Abstract
Natural plant resources with herbicidal activity may be substitutes for synthetic chemical pesticides, likewise aromatic plant extracts, especially essential oils. Essential oil from Litsea pungens has been proved to possess a strong antibacterial property. Interestingly, we found the essential oil also showed a strong allelopathic capacity. Therefore, in the present work, the chemical composition of the essential oil from the fruit of L. pungens was analyzed by gas chromatography–mass spectrometry (GC–MS). The weed control abilities of the essential oil were also further evaluated. The results show that the yield of essential oil extracted by steam distillation was 1.4%, and 17 compounds, mainly terpenoids, were identified by GC–MS. In allelopathic tests, the essential oil exhibited a negative effect on seed germination rate and seedling growth of Lolium perenne and Bidens pilosa. Moreover, chlorophyll content, malondialdehyde content, electrolyte leakage, catalase, superoxide dismutase and peroxidase of seedlings treated with essential oil were also negatively affected. This work could provide a better understanding of the rational utilization of L. pungens essential oil for crop cultivation and further development of environment-friendly herbicides.
1. Introduction
In recent years, the excessive abuse of chemical synthetic herbicides has had a negative impact on environmental pollution and human health, and has led to increasing herbicidal resistance in many weeds [1,2,3]. In agriculture, weeds constantly compete with crops for water, light and nutrients, which is one of the main causes of crop yield decline. Allelopathic weed control is one of the alternative strategies for weed management in agro-ecosystems, which can reduce the use of traditional herbicides [4]. Compared with traditional herbicides, aromatic plant essential oils have more ecological advantages in the environment and can be used as biological herbicides [5,6,7]. It is found that essential oils such as clove oil and eucalyptus oil are a promising natural resource that can be used as insect repellents, herbicides, food protectors and for environmental protection as non-toxic chemical pesticide alternatives [8,9,10]. Aghbash reported that essential oils contain bioactive compounds, such as monoterpenes and sesquiterpenoids [11], which belong to the allelochemicals, and are involved in many metabolic and ecological processes [12]. Concomitantly, many reports are interested in the development and utilization of plant essential oils as environment-friendly herbicides [5,13,14].
Litsea pungens is a species belonging to the Litsea genus and family Lauraceae, predominantly distributed in Hubei, Hunan, Guangxi, Sichuan, Guizhou, Yunnan in China. It is a pioneer herb traditionally utilized in medicine [15]. It is reported that L. pungens essential oil is a safe and environment-friendly raw material with good bactericidal activity. L. pungens fruit contains essential oil, which can be used as food flavor and cosmetics flavor, and has been widely used as raw material of advanced spices, Vitamin A, as well as violet ketone in industry [16]. So far, many scholars have conducted extensive research on the composition of this L. pungens essential oil, mainly focusing on antibacterial, antioxidant, insect inhibition, and cancer inhibition [17,18,19], while research on inhibiting the growth of weeds and the development and utilization of new herbicidal active substances are very limited.
Therefore, growth parameters as well as physiological and biochemical indicators of weed seeds treated with L. pungens essential oil by petri dishes and pot experiments were measured. To the best of our knowledge, this is the first systematic report about the potential allelopathic effect of essential oil from L. pungens.
2. Materials and Methods
2.1. Plant Materials
The fruits of L. pungens were gathered from Hongya county (29°56′75.28″ N; 103°24′33.55″ E), Sichuan, in 2019. Fresh fruit dries naturally to get dried fruit. Seeds of Lolium perenne (L. perenne) and Bidens pilosa (B. pilosa) were purchased from local seed stores.
Ethanol, Tween 80, ferrous sulfate and sodium sulfate were purchased from Chengdu Kelong Chemical Factory (Chengdu, China). All chemicals were analytical grade and used without further purification.
2.2. Isolation of the Essential Oil
The dried fruits of L. pungens were ground into fine powder and extracted by steam distillation for 6 h, respectively. After standing and layering, anhydrous sodium sulfate was added to remove water, then the essential oil was obtained and stored in a brown bottle at 4 °C until analyzed by gas chromatography–mass spectrometry and bioassays. According to the following formula to calculate the extraction yield of essential oil:
Oil yield (%, v/w) = volume of essential oil (mL)/dried weight of plant material (g) × 100%
2.3. Gas Chromatography–Mass Spectrometry Analysis
The volatile oil was analyzed qualitatively and semi-quantitatively by GC–MS. Typically, 1 µL essential oil was resolved in chromatography n-hexane (ratio of 1/10) and n-hexane was used as blank control. Gas chromatographic conditions: HP-5MS column (30 cm × 0.25 mm × 0.25 μm), using a temperature program. This was held at 60 °C for 5 min and then raised to 100 °C at the speed of 12 °C/min keeping 5 min. The temperature was raised to 160 °C at the speed of 10 °C/min and kept for 5 min. Subsequently, the temperature was increased to 280 °C at the speed of 8 °C/min and kept for 5 min. Helium was invoked as the carrier gas and the injection volume was 1 μL with no shunt. The injection port temperature was 260 °C, and the interface temperature was 220 °C. Mass spectrometry conditions: electron bombardment (EI) ion source. Electron energy was 70 eV and the electron multiplier voltage was 1.5 kV. Mass scanning ranged from 40 to 650 (m/z) full scanning [20]. In order to make the experimental spectrum closer to the library spectrum, the standard spectral modulation was used.
