1. Introduction
Poland contributes 8.3% of the total onion production in the EU and Polish exports of this vegetable constitute 2.5% of global exports, putting Poland in 9th position in the world [
1]. In 2019, onions were cultivated in Poland on an area of 25,200 ha with an average yield of 21.2 t/ha [
2]. Onion thrips,
Thrips tabaci Lindeman 1889 is a global pest [
3,
4] and also represents a serious threat to onion production in Poland [
5]. Larvae and adults of onion thrips live in the leaf sheath and chives, and their feeding causes silvery leaf spots that turn into white blotches along the leaves due to the removal of cellular content. This injury reduces photosynthetic ability and interferes with the transportation of nutrients to the bulb [
6]. The negative effects of onion thrips are reflected in both the quantity and quality of the crop [
7]. Their feeding causes the reduction of onion bulb mass and also has an indirect effect by creating an entry point for plant pathogens [
8,
9].
T. tabaci is also a vector of Iris yellow spot virus (IYSV) [
10]. Effective control of onion thrips necessitates frequent applications of insecticide [
11], with the unfortunate result of increased resistance to commonly used chemicals [
12,
13].
The demand for high-quality onions, produced to the highest standards, is growing continually. For this reason, an increasing number of onion growers are choosing more environmentally friendly methods of cultivation, which allow the use of fewer pesticides. The choice of a cultivar characterized by a higher degree of resistance to the important pest species for the particular crop is one of the most important agrotechnical measures and an element of integrated pest management (IPM).
Plant characteristics make herbivores prefer one cultivar to another. Some of the plant characteristics are perceivable before landing, such as the size, shape, scent, colour, and light reflectance of the host, while others are perceivable only after landing, such as the physical and chemical parameters of the epidermis or chemical constitution of plant sap [
14,
15,
16]. Reeves [
17] pointed out that despite a general trend of studies and reviews ignoring or downplaying the importance of insect vision in locating host plants in favour of chemical cues, perception of colour and shape can be as important, or in some cases more important, than chemical cues [
18,
19]. Studies of visual plant characteristics have largely focused on the plant colour, brightness (intensity of perceived reflected light), the polarization of foliar reflectance, saturation (hue clearness), and shape [
17,
20,
21].
Like other insects, thrips use colour, shape, size, and volatiles to locate the host plant. Colour and colour contrast are used by some thrips species to distinguish between a host and the surrounding environment [
21]. To determine which colour is most attractive to onion thrips, researchers have tested different colour sticky traps. Behavioural studies of the colour preference of
T. tabaci have provided variable results, but generally agree that greater numbers of thrips are caught by low UV-reflective white, blue, yellow, and fluorescent yellow traps than are caught by green, red, black, and high UV-reflective white traps [
22,
23,
24,
25,
26,
27]. According to Demirel and Yeldirim [
28], green is regarded as one of the least attractive colours for
T. tabaci; however, few studies have looked at differences in preference based on a green hue [
22,
29]. Onion thrips show a significant preference for both light green and mid-green over dark green [
29]. Studies on the spectral sensitivity of
T. tabaci photoreceptors suggest that its vision covers the general insect-visible spectrum between 350 and 650 nm [
30]. The last study of Rőth et al. [
27] indicated that onion thrips possess in their eyes at least two different light receptors: one with peak spectral sensitivity in the greenish-yellow region (540–570 nm) and another in the UV-A region (350–360 nm). Egri et al. [
31] established that the spectral sensitivity of the leek-associated biotype L2 of the onion thrips’ compound eye has only one striking green peak at 521 nm with only a small shoulder in the UV-A range, while the attractiveness of 350 nm light is much stronger than that of 525 nm for biotype L1 [
32].
A mechanism that includes the morphology of a physical plant characteristic by which an insect’s behaviour is disturbed, for example, feeding, mating, and oviposition, is known as antixenosis [
33]. Antixenosis is an important component of resistance because it reduces the initial infestation level; however, in monoculture, this mechanism may be broken down in the absence of the preferred host plant. In this case, pests may eventually accept a less favoured host [
14,
15]. Leaf colour has been advocated as an influential factor in determining thrips resistance in onion [
34,
35,
36,
37]. In recent years, Diaz-Montano [
10,
38], Fail et al. [
39], and Balint et al. [
40] have confirmed that antixenosis plays a role in the resistance of onion and white cabbage to onion thrips and also documented that leaf colour and leaf reflectance might positively or negatively influence colonization by
T. tabaci. The results of Rőth et al. [
27] suggest that light reflectance in the yellow region and the UV range has the most important effect on the selection of a host plant by
T. tabaci.
Varietal preference and the susceptibility of onion to
T. tabaci have been documented in Poland by Pobożniak et al. [
41], but no traits were investigated to explain this preference. In this paper, we report the result of measuring the susceptibility of existing commercial onion cultivars to
T. tabaci infestation concerning leaf colour characteristics. The detailed objectives of the study were to identify cultivars with a high level of nonpreference (antixenosis) that can be cultivated by farmers as a tool of IPM and can also be used by plant breeders as a source of resistance to onion thrips in plant-breeding programmes.
