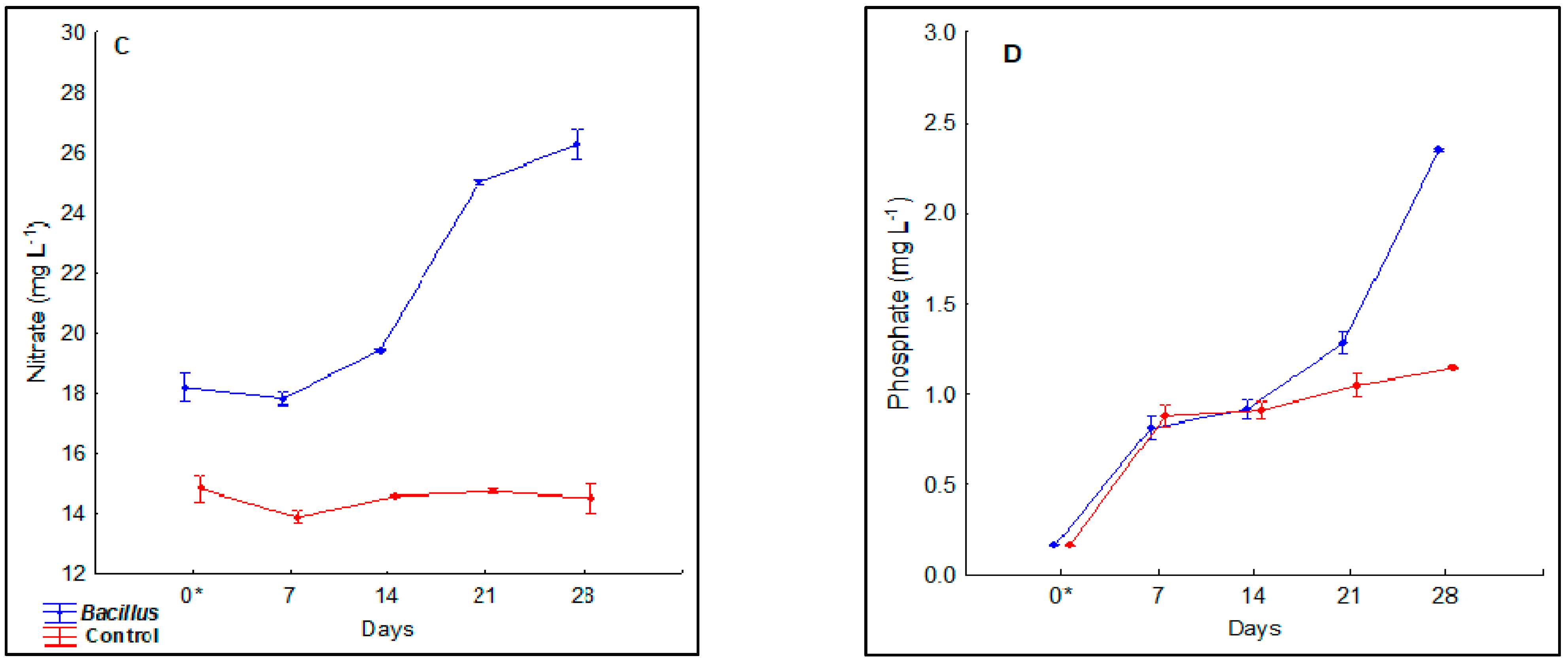
3.2. Nitrate and Phosphate Concentration Dynamics in the Deep-Water Culture Solution
3.3. Lettuce Growth and Chlorophyll Concentration
3.4. Water Chemical Analysis and Lettuce Nutrition
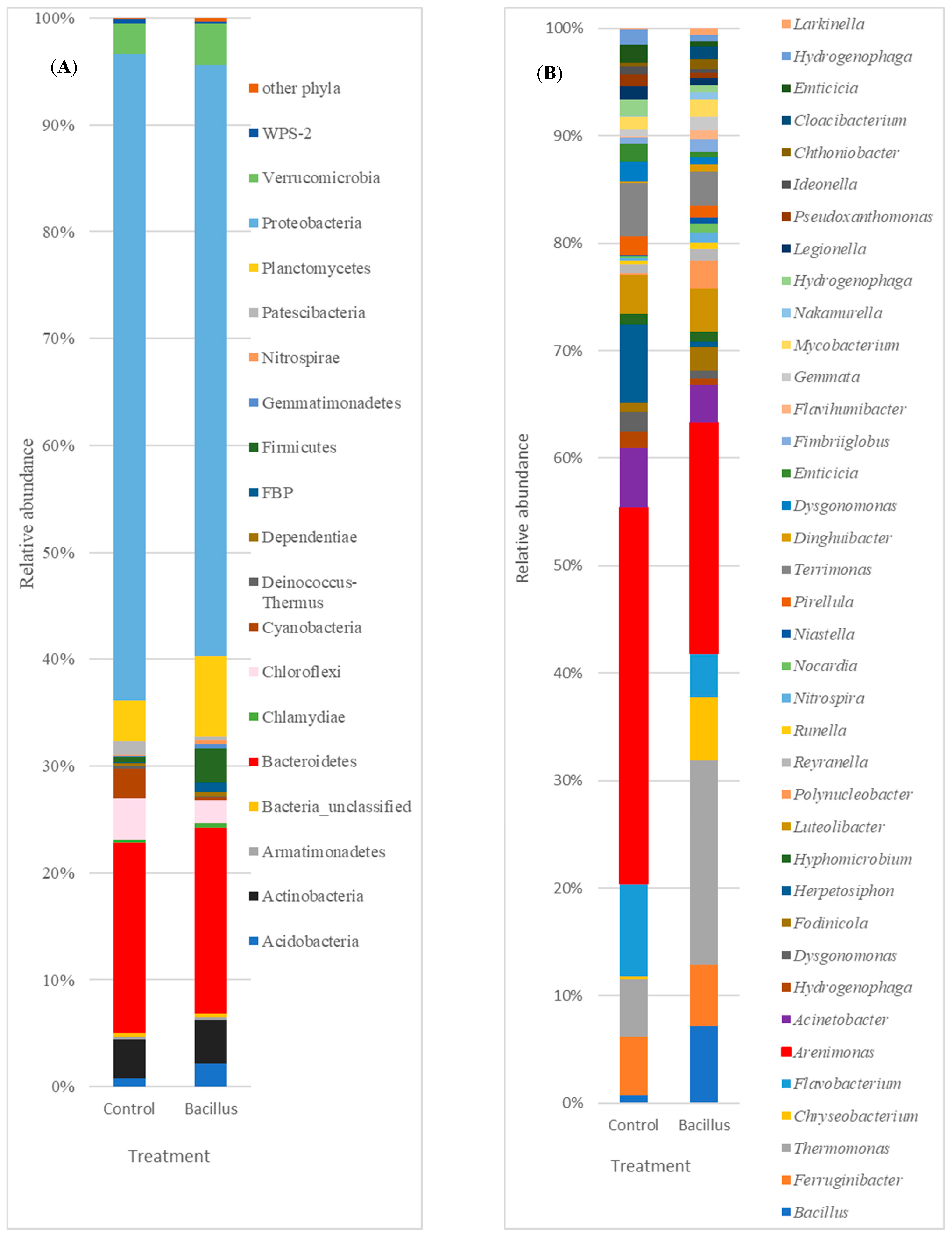
3.5. Taxonomic Assignment of Reads
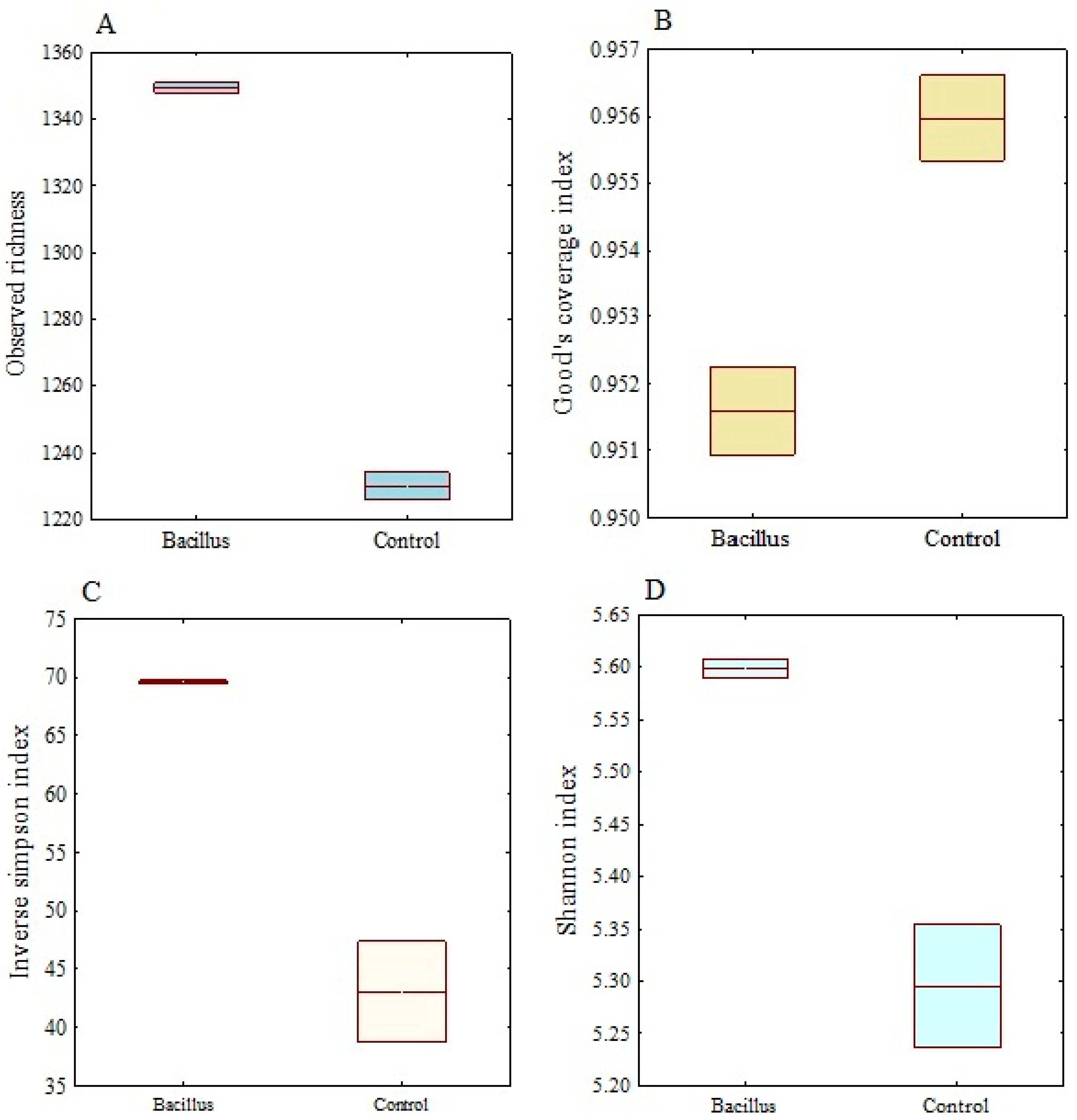
3.6. Alpha Diversity and Rarefaction
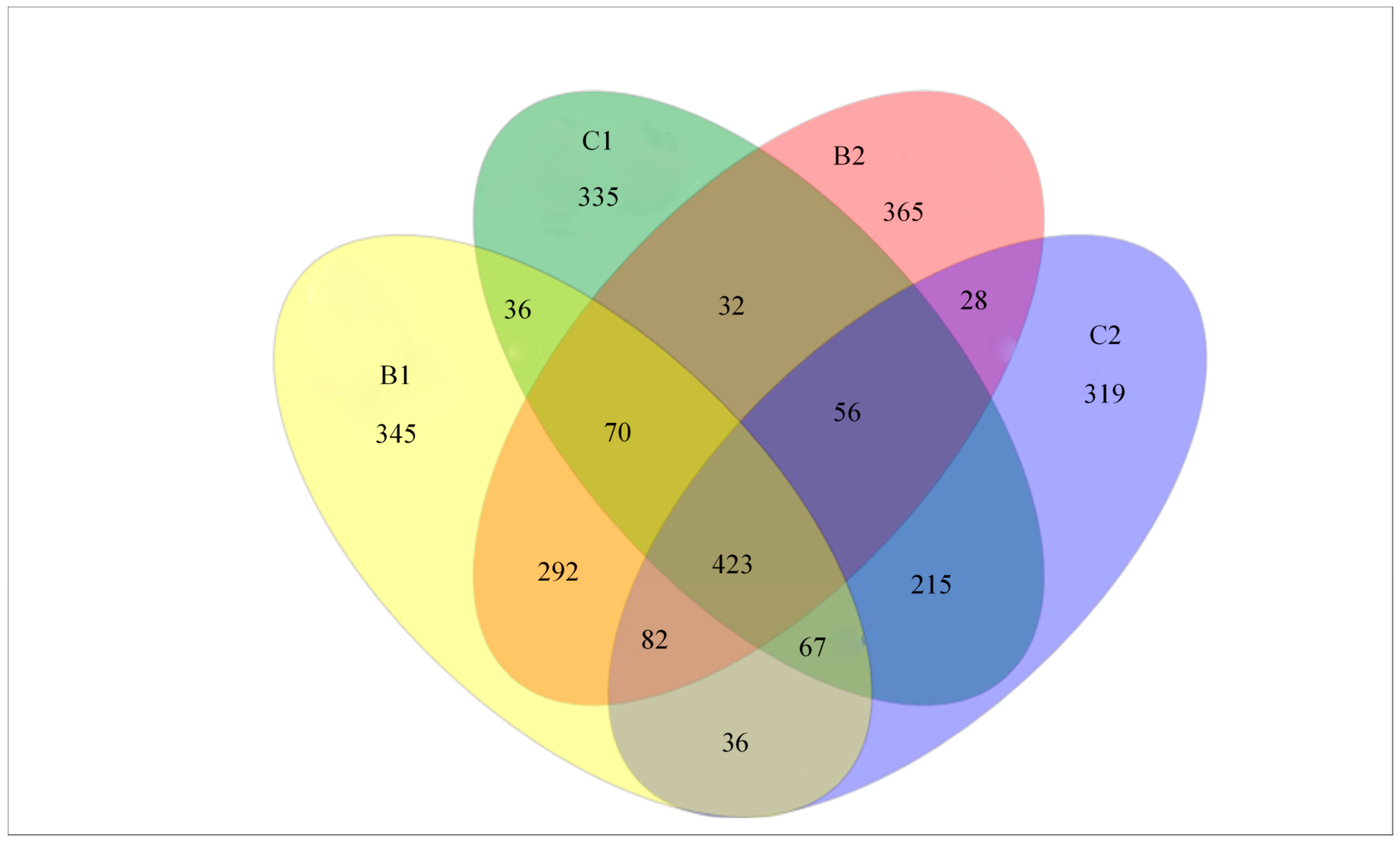
3.7. Common and Unique Bacterial Communities from Replicate Systems
4. Discussion
4.1. Water Quality Management in Fish Rearing Tanks
4.2. Lettuce Growth
4.3. Nutrient Accumulation in Lettuce and Water
4.4. Dissolved Nitrate and Phosphate in the Deep-Water Culture Growth Beds
4.5. Root Associated Core Microbiota
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rakocy, J.E.; Masser, M.P.; Losordo, T.M. Recirculating Aquaculture Tank Production Systems: Aquaponics—Integrating Fish and Plant Culture; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2006; pp. 1–16. [Google Scholar]
- Kasozi, N.; Tandlich, R.; Fick, M.; Kaiser, H.; Wilhelmi, B. Iron supplementation and management in aquaponic systems: A review. Aquac. Rep. 2019, 15, 1–10. [Google Scholar] [CrossRef]
- Joyce, A.; Timmons, M.; Goddek, S.; Pentz, T. Bacterial Relationships in Aquaponics: New Research Direction. In Aquaponics Food Production Systems; Goddek, S., Joyce, A., Kotzen, B., Burnell, G.M., Eds.; Springer: Cham, Switzerland, 2019; pp. 145–161. [Google Scholar]
- Kasozi, N.; Abraham, B.; Kaiser, H.; Wilhelmi, B. The complex microbiome in aquaponics: Significance of the bacterial ecosystem. Ann. Microbiol. 2021, 71, 1–13. [Google Scholar] [CrossRef]
- Gatesoupe, F.J. The use of probiotics in aquaculture. Aquaculture 1999, 180, 147–165. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Irianto, A.; Austin, B. Probiotics in aquaculture. J. Fish Dis. 2002, 25, 633–642. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living Rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Cerozi, B.D.S.; Fitzsimmons, K. Use of Bacillus spp. to enhance phosphorus availability and serve as a plant growth promoter in aquaponics systems. Sci. Hortic. 2016, 211, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Bartelme, R.P.; Oyserman, B.O.; Blom, J.E.; Sepulveda-Villet, O.J.; Newton, R.J. Stripping away the Soil: Plant growth promoting microbiology opportunities in aquaponics. Front. Microbiol. 