Abstract
The phenomenon of rhizobial synergy was investigated to increase the efficiency of nitrogen-fixing symbiosis of alfalfa (Medicago varia Martyn), common vetch (Vicia sativa L.) or red clover (Trifolium pratense L.). These plants were co-inoculated with the respective commercial strains Sinorhizobium meliloti RCAM1750, Rhizobium leguminosarum RCAM0626 or R. leguminosarum RCAM1365 and with the strains Mesorhizobium japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343 isolated from the relict legumes Oxytropis popoviana Peschkova and Astragalus chorinensis Bunge. The isolates mentioned above had additional symbiotic genes (fix, nif, nod, noe and nol) as well as the genes promoting plant growth and symbiosis formation (acdRS, genes associated with the biosynthesis of gibberellins and auxins, genes of T3SS, T4SS and T6SS secretion systems) compared to the commercial strains. Nodulation assays showed that in some variants of co-inoculation the symbiotic parameters of plants such as nodule number, plant biomass or acetylene reduction activity were increased. We assume that the study of microbial synergy using rhizobia of relict legumes will make it possible to carry out targeted selection of co-microsymbionts to increase the efficiency of agricultural legume–rhizobia systems.
1. Introduction
Legumes play an important role in natural ecosystems due to their ability to form symbiosis with nitrogen-fixing nodule bacteria (rhizobia). The formation of a mutually beneficial symbiosis allows the plant-microbial system to acquire new adaptive properties that increase its productivity and resistance to stress, as well as to ensure successful competition for ecological space. During co-evolution of leguminous plants and root nodule bacteria, the signaling system of interaction between symbionts arose, providing specific recognition and integration between partners. Knowledge about the mechanisms of specificity and evolution of symbiosis is necessary for science-based selection and design of highly effective plant-microbe agricultural systems in which the potential productivity of the plant is most realized. However, the complex mechanisms underlying the specificity of legume–rhizobia interactions and their optimization are still poorly understood, including due to the lack of adequate experimental models. In this respect the symbiotic systems based on the relict leguminous plants may be of particular importance.
Earlier a representative collection of microsymbionts of Miocene–Pliocene relict legumes Vavilovia formosa (Steven) Fed., Oxytropis triphylla (Pall.) DC., O. popoviana Peschkova, Astragalus chorinensis Bunge and Caragana jubata (Pall.) Poir. originating from the North Caucasus and the Baikal Lake region (Russia), as well as from the Khuvsgul Lake region in Mongolia [1,2,3,4,5] was created. A large taxonomic diversity of the collected microsymbionts was demonstrated, including representatives of the families Phyllobacteriaceae, Rhizobiaceae and Bradyrhizobiaceae. Using these microsymbionts, the phenomenon of rhizobial synergy was discovered, which was expressed in the ability of strains with different taxonomic position and complementary sets of symbiotic genes to simultaneously be present in nodules and increase the rate of nodulation, number of nodules, nitrogen-fixing activity and plant biomass [3,4]. It was also shown that strains isolated from relict symbiotic systems may have a wide range of host plants, including symbiotically narrow-specific species of Medicago sativa L. and Trifolium pratense L. [4]. These results suggested that the symbiotic systems of relict plants are formed with various groups of rhizobia, which can be co-microsymbionts and jointly participate in increasing the effectiveness of symbiosis with a wide range of host plants, including agricultural crops.
Thus, symbiotic systems of relict legumes are promising models for studying the evolution of host specificity, the impact of natural rhizobial diversity on plant growth and multipartite interactions between symbiotic partners. In addition, the discovered phenomenon of rhizobial synergy can be used as a new principle to increase the productivity of traditional crop legumes. In this regard, the aim of this work was to study the effectiveness of symbiosis upon joint inoculation of alfalfa (Medicago varia Martyn), common vetch (Vicia sativa L.) and red clover (Trifolium pratense L.) with their commercial strains and strains isolated from O. popoviana and A. chorinensis. Specific tasks were as follows: (1) To select, using a comparative whole-genome analysis, the isolates having additional genes involved in the formation of symbiosis as compared to the commercial strains Sinorhizobium meliloti RCAM1750 (alfalfa), Rhizobium leguminosarum RCAM0626 (common vetch) and R. leguminosarum RCAM1365 (red clover). Attention was paid to genes fix, nif, nod, noe and nol, as well as genes promoting plant growth and symbiosis formation (acdRS; genes associated with the biosynthesis of gibberellins and auxins; genes of T3SS, T4SS and T6SS secretion systems). (2) To study the effects of co-inoculation of alfalfa, common vetch and red clover with commercial strains and the selected isolates in the gnotobiotic plant nodulation assays.
2. Materials and Methods
2.1. Bacterial Material
Commercial strains of rhizobia: Sinorhizobium meliloti RCAM1750 nodulating alfalfa (M. varia), Rhizobium leguminosarum RCAM0626 nodulating common vetch (V. sativa) and R. leguminosarum RCAM1365 nodulating red clover (T. pratense) were used. The rhizobial strains Mesorhizobium japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343 were isolated from nodules of the relict legumes O. popoviana and Astragalus chorinensis. For this purpose, soil samples and seeds of O. popoviana and A. chorinensis were collected in the Republic of Buryatia (Baikal Lake region, Russia). Seeds were surface sterilized and scarified by treatment with 0.1% HgCl2 for 10 min and then 5% NaOCl for 8 min, rinsed carefully with sterile tap water and germinated on filter paper in Petri dishes at 25 °C in the dark for 4 days. Seedlings were transferred to three sterile plastic pots (one pot with 5 seeds) containing 250 g of soil. Plants were cultivated for 60 days in the growth chamber with 50% relative humidity and four-level illumination/temperatures mode: night (dark, 18 °C, 8 h), morning (200 μmol m−2 s−1, 20 °C, 2 h), day (400 μmol m−2 s−1, 23 °C, 12 h), evening (200 μmol m−2 s−1, 20 °C, 2 h). Illumination was performed by L 36W/77 FLUORA lamps (Osram, Munich, Germany). Then roots of individual plants were removed from soil and washed with tap water. Strains of nodule bacteria were isolated from the obtained nodules by the standard method described earlier [6] and were cultivated using modified yeast extract mannitol agar (YMA) [7] supplemented with 0.5% succinate (YMSA) [8]. Information about strains used in this work is given in Table 1. The genera Sinorhizobium and Mesorhizobium belong to fast-growing rhizobia (colonies on agar medium appear on 3–4 days), and the genus Bradyrhizobium—to slowly-growing rhizobia (colonies appear on 6–7 days). All strains were deposited in the Russian Collection of Agricultural Microorganisms (RCAM), registered in the WFCC-MIRCEN World Data Centre for Microorganisms under the number WDCM 966 [9] and stored at −80 °C in the automated Tube Store (Liconic Instruments, Mauren, Lichtenstein) as described previously [10].