The essential oil qualitative identification was based on a comparison of mass spectrometry with standards recorded by the National Institute of Standards and Technology (NIST) libraries. The properties of the compounds were verified by mass spectrometry and linear retention index compared with literature data [21]. The relative proportion of compounds was determined by area standardization method and expressed by area %.
2.4. Allelopathic Activity Assays
2.4.1. Seed Germination Experiment (Petri Dish Experiment)
The allelopathic effect of essential oil was determined by a Petri dish method with slight modifications [22]. Briefly, essential oil was decomposed in Tween 80 with different concentrations. Then essential oil solution was added into a petri dish to dissolve essential oil with different concentrations (0.125–2.000) mg/mL in Tween 80 (The final concentration was 1%). The seeds were disinfected by NaClO (0.5%) for 10 min and rinsed multiple times with distilled water. Distilled water and 1% Tween 80 were used as blank control and negative control, respectively. Two pieces of sterilized filter papers were placed in a sterilized petri dish with a diameter of 9 cm. 5 mL essential oil solution was added in the test group, and 5 mL distilled water and 5 mL 1% Tween 80 solution were added in the control group. After the filter paper was soaked, 30 seeds of L. perenne and B. pilosa were evenly spread on the filter paper, then the petri dish was covered immediately, and placed in an artificial climate chamber of (25 ± 2) °C for culture. To ensure the wetness of the filter paper, 2 mL of the test group solution and the control group solution were added to the Petri dish every 2 days after 3 days of culturing. The standard for seed germination is that the radicle or hypocotyl can break through the seed coat by 1–2 mm. The number of plant seeds germinating was observed and recorded every day for 7 consecutive days (until no more germination for 3 consecutive days). The shoot length and root length were measured with a digital display Vernier caliper on the 7th day. Each treatment was repeated 6 times.
The final germination rate, germination force, and germination index were calculated according to the following formula:
where, A = the total number of germinated seeds, A0 = the total number of tested seeds.
where, B = the number of seeds germinated at the peak of germination, A0 = the total number of tested seeds.
where, Gt = the number of germinations on the t day, Dt = the number of corresponding germination days.
Germination rate = (A/A0) × 100%
Germination force = B/A0
Germination index = ∑(Gt/Dt)
2.4.2. Pot Experiment
The further allelopathic effect of essential oil was assessed with pot assay according to the previous experiment with slight modifications [23]. In a small pot (height 7.5 cm and width 6.0 cm), 185 g of sterilized weed-free seed soil was filled, and 30 sterilized seeds were evenly sown in a pot; 10 mL of essential oil and control solutions were applied to water the pots for 2 weeks, respectively. After the emergence of the seedlings, a sprayer was used to spray 5 mL of essential oil and control solutions on each pot for one week, respectively. This was repeated for each treatment group six times. After spraying for a week, plant-related growth parameters and biochemical indicators were determined.
2.4.3. Determination of Photosynthetic Pigments
The content of photosynthetic pigments was determined according to the previous method with slight modifications [24]. We placed 0.10 g leaves into a mortar, and 2 mL of 95% ethanol was added to grind thoroughly with some appropriate amount of quartz sand and calcium carbonate. The mixture was then transferred to a 2 mL centrifuge tube and centrifuged at the speed of 10,000 rpm for 1 min. The supernatant was mixed with 95% ethanol to 50 mL and then the absorbance of the solution was determined at three wavelengths of 663 nm, 645 nm, and 440 nm, respectively:
where, V = the extract volume, Fw = the sample quality.