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahir, A.A.; Arshad, M.; Frankenberger, W.T. Plant growth promoting rhizobacteria: Applications and perspectives in agriculture. Adv. Agron. 2004, 81, 97–168. [Google Scholar]
- Martínez-Viveros, O.; Jorquera, M.A.; Crowley, D.E.; Gajardo, G.; Mora, M.L. Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Lareen, A.; Burton, F.; Schafer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Sallenave, R. Important Water Quality Parameters in Aquaponics Systems; New Mexico State University: Las Cruces, NM, USA, 2016; Volume 680, pp. 1–8. [Google Scholar]
- Emerson, K.; Russo, R.C.; Lund, R.E.; Thurston, R.V. Aqueous ammonia equilibrium calculations: Effect of pH and temperature. J. Fish. Res. Board Can. 1975, 32, 2379–2383. [Google Scholar] [CrossRef]
- Kalra, Y.P. Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1998; pp. 165–170. [Google Scholar]
- De Winter, J.C.F. Using the Student’s t-test with extremely small sample sizes. Pract. Asses. Res. Eval. 2013, 18, 1–12. [Google Scholar]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, platform-independent, community supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open-source tool for metagenomics. PeerJ 2016, 4, e2409v1. [Google Scholar] [CrossRef]
- Somerville, C.; Cohen, M.; Pantanella, E.; Stankus, A.; Lovatelli, A. Small-Scale Aquaponic Food Production. Integrated Fish and Plant Farming; Technical Paper No. 589; FAO Fisheries and Aquaculture: Rome, Italy, 2014; pp. 133–134. [Google Scholar]
- DeLong, D.P.; Losordo, T.M.; Rakocy, J.E. Tank Culture of Tilapia; Southern Regional Aquaculture Centre: Stoneville, MS, USA, 2009; p. 282. [Google Scholar]
- Reyes Yanes, A.; Martinez, P.; Ahmad, R. Towards automated aquaponics: A review on monitoring, IoT, and smart systems. J. Clean. Prod. 2020, 263, 1–15. [Google Scholar]
- Stone, N.M.; Thomforde, H.K. Understanding Your Fishpond Water Analysis Report; University of Arkansas: Pine Bluff, AR, USA, 2004. [Google Scholar]
- Tyson, R.V.; Treadwell, D.D.; Simonne, E.H. Opportunities and challenges to sustainability in aquaponic systems. HortTechnology 2011, 21, 6–13. [Google Scholar] [CrossRef]