4. Discussion
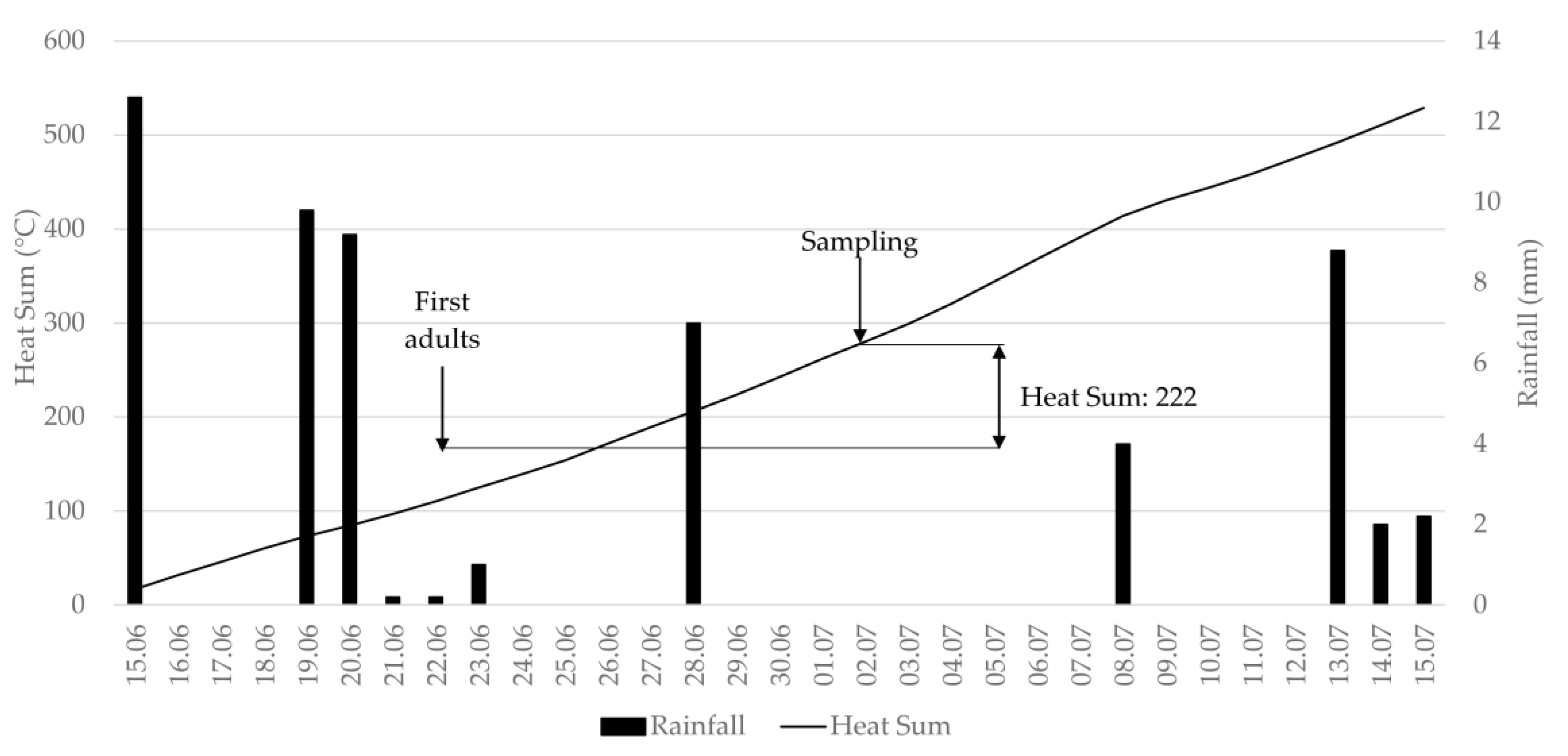
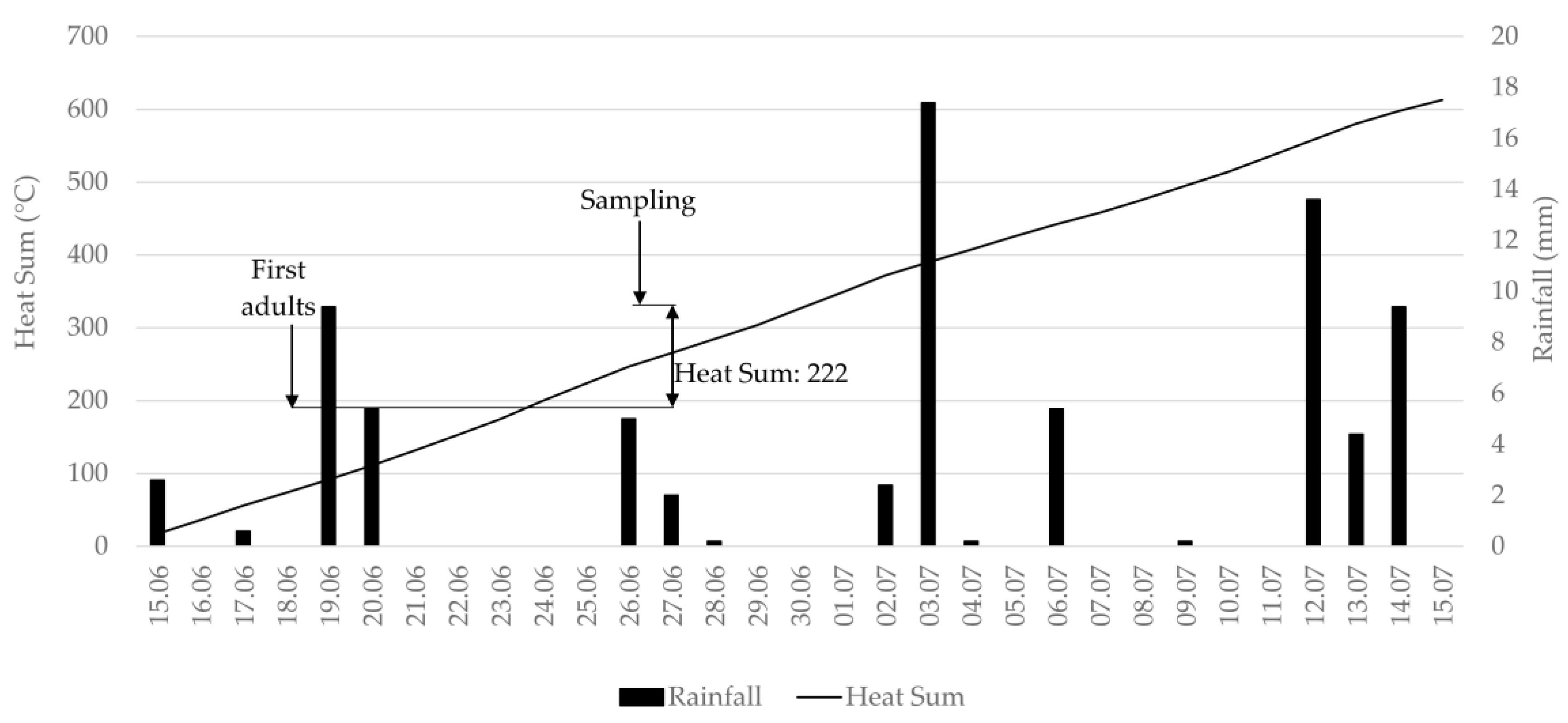
The measurement of onion thrips population size under field conditions is used by entomologists as a tool for the first stage of selecting resistant plant material. Antixenosis testing is essentially based on measuring the attractiveness of a plant genotype to colonizing adult thrips. Fail et al. [
51] proposed using the proportional abundance of thrips adults in addition to the actual count of thrips as an appropriate measure of antixenosis because it seems to be more stable under varying field conditions. Proportional abundance is also more in line with the concept that resistance of plants is relative and is based on comparison with plants lacking the resistance characters, i.e., susceptible plants [
52]. According to Fail et al. [
51], the heat sum model predicts the development of
T. tabaci well, and therefore all thrips adults that we encountered during the sampling events in both years can be considered as colonizing adults. The term antixenosis cannot be used at a later time when newly emerged individuals appear on onions.