The content of chlorophyll a (mg/g FW) = (12.72A663 − 2.59A645) × V/Fw × 1000
The content of chlorophyll b (mg/g FW) = (22.88A645 − 4.67A663) × V/Fw × 1000
The content of total chlorophyll (mg/g FW) = (Ca + Cb)
The content of carotenoids (mg/g FW) = (4.7A440 − 0.27(Ca + Cb)) × V/Fw × 1000
2.4.4. Determination of Antioxidant Enzyme Activity
The activities of enzymes were determined according to the previous method with some modifications [25]. 0.1 g of potted leaves were homogenized in 2 mL phosphate buffer (50 mM and pH = 7.8) and some quartz sand. The mixture was centrifuged at 12,000 rpm at 4 °C for 15 min, and the supernatant was obtained for determining the activity of CAT, POD, and SOD.
We mixed 1.95 mL of phosphate buffer (50 mM and pH = 7), 1 mL of H2O2 (0.2%), and 50 μL of supernatant for 40 s and the absorbance of the reaction solution was immediately determined at 240 nm. The activity of CAT was determined according to the following formula:
Activity of CAT (U/g Fw) = (A240 × Vt)/(W × Vs × 0.01 × t)
We added 2 mL of H2O2 (0.2%) and 0.95 mL of guaiacol to 10 µL of supernatant solution for 40 s. Then the absorbance of the reaction solution was determined at 470 nm to calculate the activity of POD.
Activity of POD (U/g Fw) = (A470 × Vt)/(W × Vs × 0.01 × t)
We mixed 150 µL of riboflavin (0.3 µmol/L) with 2.5 mL of methionine (13 mmol/L) and 250 µL of nitronitroblue tetrazolium chloride. Then 50 µL supernatant was added and the solution was placed in the light incubator for 20 min reacting under 4000 lux light. The non-illuminated control tube contained only buffer solution, placed in the dark as the zero adjustment. The absorbance of each reaction solution was determined at 560 nm to evaluate the activity of SOD.
Activity of SOD (U/g Fw) = ((ACK − AE) × V)/(1/2ACK × W × Vt)
2.4.5. Determination of the Content of Malondialdehyde (MDA) and Electrolyte Leakage (EL)
We ground 0.1 g leaves with 5 mL 5% triclosan abrasion in the presence of quartz sand. Then the extract solution was centrifuged at the speed of 3000 rpm for 10 min. 2 mL supernatant was mixed with 2 mL thiobarbituric acid (0.67%, w/v), then placed at 100 °C for 30 min. The absorbance of the reaction solution was measured at the wavelengths of 600 nm, 532 nm, and 450 nm, respectively.
Content of MDA (mmol/g Fw) = (6.452 × (A532 − A600) − 0.559 × A450) × Vt/Vs × W
We placed 0.1 g leaves in a graduated test tube of 10 mL distilled water, covered with a glass stopper and soaked in a dark room temperature for 24 h. The conductivity (R1) was firstly measured with a conductivity meter, then the tube was heated in a boiling water bath for 30 min. After cooling to room temperature and shaking, the conductivity (R2) was determined again. The EL was calculated according to the following formula:
EL = (R1/R2) × 100%
2.5. Statistical Analysis
All the experimental groups were conducted in triplicates. The outcomes were expressed as mean ± standard deviation (SD). Graphpad Prism 8.0 was used for statistical analysis and making graphic pictures. The significance test was analyzed by SPSS 19.0 using the analysis of variance (ANOVA) method and the significance level was considered as p < 0.05.
3. Results
3.1. Chemical Characterization Analysis of L. Pungens Essential Oil
The yield of L. pungens essential oil extracted by steam distillation was 1.4% (v/w), the density was 0.9 g/mL, which was based on dried weight, and the constituents of L. pungens essential oil were analyzed by gas chromatography-mass spectrometry (GC–MS). As shown in Table 1, 17 compounds were detected in the L. pungens essential oil, equivalent to 99.54% of the essential oil, Monoterpenes (90.43%) dominated the essential oil, and citral (29.44%) had the highest proportion of monoterpenes. Moreover, sesquiterpenes accounted for 4.68%, which were β-caryophyllene (3.74%), humulene (0.34%) and caryophyllene oxide (0.60%).

Table 1.
The chemical composition of the L. pungens essential oil.