- Lennard, W.; Goddek, S. Aquaponics: The basics. In Aquaponics Food Production Systems; Goddek, S., Joyce, A., Kotzen, B., Burnell, G.M., Eds.; Springer: Cham, Switzerland, 2019; pp. 129–131. [Google Scholar]
- Yildiz, H.Y.; Robaina, L.; Pirhonen, J.; Mente, E.; Domínguez, D.; Parisi, G. Fish welfare in aquaponic systems: Its relation to water quality with an emphasis on feed and faeces—A review. Water 2017, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, E.; Timmons, M.B.; Mattson, N.S. Quality, yield, and biomass efficacy of several hydroponic lettuce (Lactuca sativa L.) cultivars in response to high pressure sodium lights or light emitting diodes for greenhouse supplemental lighting. Horticulturae 2020, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- García, J.A.L.; Probanza, A.; Ramos, B.; Palomino, M.R.; Manero, F.J.G. Effect of inoculation of Bacillus licheniformis on tomato and pepper. Agronomie 2004, 24, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Niazi, A.; Manzoor, S.; Asari, S.; Bejai, S.; Meijer, J.; Bongcam-Rudloff, E.; Gijzen, M. Genome analysis of Bacillus amyloliquefaciens subsp. plantarum UCMB5113: A Rhizobacterium that improves plant growth and stress management. PLoS ONE 2014, 9, e104651. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative analysis of the complete genome sequence of the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Mumtaz, M.Z.; Ahmad, M.; Jamil, M.; Hussain, T. Zinc solubilizing Bacillus spp potential candidates for biofortification of maize. Microbiol. Res. 2017, 202, 51–60. [Google Scholar] [CrossRef]
- Graber, A.; Junge, R. Aquaponic systems: Nutrient recycling from fish wastewater by vegetable production. Desalination 2009, 246, 147–156. [Google Scholar] [CrossRef]
- Asher, C.J.; Loneragan, J.F. Response of plants to phosphate concentration in solution culture: I. Growth and phosphorus content. Soil Sci. 1967, 103, 225–233. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Green, S.; Dangl, J.L. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [Green Version]
- Wongkiew, S.; Park, M.R.; Chandran, K.; Khanal, S.K. Aquaponic systems for sustainable resource recovery: Linking nitrogen transformations to microbial communities. Environ. Sci. Technol. 2018, 52, 12728–12739. [Google Scholar] [CrossRef]