In two consecutive years, 2015 and 2016, a different order of the level of susceptibility of onion cultivars to T. tabaci was established. In 2015, the cultivar Wenta followed by Alibaba showed a low level of susceptibility to colonization by onion thrips, which was reflected in both low actual count and proportional abundance of thrips collected directly from plants and with a sweep net. In 2016, the cultivars Tęcza and Bila were the least susceptible. The proportional abundance of onion thrips collected directly from Tęcza in 2015 and collected from Wenta with a sweep net in 2016 also indicated a low level of attractiveness of these cultivars to onion thrips. In contrast, the cultivars most susceptible to onion thrips infestation were Niagara F1 and Bila in 2015, and Alibaba and Kristine in 2016.
The observed preference among leaves of different cultivars could have been the result of thrips choosing landing sites among the leaves of the available plants. The decision on the suitability of a plant as a host is made in the very first phase of host selection, with colonizers using both visual and chemical cues [
15]. Thrips might show a preference for a specific odour, hue, or intensity of colour in their preferred plant [
10,
53]. The selection process can be disrupted under field conditions. Thrips are relatively weak flyers, being able to determine their speed and direction only at low wind speeds [
54,
55]. As a consequence, only a very small proportion of thrips locate suitable hosts [
54]. The other reason for the lower abundance of thrips on some cultivars compared to those more colonized could be the result of more rapid take-off after landing on the leaves of a resistant variety [
51]. In our study, the low number of onion thrips on Tęcza and Wenta in two consecutive seasons may indicate host plant selection and nonpreference mechanism of resistance (antixenosis). Nonetheless, to confirm this mechanism of resistance to plant colonization by
T. tabaci, a choice test should be performed under controlled conditions.
We found distinct differences in some computed colour variables between the leaves of the onion cultivars most and least susceptible to
T. tabaci infestation, but because of the discrepancy in the results between the two years of the study, the role of visual cues for thrips must be interpreted with caution. By analysing the results for the eight onion cultivars, we have ascertained that leaf colour may influence colonization by thrips. In both years, the cultivars with the highest value for lightness (Bila and Niagara F
1 in 2015, and Alibaba and Niagara F
1 in 2016) were very attractive to
T. tabaci. In contrast, the resistant cultivars, Wenta in 2015 and Tęcza in 2016, were darker and had low L* values. A negative coordinate a* value indicates a green colour; for susceptible cultivars, namely Bila in 2015, and Alibaba and Kristine in 2016, its value decreased, so the tonality of the colour shifted to greener. In turn, the increasing positive value of coordinate b* (yellowness) indicated that the leaf colour of susceptible cultivars was oriented towards yellow. In contrast, the increasing value of a* in resistant cultivars, Wenta in 2015 and Tęcza in 2016, and lower value of b* indicated a less intense green colour oriented towards yellow-grey. The lower value of C* in these resistant cultivars means that their colour is less saturated in comparison with susceptible ones with a high chroma. Estimation of the hue difference (ΔH*
ab) and hue angle h* indicated that the colour of the cultivars most colonized by onion thrips had a more yellowish hue while that of the least infested plants was more grey. Colorimetric values of leaf colour were correlated with attractiveness to thrips: typically, higher b*, C*, and h* values and lower a* attracted more thrips; therefore, we concluded that the vivid, intense green-yellowish leaf colour of susceptible varieties might have been the cause of the observed thrips preference. The peak sensitivity of
T. tabaci photoreceptors at 540–570 nm [
27] could perhaps partially explain the preference for the light green-yellowish colour. In contrast to our results, Diaz-Montano et al. [
10] found that onion varieties resistant to
T. tabaci such Tioga, OLYSO5N5, and Peso had visually determined yellow-green leaves unlike the bluish-green foliage of susceptible cultivars SYN-G2 and Santana. These observations were usually accompanied by high b* values measured in the resistant cultivars in comparison with the susceptible ones, but some resistant varieties had a b* value very similar to that of the susceptible ones. Fail et al. [
39] found significant differences in the CIE 1976 a* and b* values between leaves forming the head of resistant and susceptible varieties of cabbage. The moderately resistant variety Blokotor had the highest negative value of coordinate a* and the lowest value of coordinate b* in comparison with two other resistant (yellowish-green) as well as three susceptible cultivars (yellowish). In other studies, Alimousavi et al. [
56], Birithia et al. [
57], and Yousefi et al. [
58] showed that onion genotypes with glossy foliage and a light green colour had lower thrips infestation in comparison with nonglossy and medium or dark green susceptible genotypes. Pobożniak [
59] found that pea cultivars with grassy green leaves were the most frequently infested by
T. tabaci. At the same time, cultivars with yellow-green leaves or dark blue-green leaves were less attractive to thrips.