Table 1.
Used commercial strains of rhizobia and strains isolated from the relict legumes originated from Baikal Lake region.
2.2. Whole Genome Sequencing of the Strains under Investigation
For the whole genome sequencing of three commercial rhizobial strains deposited in the RCAM collection (S. meliloti RCAM1750, R. leguminosarum RCAM0626 and R. leguminosarum RCAM1365) their genomic DNA was extracted using Genomic DNA Purification KIT (Thermo Fisher Scientific, EU) according to recommendation of manufacturer. Long-read whole genome sequencing was performed using a MinIon sequencer (Oxford Nanopore, England) of the Core Centrum “Genomic Technologies, Proteomics and Cell Biology” at the ARRIAM. The SQK-LSK109 Ligation Sequencing Kit with the EXP-NBD104 Native Barcoding Expansion 1–12 kit was used to prepare the library according to manufacturer instructions. The reads were basecalled and demultiplexed using the guppy_basecaller (v. 3.3.0). The Flye pipeline (v 2.6-release) was used to assemble the Nanopore reads [11]. The resulting assembly was corrected 4 times using Racon (v. 1.3.2) (https://github.com/lbcb-sci/racon, accessed on 22 March 2013) with the following modifiers (−m 8 −x −6 −g −8 −w 500), following with a single polish using the medaka (v 0.10.0) program with default parameters [12]. Search for genes in the assembled contigs was performed using the RAST annotation service [13]. All the genomes were assembled into circular replicons. Genome sequencing of the Baikal isolates Mesorhizobium japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343 was performed on a MiSeq genomic sequencer (Illumina, San Diego, CA, USA), as described earlier [3,4].
Search for homologs of the symbiotic fix, nif, nod, noe and nol genes as well as genes that promote plant growth (acdRS, gibberellin- and auxin-biosynthesis related) and genes of the T3SS, T4SS and T6SS secretion systems in annotated genomes was performed using CLC Genomics Workbench 7.5.1 software using local BLASTn and tBLASTx [14].
The whole genome sequences have been deposited to the NCBI GenBank database under accession numbers: CP050511-CP050513 for the strain S. meliloti RCAM1750; CP050553-CP050557 for the strain R. leguminosarum RCAM0626; CP050514-CP050519 for the strain R. leguminosarum RCAM1365; QKOD00000000 for the isolate M. japonicum Opo-235; MZXX00000000 for the isolate M. japonicum Opo-242; MZXW00000000 for the isolate Bradyrhizobium sp. Opo-243 and MZXV00000000 for the isolate M. kowhaii Ach-343.
2.3. Plant Nodulation Assays
Seeds of alfalfa (M. varia), common vetch (V. sativa) and red clover (T. pratense) plants were surface sterilized by H2SO4 for 10 min, washed with sterile tap water and germinated on filter paper in Petri dishes at 25 °C in the dark for 4 days. Germinated seedlings were transferred to 50 mL glass test tubes (2 seedlings per test tube) which contained 10 mL of sterile agar medium of the following composition (g/L): K2HPO4 1.0, KH2PO4 0.25, MgSO4 1.0, Ca3(PO4)2 0.2, FeSO4 0.02, H3BO3 0.005, (NH4)2MoO4 0.005, ZnSO4 0.005, MnSO4 0.002, agar for micropropagation of plants (Dia-m, Moscow, Russia) 5.0.
After planting to the test tubes, two-days old seedlings were inoculated with individual strains or with a pair of strains in the amount of 106 cells of each strain per test tube (10 tubes per inoculation variant). The cell concentration in suspensions was determined by the SmartSpec Plus Spectrophotometer (BioRad, Hercules, CA, USA). A suspension of each strain was prepared in a liquid medium of the same composition as described above (0.5 mL of each suspension per tube). In all mono-inoculation treatments an additional 0.5 mL of the liquid medium per tube was added. The uninoculated plants were used as negative control. Plants were cultivated for 24 (T. pratense), 25 (M. varia) or 26 (V. sativa) days in the growth chamber with 50% relative humidity and four-level illumination/temperatures mode: night (dark, 18 °C, 8 h), morning (200 μmol m−2 s−1, 20 °C, 2 h), day (400 μmol m−2 s−1, 23 °C, 12 h), evening (200 μmol m−2 s−1, 20 °C, 2 h). Illumination was performed by L 36W/77 FLUORA lamps (Osram, Munich, Germany). At the end of one representative experiment the nitrogen fixation of nodules in each test tube was measured by the acetylene-reduction method [15] using a gas chromatograph GC-2014 (Shimadzu, Japan). Then nodules were counted, shoots and roots were separated at the root collar and the fresh biomass of shoots and roots was immediately determined by the analytical balance Pioneer PX224 (Ohaus, Parsippany, NJ, USA). The level of acetylene reduction activity per one nodule in a tube was calculated. Strains were re-isolated from the obtained nodules (5 nodules from different tubes for each co-inoculation variant) and identified by 16S rDNA sequencing as described earlier [16]. Pictures of roots and nodules were performed by the stereo microscope Stemi 508 (Carl Zeiss, Oberkochen, Germany). The data were processed using the software STATISTICA version 10. Fisher’s least significant difference (LSD) test, standard deviation (SD) and standard error (SE) were used to evaluate differences between means.
3. Results and Discussion
3.1. Whole Genome Sequencing of the Strains under Investigation
The genomic features of the studied strains are presented in Table 2.
Table 2.
Genomic features of the studied isolates and the commercial strains.