- Schmautz, Z.; Graber, A.; Jaenicke, S.; Goesmann, A.; Junge, R.; Smits, T.H.M. Microbial diversity in different compartments of an aquaponics system. Arch. Microbiol. 2017, 199, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Eck, M.; Sare, A.R.; Massart, S.; Schmautz, Z.; Junge, R.; Smits, T.H.; Mjijakli, M.H. Exploring bacterial communities in aquaponic systems. Water 2019, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W.; He, X. Reduced dependence of rhizosphere microbiome on plant-derived carbon in 32-year long-term inorganic and organic fertilized soils. Soil Biol. Biochem. 2015, 80, 70–78. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lee, J.W.; Chandran, K.; Kim, S.; Brotto, A.C.; Khanal, S.K. Effect of plant species on nitrogen recovery in aquaponics. Bioresour. Technol. 2015, 188, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The ecology of Acidobacteria: Moving beyond genes and genomes. Front. Microbiol. 2016, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello-Yeves, P.J.; Ghai, R.; Mehrshad, M.; Picazo, A.; Camacho, A.; Rodriguez-Valera, F. Reconstruction of diverse verrucomicrobial genomes from metagenome datasets of freshwater reservoirs. Front. Microbiol. 2017, 8, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cardman, Z.; Arnosti, C.; Durbin, A.; Ziervogel, K.; Cox, C.; Steen, A.; Teske, A. Verrucomicrobia are candidates for polysaccharide-degrading bacterioplankton in an Arctic fjord of Svalbard. Appl. Environ. Microbiol. 2014, 80, 3749–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Ding, Y.; Ren, C.; Sun, Z.; Rodionov, D.A.; Zhang, W.; Yang, S.; Yang, C.; Jiang, W. Reconstruction of xylose utilization pathway and regulons in Fermicutes. BMC Genom. 2010, 11, 255–268. [Google Scholar] [CrossRef] [Green Version]
- Kuenen, J.G. Anammox bacteria: From discovery to application. Nat. Rev. Microbiol. 2008, 6, 320–326. [Google Scholar] [CrossRef]
- Li, X.Z.; Rui, J.P.; Mao, Y.J.; Yannarell, A.; Mackie, R. Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol. Biochem. 2014, 68, 392–401. [Google Scholar] [CrossRef]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium genus in the plant holobiont: Ecological, physiological, and applicative insights. In Microbial Models: From Environmental to Industrial Sustainability. Microorganisms for Sustainability; Castro-Sowinski, S., Ed.; Springer: Singapore, 2016; pp. 189–207. [Google Scholar]