3.2. Effect of L. Pungens Essential Oil on Germination and Seedling Growth (Petri Dish Experiment)
The allelopathic effect of L. pungens essential oil on seed germination of two plants (L. perenne and B. pilosa) was evaluated (Figure 1A–C). The germination rate and germination force of seeds are one of the main index reflecting the seed quality. As shown in Figure 1A,B, the essential oil inhibited (p < 0.05) the seed germination rate of both weeds significantly in concentration dependent manner. Remarkably, under the treatment of high concentration (2.000 mg/mL), it can be observed that the germination of L. perenne and B. pilosa seeds were completely inhibited. Moreover, the germination index of seeds decreased with the increase of essential oils concentration, and the germination indexes of the two seeds were completely suppressed under the highest concentration (Figure 1C). Not only did it affect seed germination, but also had a certain effect on seedling growth (Figure 1D–F). Compared with blank control, except the seedling growth of L. perenne under the concentration of 0.125 mg/mL was greater than negative control, the growth of other seedlings gradually decreased under the increase of essential oil concentration. In particular, under the highest concentration of essential oil (2.000 mg/mL), L. perenne was completely inhibited (100%). However, it can be seen from the figure that the growth of seedlings was completely inhibited at 1.000 mg/mL and 2.000 mg/mL. It also can be seen that the effect of essential oil on the growth of B. pilosa seedlings was significantly greater than that on the growth of L. perenne seedlings. In a word, essential oils have a certain dose effect on seed germination and seedling growth.

Figure 1.
Effect of different concentration of L. pungens essential oil on seed germination and seedling growth (CK1, blank control; CK2, negative control; (A), Germination rate; (B), Germination force; (C), Germination index; (D), Shoot length; (E), Root length; (F), Plant length). Note: different lowercase letters (a–g) represent statistical differences (Tukey’s test p < 0.05).
3.3. Effect of L. Pungens Essential Oil on Photosynthetic Pigments Content (Petri Dish Experiment)
Effects of different concentrations of essential oils on photosynthetic pigments of L. perenne and B. pilosa are shown in Figure 2A–C. The results show that the chlorophyll a and total chlorophyll content of the two plants decreased as the concentration of essential oil increased. Under the treatment of essential oils, the chlorophyll b content of the two plants did not change regularly, but the lowest chlorophyll content was still at the concentration of 1.000–2.000 mg/mL. It can be further seen from Figure 2 that the synthesis of photosynthetic pigments was affected after the treatment under high concentrations of essential oil (2.000 mg/mL), which in turn affected the photosynthesis of plants.

Figure 2.
Effect of different concentration of L. pungens essential oil on photosynthetic pigments content (CK1, blank control; CK2, negative control; (A), Chlorophyll a content; (B), Chlorophyll b content; (C), Total chlorophyll content). Note: different lowercase letters (a–f) represent statistical differences (Tukey’s test p < 0.05).
3.4. Pot Experiment
3.4.1. Seedling Growth Parameters
As shown in Figure 3, L. perenne caused obvious damage after spraying with different concentrations of L. pungens essential oil solution. On the fifth day of spraying, the leaves’ etiolation and necrosis appeared. Especially, the effect on L. perenne was the greatest at 2.000 mg/mL. The results in Figure 4 show that the concentration of (0.250–2.000) mg/mL of L. pungens essential oil led to a significant (p < 0.05) decrease in seedling growth. Except at shoot length of 0.125 mg/mL and root length of negative control, other treatment groups showed a dose-dependent relationship with essential oil.

Figure 3.
L. perenne in pot experiment (The concentration of potted plants from left to CK1 (blank control), CK2 (negative control), (2.000–0.125) mg/mL).

Figure 4.
Effect of different concentration of L. pungens essential oil on seedling growth (CK1, blank control; CK2, negative control; (A), Shoot length; (B), Root length; (C), Plant length). Note: different lowercase letters (a–f) represent statistical differences (Tukey’s test p < 0.05).
3.4.2. Photosynthetic Pigments Content
Chlorophyll and carotene are light-harvesting pigments, whose changes are related to photosynthesis. The results show that chlorophyll a, b, and carotenoid contents decreased with the increase of essential oil concentration (Figure 5A–D). In particular, under the concentration of 2.000 mg/mL, the difference was significant (p < 0.05) compared with the blank control, in which the content of chlorophyll a, b, total chlorophyll, and carotene decreased by 70.7%, 83.9%, 73.6%, 75%, respectively. Moreover, it can be seen from Figure 5 that except for the chlorophyll b measured after 0.125 mg/mL treatment, a similar decrease in all pigments suggests that the photosynthesis of L. perenne plants was inhibited, further affecting plant growth.

Figure 5.
Effect of different concentration of L. pungens essential oil on photosynthetic pigments content (CK1, blank control; CK2, negative control; (A), Chlorophyll a content; (B), Chlorophyll b content; (C), Total chlorophyll content; (D), Carotenoids content). Note: different lowercase letters (a–d) represent statistical differences (Tukey’s test p < 0.05).