The currently described symbiotic fix, nif, nod, noe and nol genes as well as genes that promote plant growth (acdRS, gibberellin- and auxin-biosynthesis related) and genes of the T3SS, T4SS and T6SS secretion systems involved in the formation of symbiosis were searched throughout the whole genome sequences of three commercial rhizobial strains (S. meliloti RCAM1750; R. leguminosarum RCAM0626, RCAM1365) and four strains isolated from the relict legumes O. popoviana (M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243) and A. chorinensis (M. kowhaii Ach-343). Presence of symbiotic fix, nif, nod, noe and nol genes is shown in Table 3.
Table 3.
Presence of the symbiotic genes fix, nif, nod, noe and nol in the commercial rhizobial strains (R. leguminosarum RCAM0626, RCAM1365; S. meliloti RCAM1750) and four strains isolated from the relict legumes O. popoviana and A. chorinensis (M. japonicum Opo-235, Opo-242; Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343).
The common nodABC genes necessary for legume nodulation [17] were found in all commercial strains and only in three isolates: M. japonicum Opo-235, M. japonicum Opo-242 and M. kowhaii Ach-343. These strains possessed also nif and fix genes required for nitrogen fixation and forming clusters: nifHDK and nifENB genes encoding structural and catalytic components of the nitrogenase complex [18] as well as fixABCX genes participating in electron transfer to nitrogenase [19]. At the same time, some of fixNOPQ and fixGHIS genes participating in symbiotically essential oxidase complex [20] were not observed in the commercial strains: the strain R. leguminosarum RCAM1365 lacked fixQHS; the strains R. leguminosarum RCAM0626 and S. meliloti RCAM1750 lacked fixS and fixH, respectively (Table 3). The isolate Bradyrhizobium sp. Opo-243 had only one group of fixKJLNOQPGHIS genes, which formed a single cluster. All the studied strains possessed some nol and noe genes, as well as additional nod, nif and fix genes. It should be noted that the set of symbiotic genes for each strain was unique.
Presence of the genes related to plant growth promotion (acdRS, gibberellin- and auxin-biosynthesis related genes) is shown in Table 4.
Table 4.
Presence of the genes that promote plant growth in the commercial rhizobial strains (R. leguminosarum RCAM0626, RCAM1365; S. meliloti RCAM1750) and four strains isolated from the relict legumes O. popoviana and A. chorinensis (M. japonicum Opo-235, M. japonicum Opo-242; Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343).
It was repeatedly shown that the acdS gene of rhizobia encoding enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase plays important role in nodulation process [21]. Particularly, pea plants inoculated with ACC deaminase minus mutants of Rhizobium leguminosarum bv. viciae 128C53K [22] and 1066S [23] showed decreased nodule number, nitrogenase activity and shoot biomass as compared with a wild type. The commercial strains R. leguminosarum RCAM0626 and RCAM1365 did not contain the acdS gene. At the same time, the strain S. meliloti RCAM1750, as well as the isolates Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343, had both acdS and acdR genes required for the synthesis of ACC deaminase and its regulation, respectively. The isolates M. japonicum Opo-235 and M. japonicum Opo-242 contained only acdS gene.
All isolates had significantly more genes associated with the biosynthesis of gibberellins and auxins compared to the commercial strains (Table 4). It should be noted that gibberellin- and auxin-biosynthesis related genes are commonly known as factors responsible for plant growth, the efficiency of plant–microbe interactions and symbiosis formation. For example, they are involved in the processes of nodule initiation and development [24,25,26].
The genes of T3SS, T4SS and T6SS secretion systems presented in the studied strains are shown in Table 5. The T3SS secretion system was scarcely present in either commercial or isolated strains by one fli gene in the isolates M. japonicum Opo-235 and M. kowhaii Ach-343. In contrast, all strains, with the exception of the R. leguminosarum RCAM0626, possessed multiple vir genes belonging to theT4SS playing an important role in the translocation of a wide range of virulence factors into the host cell [27,28]. No genes of the T6SS secretion system were found in the commercial strains or in the isolate Bradyrhizobium sp. Opo-243, while four remaining isolates had different sets of these genes (Table 5). Most of them (icmF, tssABCEGJKL, tagFH, vgrG, vasAEFK, clpV1, impA, vipAB, sciN, dotU, Vgr family, Hcp family, FHA domain) was presented in the isolate M. kowhaii Ach-343. The T6SS secretion system, like T3SS and T4SS, is involved in bacterial virulence due to secretion of broad classes of effectors proteins: cytotoxins; lysozymes; lipoproteins; factors of adherence to epithelial cells and penetrating system, pilus formation, conjugation, protein translocation and so on [29,30,31,32]. It is known that these secretion systems in rhizobia can affect the development of plant–microbe interactions and the host specificity of strains, as well as determine the number of nodules formed [33].
Table 5.
Presence of the secretion systems T3SS, T4SS and T6SS genes in the commercial rhizobial strains (R. leguminosarum RCAM0626, RCAM1365; S. meliloti RCAM1750) and four strains isolated from the relict legumes O. popoviana and A. chorinensis.*—commercial strain, **—isolate from the relict legumes.
3.2. Plant Nodulation Assays
The plant nodulation tests were performed on alfalfa, common vetch and red clover plants with the participation of the corresponding commercial strains and the isolates M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343. Mono- and co-inoculation of alfalfa plants showed that three out of four isolates (except for M. kowhaii Ach-343) led to a significant increase in the number of nodules in variants of joint inoculation with the commercial strain S. meliloti RCAM1750 (Table 6). In some cases, this was accompanied by an increase in the biomass of shoots and the total biomass of plants (roots and shoots). An enhanced total nitrogen-fixing activity was observed only when the strain S. meliloti RCAM1750 was co-inoculated with Bradyrhizobium sp. Opo-243 (Figure 1), although the level of activity calculated per one nodule did not differ from the commercial strain (data not shown). After co-inoculation with S. meliloti RCAM1750 and M. kowhaii Ach-343, despite a significant decrease in nodule number and nitrogen-fixing activity, an increase in the biomass of roots, shoots and total plant was observed.
Table 6.
Effect of mono- and co-inoculation of alfalfa with the commercial strain S. meliloti RCAM1750 and the isolates M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343 in the sterile plant nodulation assay. The data means ± standard errors of one representative experiment (n = 10). Different letters show significant differences between treatments (Fisher’s LSD test, p < 0.05). ↑ and ↓—significant increase or decrease, respectively, between inoculation with the strain RCAM1750 and other treatments. FW stands for fresh weight.