- Holmes, D.E.; Dang, Y.; Smith, J.A. Nitrogen cycling during wastewater treatment. Adv. Appl. Microbiol. 2019, 106, 113–192. [Google Scholar] [PubMed]
- Raaijmakers, J.M.; Brunijn, I.; Nybrow, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Bakker, P.A.H.M. Induced systemic resistance as a mechanism of disease suppression by rhizobacteria. In PGPR: Biocontrol and Biofertilization; Siddiqui, Z.A., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 39–66. [Google Scholar]
- Radzki, W.; Manero, F.G.; Algar, E.; García, J.L.; García-Villaraco, A.; Solano, B.R. Bacterial siderophores efficiently provide iron to iron-starved tomato plants in hydroponics culture. Antonie Van Leeuwenhoek 2013, 104, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Bartelme, R.P.; McLellan, S.L.; Newton, R.J. Freshwater recirculating aquaculture system operations drive biofilter bacterial community shifts around a stable nitrifying consortium of ammonia-oxidizing archaea and comammox Nitrospira. Front. Microbiol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Palatinszky, M.; Herbold, C.; Jehmlich, N.; Pogoda, M.; Han, P.; von Bergen, M.; Lagkouvardos, I.; Karst, S.M.; Galushko, A.; Koch, H.; et al. Cyanate as an energy source for nitrifiers. Nature 2015, 524, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Growth Trial 1 | Growth Trial 2 | ||
|---|---|---|---|---|
| Control | Bacillus Treatment | Control | Bacillus Treatment | |
| Temperature (°C) | 24.46 (21.29–25.56) | 24.37 (21.0–25.33) | 26.40 (26.20–26.74) | 26.26 (25.53–26.73) |
| Dissolved oxygen (mg L−1) | 6.0 (5.90–6.10) | 6.1 (5.90–6.20) | 5.5 (5.30–5.90) | 5.6 (5.20–6.0) |
| Total dissolved solids (mg L−1) | 521 (401–584) | 452 (350–541) | 398 (330–471) | 370 (331–446) |
| pH | 7.15 (6.78–7.47) | 7.23 (6.63–7.80) | 6.80 (6.61–7.09) | 6.72 (6.52–6.89) |
| Electrical conductivity (mS cm−1) | 0.69 (0.55–0.84) | 0.61 (0.50–0.71) | 0.57 (0.46– 0.78) | 0.54 (0.46–0.72) |
| Total ammonia nitrogen (mg L−1) | 0.33 (0.10–0.72) | 0.27 (0.13–0.52) | 0.14 (0.10–0.29) | 0.15 (0.04–0.35) |
| Free ammonia (mg L−1) | 0.002 (0.001–0.004) | 0.003 (0.001–0.007) | 0.0006 (0.0003–0.002) | 0.0005 (0.0001–0.002) |