3.4.3. Malondialdehyde (MDA) Content and Electrolyte Leakage (EL)
The content of malondialdehyde (MDA) is the embodiment of the degree of membrane peroxidation in plant cells. After spraying with essential oils, the visible injury observed on L. perenne plants was presumed to be due to electrolyte leakage and membrane integrity. As shown in Figure 6A,B, different concentrations of essential oils had effects on MDA content and membrane leakage of L. perenne. The results show that the membrane leakage increased with the increase of MDA concentration, which was almost proportional to the concentration. Compared with the blank control, the difference is significant (p < 0.05), especially when the concentration of essential oil is 2.000 mg/mL, where MDA content and EL is the highest. Conclusively, the increase of the content of MDA and EL indicates that the degree of peroxidation of the plant cell membrane increases, accompanying by the gradually aggravating cell membrane damage.
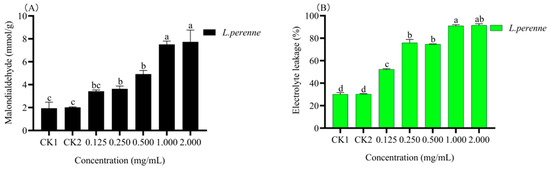
Figure 6.
Effect of different concentration of L. pungens essential oil on malondialdehyde concentration and electrolyte leakage (CK1, blank control; CK2, negative control; (A), Malondialdehyde; (B), Electrolyte leakage). Note: different lowercase letters (a–c) represent statistical differences (Tukey’s test p < 0.05).
3.4.4. Antioxidant Enzymes Activity
Catalase (CAT), superoxide dismutase (SOD), and peroxidase (POD) belong to antioxidant enzymes, and the levels of their activity are related to the growth of plants. The CAT, SOD and POD activity of L. perenne. were significantly affected (p < 0.05) in response to spraying various concentrations of essential oils (Figure 7A–C). CAT enzyme activity increased first and then decreased, and the highest activity appeared at 0. 250 mg/mL (Figure 7A). The activity of the SOD enzyme was similar to that of the CAT enzyme, and the activity increased first and then decreased, among which the highest activity was at 0.250 mg/mL (Figure 7B). As shown in Figure 7C, the POD enzyme’s activity increased with the increase of concentration, which was almost proportional to the concentration. It can be seen that the treatment of essential oil has a significant effect on antioxidant enzyme.
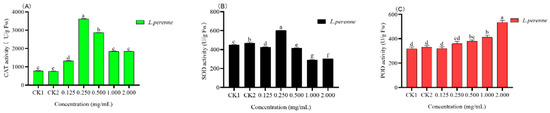
Figure 7.
Effect of different concentration of L. pungens essential oil on catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD) enzymes activity (CK1, blank control; CK2, negative control; (A), CAT activity; (B), SOD activity; (C), POD activity). Note: different lowercase letters (a–g) represent statistical differences (Tukey’s test p < 0.05).
4. Discussion
In this study, the allelopathy of the L. pungens essential oil was evaluated. As far as we know, there are relatively few studies on the chemical components of the essential oil of L. pungens. In this study, limonene (30.29%), neral (29.44%) and linalool (6.38%) are the main components of the L. pungens essential oil. According to reports, Zhang et al. found that 1, 8-cineole was the main chemical component of the essential oil in the cotyledon of L. pungens [26]. Jiang et al. also studied and found that 1, 8-cineole was the most abundant essential oil in the cotyledon and twig of L. pungens [20], indicating that the essential oils extracted from different parts of one plant were different. Due to the lack of studies on L. pungens, we compared the components of the essential oil of Litsea cubeba which belongs to the same genus and has similar appearance and smell. Si et al. found that limonene (0.7–5.3%) in the essential oil of Litsea cubeba fruit exists only as a minor constituent in the essential oil [27], then Su et al. detected and analyzed the components of essential oil in the four parts of Litsea cubeba, flower, branch and fruit, and found that the main component of essential oil in leaf, flower and branch was 1, 8-cineole, and the main components of fruit were limonal and limonene [28]. It could be seen that there are similar differences in the main components of essential oil in different parts of Litsea cubeba. These differences may be related to the synthetic and metabolic differences of secondary metabolites in different parts of plant growth. The main components of the L. pungens essential oil are similar to that of Litsea cubeba, but the contents are different. It is speculated that the differences of these components may be caused by the genetic differences of species, or may be related to the differences of growth environment.