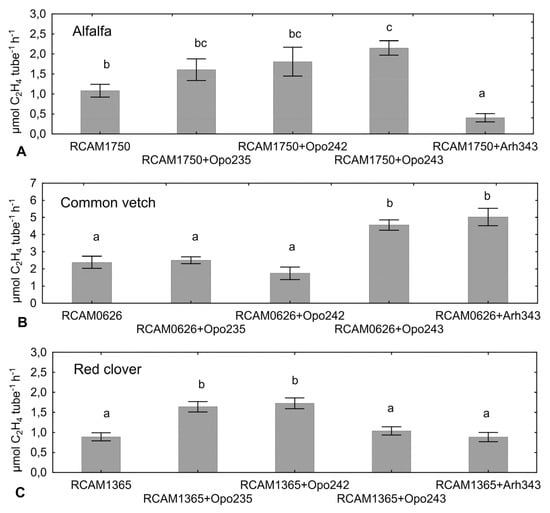
Figure 1.
Acetylene reduction activity level in variants of mono- and co-inoculation of alfalfa (A), common vetch (B) and red clover (C) with the commercial strains S. meliloti RCAM1750, R. leguminosarum RCAM0626 or R. leguminosarum RCAM1365, respectively, and the isolates from the relict legumes. Different letters show significant differences between treatments (Fisher’s LSD test, p < 0.05). Vertical bars represent SE.
The isolate M. japonicum Opo-235 co-inoculated with S. meliloti RCAM1750 contributed to significant acceleration of the nodule formation in comparison (Figure 2) and also led to the increase in plant biomass of alfalfa, although no increase in the total nitrogen-fixing activity was detected (Table 6, Figure 1).
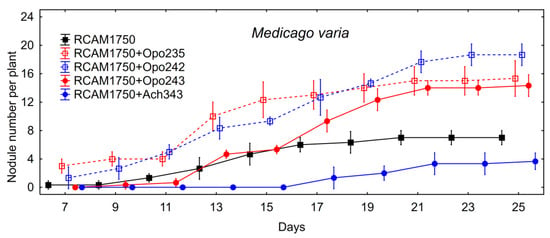
Figure 2.
Nodule formation on alfalfa roots after mono- and co-inoculation with the commercial strain S. meliloti RCAM1750 and the isolates M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 and M. kowhaii Ach-343 in the sterile plant nodulation assay. Vertical bars represent SD.
The other two isolates M. japonicum Opo-242 and Bradyrhizobium sp. Opo-243 also contributed to the formation of more nodules. However, M. japonicum Opo-242 had no effect on plant biomass, while Bradyrhizobium sp. Opo-243 increased the biomass of alfalfa shoots. It was shown that the nodules in the variant S. meliloti RCAM1750 + M. japonicum Opo-242 were small, and among them there were many tumor-like structures (Figure 3).

Figure 3.
Pictures of the alfalfa roots in the sterile nodulation assay. Variants of mono-inoculation: (A) commercial strain S. meliloti RCAM1750, (B)—isolate M. japonicum Opo-235, (C) isolate Bradyrhizobium sp. Opo-243; variants of co-inoculation: (D) S. meliloti RCAM1750 + M. japonicum Opo-242, (E) S. meliloti RCAM1750 + M. japonicum Opo-235, (F) S. meliloti RCAM1750 + Bradyrhizobium sp. Opo-243. Scale bar = 0.5 mm.
When the isolates M. japonicum Opo-235 and Bradyrhizobium sp. Opo-243 were used together with the strain S. meliloti RCAM1750, almost all nodules had a typical elongated shape, although both isolates themselves formed a small number of abnormal structures on the alfalfa roots (Figure 3). None of the isolates were detected in the alfalfa nodules and tumors either in mono-inoculation or in co-inoculation variants.
The data of plant nodulation experiment on mono- and co-inoculation of common vetch with the commercial strain R. leguminosarum RCAM0626 and the isolates are shown in the Table 7. Unlike alfalfa plants, the isolates themselves did not form nodules or tumors on the common vetch roots. Isolate M. kowhaii Ach-343 increased nodule number when co-inoculated with R. leguminosarum RCAM0626, while the isolate M. japonicum Opo-242 had no effect on this parameter. The level of total nitrogen-fixing activity enhanced upon co-inoculation with R. leguminosarum RCAM0626 and Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343 (Figure 1). However, this was associated with the formation of a significantly larger number of nodules, but not with their activity per one nodule (data not shown). An increase in the nodule number and the level of nitrogen-fixing activity had practically no effect on the plant biomass (Table 7). It is possible that some unknown growth limiting factors were presented under model gnotobiotic conditions and a potential of such symbiosis for biomass production could be more pronounced under optimal conditions. The study of the nodulation rate on the common vetch roots showed that only Bradyrhizobium sp. Opo-243 contributed to a significantly faster nodule formation compared to inoculation with R. leguminosarum RCAM0626 (Figure 4).
Table 7.
Effect of mono- and co-inoculation of common vetch with the commercial strain R. leguminosarum RCAM0626 and the isolates M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343 in the sterile plant nodulation assay. The data means ± standard errors of one representative experiment (n = 10). Different letters show significant differences between treatments (Fisher’s LSD test, p < 0.05). ↑ and ↓—significant increase or decrease, respectively, between inoculation with the strain RCAM0626 and other treatments. FW stands for fresh weight.
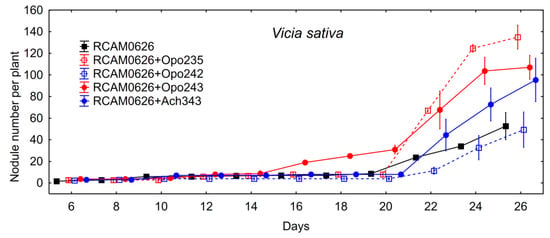
Figure 4.
Nodule formation on common vetch roots after mono- and co-inoculation with the commercial strain R. leguminosarum RCAM0626 and the isolates in the sterile plant nodulation assay. Vertical bars represent SD.