| Nitrite (mg L−1) | <0.1 | <0.1 | <0.1 | <0.1 |
| Initial Weight (g Plant−1) | Initial Height (cm Plant−1) | Final Fresh Shoot Weight (g Plant−1) | Final Root Fresh Weight (g Plant−1) | Final Height (cm Plant−1) | Fv/Fm | CCI | Shoot Dry Weight (g Plant−1) | Root Dry Weight (g Plant−1) | |
|---|---|---|---|---|---|---|---|---|---|
| Growth Trial 1 | |||||||||
| Bacillus | 3.51 ± 0.005 | 13.30 ± 0.10 | 24.84 ± 0.18 | 1.58 ± 0.004 | 34.22 ± 1.56 | 0.83 ± 0.007 | 2.20 ± 0.09 | 0.76 ± 0.03 | 0.104 ± 0.004 |
| Control | 3.50 ± 0.09 | 13.25 ± 0.15 | 20.07 ± 0.02 | 1.06 ± 0.009 | 33.49 ± 0.49 | 0.71 ± 0.01 | 2.06 ± 0.05 | 0.55 ± 0.006 | 0.09 ± 0.005 |
| p-value | 0.29 | 0.80 | 0.001 * | 0.07 | 0.69 | 0.01 * | 0.30 | 0.02 * | 0.07 |
| Growth Trial 2 | |||||||||
| Bacillus | 4.62 ± 0.05 | 12.35 ± 0.30 | 33.08 ± 1.36 | 2.11 ± 0.04 | 41.68 ± 1.98 | 0.82 ± 0.04 | 2.46 ± 0.03 | 1.47 ± 0.031 | 0.24 ± 0.01 |
| Control | 4.63 ± 0.01 | 12.63 ± 0.15 | 25.57 ± 0.53 | 1.65 ± 0.05 | 32.94 ± 0.29 | 0.72 ± 0.01 | 2.41 ± 0.004 | 0.85 ± 0.01 | 0.14 ± 0.01 |
| p-value | 0.70 | 0.49 | 0.03 * | 0.02 * | 0.01 * | 0.03 * | 0.30 | 0.001 * | 0.02 * |
| Variables | Control | Bacillus | p-Value |
|---|---|---|---|
| Calcium (mg L−1) | 27.90 ± 1.50 | 27.58 ± 3.35 | 0.94 |
| Copper (mg L−1) | 0.15 ± 0.0005 | 0.14 ± 0.03 | 0.71 |
| Iron (mg L−1) | 0.14 ± 0.002 | 0.14 ± 0.002 | 0.80 |
| Potassium (mg L−1) | 18.48 ± 0.82 | 20.31 ± 0.01 | 0.004 * |
| Magnesium (mg L−1) | 16.15 ± 0.61 | 15.17 ± 0.01 | 0.25 |
| Sodium (mg L−1) | 116.8 ± 1.60 | 98.6 ± 1.35 | 0.01 * |
| Phosphorus (mg L−1) | 1.13 ± 0.13 | 4.26 ± 0.26 | 0.008 * |
| Zinc (mg L−1) | 0.03 ± 0.002 | 0.08 ± 0.001 | 0.002 * |
| Mineral Composition | Control | Bacillus | p-Value |
|---|---|---|---|
| Calcium (g kg−1) | 51.40 ± 1.50 | 50.65 ± 0.69 | 0.69 |
| Potassium (g kg−1) | 50.95 ± 0.20 | 54.87 ± 0.73 | 0.03 * |
| Magnesium (g kg−1) | 29.86 ± 0.66 | 29.38 ± 0.26 | 0.57 |
| Sodium (g kg−1) | 25.20 ± 0.44 | 21.21 ± 1.09 | 0.07 |
| Phosphorus (g kg−1) | 4.83 ± 0.22 | 7.95 ± 0.12 | 0.007 * |
| Copper (mg kg−1) | 4.96 ± 0.09 | 4.04 ± 0.51 | 0.22 |
| Iron (mg kg−1) | 48.36 ± 1.81 | 47.99 ± 1.22 | 0.88 |
| Zinc (mg kg−1) | 6.25 ± 0.39 | 14.75 ± 1.39 | 0.02 * |