Behavioural variability between studies may reflect the geographical distribution and genetic differences of onion thrips [
31,
32]. It may also relate to differences in experimental designs. In contrast to our research, the authors of the above-mentioned studies included thrips collected throughout the entire growing season, i.e., also individuals already reproducing on onion leaves; their presence on plants cannot be considered as the result of thrips choice and could be affected by antibiosis.
Our results are concordant with those of the study carried out by Westmore et al. [
29], who reported that potato cultivars preferred by adult onion thrips during colonization for foraging usually had lighter green foliage and higher spectral reflectance in the green wavelength (552 nm). In contrast, more eggs were laid on potato leaves with darker green foliage [
29]. Understanding the factors underlying the choice of both host and oviposition is important for guiding breeding programmes for new cultivars.
The environmental conditions that a plant is exposed to, including soil properties, weather, and its phenological age, alter the optical properties of its leaves [
60]. It is highly probable that environmental stress might have affected the leaf colour of onion cultivars, and therefore colorimetric values of some onion cultivars were different in the two years. Therefore, the manager of the onion-breeding programme should plant the same check varieties each year to determine how the colour parameters may have shifted. In our experiment, the very high attractiveness of Bila for thrips infestation in 2015 might have been related to the higher lightness and higher values of b*, C*, and h* and lower value of a* for Bila compared to 2016.
Although it is difficult to ascertain which colour characteristic contributed most to the differences in susceptibility of onion leaves to onion thrips; we suspect that lightness may be one of the key factors associated with varietal attractiveness. In both years, the cultivar most attractive for thrips infestation, Niagara F1, had a very high lightness value, while the a* and b* values were similar to those of the moderately resistant varieties. In addition, the high correlation (r > 0.6) between the actual count and proportional abundance of T. tabaci, and the lightness values measured for the onion leaves in 2015 might indicate a high preference of T. tabaci for cultivars with lighter leaves.
Reflectance values within a spectral range have been correlated with thrips attraction, usually with higher reflectance attracting higher numbers of thrips. Diaz-Montano et al. [
38] found that
T. tabaci abundance and the brightness of onion leaves were significantly correlated, especially in the UV range of light (275–375 nm and 310–410 nm). The same authors created the hypothesis that
T. tabaci prefer onion cultivars that reflect a greater amount of light; it is possible that this characteristic may shelter onion thrips from heat and may make these onion cultivars a more preferable host. In addition, Fail et al. [
51] found a positive correlation between the actual count of thrips adults and the brightness of old cabbage leaves. Besides the average brightness (between 270 and 650 nm), the reflectance in the range of sensitivity of the first (275–375 nm), second (275–400 nm), and third (400–650 nm) theoretical photoreceptor systems of
T. tabaci adopted by the authors was almost equally correlated with thrips abundance.
Brightness is related to reflectance, while lightness (L*) is more related to the colour impression. A colour surface with a higher L* value usually has a higher reflectance [
61]. Reflectance is a physical measure that depends on the incidence angle, the polarization of the radiation, and the refraction index of the surface [
62]. The reflection of light from the leaf and transmission through the leaf are determined by the wavelengths of light absorbed by the various biochemical compounds in leaves (chlorophylls, carotenoids, water, cellulose and lignin, proteins, etc.) and also depend on the epidermis, waxes, cutin, and protrusions such as leaf hairs [
63,
64]. Thus, for leaves with the greatest concentrations of chlorophylls, carotenoids, water, cellulose and lignin, proteins, etc., reflectance is the smallest at a certain wavelength. Lighter leaves usually reflect much of the visible light [
64]. In our results, cultivars with lighter, green-yellowish leaves attracted thrips the most, which could be due to their higher reflectance; however, more evidence supporting this and additional work are needed to understand the relationship between the optical properties of leaves and the responses of onion thrips to different colour attributes.
It should be noted that colour is only one component that contributes to the appearance of onion plants. Shape, leaf angle, plant height, and insect or disease damage also contribute to the appearance of a plant [
56,
65,
66]. In turn, the micromorphology of the leaf epidermis, internal leaf structure, and chemical properties might affect thrips post lighting behaviour [
67,
68].






{kind=link}
{kind=link}
{kind=link}
{kind=link}