In the experiment on T. pratense, the increase in the level of nitrogen-fixing activity was detected when R. leguminosarum RCAM1365 was combined with M. japonicum Opo-235 or M. japonicum Opo-242 (Figure 1). The level of activity calculated per one nodule in these two variants was also statistically significant increased (129.1 and 125.4 nmol C2H4 nodule−1 h−1, respectively), compared to mono-inoculation with the commercial strain RCAM1365 (66.2 nmol C2H4 nodule−1 h−1).
In this regard, it is important to note that in these two variants of co-inoculation the isolates were re-isolated from nodules (the data not presented). At the same time, M. japonicum Opo-235 itself formed a small number of typical elongated nodules on the T. pratense roots containing cells of this strain, whereas the isolate M. japonicum Opo-242 formed tumors (Figure 5) without bacteria inside them. Studying the nodulation rate on red clover did not reveal any variants of co-inoculation that differed from mono-inoculation with the commercial strain R. leguminosarum RCAM1365 (data not shown).
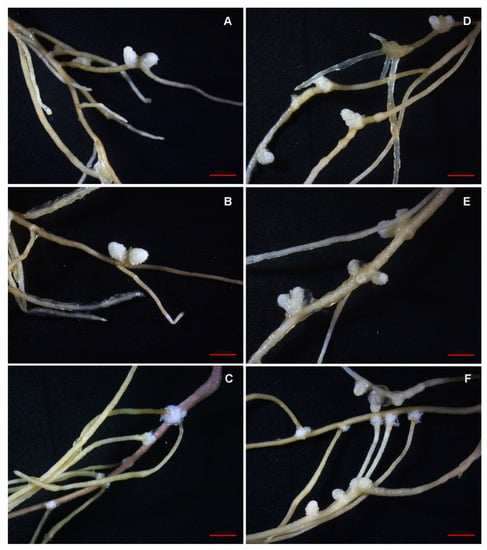
Figure 5.
Pictures of red clover nodules in the sterile nodulation assay. Variants of mono-inoculation: (A) commercial strain R. leguminosarum RCAM1365, (B) isolate M. japonicum Opo-235, (C) isolate M. japonicum Opo-242; variants of co-inoculation: (D) R. leguminosarum RCAM1365 + Bradyrhizobium sp. Opo-243, (E) R. leguminosarum RCAM1365 + M. japonicum Opo-235, (F) R. leguminosarum RCAM1365 + M. japonicum Opo-242. Scale bar = 2.0 mm.
No change in the nodule number was observed as compared to the commercial strain (Table 8). It should be noted that the increase in the level of nitrogen-fixing activity did not lead to a significant increase in plant weight.
Table 8.
Effect of mono- and co-inoculation of red clover with the commercial strain R. leguminosarum RCAM1365 and the isolates M. japonicum Opo-235, M. japonicum Opo-242, Bradyrhizobium sp. Opo-243 or M. kowhaii Ach-343 in the sterile plant nodulation assay. The data means ± standard errors of one representative experiment (n = 10). Different letters show significant differences between treatments (Fisher’s LSD test, p < 0.05). ↑ and ↓—significant increase or decrease, respectively, between inoculation with the strain RCAM1365 and other treatments. FW stands for fresh weight.
3.3. Comparison of the Genomic Data and Plant Nodulation Assays
We tried to explain the influence of the Baikal isolates on the efficiency of symbiosis with alfalfa, common vetch and red clover using a comparative genomic analysis of all strains that participated in plant nodulation experiments. Symbiotic and non-symbiotic genes that are absent in the commercial strains S. meliloti RCAM1750, R. leguminosarum RCAM0626 and R. leguminosarum RCAM1365 but are present in the isolates are shown in Table 9. Based on the analysis of the results obtained in the nodulation assay with alfalfa it can be assumed that the symbiotic genes nodLZ, nolKL and noeKL could act as the main accessory genes of the isolates, which contributed to an increase in the rate of nodulation and the efficiency of symbiosis with this plant species. The genes nodL and nodZ perform, respectively, 6-O-acetylation and fucosilation of the non-reducing end of the Nod factor (NF) and has been shown to be important for symbiosis of alfalfa and soybean with Sinorhizobium strains [34,35,36,37]. The noeK, noeL, and nolK genes associated with the expression of the nodZ gene are involved in the synthesis of the precursor of the fucosyl group from mannose [37,38,39]. The gene nolL carries out 4-O-acetylation of the fucose residue in NF, which arises as a result of the work of the gene nodZ [40]. Thus, all of the genes discussed above are responsible for the specificity of the plant–rhizobia interactions by NFs modification (acetylation and fucosilation). The nodT gene, also found in the isolates but not in S. meliloti RCAM1750 (Table 9), encodes an outer membrane lipoprotein playing a role in the secretion of NFs [41,42].
Table 9.
Symbiotic and non-symbiotic genes that are present in the isolates and absent in the commercial strains S. meliloti RCAM1750, R. leguminosarum RCAM0626 and R. leguminosarum RCAM1365. SG—symbiotic genes, NSG—non-symbiotic genes.
However, a positive effect of the gene nodT on the formation of symbiosis upon joint inoculation with the commercial strain seems unlikely. We also do not focus on the nif and fix genes of isolates that are absent in the strain S. meliloti RCAM1750, since none of the isolates was detected in the alfalfa nodules and tumors either in mono-inoculation or in co-inoculation variants.
Genes of the isolates that were absent in the commercial strain R. leguminosarum RCAM0626 are shown in Table 9. Among them, the nodPZ and nolK genes could affect the rate of common vetch nodule formation, their total number and/or nitrogen-fixing activity. The nodP gene is involved in the 6-O-sulfation of the reducing end of the NFs [17]. It was known that NodP functioned in conjunction with NodQ, which synthesizes the donor of the sulfate group [43]. It should be noted that the commercial strain R. leguminosarum RCAM0626 has the nodQ gene, although the importance of NFs sulfation for the common vetch has not been shown. The functions of nodZ and nolK genes have been described above. It is noteworthy that the nolK gene is present only in the isolate Bradyrhizobium sp. Opo-243, which alone contributed to the acceleration of nodulation on V. sativa plants (Figure 4). The nodZ and nolK genes were found by other authors in a number of R. leguminosarum strains, e.g., in many pea nodulating rhizobia the NFs carry a fucosyl group, which may indicate the importance of this modification for the tribe Fabeae [44]. Since we did not detect an increase in nitrogen-fixing activity per nodule in variants of co-inoculation of common vetch, we do not discuss the role of nif and fix genes that are present in isolates, although the strain M. kowhaii Ach-343 was re-isolated from the nodules together with the commercial strain RCAM0626.