| Treatment | Number of Raw Sequences | Number of Sequences before Chimeras | Number of Sequences after Chimeras | Chloroplast and Mitochondria Reads | Number of Reads after Screening and Filtering |
|---|---|---|---|---|---|
| Bacillus | |||||
| Sample B1 | 56,285 | 47,935 | 40,859 | 246 | 34,348 |
| Sample B2 | 20,466 | 17,971 | 15,684 | 102 | 12,696 |
| Control | |||||
| Sample C1 | 42,899 | 37,663 | 33,101 | 1547 | 28,145 |
| Sample C2 | 59,840 | 53,351 | 46,178 | 2366 | 39,980 |
| Phyla | Control | Bacillus | p-Value |
|---|---|---|---|
| Acidobacteria | 0.76 ± 0.08 | 2.21 ± 0.12 | 0.009 * |
| Actinobacteria | 3.64 ± 0.25 | 4.05 ± 0.02 | 0.246 |
| Armatimonadetes | 0.28 ± 0.01 | 0.24 ± 0.00 | 0.095 |
| Bacteria_unclassified | 0.38 ± 0.05 | 0.35 ± 0.00 | 0.542 |
| Bacteroidetes | 17.74 ± 0.74 | 17.35 ± 0.13 | 0.657 |
| Chlamydiae | 0.24 ± 0.008 | 0.42 ± 0.05 | 0.072 |
| Chloroflexi | 3.93 ± 0.09 | 2.23 ± 0.09 | 0.006 * |
| Cyanobacteria | 2.75 ± 0.11 | 0.18 ± 0.001 | 0.002 * |
| Deinococcus-Thermus | 0.27 ± 0.04 | 0.11 ± 0.001 | 0.060 |
| Dependentiae | 0.23 ± 0.02 | 0.42 ± 0.07 | 0.121 |
| FBP | 0.086 ± 0.00 | 0.89 ± 0.00 | <0.001 * |
| Firmicutes | 0.53 ± 0.06 | 3.24 ± 0.07 | 0.001 * |
| Gemmatimonadetes | 0.10 ± 0.006 | 0.37 ± 0.03 | 0.009 * |
| Nitrospirae | 0.09 ± 0.025 | 0.37 ± 0.03 | 0.022 * |
| Patescibacteria | 1.34 ± 0.08 | 0.36 ± 0.04 | 0.008 * |
| Planctomycetes | 3.75 ± 0.35 | 7.49 ± 0.24 | 0.012 * |
| Proteobacteria | 60.44 ± 1.96 | 55.35 ± 0.38 | 0.125 |
| Verrucomicrobia | 2.89 ± 0.23 | 3.89 ± 0.03 | 0.048 * |
| WPS-2 | 0.39 ± 0.08 | 0.16 ± 0.03 | 0.115 |
| Genera | Control | Bacillus | p-Value |
|---|---|---|---|
| Acinetobacter | 1.78 ± 0.03 | 1.42 ± 0.03 | 0.012 * |
| Arenimonas | 11.29 ± 0.58 | 8.67 ± 0.07 | 0.046 * |
| Bacillus | 0.24 ± 0.02 | 2.90 ± 0.08 | <0.001 * |
| Brevifollis | 0.02 ± 0.009 | 0.11 ± 0.009 | 0.018 * |
| Chryseobacterium | 0.08 ± 0.007 | 2.38 ± 0.007 | <0.001 * |
| Chthoniobacter | 0.11 ± 0.009 | 0.39 ± 0.03 | 0.013 * |
| Cloacibacterium | 0.00 ± 0.0006 | 0.49 ± 0.02 | 0.002 * |
| Defluviimonas | 0.02 ± 0.004 | 0.12 ± 0.0001 | 0.002 * |
| Dinghuibacter | 0.04 ± 0.002 | 0.25 ± 0.01 | 0.005 * |
| Dysgonomonas | 0.60 ± 0.09 | 0.28 ± 0.00003 | 0.083 |
| Emticicia | 0.53 ± 0.009 | 0.19 ± 0.03 | 0.007 * |
| Ferruginibacter | 1.76 ± 0.15 | 2.31 ± 0.04 | 0.072 |