Essential oil from a natural plant is a valuable potential for allelopathic substance [6]. There are few research works on the bioactivities of the essential oil from L. pungens. Hence, the allelopathic activity in this work was firstly report. The results show that the essential oil had a strong allelopathy effect on both L. perenne and B. pilosa, especially at the concentration of 2.000 mg/mL, the germination and seedling growth of the two kinds of seeds were completely inhibited in the Petri dish test. In addition, the pot experiment also showed that after spraying L. perenne with different concentrations of essential oil, the plants appeared yellowing, necrosis and other phenomena, which were significantly different compared with the control group. According to literature reports, this phenomenon was also found in the previous study [29]. Angelini et al. reported that four monoterpenoids in three essential oils had a strong inhibitory effect on three annual weeds and three crop seeds [1]. Other monoterpenoids and sesquiterpenes in the essential oil from Peumus boldus and Drimys winterii were also found to exhibit an inhibitory effect on seed germination and seedling growth [30]. In addition, some researchers reported that terpenoids could change the permeability of cell membrane, enzyme activity, DNA transcription and RNA translation, thus inhibiting seed germination and seedling growth [31,32]. As shown in Table 1, terpenoids were the main components in the essential oil from L. pungens. Therefore, the allelopathy capacity of the essential oil may be associated with the high content of terpenoids.
After treatment with essential oil, the leaves of L. perenne were pale yellow and short, and the chlorophyll content decreased significantly, indicating that the essential oil treatment reduced the chlorophyll content of leaves and affected photosynthesis. These results are highly consistent with previous reports that some essential oils had a negative effect on chlorophyll content [10,33,34,35,36]. In addition, other works suggested that volatile allelochemicals might affect photosynthesis by reducing the formation of photosynthetic pigments [2]. However, further investigations are needed on whether the essential oil directly or indirectly interfered with photosynthesis.
In the life of plants, the balance of generation and elimination of reactive oxygen species (ROS) was regulated by the antioxidant enzyme system. Likewise, SOD and POD could scavenge the increasing ROS under acute stress [37]. Hence, the activities of antioxidant enzymes were closely related to the plant protection system [38,39,40]. In these work, the antioxidant enzyme activity of L. perenne was sharply reduced with the spraying of essential oil from L. pungens, and the content of EL and MDA increased with the increase of the spraying time and concentration of essential oil. The accumulation of ROS could lead to membrane lipid peroxidation on the cell membrane, resulting in the loss of membrane function [41]. Moreover, the stimulation of monoterpenoids on these enzymes was found to lead to the overproduction of ROS [42]. In addition, monoterpenes also possessed strong lipophile properties, thus inducing membrane expansion, increasing fluidity, destroying membrane structure, and causing the inactivation of antioxidant enzyme system [35,43,44]. Therefore, the stimulation of monoterpenoids in the essential oil might account for the lower activity of antioxidant enzymes.
5. Conclusions
In conclusion, the essential oil of L. pungens has a strong allelopathic effect and a herbicide activity on weeds. We found that in the Petri dish experiment, the concentration of 2.000 mg/mL of essential oil completely inhibited the seed germination and seedling growth of L. perenne and B. pilosa. Secondly, in the pot experiment, it was also found that with the increase of the concentration of the essential oil of L. pungens, the chlorophyll content, EL, MDA, CAT, SOD and POD also had negative effects on the L. perenne seedlings. This experiment was the first to study the allelopathic effect of essential oil from L. pungens, which laid a foundation for the development of environment-friendly herbicides in the future and provided a new idea. Compared with commercial chemical herbicides, it is safer, non-toxic and easy to degrade. The applications of essential oils in these fields are very promising in the near future, so they are compatible with both environmental health and human health. However, further studies on allelopathic effects of single component or synergies between multiple components of L. pungens essential oil are needed to comprehensively evaluate potential herbicidal effects in order to determine its potential for large-scale use as a pesticide or herbicide.