The nodulation experiment with red clover, unlike those with alfalfa and common vetch, showed an increase in the level of nitrogen-fixing activity per one nodule caused by co-inoculations. The additional nif and fix genes of M. japonicum Opo-235 and M. japonicum Opo-242 might contributed to this parameter when co-inoculated with R. leguminosarum RCAM1365 (Table 9). Among them, nifZQXW genes associated with the synthesis of the Fe-S-cofactor, fixation of molybdenum and protection of nitrogenase from oxygen damage, as well as the nifV gene, which is involved in the maturation of nitrogenase through the synthesis of homocitrate [45,46,47,48,49]. The fixHSQ genes are involved in the synthesis of components of the oxidase complex, which is associated with the membrane and allows cells to breathe at low oxygen concentrations [20,46]. The fixKL genes, together with the fixJT genes, determine the expression of genes necessary for growth under microaerophilic conditions [20,50].
It should be noted that, in our previous work devoted to the analysis of genetic complementation in strains isolated from the relict symbiotic systems, the genes indicated above have already come into view. Thus, the nolK gene was mentioned in connection with the ability of the isolate Bradyrhizobium sp. Opo-243 to increase the rate of nodulation on the relict legume Oxytropis popoviana (host plant) when used with its paired strain M. japonicum Opo-242 isolated from the same nodule [3]. The presence of the complementary genes nifQV and fixJKL in M. japonicum Opo-235 and M. kowhaii Ach-343 was associated with an increase in the nitrogen-fixing activity of nodules on Glycyrrhiza uralensis plants co-inoculated with both of these strains. Moreover, it was shown that isolates complementing each other for these genes were localized in the same nodule cells [4].
The genes responsible for the synthesis of auxins and genes of the T6SS secretion system can also positively influence the process of nodule formation. The presence of these genes in M. kowhaii Ach-343 apparently caused a significant increase in biomass of alfalfa plants with a decrease in the number of root nodules (Table 6). The influence of secretion systems on the formation of plant–microbial relationships was previously shown on the example of two strains (M. loti 582 and M. huakuii 583) isolated from the endemic of Kamchatka Oxytropis kamtschatica [16]. These strains possessed the T3SS and T6SS genes and formed two types of nodules (typical elongated and abnormal rounded) on roots of the host plant after mono-inoculations. It was suggested that this phenomenon can be regulated by two different nodulation strategies: (1) NF dependent, by the infection thread formation, and (2) NF independent, by the direct penetration of bacteria through the host cell wall. However, it certainly cannot be ruled out that other genetic factors, which are not considered here, could influence the process of the symbiosis formation.
The mechanisms of the rhizobial synergy described above, which are associated with the genetic complementation of co-microsymbionts, can be based on interbacterial interactions involving membrane and secreted proteins. In addition, some affecting proteins can enter the environment after cell lysis. The phenomenon of bacterial synergy in the context of accelerating and increasing the efficiency of nodulation is most easily explained by the modification of NFs that occurs outside the cells or on their surface. At the same time, the cooperation between co-microsymbionts most likely depends on their spatial arrangement (rhizosphere, rhizoplane or nodule), and in the case of localization of bacteria in one nodule or in the same plant cells, the level of interaction should be higher.
4. Conclusions
Thus, it was shown that the strains isolated from relict leguminous plants differ significantly in the set of symbiotic genes, as well as the genes related to other genetic systems that affect plant–microbe interactions. Moreover, the studied isolates obtained from relict legumes have a noticeably greater diversity of such genes as compared to the commercial strains. The cooperation of rhizobial strains simultaneously present either on roots or in nodules was expressed in a change (both positive and negative) in the efficiency of symbiosis upon joint inoculation with commercial strains. It is assumed that carrying out genomic analysis of rhizobia will allow selecting complementary rhizobial strains and assessing the contribution of various genetic loci (including those with little-known functions) to the formation of plant–microbial relationships. It could be expected that the study of microbial synergy using rhizobia of relict legumes will make it possible to carry out targeted selection of co-microsymbionts for the increasing of the efficiency of agricultural legume–rhizobia systems.
Author Contributions
Conceptualization and writing, V.S. and E.A.; methodology, I.K., E.C., A.A. and O.Y.; investigation, A.S., D.K. and P.G.; resources, A.B. and A.V.; supervision, I.T. All authors have read and agreed to the published version of the manuscript.