| Fimbriiglobus | 0.20 ± 0.009 | 0.50 ± 0.03 | 0.009 * |
| Flavihumibacter | 0.04 ± 0.007 | 0.33 ± 0.01 | 0.002 * |
| Flavobacterium | 2.78 ± 0.12 | 1.67 ± 0.008 | 0.011 * |
| Fodinicola | 0.28 ± 0.02 | 0.91 ± 0.06 | 0.010 * |
| Gemmata | 0.22 ± 0.02 | 0.49 ± 0.035 | 0.023 * |
| Haloferula | 0.18 ± 0.02 | 0.08 ± 0.03 | 0.144 |
| Herbaspirillum | 0.11 ± 0.03 | 0.15 ± 0.007 | 0.257 |
| Herpetosiphon | 2.34 ± 0.04 | 0.17 ± 0.02 | <0.001 * |
| Hydrogenophaga | 0.47 ± 0.02 | 0.24 ± 0.03 | 0.026 * |
| Hyphomicrobium | 0.31 ± 0.0002 | 0.37 ± 0.05 | 0.337 |
| Ideonella | 0.23 ± 0.009 | 0.11 ± 0.01 | 0.015 * |
| Larkinella | 0.02 ± 0.004 | 0.23 ± 0.005 | 0.001 * |
| Legionella | 0.42 ± 0.02 | 0.30 ± 0.03 | 0.059 |
| Luteolibacter | 1.18 ± 0.09 | 1.62 ± 0.05 | 0.056 |
| Mycobacterium | 0.37 ± 0.03 | 0.65 ± 0.005 | 0.012 * |
| Nakamurella | 0.03 ± 0.003 | 0.29 ± 0.03 | 0.015 * |
| Niastella | 0.02 ± 0.009 | 0.22 ± 0.02 | 0.012 * |
| Nitrospira | 0.09 ± 0.03 | 0.37 ± 0.03 | 0.023 * |
| Nocardia | 0.06 ± 0.009 | 0.35 ± 0.01 | 0.002 * |
| Pirellula | 0.55 ± 0.04 | 0.44 ± 0.02 | 0.127 |
| Polynucleobacter | 0.05 ± 0.01 | 1.08 ± 0.06 | 0.003 * |
| Pseudomonas | 0.15 ± 0.01 | 0.08 ± 0.02 | 0.089 |
| Pseudoxanthomonas | 0.35 ± 0.03 | 0.20 ± 0.02 | 0.050 |
| Reyranella | 0.28 ± 0.04 | 0.42 ± 0.03 | 0.090 |
| Runella | 0.09 ± 0.009 | 0.24 ± 0.01 | 0.010 * |
| Sphingomonas | 0.10 ± 0.0004 | 0.14 ±0.003 | 0.006 * |
| Terrimonas | 1.61 ± 0.08 | 1.32 ± 0.02 | 0.062 |
| Thermomonas | 1.72 ± 0.10 | 7.70 ± 0.33 | 0.003 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasozi, N.; Kaiser, H.; Wilhelmi, B. Effect of Bacillus spp. on Lettuce Growth and Root Associated Bacterial Community in a Small-Scale Aquaponics System. Agronomy 2021, 11, 947. https://doi.org/10.3390/agronomy11050947
Kasozi N, Kaiser H, Wilhelmi B. Effect of Bacillus spp. on Lettuce Growth and Root Associated Bacterial Community in a Small-Scale Aquaponics System. Agronomy. 2021; 11(5):947. https://doi.org/10.3390/agronomy11050947
Chicago/Turabian StyleKasozi, Nasser, Horst Kaiser, and Brendan Wilhelmi. 2021. "Effect of Bacillus spp. on Lettuce Growth and Root Associated Bacterial Community in a Small-Scale Aquaponics System" Agronomy 11, no. 5: 947. https://doi.org/10.3390/agronomy11050947