Author Contributions
Formal analysis, S.F.; Investigation, Q.K., J.L., S.L. and X.W.; Methodology, T.X.; Project administration, M.Y. and T.C.; Software, H.X. and X.Z.; Supervision, C.D.; Writing—review & editing, L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Written informed consent has been obtained from all participants.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Angelini, L.G.; Carpanese, G.; Cioni, P.L.; Morelli, I.; Macchia, M.; Flamini, G. Essential oils from Mediterranean lamiaceae as weed germination inhibitors. J. Agr. Food Chem. 2003, 51, 6158–6164. [Google Scholar] [CrossRef]
- Batish, D.R.; Setia, N.; Singh, H.P.; Kohlia, R.K. Phytotoxicity of lemon-scented eucalypt oil and its potential use as a bioherbicide. Crop Prot. 2004, 23, 1209–1214. [Google Scholar] [CrossRef]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Ercoli, L.; Masoni, A.; Pampana, S.; Arduini, I. Allelopathic effects of rye, brown mustard and hairy vetch on redroot pigweed, common lambsquarter and knotweed. Allelopathy J. 2007, 19, 249–256. [Google Scholar]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorgan. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, H.P.; Mittal, S. Phytotoxic effects of volatile oil from Artemisia scoparia against weeds and its possible use as a bioherbicide. Ind. Crops Prod. 2010, 32, 54–61. [Google Scholar] [CrossRef]
- Mekky, M.S.; Hassanien, A.M.; Kamel, E.M.; Ismail, A.E.A. Allelopathic effect of Ocimum basilicum L. extracts on weeds and some crops and its possible use as new crude bio-herbicide. Ann. Agric. Sci. 2019, 64, 211–221. [Google Scholar] [CrossRef]
- Ahuja, N.; Batish, D.R.; Singh, H.P.; Kohli, R.K. Herbicidal activity of eugenol towards some grassy and broad-leaved weeds. J. Pest Sci. 2015, 88, 209–218. [Google Scholar] [CrossRef]
- Feldlaufer, M.F.; Ulrich, K.R. Essential Oils as Fumigants for Bed Bugs (Hemiptera: Cimicidae). J. Entomol. Sci. 2015, 50, 129–137. [Google Scholar] [CrossRef]
- Laosinwattana, C.; Wichittrakarn, P.; Teerarak, M. Chemical composition and herbicidal action of essential oil from Tagetes erecta L. leaves. Ind. Crops Prod. 2018, 126, 129–134. [Google Scholar] [CrossRef]
- Aghbash, B.N.; Pouresmaeil, M.; Dehghan, G.; Nojadeh, M.S.; Mobaiyen, H.; Maggi, F. Chemical composition, antibacterial and radical scavenging activity of essential oils from Satureja macrantha C.A.Mey. At different growth stages. Foods 2020, 9, 494. [Google Scholar] [CrossRef]
- Saric-Krsmanovic, M.; Umiljendic, J.G.; Radivojevic, L.; Rajkovic, M.; Santric, L.; Durovic-Pejcev, R. Chemical composition of Ambrosia trifida essential oil and phytotoxic effect on other plants. Chem. Biodivers. 2020, 17, e1900508. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest. Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, J.A.; Burke, I.C.; Everman, W.J.; Vangessel, M.J.; Davis, V.M.; Sikkema, P.H. Potential corn yield losses from weeds in North America. Weed Technol. 2016, 30, 979–984. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Lee, K.E.; Bajpai, V.K.; Gajurel, P.R.; Gu, K.S.; Kumar, P. Ethnopharmacological properties and medicinal uses of Litsea cubeba. Plants-Basel 2019, 8, 150. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y. Chemical composition and antibacterial activity of essential oils from different parts of Litsea cubeba. Chem. Biodivers. 2010, 7, 229–235. [Google Scholar] [CrossRef]
- Hu, W.; Li, C.; Dai, J.; Lin, L. Antibacterial activity and mechanism of Litsea cubeba essential oil against methicillin-resistant Staphylococcus aureus (MRSA). Ind. Crops Prod. 2019, 130, 34–41. [Google Scholar] [CrossRef]
- Yang, K.; Wang, C.F.; You, C.X.; Geng, Z.F.; Sun, R.Q.; Guo, S.S.; Du, S.S.; Liu, Z.L.; Deng, Z.W. Bioactivity of essential oil of Litsea cubeba from China and its main compounds against two stored product insects. J. Asia-Pac. Entomol. 2014, 17, 459–466. [Google Scholar] [CrossRef]
- Zheng, K.W.; Li, W.; Fu, B.Q.; Ren, Q.L.; Yang, F.; Qin, C.Q. Physical, antibacterial and antioxidant properties of chitosan films containing hardleaf oatchestnut starch and Litsea cubeba oil. Int. J. Biol. Macromol. 2018, 118, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Akhtar, Y.; Bradbury, R.; Zhang, X.; Isman, M.B. Comparative toxicity of essential oils of L. pungens and Litsea cubeba and blends of their major constituents against the cabbage looper, Trichoplusia ni. J. Agric. Food Chem. 2009, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Ibrahim, M.; Zahiruddin, S.; Parveen, R.; Khan, W.; Ahmad, S.; Shrivastava, B.; Shrivastava, A.K. TLC-bioautography identification and GC-MS analysis of antimicrobial and antioxidant active compounds in Musa × paradisiaca L. fruit pulp essential oil. Phytochem. Anal. 2019, 30, 332–345. [Google Scholar] [CrossRef]