Funding
The article was made with support of the Ministry of Science and Higher Education of the Russian Federation in accordance with agreement № 075-15-2020-920 date 16 November 2020 on providing a grant in the form of subsidies from the Federal budget of Russian Federation. The grant was provided for state support for the creation and development of a World-class Scientific Center “Agrotechnologies for the Future”.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Safronova, V.I.; Kimeklis, A.K.; Chizhevskaya, E.P.; Belimov, A.A.; Andronov, E.E.; Pinaev, A.G.; Pukhaev, A.R.; Popov, K.P.; Tikhonovich, I.A. Genetic diversity of rhizobia isolated from nodules of the relic species Vavilovia formosa (Stev.) Fed. Antonie Leeuwenhoek 2014, 105, 389–399. [Google Scholar] [CrossRef]
- Safronova, V.; Belimov, A.; Sazanova, A.; Kuznetsova, I.; Popova, J.; Andronov, E.; Verkhozina, A.; Tikhonovich, I. Does the Miocene-Pliocene relict legume Oxytropis triphylla form nitrogen-fixing nodules with a combination of bacterial strains? Int. J. Environ. Stud. 2017, 74, 706–714. [Google Scholar] [CrossRef]
- Safronova, V.; Belimov, A.; Sazanova, A.; Chirak, E.; Verkhozina, A.; Kuznetsova, I.; Andronov, E.; Puhalsky, J.; Tikhonovich, I. Taxonomically different co-microsymbionts of a relict legume Oxytropis popoviana have complementary sets of symbiotic genes and together increase the efficiency of plant nodulation. MPMI 2018, 31, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Safronova, V.; Belimov, A.; Sazanova, A.; Chirak, E.; Kuznetsova, I.; Andronov, E.; Pinaev, A.; Tsyganova, A.; Seliverstova, E.; Kitaeva, A.; et al. Two broad host range rhizobial strains isolated from relict legumes have various complementary effects on symbiotic parameters of co-inoculated plants. Front. Microbiol. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Sazanova, A.L.; Safronova, V.I.; Kuznetsova, I.G.; Karlov, D.S.; Belimov, A.A.; Andronov, E.E.; Chirak, E.R.; Popova, J.P.; Verkhozina, A.V.; Willems, A.; et al. Bosea caraganae sp. nov. a new species of slow-growing bacteria isolated from root nodules of the relict species Caragana jubata (Pall.) Poir. originating from Mongolia. Int. J. Syst. Evol. Microbiol. 2019, 69, 2687–2695. [Google Scholar] [CrossRef] [PubMed]
- Novikova, N.; Safronova, V. Transconjugants of Agrobacterium radiobacter harbouring sym genes of Rhizobium galegae can form an effective symbiosis with Medicago sativa. FEMS Microbiol. Lett. 1992, 93, 261–268. [Google Scholar] [CrossRef]
- Vincent, J.M. A manual for the practical study of root nodule bacteria. In IBP Handbook No. 15; Blackwell Scientific: Oxford, UK; Edinburgh, UK, 1970; pp. 73–97. [Google Scholar]
- Safronova, V.I.; Kuznetsova, I.G.; Sazanova, A.L.; Kimeklis, A.K.; Belimov, A.A.; Andronov, E.E.; Pinaev, A.G.; Chizhevskaya, E.P.; Pukhaev, A.R.; Popov, K.P.; et al. Boseavaviloviae sp. nov. a new species of slow-growing rhizobia isolated from nodules of the relict species Vavilovia formosa (Stev.) Fed. Antonie Leeuwenhoek 2015, 107, 911–920. [Google Scholar] [CrossRef]
- CCINFO/World Data Centre for Microorganisms. Available online: http://www.wfcc.info/ccinfo/collection/by_id/966 (accessed on 22 March 2013).
- Safronova, V.; Tikhonovich, I. Automated cryobank of microorganisms: Unique possibilities for long-term authorized depositing of commercial microbial strains. In Microbes in Applied Research: Current Advances and Challenges; Mendez-Vilas, A., Ed.; World Scientific Publishing Co.: Hackensack, NJ, USA, 2012; pp. 331–334. [Google Scholar]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P. Assembly of long error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- GitHub-Nanoporetech/Medaka: Sequence Correction Provided by ONT Research. Available online: https://github.com/nanoporetech/medaka (accessed on 1 March 2019).
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 28 April 2021).
- Turner, G.L.; Gibson, A.H. Measurement of nitrogen fixation by indirect means. In Methods for Evaluating Biological Nitrogen Fixation; Bergensen, F.J., Ed.; Wiley: Toronto, ON, Canada, 1980; pp. 111–138. [Google Scholar]
- Safronova, V.I.; Guro, P.V.; Sazanova, A.L.; Kuznetsova, I.G.; Belimov, A.A.; Yakubov, V.V.; Chirak, E.R.; Afonin, A.M.; Gogolev, Y.V.; Andronov, E.E.; et al. Rhizobial Microsymbionts of Kamchatka Oxytropis Species Possess Genes of the Type III and VI Secretion Systems, Which Can Affect the Development of Symbiosis. MPMI 2020, 33, 1232–1241. [Google Scholar] [CrossRef]
- Wais, R.J.; Keating, D.H.; Long, S.R. Structure-function analysis of Nod factor-induced root hair calcium spiking in Rhizobium-legume symbiosis. Plant Physiol. 2002, 129, 211–224. [Google Scholar] [CrossRef]
- Dos Santos, P.C.; Fang, Z.; Mason, S.W.; Setubal, J.C.; Dixon, R. Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes. BMC Genom. 2012, 13, 162. [Google Scholar] [CrossRef]
- Edgren, T.; Nordlund, S. The fixABCX genes in Rhodospirillum rubrum encode a putative membrane complex participating in electron transfer to nitrogenase. J. Bacteriol. 2004, 186, 2052–2060. [Google Scholar] [CrossRef]
- Fischer, H.-M. Genetic regulation of nitrogen fixation in Rhizobia. Microbiol. Rev. 1994, 58, 352–386. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Brigido, C.; Glick, B.R.; Rossi, M.J. The role of rhizobial ACC deaminase in the nodulation process of leguminous plants. Int. J. Agron. 2016, 2016, 1369472. [Google Scholar] [CrossRef]
- Ma, W.; Guinel, F.C.; Glick, B.R. Rhizobium leguminosarum biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Appl. Environ. Microb. 2003, 69, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Belimov, A.A.; Zinovkina, N.Y.; Safronova, V.I.; Litvinsky, V.A.; Nosikov, V.V.; Zavalin, A.A.; Tikhonovich, I.A. Rhizobial ACC deaminase contributes to efficient symbiosis with pea (Pisum sativum L.) under single and combined cadmium and water deficit stress. Environ. Exper. Bot. 2019, 167, 103859. [Google Scholar] [CrossRef]
- Hayashi, S.; Gresshoff, P.M.; Ferguson, B.J. Mechanistic action of gibberellins in legume nodulation. J. Integr. Plant Biol. 2014, 56, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Nett, R.S.; Montanares, M.; Marcassa, A.; Lu, X.; Nagel, R.; Charles, T.C.; Hedden, P.; Rojas, M.C.; Peters, R.J. Elucidation of gibberellin biosynthesis in bacteria reveals convergent evolution. Nat. Chem. Biol. 2017, 13, 69–74. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [PubMed]
- Fauvart, M.; Michiels, J. Rhizobial secreted proteins as determinants of host specificity in the rhizobium-legume symbiosis. FEMS Microbiol. Lett. 2008, 285, 1–9. [Google Scholar] [CrossRef]
- Wallden, K.; Rivera-Calzada, A.; Waksman, G. Type IV secretion systems: Versatility and diversity in function. Cell Microbiol. 2010, 12, 1203–1212. [Google Scholar] [CrossRef]
- Cascales, E.; Christie, P.J. The versatile bacterial type IV secretion systems. Nat. Rev. Microbiol. 2015, 1, 137–149. [Google Scholar] [CrossRef]
- Notti, R.Q.; Stebbins, C.E. The Structure and Function of Type III Secretion Systems. Microbiol. Spectr. 2016, 4, 4. [Google Scholar] [CrossRef]
- Okazaki, S.; Tittabutr, P.; Teulet, A.; Thouin, J.; Fardoux, J.; Chaintreuil, C.; Gully, D.; Arrighi, J.F.; Furuta, N.; Miwa, H.; et al. Rhizobium-legume symbiosis in the absence of Nod factors: Two possible scenarios with or without the T3SS. ISME J. 2016, 10, 64–74. [Google Scholar] [CrossRef]
- Russell, A.B.; Peterson, S.B.; Mougous, J.D. Type VI secretion system effectors: Poisons with a purpose. Nat. Rev. Microbiol. 2014, 51, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.S.; Sadowsky, M.J. Secretion systems and signal exchange between nitrogen-fixing rhizobia and legumes. Front. Plant Sci. 2015, 6, 491. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lara, I.M.; Geiger, O. The nodulation protein NodG shows the enzymatic activity of an 3-oxoacyl-acyl carrier protein reductase. MPMI 2001, 14, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Rodpothong, P.; Sullivan, J.T.; Songsrirote, K.; Sumpton, D.; Cheung, K.W.; Thomas-Oates, J.; Radutoiu, S.; Stougaard, J.; Ronson, C.W. Nodulation gene mutants of Mesorhizobium loti R7A-nodZ and nolL mutants have host-specific phenotypes on Lotus spp. MPMI 2009, 22, 1546–1554. [Google Scholar] [CrossRef]
- Sugawara, M.; Epstein, B.; Badgley, B.D.; Unno, T.; Xu, L.; Reese, J.; Gyaneshwar, P.; Denny, R.; Mudge, J.; Bharti, A.K.; et al. Comparative genomics of the core and accessory genomes of 48 Sinorhizobium strains comprising five genospecies. Genome Biol. 2013, 14, R17. [Google Scholar] [CrossRef]
- Wang, D.; Couderc, F.; Tian, C.F.; Gu, W.; Liu, L.X.; Poinsot, V. Conserved composition of Nod factors and exopolysaccharides produced by different phylogenetic lineage Sinorhizobium strains nodulating soybean. Front. Microbiol. 2018, 9, 2852. [Google Scholar] [CrossRef]
- Kaneko, T.; Maita, H.; Hirakawa, H.; Uchiike, N.; Minamisawa, K.; Watanabe, A.; Sato, S. Complete genome sequence of the soybean symbiont Bradyrhizobium japonicum strain USDA6T. Genes 2011, 2, 763–787. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Van Montagu, M.; Holsters, M. The nodulation gene nolK of Azorhizobium caulinodans is involved in the formation of GDP-fucose from GDP-mannose. FEBS Lett. 1997, 409, 312–316. [Google Scholar] [CrossRef]
- Corvera, A.; Promé, D.; Promé, J.-C.; Martínez-Romero, E.; Romero, D. The nolL Gene from Rhizobium etli Determines Nodulation Efficiency by Mediating the Acetylation of the Fucosyl Residue in the Nodulation Factor. MPMI 1999, 12, 236–246. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pinto, F.G.; Chueire, L.M.; Vasconcelos, A.T.; Nicolás, M.F.; Almeida, L.G.; Souza, R.C.; Menna, P.; Barcellos, F.G.; Megías, M.; Hungria, M. Novel genes related to nodulation, secretion systems, and surface structures revealed by a genome draft of Rhizobium tropici strain PRF 81. Funct. Integr. Genom. 2009, 9, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Rivilla, R.; Sutton, J.M.; Downie, J.A. Rhizobium leguminosarum NodT is related to a family of outer-membrane transport proteins that includes TolC, PrtF, CyaE and AprF. Gene 1995, 161, 27–31. [Google Scholar] [CrossRef]
- Perret, X.; Staehelin, C.; Broughton, W.J. Molecular basis of symbiotic promiscuity. Microbiol. Mol. Biol. Rev. 2000, 64, 180–201. [Google Scholar] [CrossRef]
- Debelle, F.; Moulin, L.; Mangin, B.; Dénarié, J.; Boivin, C. Nod-Genes and Nod-signals and the evolution of the rhizobium-legume symbiosis. Acta Biochim. Pol. 2001, 48, 359–365. [Google Scholar] [CrossRef]
- Barnett, M.J.; Fisher, R.F.; Jones, T.; Komp, C.; Abola, A.P.; Barloy-Hubler, F.; Bowser, L.; Capela, D.; Galibert, F.; Gouzy, J.; et al. Nucleotide sequence and predicted functions of the entire Sinorhizobium meliloti pSymA megaplasmid. Proc. Natl. Acad. Sci. USA 2001, 98, 9883–9888. [Google Scholar] [CrossRef]
- Black, M.; Moolhuizen, P.; Chapman, B.; Barrero, R.; Howieson, J.; Hungria, M.; Bellgard, M. The genetics of symbiotic nitrogen fixation: Comparative genomics of 14 Rhizobia strains by resolution of protein clusters. Genes 2012, 3, 138–166. [Google Scholar] [CrossRef]
- Boyd, E.S.; Costas, A.M.G.; Hamilton, T.L.; Mus, F.; Peters, J.W. Evolution of molybdenum nitrogenase during the transition from anaerobic to aerobic metabolism. J. Bacteriol. 2015, 197, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J.; Jones, R.; Woodley, P.R.; Wilborn, J.R.; Robson, R.L. Nucleotide sequence and genetic analysis of the Azotobacter chroococcum nifUSVWZM gene cluster, including a new gene (nifP) which encodes a serine acetyltransferase. J. Bacteriol. 1991, 173, 5457–5469. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Burgess, B.K. Evidence for the direct interaction of the nifW gene product with the MoFe protein. J. Biol. Chem. 1996, 271, 9764–9770. [Google Scholar] [CrossRef] [PubMed]
- Ermilova, E.V. Molecular Aspects of Adaptation of Prokaryotes; Khimizdat: St. Petersburg, Russia, 2012; p. 344. (In Russian) [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).