- Benchaa, S.; Hazzit, M.; Abdelkrim, H. Allelopathic effect of Eucalyptus citriodora essential oil and its potential use as bioherbicide. Chem. Biodivers. 2018, 15, e1800202. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Chemical profiling, cytotoxicity and phytotoxicity of foliar volatiles of Hyptis suaveolens. Ecotox. Environ. Safe 2019, 171, 863–870. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method. Enzymol. 1987, 148, 350–382. [Google Scholar]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, H.L.; Wang, Y. Study on the chemical constituents of the essential oil from the leaves of L. pungens Hemsl. Nat. Prod. Res. 1992, 4, 20–23. [Google Scholar]
- Si, L.; Chen, Y.; Han, X.; Zhan, Z.; Tian, S.; Cui, Q.; Wang, Y. Chemical composition of essential oils of Litsea cubeba harvested from its distribution areas in China. Molecules 2012, 8, 7057. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Ho, C.L. Essential oil compositions and antimicrobial activities of various parts of Litsea cubeba from Taiwan. Nat. Prod. Commun. 2016, 11, 515–518. [Google Scholar] [CrossRef]
- Ben, K.S.; Lins, L.; Hanafi, M.; Bettaieb, R.I.; Deleu, M.; Fauconnier, M.L.; Ksouri, R.; Jijakli, M.H.; Clerck, C. Cynara cardunculus crude extract as a powerful natural herbicide and insight into the mode of action of its bioactive molecules. Biomolecules 2020, 10, 209. [Google Scholar]
- Verdeguer, M.; García-Rellán, D.; Boira, H.; Pérez, E.; Gandolfo, S.; Blazquez, M.A. Herbicidal activity of Peumus boldus and Drimys winterii essential oils from Chile. Molecules 2011, 16, 403–411. [Google Scholar] [CrossRef] [PubMed]
- El-Shora, H.M.; Abd El-Gawad, A.M. Evaluation of allelopathic potential of Rumex dentatus root extract and allelochemicals on Cicer arietinum. J. Physiol. Biochem. 2014, 10, 167–180. [Google Scholar]
- Marco, C.A.; Teixeira, E.; Simplício, A.; Oliverira, C.; Costa, J.; Feitosa, J. Chemical composition and allelopathyc activity of essential oil of Lippia sidoides cham. Chil. J. Agr. Res. 2012, 72, 157–160. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Setia, N.; Kohli, R.K.; Kaur, S.; Yaday, S.S. Alternative control of little seed canary grass using eucalypt oil. Agron Sustain. Dev. 2007, 27, 171–177. [Google Scholar] [CrossRef]
- Mohammad, P.; Mohsen, S.N.; Ali, M.; Filippo, M. Exploring the bio-control efficacy of Artemisia fragrans essential oil on the perennial weed Convolvulus arvensis: Inhibitory effects on the photosynthetic machinery and induction of oxidative stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar]
- Poonpaiboonpipat, T.; Pangnakorn, U.; Suvunnamek, U.; Teerarak, M.; Charoenying, P.; Laosinwattana, C. Phytotoxic effects of essential oil from Cymbopogon citratus and its physiological mechanisms on barnyardgrass (Echinochloa crus-galli). Ind. Crops Prod. 2013, 41, 403–407. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Ramezani, H.; Kohli, R.K. Comparative phytotoxicity of four monoterpenes against Cassia occidentalis. Ann. Appl. Biol. 2002, 141, 111–116. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Mutlu, S.; Atici, O.; Nalbantoglu, B. Effects of salicylic acid and salinity on apoplastic antioxidant enzymes in two wheat cultivars differing in salt tolerance. Bio. Plantarum. 2009, 53, 334–338. [Google Scholar] [CrossRef]
- Mutlu, S.; Atici, O.; Kaya, Y. Effect of cement dust on the diversity and the antioxidant enzyme activities of plants growing around a cement factory. Fresen. Environ Bull. 2009, 18, 1823–1827. [Google Scholar]
- Mutlu, S.; Atici, O. Allelopathic effect of Nepeta meyeri Benth. extracts on seed germination and seedling growth of some crop plants. Acta Physiol. Plant. 2009, 31, 89–93. [Google Scholar] [CrossRef]
- Amiri, H. Volatile constituents and antioxidant activity of flowers, stems and leaves of Nasturtium officinale R. Br. Nat. Prod. Res. 2012, 26, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Bailly, C.; Gniazdowska, A.; Come, D.; Corbineau, F.; Bogatek, R. Induction of oxidative stress by sunflflower phytotoxins in germinating mustard seeds. J. Chem. Ecol. 2007, 33, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Turina, A.V.; Perillo, M.A. Monoterpenes affect chlorodiazepoxide–micelle interaction through micellar dipole potential modififications. BBA-Biomembranes 2003, 1616, 112–120. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).